
Дисбаланс мышц плеча — лечение, симптомы, причины, диагностика
Мышечный дисбаланс встречается достаточно часто у людей, даже у спортсменов мирового класса. Мышцы передней части бедра (четырехглавая мышца) довольно часто сильнее, чем мышцы задней части бедра. А верхние трапециевидные мышцы, как правило, сильнее, чем нижние трапециевидные. Передние дельтовидные мышцы сильнее, чем задние дельтовидные. Развивают, как правило, мышцы, которые человек видит и они находятся спереди. Плечо является классическим примером части тела, которую стараются развивать в первую очередь. Неудивительно, что 30% всех травм при силовых тренировках возникает именно в плечевом суставе.
Мышцы плеча можно разделить на две группы:
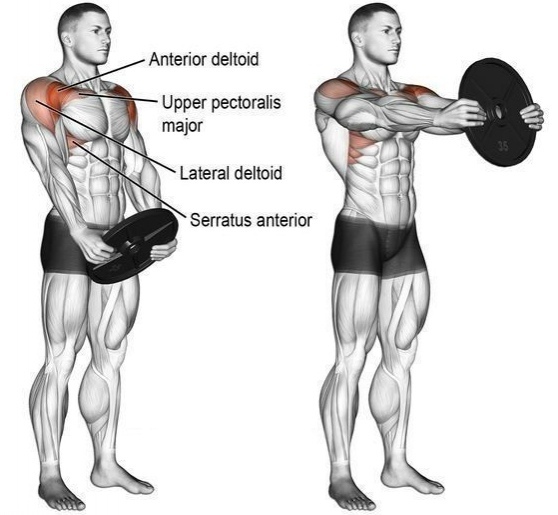
- Сгибатели и разгибатели и к ним относятся дельтовидная мышца, надостная и подостная мышцы, малая и большая круглые мышцы, подлопаточная мышца Наиболее сложные и многообразные функции имеет дельтовидная мышца. Дельтовидная мышца состоит из трех пучков
- Передний пучок дельтовидной мышцы — расположен в передней части плеча ,боковой пучок дельты в средней части плеча и задний пучок в задней части плеча
Данные исследования бодибилдеров показали, что сила передней дельтовидной мышцы могла превосходить таковую у обычных людей в 5 раз. А боковые дельтовидные мышцы могли быть сильнее в 3 раза. В тоже время, как ни странно, задние дельтовидные мышцы были сильнее лишь на 15%.
Мышечный дисбаланс между передней и задней дельтой отчасти можно объяснить акцентом на упражнения, при которых задействуются передние дельты и отсутствие упражнений для задней дельты.
Координированная работа мышц плеча позволяет выполнять необходимые движения и сохранять стабильность плечевого сустава. Нарушение взаимодействия мышц стабилизирующих плечо с мышцами, выполняющими движения, называется мышечным дисбалансом плеча. Такое нарушение создает проблемы при движении, так как мышцы, отвечающие за движения, сильнее мышц отвечающих за стабильность и это приводит к нестабильности.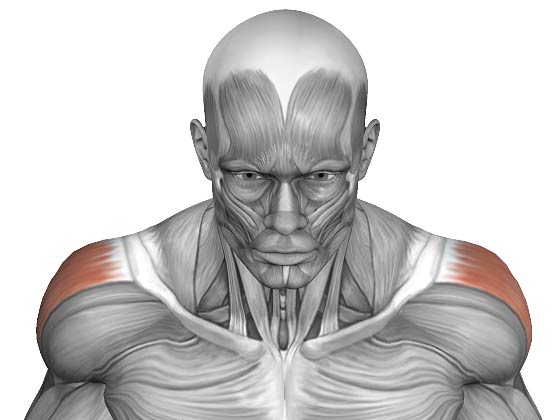
В большинстве случаев мышечный дисбаланс может возникнуть из-за повторяющихся движений. Наиболее распространенным является чрезмерное усиление мышц внутреннего ротатора плеча на фоне слабого развития мышц внешнего ротатора. Внутренние мышцы ротаторы укрепляются упражнениями на грудные мышцы и упражнениями для спины, а наружные группы мышц ротаторов прорабатываются редко. Это приводит к дисбалансу мышц вращательной манжеты и повреждению, как мышц ротаторной манжеты, так и сухожилий. Такой дисбаланс можно устранить с помощью специальных упражнений для укрепления мышц наружной ротации.
Причины
Чаще всего дисбаланс мышц плеча возникает вследствие повторяющихся движений с неравномерной нагрузкой различных групп мышц. Кроме того, причиной дисбаланса мышц плеча могут быть травмы плеча. Также способствовать развитию дисбаланса мышц плеча могут быть повреждения сухожилий и связок или длительные статические нагрузки, плохая осанка. Например, при профессиях связанных с длительным сидением (офисные работники, водители, работающие за компьютером) довольно часто возникает нарушение баланса мышц плеча. В то время как внутренние ротаторы подтянуты, внешние ротаторы в растянутом состоянии и происходит их постепенное ослабление, это является результатом позы и опоры при сидении. В таких случаях необходимо растяжение внутренних ротаторов и укрепление внешних ротаторов с помощью упражнений.
Симптомы и диагностика
Нарушение баланса мышц плеча, как правило, проявляются такими симптомами, как боли, жжение в области плеча, усталость, уменьшение диапазона движений в плече. Наличие таких симптомов дает возможность врачу предположить наличие дисбаланса мышц плеча. Для исключения органических изменений в плече назначаются инструментальные методы обследования, такие как МРТ, особенно если есть подозрения на наличие повреждений вращательной манжеты. При наличии нарушений чувствительности возможно назначение ЭМГ и ЭНМГ, для выявления возможного нарушения проводимости по нервным волокнам.
Лечение
Наиболее эффективным методом лечения дисбаланса мышц плеча является ЛФК. Программа упражнений позволяет укрепить ослабленные мышцы и расслабить мышцы находящиеся в постоянном напряжении. Кроме ЛФК возможно использование физиопроцедур, которые помогают уменьшить болевые проявления. Массаж позволяет уменьшить мышечный спазм и улучшить кровообращение в мышцах. Большое значение в лечение мышечного дисбаланса мышц плеча имеет изменение осанки во время работы особенно если работа пациента связана с длительным сидением.
Как правило, лечение мышечного дисбаланса консервативное. Но в тех случаях когда дисбаланс мышц тяжелый и привел к осложнениям (например нестабильности) то в таких случаях проводится артроскопическая операция для укрепления стабильности плечевого сустава.
Дельтовидные мышцы их строение и функции | Фактор Силы
Дельтовидная мышца в значительной мере способствует укреплению плечевого сустава. Образуя ярко выраженную выпуклость, она обусловливает форму всей области сустава. Между дельтовидной и большой грудной мышцами находится хорошо видная на коже борозда. Задний край дельтовидной мышцы также легко может быть определен на Живом человеке.
Анатомически в дельтовидной мыше выделяют три пучка:
- передний;
- средний (боковой)
- задний.
Однако, по результатам электромиографических исследований, в ней можно выделить как минимум семь групп волокон, функционирующих независимо друг от друга
Строение дельтовидной мышцы плечаДельтовидная мышца располагается над плечевым суставом. Она начинается от ости лопатки, акромиона и акро-миального конца ключицы, а прикрепляется на плечевой кости к дельтовидной бугристости.
Мышца имеет многочисленные соединительнотканные прослойки, по отношению к которым отдельные ее пучки идут под некоторым углом. Эта особенность строения относится главным образом к средней части мышцы, делает ее многоперистой и способствует увеличению подъемной силы.
Функции дельтовидной мышцы плеча
Функции дельтовидной мышцы сложны и многообразны. Если попеременно работают то передняя, то задняя части мышцы, то происходит сгибание и разгибание конечности. Если же напрягается вся мышца, то ее передняя и задняя части действуют одна по отношению к другой под некоторым углом и направление их равнодействующей совпадает с направлением волокон средней части мышцы. Таким образом, напрягаясь целиком, эта мышца производит отведение плеча.
При сокращении дельтовидная мышца вначале несколько поднимает плечевую кость, отведение же этой кости наступает после того, как ее головка упирается в свод плечевого сустава. Когда тонус этой мышцы очень велик, плечо при спокойном стоянии несколько отведено. Поскольку мышца прикрепляется к дельтовидной бугристости, располагающейся снаружи и спереди в верхней половине плечевой кости, она может участвовать также и во вращении ее вокруг вертикальной оси, а именно: передняя, ключичная, часть мышцы не только поднимает руку кпереди (сгибание), но и пронирует ее, а задняя часть не только разгибает, но и супинирует.
Если передняя часть дельтовидной мышцы работает совместно со средней, то по правилу параллелограмма сил мышца сгибает и несколько отводит руку. Если же средняя часть работает совместно с задней, то происходит одновременно разгибание и отведение руки. Плечо силы этой мышцы, при котором ей приходится работать, меньше, чем плечо силы тяжести.
Антагонистами при отведении руки выступают большая грудная и широчайшая мышца спины.
Передние пучки участвуют в боковом отведении руки при наружном вращении плеча. В сгибании плеча их роль невелика, но они помогают в этом движении большой грудной мышце (локоть чуть ниже плеча). Содействуют мышцам: подключичной, большой грудной и широчайшей спины при внутреннем вращении плеча.
Латеральные пучки участвуют в боковом отведении плеча при его положении во внутреннем вращении и в горизонтальном отведении при его наружном вращении, но практически не участвуют в горизонтальном разгибании плеча (при его внутреннем вращении).
Задние пучки принимают большое участие в горизонтальном разгибании, особенно по причине малого участия широчайшей мышцы спины в этом движении в горизонтальной плоскости. Другие горизонтальные разгибатели — подостная и малая круглая мышцы — также работают вместе с задней порцией дельтовидной мышцы как наружные ротаторы, антагонистично внутренним ротаторам — большим грудным мышцам и широчайшим. Задняя порция дельтовидной мышцы также принимает большое участие в переразгибании плеча, при поддержке длинной головки трицепса.
Сморите так жеИнтересный способ сжигать калории))))
Описание ФОЛЛИТРОПИН ДЕЛЬТА показания, дозировки, противопоказания активного вещества FOLLITROPIN DELTA
Лекарственное средство обладает выраженной гонадотропной активностью, в связи с чем при применении препарата могут развиваться нежелательные реакции от легкой до тяжелой степени тяжести.Применение гонадотропинов требует присутствия квалифицированного медицинского персонала, а также соответствующего оборудования.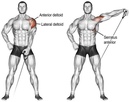 Перед началом применения и в процессе терапии фоллитропином дельта следует контролировать состояние яичников (УЗИ, в т.ч. с определением концентрации эстрадиола в плазме крови).
Перед началом применения и в процессе терапии фоллитропином дельта следует контролировать состояние яичников (УЗИ, в т.ч. с определением концентрации эстрадиола в плазме крови).
У пациенток, проходящих процедуру контролируемой стимуляции роста фолликулов, может наблюдаться увеличение яичников и существует риск развития СГЯ. При соблюдении режима дозирования в сочетании с мониторингом проводимой терапии возможно минимизировать риск возникновения вышеуказанных реакций.
В определенной степени, увеличение яичников представляет ожидаемую реакцию на проведение контролируемой гиперстимуляции яичников и чаще развивается у пациенток с СПКЯ, обычно регрессирует самостоятельно, без дополнительной терапии. В отличие от неосложненного увеличения яичников, клинические симптомы СГЯ могут проявляться с нарастающей выраженностью. Они включают в себя: значительное увеличение яичников, высокую концентрацию эстрогенов в плазме крови, а также увеличение проницаемости сосудов, что может привести к накоплению жидкости в брюшной, плевральной и, реже, перикардиальной полости. Для снижения риска развития СГЯ необходимо проводить тщательный и частый мониторинг созревания фолликулов. В тяжелых случаях СГЯ возможны следующие симптомы: боль в животе, вздутие живота, значительное увеличение яичников, увеличение массы тела, одышка, олигурия и нарушения со стороны желудочно-кишечного тракта, включая тошноту, рвоту и диарею. При клиническом обследовании возможно выявление гиповолемии, гемоконцентрации, электролитных нарушений, асцита, гемоперитонеума, экссудативного плеврита, гидроторакса, острого респираторного дистресс-синдрома. Очень редко СГЯ тяжелой степени может осложняться перекрутом яичника или тромбоэмболическими осложнениями, такими как тромбоэмболия легочной артерии, ишемический инсульт или инфаркт миокарда.
Чрезмерная реакция яичников на введение гонадотропинов редко приводит к развитию СГЯ, если не вводится ХГЧ с целью окончательного созревания фолликулов. СГЯ может быть более тяжелым и затяжным при развитии беременности.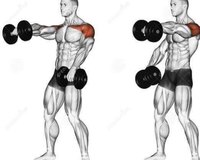 Поэтому в случае СГЯ не следует вводить ХГЧ, а пациентку следует предупредить о необходимости воздерживаться от половых контактов или использовать барьерные методы контрацепции не менее 4 дней. СГЯ может быстро прогрессировать (в течение от 24 ч до нескольких дней), становясь серьезным медицинским осложнением. Наиболее часто СГЯ развивается после завершения гормональной терапии, позднее развитие СГЯ возможно вследствие гормональных изменений при беременности. Поэтому необходимо тщательное наблюдение за пациентками, по крайней мере, в течение не менее 2-х недель после окончательного созревания фолликулов.
Поэтому в случае СГЯ не следует вводить ХГЧ, а пациентку следует предупредить о необходимости воздерживаться от половых контактов или использовать барьерные методы контрацепции не менее 4 дней. СГЯ может быстро прогрессировать (в течение от 24 ч до нескольких дней), становясь серьезным медицинским осложнением. Наиболее часто СГЯ развивается после завершения гормональной терапии, позднее развитие СГЯ возможно вследствие гормональных изменений при беременности. Поэтому необходимо тщательное наблюдение за пациентками, по крайней мере, в течение не менее 2-х недель после окончательного созревания фолликулов.
У женщин с известными факторами риска развития тромбоэмболических осложнений такими, как индивидуальная или семейная предрасположенность, ожирение (ИМТ >30 кг/м2) или тромбофилия, могут иметь повышенный риск венозных или артериальных тромбоэмболических осложнений во время или после лечения гонадотропинами. В таких случаях польза их применения должна быть сопоставлена с возможным риском. Следует учитывать, что сама беременность также повышает риск развития тромбоэмболических осложнений.
При проведении ВРТ описаны случаи развития перекрута яичника. Причиной перекрута яичника могут быть СГЯ. беременность, проведенные ранее хирургические вмешательства на брюшной полости, перекрут яичника в анамнезе, наличие кисты яичника в настоящее время или в анамнезе, а также СПКЯ. Повреждения яичника вследствие нарушения притока крови могут быть уменьшены при своевременной диагностике и проведении экстренной деторсии яичника.
При многоплодной беременности отмечается повышенный риск неблагоприятных материнских и перинатальных исходов. В случае проведения ВРТ вероятность возникновения многоплодной беременности зависит от числа введенных эмбрионов, их качества и возраста пациентки. В редких случаях отмечались случаи беременности двойней даже при переносе одного эмбриона. Пациентка должна быть предупреждена о потенциальном риске многоплодной беременности до начала лечения.
Частота прерывания беременности (выкидыша, самопроизвольного аборта) у пациенток, которым проводится контролируемая стимуляция яичников в рамках программ ВРТ, выше, чем у здоровых женщин.
У пациенток с заболеваниями маточных труб в анамнезе, как при естественном зачатии, так и при лечении бесплодия, имеется высокий риск возникновения эктопической беременности. Распространенность внематочной беременности после ВРТ выше по сравнению с общей популяцией.
Имеется информация о новообразованиях яичников и других органов репродуктивной системы, как доброкачественных, так и злокачественных, у женщин после проведения нескольких курсов лечения бесплодия. В настоящее время не установлено, увеличивает ли лечение гонадотропинами исходный риск этих опухолей у женщин с бесплодием.
Распространенность врожденных пороков развития плода при использовании ВРТ несколько выше, чем при естественном зачатии. Считается, что это может быть связано с индивидуальными особенностями родителей (возрастом матери, характеристиками спермы) и многоплодной беременностью.
Возмещение и компенсация: Delta Airlines — AirHelp
Все рейсы, выполняемые Delta Airlines из аэропортов в ЕС, должны соответствовать законодательству Европейского союза, которое защищает права авиапассажиров в случае задержки рейсов.
Согласно Регламенту ЕС №261 в случае задержки рейса авиакомпания обязана обеспечить пассажиров питанием и напитками и предоставить доступ к средствам связи. Помимо этого, при необходимости авиакомпания должна разместить пассажиров в гостинице и предложить другие рейсы.
Более того, если рейс задержан по вине а/к Delta Airlines, согласно закону она обязана выплатить пассажирам компенсацию, при условии что задержка составила минимум 3 часа. Более подробная информация приведена в справочнике по выплате компенсации за задержку рейса.
Поэтому, если вы прибываете в аэропорт прилета с опозданием на 3 часа или более, стоит проверить свой рейс.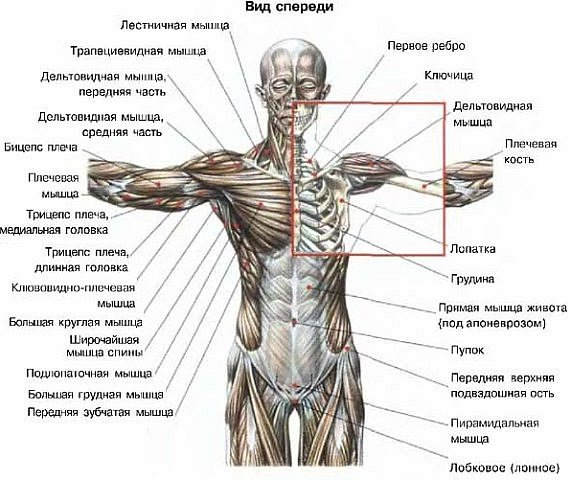 Возможно, вам полагается компенсация от а/к Delta Airlines за задержку рейса. Не упустите возможность получить до 600 евро на человека.
Возможно, вам полагается компенсация от а/к Delta Airlines за задержку рейса. Не упустите возможность получить до 600 евро на человека.
Мы постарались максимально упростить процесс подачи заявки на компенсацию за задержку рейсов Delta Airlines. Воспользовавшись специальным инструментом проверки рейсов и услугами наших специалистов по правам авиапассажиров, вы сможете уточнить время задержки и проверить наличие исключительных обстоятельств, влияющих на ваше право на компенсацию. Как правило, заявка на компенсацию может быть подана в отношении рейсов, которые были отменены или задержаны в течение последних 3 лет.
В Европе Регламент EC №261 защищает права всех пассажиров в случае отмены рейса, особенно если это сделано без предварительного уведомления.
Если а/к Delta Airlines отменяет рейс менее чем за 14 дней до запланированной даты вылета из аэропорта на территории ЕС, пассажиры вправе требовать от Delta Airlinesкомпенсацию за отмену рейса в размере до 600 евро на человека.
Исключение составляют случаи, когда рейс отменяется по причине действия исключительных обстоятельств, в результате наступления которых а/к Delta Airlines была вынуждена отменить рейс. В соответствии с действующим законодательством авиакомпания обязана выплатить компенсацию, только если рейс был отменен по её вине. При этом авиакомпания должна принять меры, гарантирующие доставку пассажиров в конечный пункт назначения, или выплатить им 100% стоимости билетов.
Авиакомпании (включая Delta Airlines) не обязаны выплачивать компенсацию, если они могут предложить билеты на другой рейс, время прилета которого практически идентично времени прилета отмененного рейса. Сведения о действующих временных ограничениях приведены в нашем справочнике по выплате компенсации в случае отмены рейса.
Несмотря на что Delta Airlines не европейская авиакомпания, она обязана соблюдать законодательство ЕС при вылете из аэропорта на территории Европейского союза. Это означает, что пассажиры, вылетающие из аэропорта на территории ЕС рейсом Delta Airlines, защищены Регламентом EC №261, представляющим собой один из наиболее исчерпывающих наборов прав авиапассажиров в мире.
Это означает, что пассажиры, вылетающие из аэропорта на территории ЕС рейсом Delta Airlines, защищены Регламентом EC №261, представляющим собой один из наиболее исчерпывающих наборов прав авиапассажиров в мире.
Таким образом, пассажиры вправе требовать от Delta Airlines выплаты компенсации за задержку, отмену или овербукинг рейса, при условии что эти обстоятельства возникли по вине Delta Airlines.
К сожалению, такие права не предусмотрены для тех, кто летит из-за пределов ЕС. Пассажиры Delta Airlines, которые прилетают в ЕС, не обладают правами, предусмотренными Регламентом ЕС №261, если аэропорт вылета находится за пределами ЕС. В этом случае следует ознакомиться с другими правами авиапассажиров.
Какие рейсы Delta Airlines подпадают под действие Регламента EC №261?
В соответствии с EC №261 все суммы в таблице указаны в евро.
Как и в случае с любым другим законом, предусмотрен ряд исключений и особых обстоятельств, которые могут повлиять на размер компенсации, выплачиваемой в каждом конкретном случае. Воспользуйтесь бесплатным калькулятором AirHelp, чтобы узнать сумму причитающейся вам компенсации.
Размещение людей с ограниченными физическими возможностями
Туристские гостиничные комплексы «Измайлово» («Гамма», «Дельта») приглашают в свои номера людей с ограниченными возможностями.
Вход в гостиницы оборудован пандусом, по которому без труда можно подняться на коляске. Если понадобится помощь, сотрудники отелей с удовольствием проводят вас до номера.
В широкие лифты поместятся инвалидная коляска с владельцем и сопровождающий. Просторные коридоры гостиниц рассчитаны на комфортный проезд на коляске без опасения задеть стены.
В отелях «Гамма» и «Дельта» предусмотрены номера для людей с ограниченными возможностями. Однокомнатные номера с прихожей — светлые и просторные. В широкий дверной проем без труда пройдет инвалидная коляска. Кровати занижены — на них будет удобно лечь самостоятельно.
Помимо специальных удобств, номера для инвалидов имеют всё необходимое для отдыха и работы. В вашем распоряжении — письменный стол, ЖК-телевизор, спутниковое телевидение, мини-холодильник и сейф.
Важная деталь номера — ванная комната. Здесь установлены поручни для рук, чтобы пользоваться ванной было удобно. Сантехника (ванна, унитаз и раковина) специально занижены. Доступен традиционный банный набор — фен, полотенца, средства личной гигиены, тапочки.
Телефон есть как в спальной комнате, так и в ванной. Если понадобится помощь или возникнут вопросы, позвоните — вам помогут сотрудники гостиницы. Для инвалидов, как и для остальных гостей Туристских гостиничных комплексов «Измайлово» («Гамма», «Дельта»), работает здравпункт.
Электронные замки избавят от необходимости пользоваться обычными ключами — двери легко открываются специальными картами.
Специальные номера для людей с ограниченными возможностями — обязательный атрибут современной гостиницы. Для инвалидов Туристские гостиничные комплексы «Измайлово» предлагают комфортные условия и безупречный сервис.
Посмотреть номера для людей с ограниченными возможностями в отеле ГаммаПосмотреть номера для людей с ограниченными возможностями в отеле Дельта
| 02.1.001 | Самозажигающийся огонь на спасательном круге после включения должен гореть не менее чем | 2 часа |
| 02.1.002 | С какой высоты можно сбрасывать спасательный круг с самозажигающимся огнем не опасаясь его повреждения? |  org/Answer»>30 метров org/Answer»>30 метров |
| 02.1.003 | Автоматически действующая дымовая шашка, прикрепленная к спасательному кругу, должна давать дым как минимум | 15 минут |
| 02.1.004 | Может ли загореться спасательный жилет после того, как был полностью охвачен пламенем в течение короткого времени (1-2 сек.)? | Нет |
| 02.1.005 | Требуется ли снимать спасательный жилет перед тем, как залезть в спасательный плот (шлюпку) из воды? | Нет |
| 02.1.006 | За какое время согласно требованиям конвенции СОЛАС-74 должен быть надет гидрокостюм без посторонней помощи? | Не более 2 минут |
| 02.1.007 | Если в гидрокостюме пробежать через пламя (1- 2 сек.) | Гидрокостюм не должен загореться |
| 02.1.008 | Обеспечивает ли в гидрокостюм спуск на надувной спасательный плот (шлюпку) по штормтрапу? | |
| 02.1.009 | Допускает ли конструкция гидрокостюма прыжок в воду с высоты 2,5 м без повреждения или смещения при этом гидрокостюма или его оборудования и без телесных повреждений? | Да |
| 02.1.010 | Допускает ли конструкция гидрокостюма прыжок в воду с высоты 3,5 м без повреждения или смещения при этом гидрокостюма или его оборудования и без телесных повреждений? | Да |
| 02.1.011 | Допускает ли конструкция гидрокостюма прыжок в воду с высоты 4,5 м без повреждения или смещения при этом гидрокостюма или его оборудования и без телесных повреждений? | Да |
| 02.1.012 | Продолжительность нахождения человека в ледяной воде (+2° С) в гидрокостюме изготовленном из материала обладающего теплозащитными свойствами до легкой степени гипотермии | 6 часов |
| 02.1.013 | Температура воды +3°С Позволяет ли гидрокостюм, изготовленный из материала, обладающего теплоизоляционными свойствами, находится в воде в течение 5 часов до развития легкой степени гипотермии? | |
| 02.1.014 | В течение какого времени обеспечивается достаточная теплозащита человека в воде в гидрокостюме изготовленном из материала, не обладающего теплоизоляционными свойствами, при температуре воды 5 градусов | Не менее часа |
| 02.1.016 | Температура воды +4°С. Позволяет ли гидрокостюм, изготовленный из материала, обладающего теплоизоляционными свойствами, находится в воде в течение 6 часов? | Да |
| 02.1.017 | Температура воды +2°С. Позволяет ли гидрокостюм, изготовленный из материала, обладающего теплоизоляционными свойствами, находится в воде в течение 7 часов? | Нет, возникает угроза переохлаждения |
| 02.1.018 | На судне спасательные круги должны размещаться | На всех открытых палубах |
| 02.1.019 | Спасательный жилет переворачивает тело потерявшего сознание человека с лицом находящимся в воде в положение при котором его рот находится над водой за время |  org/Answer»>не более 5 секунд org/Answer»>не более 5 секунд |
| 02.1.020 | Где на судне устанавливаются спасательные круги с дымовой шашкой? | На ходовом мостике |
| 02.1.021 | Где на судне расположены спасательные круги с самозажигающимися огнями? | Равномерно по обоим бортам судна |
| 02.1.022 | Дополнительные спасательные жилеты на пассажирском судне должны храниться | На палубе в местах сбора пассажиров |
| 02.1.023 | Символ, изображённый на рисунке, обозначает | Место хранения детских спасательных жилетов |
| 02.1.024 | Символ, изображённый на рисунке, обозначает | Место хранения спасательных гидрокостюмов |
| 02.1.025 | Символ, изображённый на рисунке, обозначает | Место установки спасательного круга с огнём |
| 02.1.026 | Символ, изображённый на рисунке, обозначает | Место установки спасательного круга с линем |
| 02.1.027 | Можно ли в спасательном жилете прыгать, сложив руки над головой, с высоты 1м без получения телесных повреждений и при этом – без смещения или повреждения жилета или его оборудования? | Да |
| 02.1.028 | Можно ли в спасательном жилете прыгать, сложив руки над головой, с высоты 2,5 м без получения телесных повреждений и при этом – без смещения или повреждения жилета или его оборудования? | Нет |
| 02.1.029 | Можно ли в спасательном жилете прыгать, сложив руки над головой, с высоты 4,5 м без получения телесных повреждений и при этом – без смещения или повреждения жилета или его оборудования? | Нет |
| 02.1.030 | Можно ли в спасательном жилете прыгать, наложив руки на жилет, с высоты 2,5 м без получения телесных повреждений и при этом – без смещения или повреждения жилета или его оборудования? | Да |
| 02.1.031 | Можно ли в спасательном жилете прыгать, наложив руки на жилет, с высоты 4,5 м без получения телесных повреждений и при этом – без смещения или повреждения жилета или его оборудования? | Да |
| 02.1.032 | Укажите, какие спасательные жилеты могут поставляться на суда? | Для младенцев, детей и взрослых |
| 02.1.033 | Должен ли спасательный жилет обеспечиваться каким-либо средством для осуществления связки людей, находящихся в воде? | Да |
| 02.1.034 | Продолжительность горения сигнального огня спасательного жилета составляет | Не менее 8 часов |
| 02.1.035 | Какой вес должен иметь спасательный круг, если он должен быть укомплектован светодымящимся буем? | Не менее 4 кг |
| 02.1.036 | Дымовые шашки спасательных кругов должны давать дым как минимум в течение |  org/Answer»>15 минут org/Answer»>15 минут |
| 02.1.037 | Какое индивидуальное спасательное средство должно находиться у парадного трапа? | Спасательный круг с линем |
| 02.2.001 | Как безопаснее прыгать в спасательном жилете за борт? | Вниз ногами, прижав к груди спасательный жилет и оттягивая его вниз |
| 02.2.002 | Спасательные круги, используемые на судах, оснащаются |
|
| 02.2.003 | Какие типы спасательных средств считаются индивидуальными? |
|
| 03.1.001 | Можно ли сбрасывать контейнер спасательного плота в воду с высоты 15 м безопасно для оборудования и конструкции плота | Да |
03. 1.002 1.002 | Можно ли сбрасывать контейнер спасательного плота в воду с высоты 18 м безопасно для оборудования и конструкции плота | Да |
| 03.1.003 | Можно ли сбрасывать контейнер спасательного плота в воду с высоты 20 м безопасно для оборудования и конструкции плота | Да, только при условии, что плот испытан для сбрасывания с этой высоты |
| 03.1.004 | Можно ли сбрасывать контейнер спасательного плота в воду с высоты 25 м безопасно для оборудования и конструкции плота | Да, только при условии, что плот испытан для сбрасывания с этой высоты |
| 03.1.005 | Пусковое устройство и приспособления моторных спасательных шлюпок должны обеспечивать пуск двигателя при температуре окружающей среды минус 15°С в течение | 2 минут |
| 03.1.006 | Допускается ли прыгать на спасательный плот, в котором нет людей, не опасаясь его повредить с высоты 2,5м? | Да |
| 03.1.007 | Допускается ли прыгать на спасательный плот, в котором нет людей, не опасаясь его повредить с высоты 3,5м? | 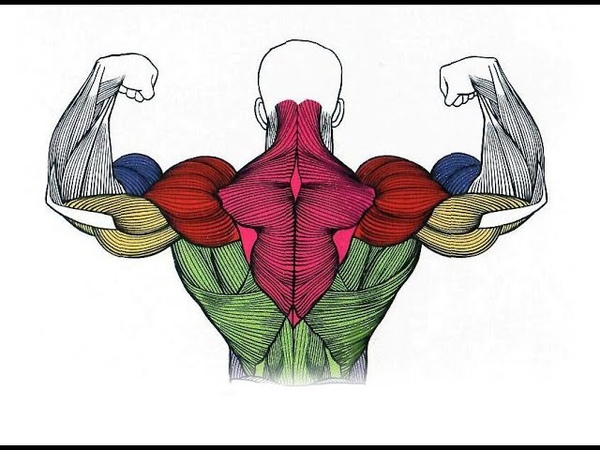 org/Answer»>Да org/Answer»>Да |
| 03.1.008 | Допускается ли прыгать на спасательный плот, в котором нет людей, не опасаясь его повредить с высоты 2,5м? | Да |
| 03.1.011 | Символ, изображённый на рисунке, обозначает | Место расположения надувного спасательного плота |
| 03.1.012 | Символ, изображённый на рисунке, обозначает | Место установки спасательной шлюпки |
| 03.1.013 | Символ, изображённый на рисунке, обозначает | Место установки дежурной шлюпки |
| 03.1.014 | Символ, изображённый на рисунке, обозначает | Место хранения теплозащитных средств |
| 03.1.015 | Посадка людей в спасательную шлюпку на грузовом судне должна быть совершена в течение… с момента подачи команды к посадке | 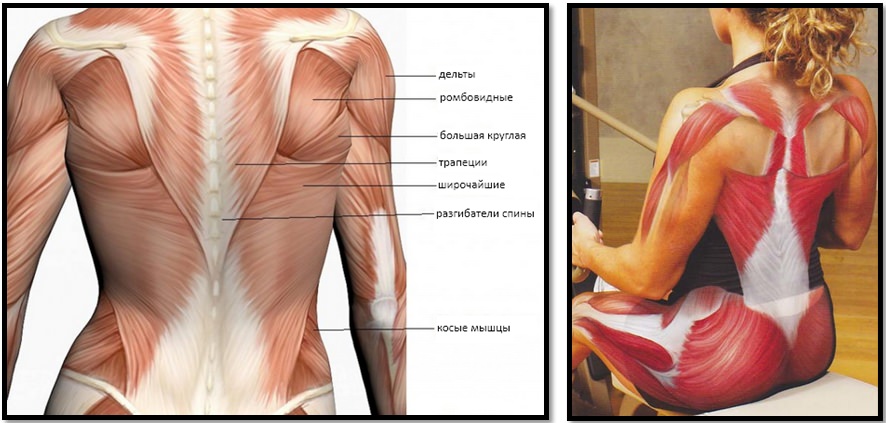 org/Answer»>3 минут org/Answer»>3 минут |
| 03.1.017 | Укажите период времени в течение которого гарантированно спасательный плот способен оставаться на плаву, выдерживая влияние окружающей среды | 30 дней |
| 03.1.018 | Применяются ли на грузовых судах спасательные плоты вместимостью менее 6 человек? | Не применяются |
| 03.2.001 | Какие способы, из числа указанных в ответах, наиболее безопасны для попадания на надувной спасательный плот, стоящий у борта судна? |
|
| 03.2.002 | Закончив посадку в спасательную шлюпку, следует немедленно |
|
| 03.2.003 | Конструкция полностью закрытой спасательной шлюпки должна обеспечивать |
|
| 03.2.004 | На каждом пассажирском судне должны быть предусмотрены коллективные спасательные средства следующих типов |
|
| 03.2.005 | На каждом грузовом судне должны быть предусмотрены коллективные спасательные средства следующих типов |
|
| 04.1.003 | Для подъёма людей в спасательную шлюпку из воды используют | Посадочный трап |
| 04.1.006 | Для удержания спасательного плота у борта судна следует использовать | Фалинь |
| 04.1.007 | Какие пиротехнические средства находятся внутри каждой спасательной шлюпки? |  org/Answer»>Парашютные ракеты, фальшфейеры, дымовые шашки org/Answer»>Парашютные ракеты, фальшфейеры, дымовые шашки |
| 04.1.010 | Есть ли в спасательной шлюпки рыболовные принадлежности? | Да |
| 04.1.012 | На какое количество человек в спасательной шлюпке имеются теплозащитные средства? | 10% от общего числа людей |
| 04.1.015 | Входят ли тёплые одеяла в перечень снабжения спасательной шлюпки? | Нет |
| 04.1.016 | На сколько суток мореплавания рассчитан надувной спасательный плот? | 30 суток |
| 04.1.017 | Штатное устройство для сбора дождевой воды на надувном спасательном плоту находится | На тенте плота |
| 04.1.018 | Плавучий нож, предназначенный для экстренного обрезания фалиня, в спасательном плоту находится | 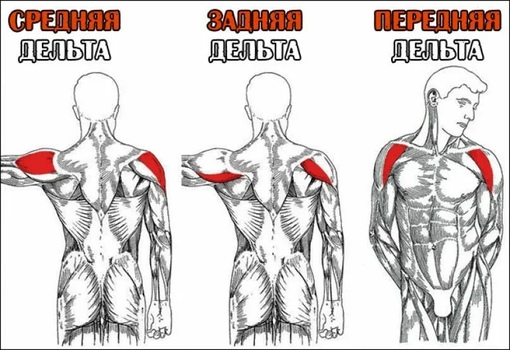 org/Answer»>Снаружи на тенте плота org/Answer»>Снаружи на тенте плота |
| 04.1.019 | Сколько комплектов аптечки первой помощи упаковано внутри спасательного плота? | Один комплект на всех |
| 04.1.020 | Какое количество пресной воды упаковано внутри спасательного плота из расчёта на одного человека? | 1,5 литра на человека |
| 04.1.023 | Внутри плота имеется электролампочка, которая автоматически включается при раскрытии тента. Выключение этой лампочки | Осуществляется при помощи выключателя |
| 04.1.025 | Снабжение спасательной шлюпки должно включать пресную. воду из расчёта… литров на каждого человека | 3 литра |
| 04.2.001 | В штатное снабжение спасательного плота входит |
|
| 04.2.002 | Укажите что из перечисленного в ответах входит в состав снабжения спасательного плота |
|
| 04.2.003 | Укажите инструкции, которые входят в обычное снабжение спасательного плота |
|
| 04.2.004 | Укажите информационные материалы, которые входят в обычное снабжение спасательной шлюпки |
|
| 06.1.001 | Для уменьшения дрейфа и бортовой качки спасательной шлюпки (спасательного плота) следует | Держаться против волны, используя плавучий якорь |
| 06.1.002 | Как используются запасы воды и пищи на спасательном плоту в первые сутки? | 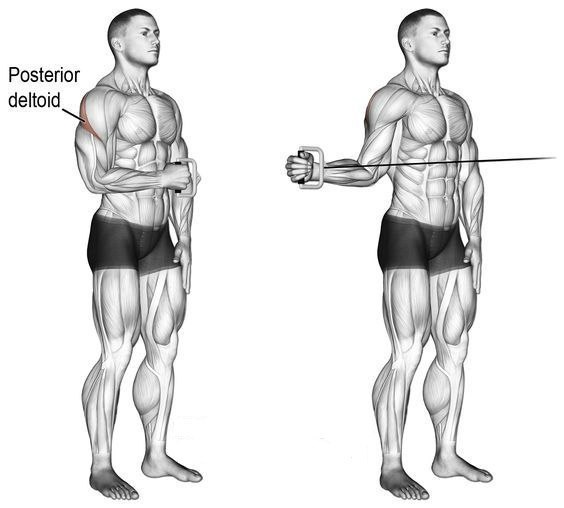 org/Answer»>В режиме экономии org/Answer»>В режиме экономии |
| 06.1.005 | Можно ли пить морскую воду? | Категорически запрещено |
| 06.1.008 | Резкому усилению потерь тепла организмом при нахождении человека в плоту способствует | Ветер |
| 06.1.009 | Известно, что при нахождении человека в воде его тело остывает существенно быстрее чем на воздухе. Во сколько раз вода быстрее забирает тепло человеческого организма, чем воздух? | Более чем в 20 раз |
| 06.1.014 | Экипаж покинул судно, терпящее бедствие. УКВ радиостанция включена в спасательном плоту (шлюпке) и используется для того чтобы привлечь внимание судов, которые могут находиться в данном районе. Укажите, на какой канал должна быть настроена УКВ радиостанция для передачи сигнала бедствия MAYDAY | УКВ канал 16 |
| 06.1.016 | Можно ли нормально дышать внутри плота, если закрыть оба входа? |  org/Answer»>Да org/Answer»>Да |
| 06.1.017 | Можно ли использовать для еды мясо пойманных при нахождении в спасательном средстве морских птиц? | Да, все морские птицы съедобны |
| 06.1.018 | Спасательные вертолёты оборудованы подъёмными устройствами для подбора людей из воды, снятия с аварийного судна или спасательного средства. Во избежание опасного для жизни разряда статического электричества прикасаться к опускаемому с вертолёта средству можно | После того как подъёмное устройство имело контакт с судовыми конструкциями или водой |
| 06.2.001 | Уменьшить влияние жары на людей спасательном средстве можно |
|
| 06.2.002 | Укажите наиболее надёжные признаки, по которым находясь в спасательном средстве, можно определить факт смерти человека |
|
| 06.2.003 | Какое штатное оборудование предназначено для управления закрытой спасательной шлюпкой на ходу? |
|
| 06.2.004 | Укажите меры безопасности, которые целесообразно применять при нахождении в шлюпке в местах возможного появления акул |
|
| 06.2.005 | Для уменьшения потери тепла, находясь в воде, следует |
|
| 06.2.007 | Как спасти от переохлаждения весь экипаж спасательной шлюпки (плота) при отсутствии должного количества теплозащитных мешков |
|
| 06.2.008 | Для того чтобы избежать теплового удара и солнечных ожогов при нахождении в спасательном плоту следует |  org/Answer»> org/Answer»>
|
| 06.2.009 | Что делать, если судороги ноги возникли во время нахождения в воде? |
|
| 06.2.010 | В случае отсутствия питьевой воды на спасательном средстве её можно заменить |
|
| 06.2.011 | Наиболее опасные последствия морской болезни для человека находящегося на аварийном спасательном средстве |
|
| 06.2.012 | Не пригодны для еды морские рыбы |
|
| 07.1.001 | Запас воздуха в дыхательном аппарате, который входит в комплект снаряжения пожарного, должен обеспечить безопасную работу в течение, как минимум | 30 минут |
| 07.1.003 | Согласно требованиям ИМО минимальное количество аварийных дыхательных устройств (аппаратов EEBD — Emergency Escape Breathing Device) на каждой палубе любого типа грузового судна должно составлять | Не менее 2-х |
| 07.1.004 | Аварийные дыхательные устройства (аппараты EEBD — Emergency Escape Breathing Device) должны иметь запас воздуха, достаточный для обеспечения дыхания персонала в течение некоторого времени. Укажите минимально допустимую продолжительность | 10 минут |
| 07.1.007 | Какие меры предосторожности необходимо применять перед входом в помещение с опасными грузами, способными выделять ядовитые, едкие или воспламеняющиеся пары,? | Произвести контроль газовоздушной среды |
 org/Question»>
org/Question»> org/Question»>
org/Question»>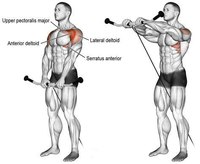 org/Question»>
org/Question»>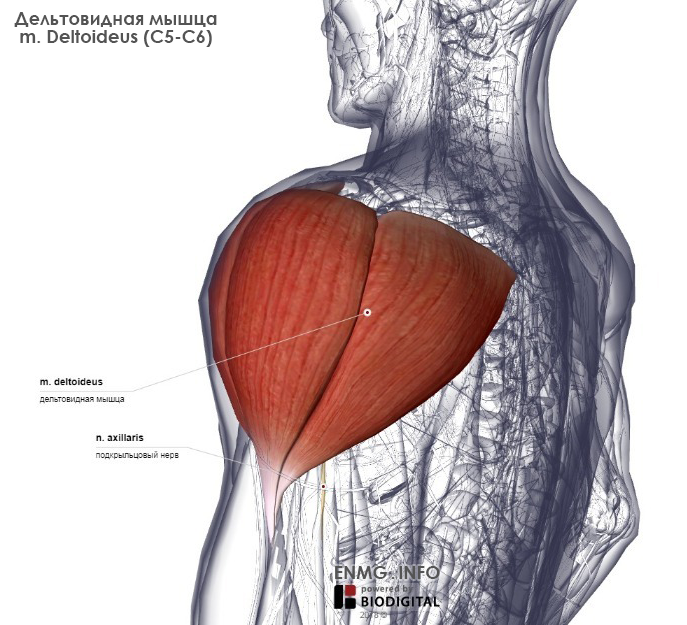 org/Question»>
org/Question»>Упражнение дельта. Качаем заднюю дельту
можно разделить на 2 категории: уп-раж-не-ния на переднюю дельту и упражнения на заднюю и среднюю дельту.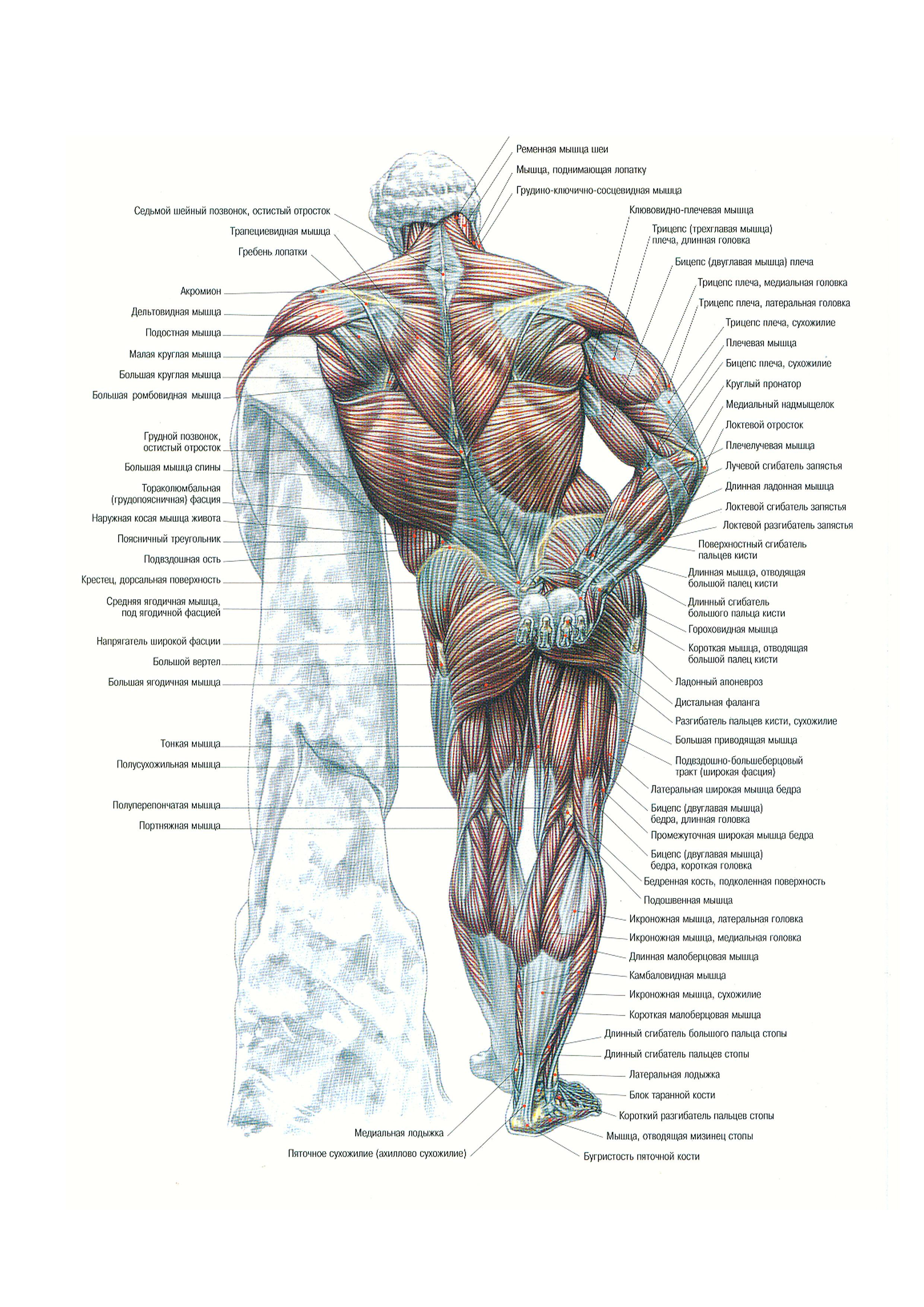 Эффективность упражнений на плечи заключается в их способности це-ле-нап-рав-лен-но «бомбить» целевую мышечную группу. Для передней дельты такими упражнениями являются различные вариации жимов штанги или гантелей, поскольку «украсть» нагрузку у передней дельты некому, в связи с чем, начинающим атлетам ре-ко-мен-ду-ет-ся тре-ни-ро-вать в первую очередь именно этот пучок дельт. Для тренинга средней дельты так же существует базовое упражнение, поэтому и эту дельту можно начинать тренировать с самого начала, а вот заднюю дельту можно качать только изолирующими уп-раж-не-ни-ями, поэтому тренировать её нужно уже на продвинутом уровне, но именно на эту дельту стоит делать упор!
Эффективность упражнений на плечи заключается в их способности це-ле-нап-рав-лен-но «бомбить» целевую мышечную группу. Для передней дельты такими упражнениями являются различные вариации жимов штанги или гантелей, поскольку «украсть» нагрузку у передней дельты некому, в связи с чем, начинающим атлетам ре-ко-мен-ду-ет-ся тре-ни-ро-вать в первую очередь именно этот пучок дельт. Для тренинга средней дельты так же существует базовое упражнение, поэтому и эту дельту можно начинать тренировать с самого начала, а вот заднюю дельту можно качать только изолирующими уп-раж-не-ни-ями, поэтому тренировать её нужно уже на продвинутом уровне, но именно на эту дельту стоит делать упор!
Рекомендуемые материалы: как накачать плечи ; специализация на плечах ; уп-раж-не-ния на плечи ;
Эффективные упражнения на плечи в основном представлены базовыми упражнениями, но это не значит, что изолирующие упражнения не эффективны, просто их имеет смысл применять только на продвинутом уровне. Суть в том, что базовые упражнения создают стресс, позволяют прогрессировать нагрузку, благодаря чему вырабатываются факторы роста, поэтому использовать изоляцию без базы бессмысленно. В тоже время, ис-поль-зо-вать и базовые, и изолирующие упражнения для новичков будет слишком сложно, мышцы просто не «переварят» такую нагрузку, поэтому-то начинающим атлетам и следует использовать только базовые упражнения. В самом начале, когда атлет использует круговые тренировки, упражнение должно быть только 1, через 2-3 месяца, когда атлет перейдет на сплит, количество упражнений можно увеличить до 2-3.
Если речь идет о бодибилдинге, то предпочтительнее использовать упражнения с гантелями, поскольку, при использовании гантель, даже в жимах атлет может за-дей-с-т-во-вать не только передний пучок дельт, но ещё и средний. Рабочей дельтой всегда является та, которая находится над плечевой костью, а использование гантель позволяет так развернуть локти, чтобы над костью находилась не только верхняя, но и средняя дельтовидная мышца.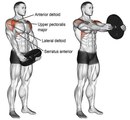 В тягах и махах ключевым моментом, определяющим эффективность упражнений, является положение локтей относительно кистей и локтей относительно корпуса. Так же важно заметить, что, чем шире Вы будете разводить руки, тем большую нагрузку будут получать именно дельты, и, чем уже будет амплитуда, тем большую нагрузку получит трапециевидная мышца.
В тягах и махах ключевым моментом, определяющим эффективность упражнений, является положение локтей относительно кистей и локтей относительно корпуса. Так же важно заметить, что, чем шире Вы будете разводить руки, тем большую нагрузку будут получать именно дельты, и, чем уже будет амплитуда, тем большую нагрузку получит трапециевидная мышца.
Жим штанги стоя – это одно из лучших силовых упражнений, позволяющее не только использовать большой рабочий вес, но ещё и эффективно прогрессировать нагрузку. Не смотря на свою эффективность упражнение является очень травмоопасным, настолько, что его исключили из соревновательной программы, поскольку именно во время выполнения жима штанги стоя очень многие атлеты получали травмы. Этот факт стоит учесть, особенно в том случае, если у Вас слабая спина, в таком случае упражнение лучше выполнять сидя.
Жим Арнольда является так же базовым упражнением, поскольку задействует нес-коль-ко суставов, но это упражнение не предполагает использование большого рабочего веса, поскольку плечевой сустав работает по очень опасной амплитуде. Тем ни менее, это очень эффективное упражнение для плеч, поскольку дельта отрабатывает по всей амплитуде движения, кроме того, нагрузку так же получает и верхний сегмент грудных мышц, для чего, в общем-то, Арнольд Шварценеггер и разработал этот вид жима.
Махи перед собой представляют собой изолирующее упражнение на переднюю дельту, которое можно выполнять двумя руками сразу, или же чередую руки. Плюсом упражнения является то, что оно создает необычную нагрузку для передней дельты, так как обычно передняя дельта выполняет функцию жима, а в данном случае атлет гантель тянет вверх. Упражнение не предполагает «читинга», а для большей отдачи атлету вообще рекомендуется прилагать усилие не только в позитивной фазе, но так же ещё и подконтрольно опускать гантель в исходное положение.
средняя дельтаТяга штанги к подбородку – это одно из лучших упражнений на плечи, поскольку оно одновременно является базовым и целенаправленно грузит средний пучок дельтовидных мышц. В бодибилдинге ключевыми пучками являются средний и задний, поскольку именно они формируют ширину плечевого пояса, так что наличие базового упражнения для развития хотя бы одного из них является очень позитивным моментом. Если вы хотите акцентировать нагрузку именно на средней дельте, Вам в этом упражнении нужно использовать широкий хват.
Махи стоя – это очень эффективное изолирующее упражнение для развития средней дельтовидной мышцы, плюсом которого является возможность «читинговать». Стоит заметить, что «читинговать» начинающим атлетам смысла нет, для того, чтобы было возможно использовать этот суперприем, нужно уметь чувствовать мышцы, но продвинутым атлетам «читинг» позволит нарастить ни один килограмм мышечной массы. Если Вы будете использовать «читинг», тогда Вам нужно особенное внимание уделить разминке , иначе травма неизбежна!
задняя дельтаМахи в наклоне – это, как Вы понимаете, изолирующее упражнение, но для тренировки задней дельты не существует более «базовых» упражнений, поэтому можно сказать, что это условно-базовое упражнение для тренировки задней дельтовидной мышцы. Здесь ключевыми моментами являются те же факторы, что и в махах стоя, плюс к ним добавляется такой фактор, как выведение локтей вперед, что позволяет поднять заднюю дельтовидную мышцу над осью плечевой кости, благодаря чему нагрузка и ложится на заднюю дельту.
Отведение в тренажере – это очень эффективное упражнение для плеч, позволяющее сфокусировать нагрузку на задней дельтовидной мышцы, плюсом является от-но-си-тель-ная легкость. Легкость не в том смысле, что упражнение легко выполнять, а в том, что технически упражнение достаточно простое. Основные моменты, на которые атлет должен обращать внимание, это длина амплитуды движения и скорость выполнения упражнения, поскольку и позитивная и негативная фазы должны быть подконтрольными.
- Главная проблема многих посетителей тренажерных залов, это наличие хорошей передней дельты, неплохой средней дельты и слабого заднего пучка дельтовидной мышцы.

- Знайте, что у вас непропорциональное развитие, если когда вы стоите расслабленным, лицевая сторона ладоней обращена больше назад.
- При выполнении подъемов рук вперед, поднимайте вес на угол 110˚ вместо 90˚, на которых обычно останавливается большинство людей.
- Максимизируйте работу задних дельт, удерживая плечи отведенными по сторонам на угол 30-45˚.
- Большинство тяжелоатлетов выполняют боковые подъемы неправильно. Запястье, локоть и плечо должны быть на одном уровне в верхней точке, и острая сторона локтя должна быть направлена строго назад.
- Люди, имеющие гипертрофированные верхние трапециевидные мышцы, поджимают гантели при боковых подъемах. Во избежание этого, подумайте о выталкивании гантелей от себя.
3D Дельты
Ключом к развитию дельт, имеющих трехмерный внешний вид, — пропорциональное развитие всех трех пучков дельтовидных мышц. Это позволит вашим плечам выглядеть полными, при обзоре спереди, сбоку или сзади.
Косметически, самая распространенная схема неправильного развития дельт — это хорошо развитые передние дельты, среднеразвитые средние дельты и серьезно отстающие задние дельтоиды.
Это обычно проявляется значительным поворотом плеч во внутрь. Другими словами, ваша лицевая сторона ладоней сильнее повернута назад, когда вы расслаблены.
Такое возможно, если:
- вы любите жим штанги лежа и тренировки груди в целом
- вы склонны уделять больше тренировать мышцы, которые видите в зеркале
Еще вариант тренировки плеч без нагрузки на передние дельты, отдавая приоритет развитию средней дельты пока она уже становится толстой, и передняя дельта остается недоразвитой, особенно при виде сбоку.
Оценивайте ваши селфи
Прежде чем начать тренировку плеч, следует выяснить, какой из пучков дельтовидных отстает в развитии. Чтобы расставить приоритеты.
Сделайте свой снимок с руками по сторонам и руками, обращенными к телу. Сделайте дополнительные снимки спереди, сзади и сбоку. Так вы сможете оценить увидеть себя со всех сторон, а потом через некоторое время сравнить результаты тренировок. При виде спереди плечи должны иметь округлый вид.
Если ваши плечи имеют узкую костную структуру и/или у вас широкая структура бедра, то красиво округлые дельты – то, что вам необходимо. Вам следует качать дельты, уделяя особое внимание средним пучкам дельтовидных при помощи боковых подъемов.
С другой стороны, если вы смотрите сбоку, передняя часть плеч недостаточно выпуклая, тогда нужно сосредоточиться на передней дельте. Используйте для тренировки передних дельтовидных мышц разные варианты подъемов гантелей и штанги спереди.
Если нужно накачать задние дельты, используйте тягу гантелей в наклоне для тренировки задней части плеч.
Двойной бицепс сзади
Имейте в виду, когда оцениваете вашу заднюю дельту, это обычно короткая мышца с отличающимся от передней дельты внешним видом. Поэтому не ожидайте от них такого же внешнего вида, как у передних дельт. Чтобы оценить на сколько хорошо прокачана задняя дельта, используйте позу двойной бицепс сзади.
Выстраиваем план тренировки дельтовидных мышц
Давайте вспомним некоторые основы в тренировке плеч, пред тем как погрузиться в их специфику. Я могу предположить, что вы тренируете плечи каждые 5-7 дней.
Тренировать плечи можно в комплексе с другими группами мышц. Но если ваши плечи отстают в развитии, то не качайте их после грудных мышц. Лучше тренировать эти группы в разные дни с перерывом в 2-3 дня. Так у вас будет больше сил и энергии для тренировки плеч. Это позволит подымать больше вес с правильной техникой.
Задняя дельта — это тянущая мышца, поэтому в «день плеч» имеет смысл тренировать дельтовидные вместе со спиной. Потому что задняя дельта функционирует подобно мышцам спины, они помогают тянуть ваши руки вниз и/или назад.
Так что, когда выполняете приведение плеч (подтягивание), растягивание плеч (тяга гантелей), тягу в наклоне (тяга штанги обычным хватом), ваши задние дельты серьезно вовлечены, хотите вы этого или нет.
Упражнения на плечи, Сеты и Повторения
1. Задние дельты
Чтобы накачать заднюю дельтовидную мышцу, должно хватать 3-4 рабочих подходов (без учета разминочного) в изолирующем упражнении.
Может показаться, что одного упражнения мало, но помните, что одна из функций задних дельт – внешнее вращение плечевых костей. Так что, если вы выполняете несколько упражнений с внешним вращением плеч каждую неделю (как должно быть) во время тренировок спины, то несколько сетов одного упражнения, скорее всего, будет достаточно.
Однако если ваши задние дельты серьезно отстают в развитии, тогда выполняйте два упражнения, изолирующих задние дельты. Выполняйте по три сета каждого упражнения. Вот три эффективных упражнения на выбор:
- Разводка на задние дельты в блоке стоя
- Махи или тяги гантелей в наклоне на задние дельты с упор головой в наклонную скамью
- Тяга гантелей в наклоне с вращением Видео с техникой выполнения упражнений
Обратите внимание, что это не упражнение на трицепс. Это движение, при котором вы не сгибаете локти, и тяните рук назад, используя только задние дельты.
Остерегайтесь подъема лопатки, чтобы изолировать заднюю дельту, плечи должны всегда быть опущены вниз. Имейте в виду, что функция задней дельты – просто двигать плечевую кость назад, не сводя лопатки вместе.
Независимо от того, какие упражнения для задних дельт вы выполняете, существует несколько биомеханических принципов, которые следует учитывать. Для начинающих, чтобы оптимизировать работу задних дельт, тренируйте их при оптимальной линии тяги, которая образуется при отведении плеч (по сторонам) на 35-40˚.
Также необходимо поднимать руки с гантелями на одной линии с плечами, направляя мизинцы вверх. Это выровняет заднюю дельту, так чтобы она выполняла максимально возможную работу.
Когда изолируется задняя дельта, проблемой для большинства людей является фактическая изоляция дельты для качественного сокращения мышцы.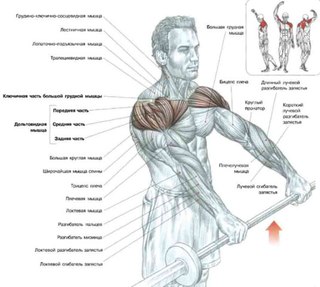 По этой причине чаще чаще выполняют упражнения с легким весом с числом повторений 10-15. Выполнение упражнения с тяжелым весом с небольшим числом повторений более эффективно, но подождите пока улучшиться нервно-мышечная связь задних дельт, прежде чем переходить к большими весами.
По этой причине чаще чаще выполняют упражнения с легким весом с числом повторений 10-15. Выполнение упражнения с тяжелым весом с небольшим числом повторений более эффективно, но подождите пока улучшиться нервно-мышечная связь задних дельт, прежде чем переходить к большими весами.
2. Передние дельты
Жим штанги стоя с груди
Для тренировки передней дельтовидной мышцы обычно используют жим штанги с груди. Это хорошее упражнение для тренировки плеч. Это базовое упражнение на плечи.
Жим на передние дельты можно делать со штангой и с гантелями. Нужно обязательно менять упражнения, чтобы включить в работу стабилизаторы мышц плеча, чтобы плечи были здоровыми
Кроме того, жим для передних дельт, как приседания для квадрицепсов. С другой стороны, подъемы вперед изолируют передние дельты подобно тому, как выпрямление ноги изолирует квадрицепс.
Если передние дельты – ваша сильная сторона, тогда фокусируйтесь на жиме вверх, с периодичными подъемами перед собой в качестве дополнения. Это в комбинации с жимом лежа, все, что необходимо для тренировки дельт.
С другой стороны, если вам нужно увеличить ваши передние дельты, тогда лучше выполнять одно упражнение подъем гантелей или штанги вперед в дополнении к жиму штанги над головой.
Два лучших упражнения для жима вверх – базовый жим штанги вверх (сидя или стоя) и базовый жим гантелей, выполненный сидя или стоя. Хотя многие схемы сет/повторения жима вверх работают, лучше остановиться на 4*6-10, хотя, 5*5 и 3*8-12 тоже хороший вариант, который должен быть выполнен правильно.
Когда речь идет об изоляции передних дельт от задних, вариант подъемов гантелей вперед просто отличный, но подъемы вперед штанги являются более эффективными.
Независимо от того, какой вариант подъемов вперед вы выберете, убедитесь, что гриф находится под углом 110˚, приблизительно на уровне верхней части вашего лба. Это пик, в котором происходит сокращение передних дельт, а не при 90˚, на которых останавливается большинство людей.
При выполнении вариантов подъемов вперед, предпочтительно выбрать диапазон повторений 3*10-12, иногда берите больший вес и выполняйте 6-8 повторений, иногда меньший и делайте 12-15,
3. Средние дельты
Если вы хотите иметь плечи, как у бодибилдеров, вам следует всегда включать варианты подъемов через стороны для подчеркивания средних дельт, наиболее фундаментальным из которых является махи гантелей через стороны.
Несмотря на то, что большинство людей выполняют махи гантелей через стороны, многие делают это неправильно.
Для оптимального напряжения средних пучков дельтовидных мышц, ладони должны быть обращены вниз в верхней точке. Или, если действительно хотите хорошо их нагрузить, наклоните выступающую часть гантелей вниз на несколько градусов, так чтобы внутренняя сторона руки немного повернулась вверх.
Также запястья, локти и плечи в верхней точке должны быть на одном уровне. Кроме того, острая часть вашего локтя должна быть направлена строго назад, а не в пол.
В верхней точке движения, большинство людей склонны наклонять выступающую сторону гантелей слишком высоко, из-за чего их запястья оказываются выше локтей. Это потому что модель этого движения позволяет больше доминировать передней дельте, чтобы помочь вам больше, чем вы того хотите.
Люди с очень сильными верхними трапециевидными мышцами также склонны поджимать гантели при угле финишной позиции близком к 90˚. Чтобы избежать этого, думайте о выталкивании гантелей от себя в стороны.
Для разнообразия, вы можете менять точку максимального напряжения среднего пучка дельтовидной мышцы, выполняя один из следующих вариантов упражнения:
- Лягте на бок на наклонной скамье. Выполняйте махи гантелей, поднимая руку до угла 90˚, где происходит пик сокращения средних дельт.
- Наклоните ваш торс в сторону с весом как на видео:
Если вам необходимо увеличить средние дельты, обычно выполняют три подхода на два различных упражнения на отведение плеч (боковые подъемы). Выполняйте одно упражнение в три сета по 8-12 повторений, и другое в три сета по 12-15. И как обычно, выполняйте иногда более сложные или более легкие.
Выполняйте одно упражнение в три сета по 8-12 повторений, и другое в три сета по 12-15. И как обычно, выполняйте иногда более сложные или более легкие.
Две Непрерывные Тренировки Плеч
Существует две фундаментальные, серьезные программы тренировки плечей, каждая нацелена на различные недостатки плеч.
- Для отстающих средних и задних дельт (но хороших развитых передних дельт):
- Для отстающих передних дельт (но хороших средних дельт):
Что отличает бодибилдера от простого человека? Широкие плечи! Тогда почему ты до сих пор ударно их не качаешь? Как правило, новички начинают думать, что перед накачкой дельт необходимо для начала раскачать грудь и спину. На самом деле это не так. Мощные развитые дельты — это фундамент для последующей проработки больших мышц корпуса. И это утверждение легко объясняется анатомией.
Передние дельты наравне с грудью участвуют в жимах лежа, а задние в той же степени участвуют в тягах, помогая работе широчайших. Если же эти пучки дельт окажутся слабыми и неразвитыми, ни о каком прогрессе в жимах и упражнениях на спину речи быть не может. В этом случае они окажутся слабым звеном, не дающим прогрессировать другим мышечным группам. Именно поэтому дельтами необходимо заниматься с самого первого дня пребывания в спортивном зале. Если никогда их не тренировать, мышечная ткань огрубеет и даже пампингу будет не под силу ее раскачать.
Перед началом тренинга дельт необходимо четко знать их анатомию. Для начала следует понять, что дельтовидные состоят из 3-х пучков: переднего, среднего и заднего. Для каждого из них существуют отдельные упражнения, позволяющие прицельно нагрузить их. Итак, начнем по порядку.
Лучшие упражнения на передние пучки дельт
Анатомическая функция этих пучков представляет собой подъем рук (локтей) перед собой и отведение их к оси тела. Передние пучки активно участвуют в жимовых упражнениях (для груди и плечевого пояса).
Лучшие базовые упражнения на передние пучки дельт:
Лучшие изолированные упражнения на передние дельты:
- Подъемы на нижнем блоке перед собой
Лучшие упражнения на средние пучки дельт
Их анатомическая функция заключается в подъеме рук (локтей) в стороны.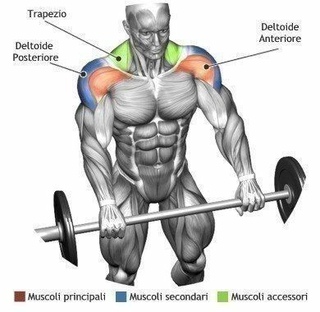 Именно поэтому лучшим упражнением для их прокачки является жим штанги сидя, который сопровождается подъемом локтей кверху.
Именно поэтому лучшим упражнением для их прокачки является жим штанги сидя, который сопровождается подъемом локтей кверху.
Лучшие базовые упражнения на средние дельты:
Лучшие изолированные упражнения на средние дельты:
- Подъем одной рукой в сторону на нижнем блоке
Лучшие упражнения на здание пучки дельт
Анатомическая функция этих пучков заключается в отведении рук назад. Именно поэтому они принимают непосредственное участие во всех тяговых упражнениях на спину. Это объясняется тем, что все тяги предполагают как раз отведение локтей назад. Максимальное напряжение задних пучков дельт наблюдается при отведении рук назад, которые на старте перпендикулярны вертикально оси тела.
Лучшие базовые упражнения на задние дельты:
Лучшие изолированные упражнения на задние дельты:
Оцените полезность статьи:Хотите ли вы иметь массивные плечи, как у лучших бодибилдеров или просто желаете иметь отчетливый рельеф, вам нужно прорабатывать мышцы плеча, делая упражнения на задние дельты.
Прежде чем приступать к выполнению упражнений, нужно разобраться, над чем собственно предстоит работать, давайте рассмотрим анатомию плеча.
Дельтоиды – мышцы, состоящие из трех пучков мышечных волокон:
- передний (фронтальный) пучок;
- средний (боковой) пучок;
- задний (тыльный) пучок.
Каждый из них позволяет нам отводить руку вперед, в сторону или назад соответственно. Косвенно дельты работают во многих упражнениях совсем на другие группы мышц, передний пучок дельт у многих спортсменов хорошо развит благодаря базовым упражнениям на грудные мышцы.
Но для достижения лучших результатов плечи нужно тренировать отдельно, причем, на каждый мышечный пучок есть свои упражнения (изолирующие), потому как средние и задние дельты тренировать крайне непросто.
Мужчины мечтают раскачать широкие плечи, а вот женщины наоборот, боятся стать широкоплечими и мужеподобными. Но спешим вас успокоить, для достижения такого эффекта вам придется ну очень усердно поработать, так как накачать массивные плечи очень непросто.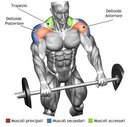 Если вы регулярно будете включать в тренировочную программу упражнения для дельтовидных мышц без больших весов, вы получите красивый рельеф спортивных женских рук и плеч.
Если вы регулярно будете включать в тренировочную программу упражнения для дельтовидных мышц без больших весов, вы получите красивый рельеф спортивных женских рук и плеч.
Тренировка фронтального пучка
Как уже говорилось выше, передние дельты работают во многих упражнениях, где нужно поднять снаряд вверх или перед собой, потому, если вы не преследуете цель сильно раскачать плечи, можете упустить отдельную проработку этого пучка.
Если же вы настроены заполучить широкие массивные плечи, в вашу тренировочную программу должны войти упражнения на передние дельты, изложенные ниже.
Армейский жим (жим штанги с груди) – это базовое упражнение, в нем больше всего задействован передний пучок дельт, чуть меньше – средний и задний, а также мышцы груди и трицепсы.
Есть вариант выполнения жима стоя и сидя. В первом случае включается много мышц-стабилизаторов, но, если у вас проблемы со спиной, вам больше подойдет второй вариант. Обязательно убедитесь, что у вас есть опора в виде спинки скамьи, чтобы снизить нагрузку на межпозвоночные диски.
Выполняется жим следующим образом: зафиксируйте штангу на стойках, зафиксированных примерно на уровне ваших плеч. Возьмите штангу со стоек, положите чуть ниже ключиц, сделайте шаг назад. Стараясь держать спину все время прямой, на вдохе выжмите штангу вверх над головой. Руки в верхней точке должны быть совсем немного присогнуты.
Плавно и осознанно (не роняя) опустите штангу в исходное положение. В процессе выполнения упражнения смотрите только вперед. Если вы начнете блуждать взглядом вверх или в стороны, вероятнее всего, потеряете равновесие. Держите в тонусе мышцы кора на протяжении всей работы.
Жим сидя выполняется аналогично . Для разнообразия можно сменить штангу на гантели, так вы подключите к работе еще больше мелких мышц стабилизаторов.
Подъемы снаряда перед собой – изолированное упражнение на передние дельты. Может выполняться с гантелями, штангой, блином или другим доступным снарядом. Возьмите в руки, например, гантели, сведите их вместе, ноги поставьте на ширине плеч. На вдохе поднимите снаряд до уровня чуть выше параллели с полом. На выдохе подконтрольно опустите руки со снарядом в исходное положение.
Возьмите в руки, например, гантели, сведите их вместе, ноги поставьте на ширине плеч. На вдохе поднимите снаряд до уровня чуть выше параллели с полом. На выдохе подконтрольно опустите руки со снарядом в исходное положение.
Не раскачивайте корпус во время тренинга, так вы распределяете нагрузку между другими группами мышц, а передние дельты остаются непроработанными. Жим гантелей сидя выполняется так: сядьте на скамью, плотно прижав к спину так, чтобы не было ни прогиба, ни сутулости. Возьмите гантели примерно на уровне ушей. Вдыхая, поднимите гантели вверх и немного сведите их вместе, не выворачивая при этом ни кисти, ни руки. На выдохе опустите гантели в исходное положение.
Прокачка средних дельт
Для качественной проработки боковых дельт нужно потратить больше времени и уделить больше внимания изолирующим упражнениям.
Вот некоторые упражнения на средние дельты:
- Жим Арнольда сидя с гантелями — база.
Возьмите снаряды (на 10-12 повторений) обратным хватом и расположите их перед собой на согнутых в локтях руках так, чтобы предплечья были перпендикулярны полу, а гантели – на уровне головы. Выжмите гантели вверх над головой, разворачивая кисти на 180 градусов. Выдыхая, опустите руки в исходное положение, снова разворачивая кисти. - Разводка гантелей в стороны стоя/сидя – изолирующее упражнение.
Станьте перед зеркалом, чтобы контролировать технику. Возьмите в руки гантели, расположите их по бокам от корпуса. Корпус совсем немного наклоните вперед и опустите плечи. Делая вдох, поднимайте гантели до уровня чуть выше плеч. В верхней точке снаряды должны быть немного наклонены вперед. На выдохе плавно опустите снаряды вниз. Не размахивайте руками, как крыльями, делайте осознанные движения, чувствуя, чем вы выполняете каждое из них. - Жим штанги из-за головы – довольно травмоопасное базовое упражнение, которое позволяет проработать средние дельты.
Выполнять его можно стоя и сидя. Для начала стоит попробовать его в Смите, чтобы снизить риск получения травмы.
Для его выполнения опустите штангу примерно до середины затылка. Выжмите снаряд вверх, затем опустите его в исходное положение, но не заваливайте его на плечи. Не стоит опускать штангу слишком низко, это снизит шансы получить травму.
Тренировка тыльного пучка дельт
Эта часть дельтовидных мышц делает плечи визуально шире, если смотреть на них сбоку. Упражнения на дельты для результативности требуют от вас двух вещей: во-первых, четкого соблюдения техники, во-вторых, большого количества повторений. Техника, как и в любом другом упражнении, позволяет нагрузить те мышцы, которые мы хотим, и не дать нагрузке разойтись по мышцам спины.
Большое количество повторений важно, потому что задние дельты хорошо приспособлены к длительным нагрузкам, их нужно долго и нудно закачивать.
А делать вы это будете в таких упражнениях:
- Разводка на наклонной скамье.
С гантелями лягте животом вниз на скамью, зафиксированную под углом 30 градусов к горизонту. Руки с гантелями выведите перед собой (держите гантели прямым хватом). Выполняйте разводку рук, при этом поворачивая кисти на 90 градусов. - Тяга штанги (гантелей) лежа на наклонной скамье.
Положение тела такое же, как в предыдущем упражнении. Разница в том, что здесь вы держите перед собой штангу в исходном положении, из которого поднимаете ее до уровня груди. В верхней точке ваши предплечья должны образовать с плечами прямой угол - Тяга в блоке/кроссовере.
Взяв закрепленную рукоять тренажера, поднимите руки, до параллели с полом. Тяните на себя рукоять, отводя назад локти и сохраняя положение рук, параллельное полу.
Тренировать дельты можно раз в неделю. Если у вас они быстро восстановятся, через два-три дня можно добавить еще одну тренировку отдельно на дельты или на плечи совместно с другой мышечной группой.
Обязательно включайте в программу проработки плеча базовые упражнения для дельты, но начинайте с изоляционных.

Подбирайте такой вес снарядов, чтобы вы могли соблюдать технику выполнения упражнения для дельт, но после тренировки ваши руки с трудом поднимали шейкер с гейнером.
Обязательно разогревайте все тело перед тренировкой, но особенно – плечевые и локтевые суставы во избежание получения травм.
Когда у мужчины широкие и массивные плечи – это смотрится мужественно и красиво. Но, к сожалению, мало кто из мужчин знает, как правильно тренироваться, чтобы раскачать большие плечи. Поэтому, сейчас я вам расскажу: как накачать средние и задние дельты и покажу лучшие упражнения .
Дельтовидные мышцы (дельты) – это поверхностные мышцы плеча, которые несут ответственность за выполнение движения рук в плечевом суставе (двигают вверх, вниз, вправо, влево, по кругу). Данные мышцы состоят из трех пучков:
- передний
- средний
- задний
Как вы уже поняли из названия, в данной статье мы затронем только 2 пучка (средний и задний). Передний трогать не будем, по той причине, что у подавляющего большинства людей данный пучок хорошо развит (у некоторых даже слишком хорошо). Конечно, чтобы плечо было большим и гармоничным нужно прокачивать все 3 пучка. Но, если передний преобладает в развитии, то на нем можно не акцентировать нагрузку. Свою нагрузку он получит при выполнении различных жимов.
В большей степени, именно средние и задние дельты дают ширину и массивность плечам. Если вы игнорируете данные участки, то никогда не сможете раскачать большие плечи. Очень часто спортсмены забывают про заднюю дельту (не делают на нее специализированных упражнений), и в итоге получается такое полу обрезанное плечо (не гармонично развитое).
Сразу скажу, что накачать среднюю и заднюю часть плеча можно не только в тренажерном зале, но и в домашних условиях . Для этого вам понадобятся гантели. В данной статье я приведу ряд эффективных упражнений для средних и задних дельт, а так же дам вам несколько рабочих тренировочных программ для тренировок в домашних условиях и в тренажерном зале.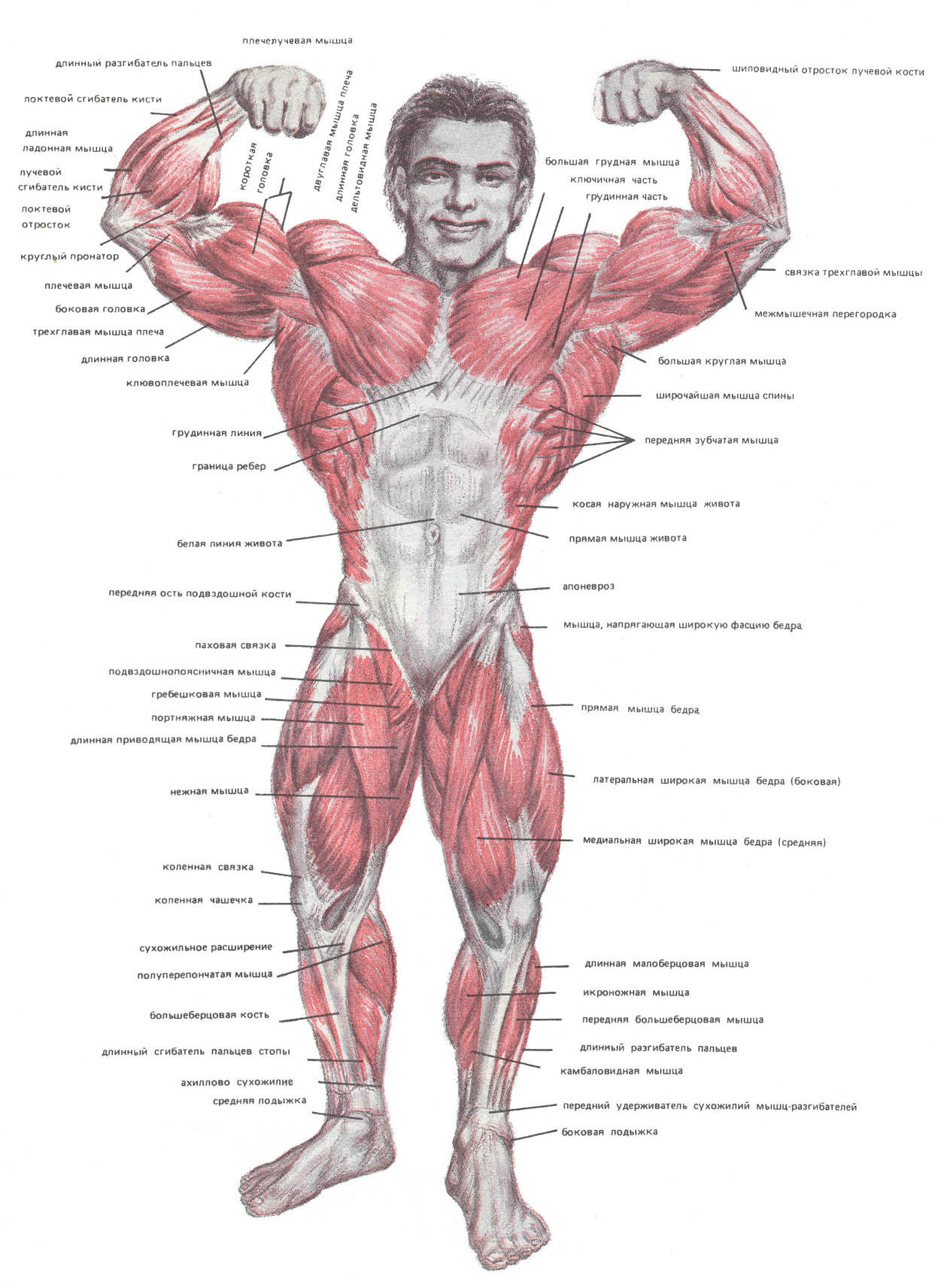 Вам остается только выбрать подходящую схему, под ваши характеристики.
Вам остается только выбрать подходящую схему, под ваши характеристики.
Для начала давайте рассмотрим основные причины, по которым ваши плечи не растут:
№1. Отсутствие целевой нагрузки на нужные участки.
Как правило, когда не опытный человек тренирует плечи, то в его программе присутствует усиленная нагрузка на переднюю дельту, на порядок меньше нагрузки на среднюю и минимум на заднюю (или на заднюю дельту нагрузки вообще может не наблюдаться). Банальная программа на плечи не опытного спортсмена может выглядеть так:
В лучшем случае:
№2. Тяга штанги к подбородку узким хватом (передняя дельта)
№3. Махи гантелями в стороны (средняя дельта)
№4. Махи гантелями в наклоне (задняя дельта)
В худшем случае:
№1. Армейский жим штанги стоя (передняя дельта)
№2. Махи гантелями перед собой (передняя дельта)
№3. Тяга штанги к подбородку узким хватом (передняя дельта)
№4. Махи гантелями в стороны (средняя дельта)
Как видите, основная нагрузка ложится именно на передние дельты, которые и так хорошо развиты. Привел такие примеры по той причине, что я примерно понимаю, как строятся программы у людей, так как работаю персональным тренером.
№2. Отсутствие базовых упражнений.
Я работал с очень многими людьми, и на своей практике очень редко встречал таких спортсменов, которые используют базовые упражнения на средние и задние дельты. Если на средние пучки еще дают базовую нагрузку в виде жимов штанги (или гантелей) из-за головы, то на задние пучки плеча базовые тяги практически никто не делает. Чаще всего средние и задние пучки дельт получают свою нагрузку в виде изолированных упражнений (махи гантелями, махи в блоке). Изолированные упражнения – это, конечно, хорошо. Но, без базы толку будет мало. Так как именно базовые упражнения с большими весами дают мощный толчок в плане роста.
№3. Слишком легкие рабочие веса и слишком много повторений.
Вы, наверное, слышали, как многие говорят, что плечи мелкие мышечные группы, которые любят многоповторку, и не нужно брать большой вес – главное чувствовать. Да, я согласен с тем, что нужно чувствовать мышечную группу. Но, на счет многоповторки – это бред. Дельты такие же мышцы, как и грудь, спина, трицепс. Чтобы они росли, их нужно тяжело тренировать. Я не отрицаю того, что каждый индивидуален и кому то больше подойдет тренинг в многоповторном режиме. Но, у подавляющего большинства дельты начнут расти только тогда, когда в программе будут присутствовать тяжелые базовые упражнения на 6 – 8 повторений + тренинг в отказ.
Да, я согласен с тем, что нужно чувствовать мышечную группу. Но, на счет многоповторки – это бред. Дельты такие же мышцы, как и грудь, спина, трицепс. Чтобы они росли, их нужно тяжело тренировать. Я не отрицаю того, что каждый индивидуален и кому то больше подойдет тренинг в многоповторном режиме. Но, у подавляющего большинства дельты начнут расти только тогда, когда в программе будут присутствовать тяжелые базовые упражнения на 6 – 8 повторений + тренинг в отказ.
Вы можете совмещать многоповторку и малоповторку. Это будет еще лучше. Например, можно делать базовые упражнения на 6 – 8 повторений, а изолированные на 10 – 12 (возможно 15) повторений. В этом случае вы получите необходимый стресс для дальнейшего мышечного роста + эффект от пампинга.
№4. Нет прогрессии нагрузок.
Если ваша схема тренировок и рабочий вес на снарядах не меняются месяцами, то и вашим дельтам незачем расти. Чтобы был рост, нужен новый стресс, который создается . У новичков это очень частая ошибка.
№5. Вы не чувствуете мышечное сокращение.
Как правило, такое происходит тогда, когда человек пренебрегает правильной техникой выполнения упражнений. Если вы только пришли в тренажерный зал, то не нужно гнаться за весами. Берите мелкий вес и отрабатывайте технику. Сначала научитесь чувствовать мышечную группу, а потом начинайте работать с тяжелыми весами.
Можно разобрать еще более мелкие ошибки, но тогда статья получится очень длинная. Поэтому, остановимся на 5-ти самых важных. Ну что ж, теперь перейдем к самым эффективным упражнениям на плечи. Сразу скажу, что существует много разных упражнений, и все перечислять я не буду. Сделаю акцент только на самые лучшие (мое мнение).
Лучшие упражнения на средние дельты:
№1. Тяга штанги к подбородку широким хватом (базовое упражнение)
№2. Жим гантелей из-за головы (базовое упражнение)
№3. Жим штанги из-за головы (базовое упражнение)
№4. Махи гантелями в стороны (изолированное упражнение)
№5. Боковые махи в блочной раме (изолированное упражнение)
Боковые махи в блочной раме (изолированное упражнение)
Лучшие упражнения на задние дельты:
№1. Тяга штанги в наклоне с акцентом на задние дельты (базовое упражнение)
№2. Тяга штанги за спиной с акцентом на задние дельты (базовое упражнение)
№3. Обратные разведения в тренажере для грудных мышц (изолированное упражнение)
№4. Махи гантелями в наклоне (изолированное упражнение)
№5. Горизонтальная протяжка в блоке (изолированное упражнение)
Хочу сказать, что каждое из этих упражнений задействует все 3 пучка дельт. Но, акцентированная большая часть нагрузки уходит именно в целевую мышцу. К примеру, тяга штанги к подбородку широким хватом максимально воздействует на средние дельты, в меньшей степени на задние и минимум нагрузки получают передние. Поэтому, данное упражнение относим к лучшим упражнениям на среднюю часть плеча.
Теперь пришло время разобрать самые эффективные программы тренировок для роста больших и массивных плеч. Разберем тренировки как в тренажерке, так и дома.
Как накачать средние и задние дельты в тренажерном зале:
Программа №1
В данной схеме мы совмещаем среднюю часть плеча с тренировкой грудных мышц, а заднюю часть плеча с тренировкой спины. Данную программу нельзя использовать новичкам, стаж которых меньше 6 месяцев (для таких спортсменов существую свои программы).
Грудные мышцы + Средние дельты:
Упражнение на грудь №1
Упражнение на грудь №2
Упражнение на грудь №3
Мышцы спины + Задние дельты:
Упражнение на спину №1
Упражнение на спину №2
Упражнение на спину №3
Обратные разведения в тренажере для грудных мышц – 1*15/3*8
Программа №2
В данной схеме плечи тренируются в отдельный день. Идеальный вариант для тех людей, у кого средний и задний пучок дельт сильно отстают в росте. Первым упражнением ставьте ту дельту, которая отстает больше (в данной программе я поставил первым упражнением заднюю дельту, так как в большинстве случаев именно она развита хуже всего). Противопоказано новичкам.
Противопоказано новичкам.
Тяга штанги в наклоне с акцентом на задние дельты – 1*15/1*10/3*6
Тяга штанги к подбородку широким хватом – 1*15/1*10/3*6
Тяга штанги за спиной с акцентом на задние дельты – 1*15/3*8
Жим гантелей из-за головы – 1*15/3*8
Махи гантелями в наклоне – 3*10/1*15
Махи гантелями в стороны – 3*10/1*15
Горизонтальная протяжка в блоке – 3*15
Боковые махи в блочной раме – 3*15
Как накачать средние и задние дельты в домашних условиях:
Программа №1
Данная программа подойдет новичкам, так как здесь присутствует умеренная нагрузка, которая даст нужный стресс для роста мышц и не приведет к перетренированности.
Жим гантелей из-за головы – 1*20/1*15/3*8
Махи гантелями в стороны – 3*12
Тяга гантелей в наклоне с акцентом на задние дельты – 1*20/1*15/3*8
Махи гантелями в наклоне – 3*12
Программа №2
Данная программа подойдет продвинутым спортсменам, которые добились хоть каких-то успехов в плане мышечного роста. Новичкам противопоказано, так как может привести к перетренированности.
Влияние человека на морфологию дельты в глобальном масштабе привело к чистому увеличению площади суши
Дельты рек входят в число наиболее экономически и экологически ценных сред на Земле. Даже при отсутствии подъема уровня моря дельты становятся все более уязвимыми для прибрежных опасностей, поскольку уменьшение количества наносов и изменение климата изменяют их запас наносов, влияя на морфологию дельты и, возможно, приводя к эрозии. Однако взаимосвязь между балансом отложений в дельте, океанографическими силами волн и приливов и морфологией дельты остается плохо определенной количественно.Здесь мы показываем, как морфология около 11000 прибрежных дельт по всему миру, от небольших дельт залива до мегадельт, пострадала от строительства плотин на реках и обезлесения. Мы представляем модель, которая показывает, что морфология современной дельты варьируется в зависимости от преобладания волн (около 80 процентов), приливов (около 10 процентов) и рек (около 10 процентов), но что наиболее крупные дельты — это приливы и реки. — преобладают. За последние 30 лет, несмотря на повышение уровня моря, дельты во всем мире испытали чистый прирост земли в размере 54 ± 12 квадратных километров в год (2 стандартных отклонения), причем на 1 процент крупнейших дельт приходится 30 процентов всей чистой суши. прирост площади.Люди являются важной движущей силой этого чистого прироста земель — 25 процентов роста дельты можно отнести к вызванному обезлесением увеличению поступления речных наносов. Тем не менее, для почти 1000 дельт строительство плотин привело к серьезному (более чем на 50 процентов) сокращению потока антропогенных наносов, вызвав коллективную потерю 12 ± 3,5 квадратных километров в год (2 стандартных отклонения) дельтовых земель. Не все дельты теряют землю в результате строительства плотин на реках: дельты, переходящие к преобладанию приливов, в настоящее время приобретают сушу, вероятно, из-за заполнения русел.Однако с учетом ожидаемого ускоренного подъема уровня моря недавние приросты суши вряд ли сохранятся на протяжении XXI века. Понимание перераспределения отложений волнами и приливами будет иметь решающее значение для успешного прогнозирования антропогенных изменений дельт как на местном, так и на глобальном уровне.
— преобладают. За последние 30 лет, несмотря на повышение уровня моря, дельты во всем мире испытали чистый прирост земли в размере 54 ± 12 квадратных километров в год (2 стандартных отклонения), причем на 1 процент крупнейших дельт приходится 30 процентов всей чистой суши. прирост площади.Люди являются важной движущей силой этого чистого прироста земель — 25 процентов роста дельты можно отнести к вызванному обезлесением увеличению поступления речных наносов. Тем не менее, для почти 1000 дельт строительство плотин привело к серьезному (более чем на 50 процентов) сокращению потока антропогенных наносов, вызвав коллективную потерю 12 ± 3,5 квадратных километров в год (2 стандартных отклонения) дельтовых земель. Не все дельты теряют землю в результате строительства плотин на реках: дельты, переходящие к преобладанию приливов, в настоящее время приобретают сушу, вероятно, из-за заполнения русел.Однако с учетом ожидаемого ускоренного подъема уровня моря недавние приросты суши вряд ли сохранятся на протяжении XXI века. Понимание перераспределения отложений волнами и приливами будет иметь решающее значение для успешного прогнозирования антропогенных изменений дельт как на местном, так и на глобальном уровне.
Гамма-дельта (γδ) Т-клеток: друг или враг в развитии рака? | Журнал трансляционной медицины
Борн В., Майлз К., Уайт Дж., О’Брайен Р., Фрид Дж. Х., Маррак П., Капплер Дж., Кубо Р. Т..Пептидные последовательности дельта- и гамма-цепей Т-клеточного рецептора идентичны предсказанным X- и гамма-белкам. Природа. 1987; 330: 572–4.
CAS PubMed Статья Google Scholar
Hayday AC, Saito H, Gillies SD, Kranz DM, Tanigawa G, Eisen HN, Tonegawa S. Структура, организация и соматическая перестройка гамма-генов Т-клеток. Клетка. 1985. 40: 259–69.
CAS PubMed Статья Google Scholar
Holtmeier W, Kabelitz D. γδ Т-клетки связывают врожденный и адаптивный иммунные ответы. В: Kabelitz D, Schröder JM, редакторы. Механизмы эпителиальной защиты, т. 86. Базель: Karger Publishers; 2005. с. 151–83.
γδ Т-клетки связывают врожденный и адаптивный иммунные ответы. В: Kabelitz D, Schröder JM, редакторы. Механизмы эпителиальной защиты, т. 86. Базель: Karger Publishers; 2005. с. 151–83.
Google Scholar
Вантуроут П., Хейдей А. Шесть лучших: уникальный вклад гаммадельта-Т-клеток в иммунологию. Nat Rev Immunol. 2013; 13: 88–100.
CAS PubMed PubMed Central Статья Google Scholar
Дар А.А., Патил Р.С., Чиплункарь СВ. Понимание взаимосвязи между толлинг-подобными рецепторами и ответами гамма-дельта-Т-клеток. Фронт Иммунол. 2014; 5: 366.
PubMed PubMed Central Статья CAS Google Scholar
Bonneville M, Chen ZW, Dechanet-Merville J, Eberl M, Fournie JJ, Jameson JM, et al. Чикаго 2014-30 годы гаммадельта Т-клеток. Cell Immunol. 2015; 296: 3–9.
CAS PubMed Статья Google Scholar
Ямасаки А., Ониши Х., Морисаки Т., Катано М. Индукция цитотоксических Т-лимфоцитов импульсными гаммадельта-клетками CEA, выделенными от пациентов с запущенным раком. Anticancer Res. 2011; 31: 2419–24.
CAS PubMed Google Scholar
Рей М., Пеннингтон Д. Д., Сильва-Сантос Б. Возникающая роль Т-лимфоцитов гаммадельта в опухолях: значение для иммунотерапии рака. Cancer Res. 2015; 75: 798–802.
CAS PubMed Статья Google Scholar
Сильва-Сантос Б., Серре К., Норелл Х. Т-клетки гаммадельта при раке. Nat Rev Immunol. 2015; 15: 683–91.
CAS PubMed Статья Google Scholar
Сильва-Сантос Б. Содействие ангиогенезу в микроокружении опухоли: секретная жизнь лимфоидных IL-17-продуцирующих мышиных Т-клеток гаммадельта.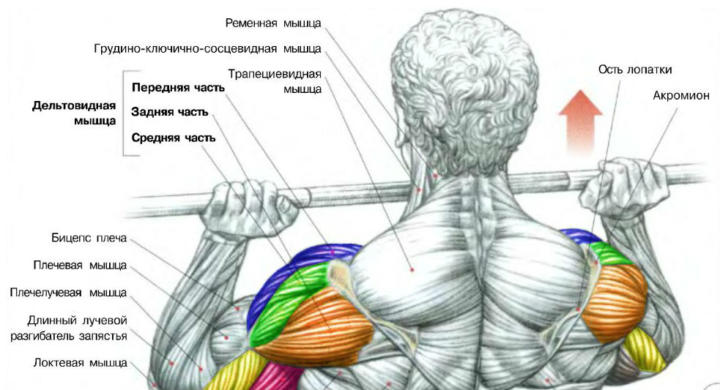 Eur J Immunol. 2010; 40: 1873–6.
Eur J Immunol. 2010; 40: 1873–6.
CAS PubMed Статья Google Scholar
Патил Р.С., Шах С.У., Шриханде С.В., Гоэль М., Дикшит Р.П., Чиплункар С.В. Гаммадельта-Т-клетки, продуцирующие IL17, вызывают ангиогенез и связаны с плохой выживаемостью у пациентов с раком желчного пузыря. Int J Cancer. 2016; 139: 869–81.
PubMed Статья CAS Google Scholar
Ву Х, Ян Т., Лю Х, Го Дж. Н., Се Т, Дин И и др. IL-17 способствует ангиогенезу опухоли посредством активации пути Stat3 VEGF при раке желудка.Tumor Biol. 2016; 37: 5493–501.
CAS Статья Google Scholar
Пан Б., Шен Дж., Цао Дж., Чжоу Й., Шан Л., Джин С. и др. Интерлейкин-17 способствует ангиогенезу, стимулируя выработку VEGF раковыми клетками через сигнальный путь STAT3 / GIV при немелкоклеточном раке легкого. Научный доклад 2015; 5: 16053.
CAS PubMed PubMed Central Статья Google Scholar
Ян Дж., Хуанг Дж. Врожденные клетки γδT17 преобразуют вызванное раком воспаление в иммуносупрессию через клетки-супрессоры миелоидного происхождения. Онкоиммунология. 2014; 3: e953423.
PubMed PubMed Central Статья Google Scholar
Kong X, Sun R, Chen Y, Wei H, Tian Z. Клетки γδT приводят в действие иммунотолерантность, вызываемую миелоидными клетками-супрессорами, опосредованное CD8 + T-клетками при вирусе гепатита B. J Immunol. 2014; 193: 1645–53.
CAS PubMed Статья Google Scholar
Wu P, Wu D, Ni C, Ye J, Chen W., Hu G и др. Клетки GammadeltaT17 способствуют накоплению и размножению миелоидных супрессорных клеток при колоректальном раке человека. Иммунитет. 2014; 40: 785–800.
CAS PubMed PubMed Central Статья Google Scholar
Qu P, Wang LZ, Lin PC. Расширение и функции миелоидных супрессорных клеток в микроокружении опухоли.Cancer Lett. 2016; 380: 253–6.
CAS PubMed Статья Google Scholar
Каракашева Т.А., Уолдрон Т.Дж., Ерусланов Э., Ким С.Б., Ли Дж.С., О’Брайен С., Хикс П.Д., Басу Д., Сингхал С., Малаваси Ф., Рустги А.К. CD38-экспрессирующие миелоидные супрессорные клетки способствуют росту опухоли на мышиной модели рака пищевода. Cancer Res. 2015; 75: 4074–85.
CAS PubMed PubMed Central Статья Google Scholar
Chafe SC, Lou Y, Sceneay J, Vallejo M, Hamilton MJ, McDonald PC, Bennewith KL, Moller A, Dedhar S. Карбоангидраза IX способствует мобилизации миелоидных клеток-супрессоров и созданию метастатической ниши путем стимуляции выработки G-CSF . Cancer Res. 2015; 75: 996–1008.
CAS PubMed Статья Google Scholar
Кабелиц Д., Маришен Л., Оберг Н-Х, Хольтмайер В., Веш Д. Защита эпителия с помощью γδ Т-клеток.Int Arch Allergy Immunol. 2005. 137: 73–81.
CAS PubMed Статья Google Scholar
Willcox CR, Pitard V, Netzer S, Couzi L, Salim M, Silberzahn T, Moreau JF, Hayday AC, Willcox BE, Dechanet-Merville J. Наблюдение за цитомегаловирусом и опухолевым стрессом путем связывания гаммадельта T человека рецептор клеточного антигена к рецептору эндотелиального протеина С. Nat Immunol. 2012; 13: 872–9.
CAS PubMed Статья Google Scholar
Wu D, Wu P, Qiu F, Wei Q, Huang J. Подмножества гаммадельтаТ-клеток человека и их участие в опухолевом иммунитете. Cell Mol Immunol. 2017; 14: 245–53.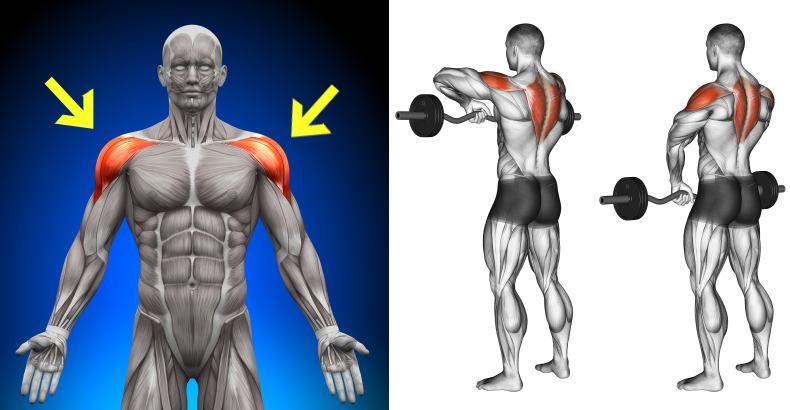
CAS PubMed Статья Google Scholar
Adams EJ, Gu S, Luoma AM. Гамма-дельта-Т-клетки человека: эволюция и распознавание лигандов. Cell Immunol. 2015; 296: 31–40.
CAS PubMed PubMed Central Статья Google Scholar
Димова Т., Брауэр М., Госселин Ф., Тассиньон Дж., Лео О., Доннер С. и др. Эффекторные Т-клетки Vgamma9Vdelta2 доминируют в репертуаре фетальных Т-клеток гаммадельта человека. Proc Natl Acad Sci USA. 2015; 112: E556–65.
CAS PubMed PubMed Central Статья Google Scholar
Ди Карло Э, Бокка П., Эмионит Л, Чилли М., Чиполлоне Дж., Моранди Ф. и др. Механизмы противоопухолевой активности Т-клеток Vgamma9Vdelta2 человека в сочетании с золедроновой кислотой на доклинической модели нейробластомы.Mol Ther. 2013; 21: 1034–43.
PubMed PubMed Central Статья CAS Google Scholar
Braza MS, Klein B. Противоопухолевая иммунотерапия с использованием Т-лимфоцитов Vgamma9Vdelta2: от скамьи к постели. Br J Haematol. 2013; 160: 123–32.
CAS PubMed Статья Google Scholar
Алмейда А.Р., Коррейя Д.В., Фернандес-Платцгуммер А., да Силва С.Л., да Силва М.Г., Аньос Д.Р. и др.Delta one T-клетки для иммунотерапии хронического лимфолейкоза: расширение / дифференциация клинического уровня и доклиническое подтверждение концепции. Clin Cancer Res. 2016; 22: 5795–804.
CAS PubMed Статья Google Scholar
Siegers GM, Dhamko H, Wang XH, Mathieson AM, Kosaka Y, Felizardo TC, et al. Человеческие Т-клетки Vdelta1 gammadelta, размноженные из периферической крови, проявляют специфическую цитотоксичность в отношении В-клеточных клеток, происходящих из хронического лимфоцитарного лейкоза. Цитотерапия. 2011; 13: 753–64.
Цитотерапия. 2011; 13: 753–64.
CAS PubMed Статья Google Scholar
Манган Б.А., Данн М.Р., О’Рейли В.П., Данн П.Дж., Эксли М.А., О’Ши Д. и др. Передний край: ограничение CD1d и секреция цитокинов Th2 / Th3 / Th27 Т-клетками Vdelta3 человека. J Immunol. 2013; 191: 30–4.
CAS PubMed PubMed Central Статья Google Scholar
Ramirez K, Witherden DA, Havran WL.Все руки на DE (T) C: резидентные в эпителии γδ Т-клетки отвечают на повреждение ткани. Cell Immunol. 2015; 296: 57–61.
CAS PubMed PubMed Central Статья Google Scholar
Siegers GM, Lamb LS Jr. Цитотоксические и регуляторные свойства циркулирующих Т-клеток Vdelta1 + gammadelta: новый игрок в области клеточной терапии? Mol Ther. 2014; 22: 1416–22.
CAS PubMed PubMed Central Статья Google Scholar
Зал JE. Учебник медицинской физиологии Гайтона и Холла. Филадельфия: Elsevier Health Sciences; 2015.
Google Scholar
Веш Д., Петерс С., Зигерс Г.М. Гамма-дельта-Т-регуляторные клетки человека при раке: факт или вымысел? Фронт Иммунол. 2014; 5: 598.
PubMed PubMed Central Статья CAS Google Scholar
Пол С., Лал Г. Регуляторные и эффекторные функции гамма-дельта (γδ) Т-клеток и их терапевтический потенциал в адоптивной клеточной терапии рака.В J Cancer. 2016; 139: 976–85.
CAS Google Scholar
Йе Дж, Ма К., Ван Ф., Сюэ ЕС, Тот К., Хуанг Й. и др. Специфическое привлечение регуляторных Т-клеток гаммадельта при раке груди человека. Cancer Res. 2013; 73: 6137–48.
CAS PubMed PubMed Central Статья Google Scholar
Lafont V, Sanchez F, Laprevotte E, Michaud H-A, Gros L, Eliaou J-F, et al.Пластичность клеток γδT: влияние на противоопухолевый ответ. Фронт Иммунол. 2014; 5: 622.
PubMed PubMed Central Статья CAS Google Scholar
Presti LE, Dieli F, Meraviglia S. Проникающие в опухоль гаммадельта-Т-лимфоциты: патогенная роль, клиническое значение и дифференциальное программирование в микросреде опухоли. Фронт Иммунол. 2014; 5: 607.
PubMed PubMed Central Статья CAS Google Scholar
Caccamo N, La Mendola C, Orlando V, Meraviglia S, Todaro M, Stassi G и др. Дифференциация, фенотип и функция продуцирующих интерлейкин-17 человеческих Т-клеток Vgamma9Vdelta2. Кровь. 2011; 118: 129–38.
CAS PubMed Статья Google Scholar
Ness-Schwickerath KJ, Jin C, Morita CT. Требования к цитокинам для дифференцировки и размножения человеческих Т-клеток Vgamma2 Vdelta2, продуцирующих IL-17A и IL-22. J Immunol.2010. 184: 7268–80.
CAS PubMed PubMed Central Статья Google Scholar
Wesch D, Glatzel A, Kabelitz D. Дифференциация покоящихся γδ Т-клеток периферической крови человека по Th2- или Th3-фенотипу. Cell Immunol. 2001; 212: 110–7.
CAS PubMed Статья Google Scholar
Казетти Р., Аграти С., Уоллес М., Сакки А., Мартини Ф, Мартино А. и др.Передний край: TGF-beta1 и IL-15 индуцируют FOXP3 + гаммадельта-регуляторные Т-клетки в присутствии стимуляции антигеном. J Immunol. 2009; 183: 3574–7.
CAS PubMed Статья Google Scholar
Коррейя Д.В., Фогли М., Хадспет К., да Силва М.Г., Мавилио Д., Сильва-Сантос Б. Дифференциация Т-клеток Vdelta1 + периферической крови человека, экспрессирующих природный рецептор цитотоксичности NKp30 для распознавания лимфоидных лейкозных клеток.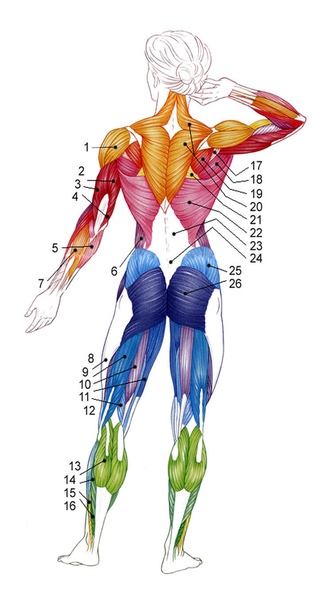 Кровь. 2011; 118: 992–1001.
Кровь. 2011; 118: 992–1001.
CAS PubMed Статья Google Scholar
Дево С., Руссо Б., Нетцер С., Питар В., Паруассен С., Хайраллах С. и др. Антиметастатический потенциал человеческих Т-клеток Vdelta1 (+) гаммадельта в ортотопической мышиной модели ксенотрансплантата карциномы толстой кишки. Cancer Immunol Immunother. 2013; 62: 1199–210.
CAS PubMed Статья Google Scholar
Peng G, Wang HY, Peng W., Kiniwa Y, Seo KH, Wang RF. Проникающие в опухоль гаммадельта-Т-клетки подавляют функцию Т- и дендритных клеток с помощью механизмов, контролируемых уникальным сигнальным путем толл-подобных рецепторов. Иммунитет. 2007. 27: 334–48.
CAS PubMed Статья Google Scholar
Niu C, Jin H, Li M, Xu J, Xu D, Hu J, et al. Анализ in vitro пролиферативной способности и цитотоксических эффектов ex vivo индуцированных естественных клеток-киллеров, индуцированных цитокинами клеток-киллеров и гамма-дельта Т-клеток.BMC Immunol. 2015; 16:61.
PubMed PubMed Central Статья CAS Google Scholar
Дхар С., Чипланкар С.В. Лизис сенсибилизированных аминобисфосфонатом опухолевых клеток молочной железы MCF-7 Т-клетками Vgamma9 Vdelta2. Cancer Immun. 2010; 10: 10.
PubMed PubMed Central Google Scholar
Viey E, Fromont G, Escudier B, Morel Y, Da Rocha S, Chouaib S, et al.Активированные фосфостимом γδ Т-клетки убивают аутологичную метастатическую почечно-клеточную карциному. J Immunol. 2005; 174: 1338–47.
CAS PubMed Статья Google Scholar
Александр А.А., Маниар А., Каммингс Дж.С., Хеббелер А.М., Шульце Д.Х., Гастман Б.Р. и др. Активированные изопентенилпирофосфатом CD56 + γδ Т-лимфоциты проявляют сильную противоопухолевую активность в отношении плоскоклеточной карциномы человека. Clin Cancer Res. 2008; 14: 4232–40.
Clin Cancer Res. 2008; 14: 4232–40.
CAS PubMed PubMed Central Статья Google Scholar
Todaro M, D’Asaro M, Caccamo N, Iovino F, Francipane MG, Meraviglia S и др. Эффективное уничтожение стволовых клеток рака толстой кишки человека γδ Т-лимфоцитами. J Immunol. 2009. 182: 7287–96.
CAS PubMed Статья Google Scholar
Dieli F, Vermijlen D, Fulfaro F, Caccamo N, Meraviglia S, Cicero G, et al. Нацеливание на человеческие γδ Т-клетки с золедронатом и интерлейкином-2 для иммунотерапии гормонорезистентного рака простаты.Cancer Res. 2007. 67: 7450–7.
CAS PubMed PubMed Central Статья Google Scholar
Dokouhaki P, Schuh NW, Joe B., Allen CA, Der SD, Tsao MS, et al. NKG2D регулирует продукцию растворимого TRAIL размноженными ex vivo человеческими гаммадельта-Т-клетками. Eur J Immunol. 2013; 43: 3175–82.
CAS PubMed Статья Google Scholar
Li Z, Xu Q, Peng H, Cheng R, Sun Z, Ye Z.IFN-γ увеличивает чувствительность клеточных линий HOS и U2OS к опосредованному γδ T-клеткам уничтожению через лигандный путь Fas / Fas. Int Immunopharmacol. 2011; 11: 496–503.
CAS PubMed Статья Google Scholar
Фишер Дж. П., Ян М., Хойджерджанс Дж., Картер Л., Аболхассани А., Фрош Дж. И др. Убийство нейробластомы Vdelta2- и Vdelta2-отрицательных gammadeltaT-клеток после размножения искусственными антигенпрезентирующими клетками.Clin Cancer Res. 2014; 20: 5720–32.
CAS PubMed PubMed Central Статья Google Scholar
Couzi L, Pitard V, Sicard X, Garrigue I, Hawchar O, Merville P и др. Зависимая от антител антицитомегаловирусная активность Т-клеток гаммадельта человека, экспрессирующих CD16 (FcgammaRIIIa). Кровь. 2012; 119: 1418–27.
Кровь. 2012; 119: 1418–27.
CAS PubMed Статья Google Scholar
Zheng J, Guo Y, Ji X, Cui L, He W. Новый антителоподобный слитый белок TCRgammadelta-Ig проявляет противоопухолевую активность против карциномы яичников человека. Cancer Lett. 2013; 34: 1150–8.
Google Scholar
Гао Й., Ян В., Пан М., Скалли Е., Жирарди М., Аугенлихт Л. Х. и др. Т-клетки γδ являются одним из первых источников интерферона γ при противоопухолевом иммунитете. J Exp Med. 2003; 198: 433–42.
CAS PubMed PubMed Central Статья Google Scholar
Ramstead AG, Джутила Массачусетс. Комплексная роль цитокинов и факторов роста Т-клеток гаммадельта в развитии рака. J Interferon Cytokine Res. 2012; 32: 563–9.
CAS PubMed PubMed Central Статья Google Scholar
Li H, Luo K, Pauza CD. TNF-α является положительным регуляторным фактором для Т-клеток Vγ2Vδ2 человека. J Immunol. 2008. 181: 7131–7.
CAS PubMed PubMed Central Статья Google Scholar
Sedlak C, Patzl M, Saalmuller A, Gerner W. IL-12 и IL-18 индуцируют продукцию гамма-интерферона и de novo экспрессию CD2 в Т-клетках гаммадельта свиней. Dev Comp Immunol. 2014; 47: 115–22.
CAS PubMed Статья Google Scholar
Хуанг И, Гетахун А., Хайзер Р.А., Детанико Т.О., Ависзус К., Кирхенбаум Г.А. и др. Gammadelta T-клетки формируют доиммунные популяции периферических B-клеток. J Immunol. 2016; 196: 217–31.
CAS PubMed Статья Google Scholar
Caccamo N, Todaro M, La Manna MP, Sireci G, Stassi G, Dieli F. IL-21 регулирует дифференцировку субпопуляции Т-клеток гаммадельта человека, обладающих активностью хелперов В-клеток. PLoS ONE. 2012; 7: e41940.
PLoS ONE. 2012; 7: e41940.
CAS PubMed PubMed Central Статья Google Scholar
Bansal RR, Mackay CR, Moser B, Eberl M. IL-21 усиливает потенциал Т-клеток гаммадельта человека по оказанию помощи В-клеткам. Eur J Immunol. 2012; 42: 110–9.
CAS PubMed Статья Google Scholar
Брандес М., Виллиманн К., Мозер Б. Профессиональная функция презентации антигена Т-клетками гаммадельта человека. Наука. 2005; 309: 264–8.
CAS PubMed Статья Google Scholar
Муто М., Багдади М., Маэкава Р., Вада Х., Сейно К. Миелоидные молекулярные характеристики человеческих гаммадельта-Т-клеток подтверждают их приобретение опухолевой антиген-презентирующей способности.Cancer Immunol Immunother. 2015; 64: 941–9.
CAS PubMed Статья Google Scholar
Мао Ц., Моу Х, Чжоу И, Юань Г, Сюй Ц., Лю Х. и др. Активированные опухолью TCRγδ + Т-клетки от пациентов с раком желудка индуцируют противоопухолевый иммунный r-ответ TCRαβ + Т-клеток через их антигенпредставляющие клеточно-подобные эффекты. J Immunol Res. 2014; 2014: 593562.
PubMed PubMed Central Статья CAS Google Scholar
Van Acker HH, Anguille S, Van Tendeloo VF, Lion E. Расширение возможностей гамма-дельта-Т-клеток с противоопухолевым иммунитетом с помощью иммунотерапии на основе дендритных клеток. Онкоиммунология. 2015; 4: e1021538.
PubMed PubMed Central Статья CAS Google Scholar
Мозер Б., Эберл М. Т-АПК гаммадельты: новый инструмент для иммунотерапии? Cell Mol Life Sci. 2011; 68: 2443–52.
CAS PubMed Статья Google Scholar
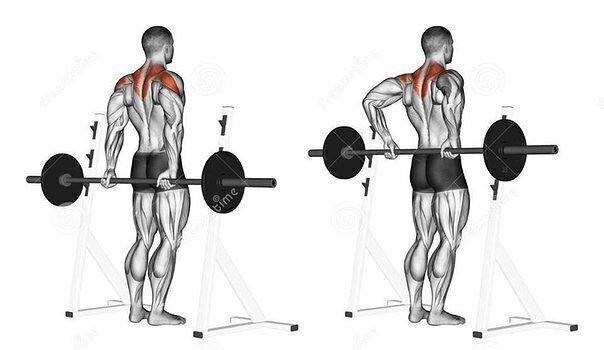
Munz C, Steinman RM, Fujii S. Созревание дендритных клеток с помощью врожденных лимфоцитов: скоординированная стимуляция врожденного и адаптивного иммунитета. J Exp Med. 2005; 202: 203–7.
PubMed PubMed Central Статья CAS Google Scholar
Конти Л., Касетти Р., Кардоне М., Варано Б., Мартино А., Беларделли Ф. и др. Взаимное активирующее взаимодействие между дендритными клетками и γδ T-клетками, стимулированными памидронатом: роль CD86 и воспалительных цитокинов.J Immunol. 2005; 174: 252–60.
CAS PubMed Статья Google Scholar
Devilder MC, Maillet S, Bouyge-Moreau I, Donnadieu E, Bonneville M, Scotet E. Усиление антиген-стимулированной продукции цитокинов V гамма 9V дельта 2 Т-клетками незрелыми дендритными клетками (DC) и обратный эффект по созреванию постоянного тока. J Immunol. 2006; 176: 1386–93.
CAS PubMed Статья Google Scholar
Maniar A, Zhang X, Lin W, Gastman BR, Pauza CD, Strome SE и др. Человеческие γδ Т-лимфоциты индуцируют сильную противоопухолевую цитотоксичность, опосредованную NK-клетками, через взаимодействие с CD137. Кровь. 2010; 116: 1726–33.
CAS PubMed PubMed Central Статья Google Scholar
Вакита Д., Сумида К., Ивакура Ю., Нисикава Х., Окури Т., Чамото К. и др. Проникающие в опухоль гаммадельта-Т-клетки, продуцирующие IL-17, поддерживают прогрессирование опухоли, способствуя ангиогенезу.Eur J Immunol. 2010; 40: 1927–37.
CAS PubMed Статья Google Scholar
Кимура Ю., Нагаи Н., Цунекава Н., Сато-Мацусита М., Йошимото Т., Куа Д.Д. и др. CD30 (+) Vdelta1 Т-клетки, продуцирующие IL-17A, управляют индуцированным воспалением прогрессированием рака. Cancer Sci. 2016; 107: 1206–14.
2016; 107: 1206–14.
CAS PubMed PubMed Central Статья Google Scholar
Ян Дж., Вайнберг РА. Эпителиально-мезенхимальный переход: на перекрестке развития и метастазирования опухоли. Dev Cell. 2008; 14: 818–29.
CAS PubMed Статья Google Scholar
Kuhl AA, Pawlowski NN, Grollich K, Blessenohl M, Westermann J, Zeitz M, et al. Периферические Т-клетки гаммадельта человека обладают регуляторным потенциалом. Иммунология. 2009; 128: 580–8.
PubMed PubMed Central Статья CAS Google Scholar
Ронг Л., Ли К., Ли Р., Лю Х.М., Сунь Р., Лю XY. Анализ инфильтрирующих опухоль гамма-дельта Т-клеток при раке прямой кишки. Мир Дж. Гастроэнтерол. 2016; 22: 3573–80.
CAS PubMed PubMed Central Статья Google Scholar
Вистуба-Хампрехт К., Ди Бенедетто С., Шиллинг Б., Сукер А., Шадендорф Д., Гарбе С. и др. Фенотипическая характеристика и прогностическое влияние циркулирующих Т-клеток гаммадельта и алфавита при метастатической злокачественной меланоме.Int J Cancer. 2016; 138: 698–704.
CAS PubMed Статья Google Scholar
Wistuba-Hamprecht K, Martens A, Haehnel K, Foppen MG, Yuan J, Postow MA, et al. Доля переносимых кровью Vδ1 + и Vδ2 + Т-клеток связана с общей выживаемостью пациентов с меланомой, получавших ипилимумаб. Eur J Cancer. 2016; 64: 116–26.
CAS PubMed PubMed Central Статья Google Scholar
Мао И, Инь С., Чжан Дж., Ху И, Хуан Б., Цуй Л. и др. Новый эффект ИЛ-4 на Т-клетки гаммадельта человека: стимулирование регуляторных Т-клеток Vdelta1 посредством продукции ИЛ-10 и ингибирование функции Т-клеток Vdelta2.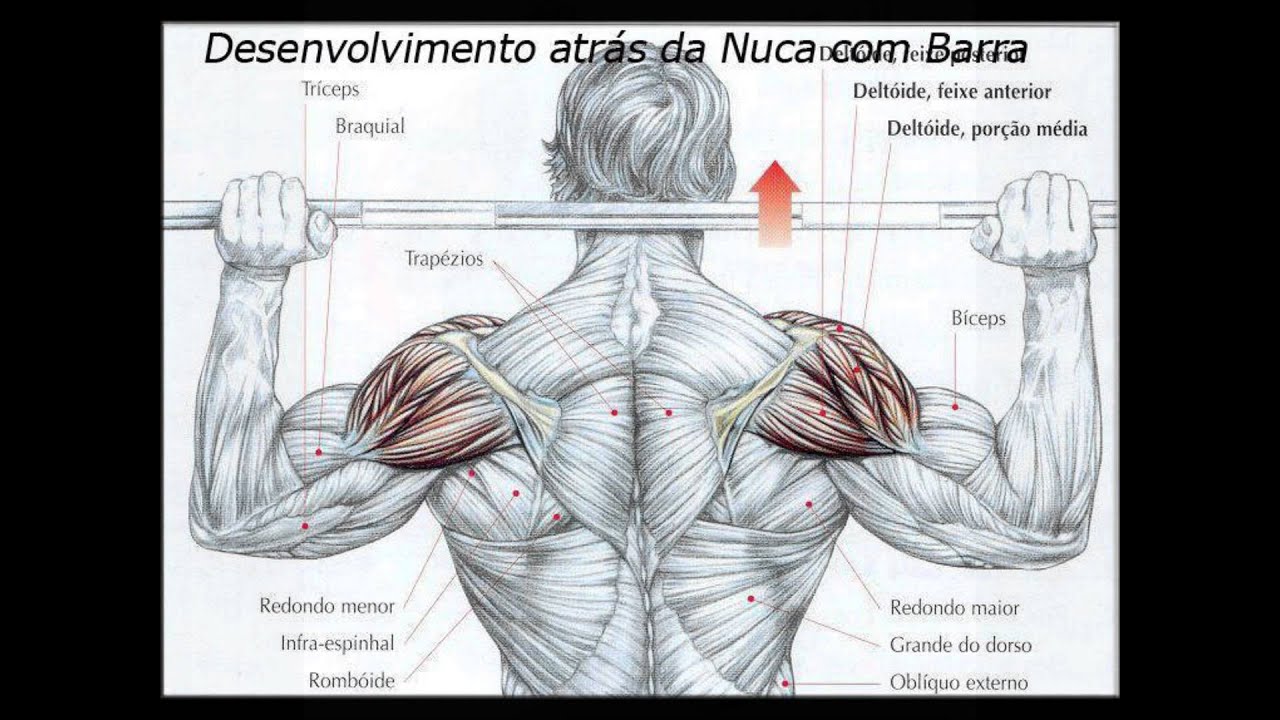 Cell Mol Immunol. 2016; 13: 217–28.
Cell Mol Immunol. 2016; 13: 217–28.
CAS PubMed Статья Google Scholar
Дейли Д., Замбиринис С.П., Зейферт Л., Аккад Н., Мохан Н., Верба Г. и др. Т-клетки гаммадельта поддерживают онкогенез поджелудочной железы, сдерживая активацию Т-клеток алфавита.Клетка. 2016; 66: 1485–99.
Артикул CAS Google Scholar
Йе Дж, Ма К., Сюэ ЕС, Эйкхофф К.С., Чжан Ю., Варварес М.А. и др. Регуляторные Т-клетки γδ опухолевого происхождения подавляют врожденный и адаптивный иммунитет за счет индукции иммунного старения. J Immunol. 2013; 190: 2403–14.
CAS PubMed PubMed Central Статья Google Scholar
Ли Дж, Лю Дж, Мао Х, Тан Цюй, Лу Х.Блокада рецептора IL-7 ингибирует продуцирующие IL-17 гаммадельта-клетки и подавляет развитие меланомы. Воспаление. 2014; 37: 1444–52.
CAS PubMed Статья Google Scholar
Welte T, Zhang XH. Интерлейкин-17 может способствовать прогрессированию рака груди на нескольких стадиях заболевания. Медиаторы Inflamm. 2015; 2015: 804347.
PubMed PubMed Central Статья CAS Google Scholar
Cai Y, Xue F, Fleming C, Yang J, Ding C, Ma Y, et al. Дифференциальная потребность в развитии и периферическая регуляция дермальных клеток Vgamma4 и Vgamma6T17 при здоровье и воспалении. Nat Commun. 2014; 5: 3986.
CAS PubMed PubMed Central Статья Google Scholar
Бунт С.К., Синха П., Клементс В.К., Лейпс Дж., Остранд-Розенберг С. Воспаление индуцирует происходящие из миелоида супрессорные клетки, которые способствуют прогрессированию опухоли.J Immunol. 2006; 176: 284–90.
CAS PubMed Статья Google Scholar
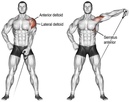
Рей М., Гонсалвес-Соуза Н., Ланка Т., Томпсон Р.Г., Менсурадо С., Балквилл Ф.Р. и др. Мышиные CD27 (-) Vgamma6 (+) гаммадельта Т-клетки, продуцирующие IL-17A, способствуют росту рака яичников за счет мобилизации малых перитонеальных макрофагов протопухи. Proc Natl Acad Sci USA. 2014; 111: E3562–70.
CAS PubMed PubMed Central Статья Google Scholar
Coffelt SB, Kersten K, Doornebal CW, Weiden J, Vrijland K, Hau CS и др. Гаммадельта-Т-клетки и нейтрофилы, продуцирующие IL-17, участвуют в сговоре, способствуя метастазированию рака груди. Природа. 2015; 522: 345–8.
CAS PubMed PubMed Central Статья Google Scholar
Саббионе Ф., Габеллони М.Л., Эрнст Дж., Гори М.С., Саламоне Дж., Олеастро М. и др. Нейтрофилы подавляют функцию Т-клеток гаммадельта. Eur J Immunol. 2014; 44: 819–30.
CAS PubMed Статья Google Scholar
Калян С., Чандрасекаран В., Квабиус Е.С., Линдхорст Т.К., Кабелиц Д. Поглощение азот-бисфосфонатов нейтрофилами приводит к подавлению гаммадельта-Т-клеток периферической крови человека. Cell Mol Life Sci. 2014; 712: 335–46.
Google Scholar
Кобаяши Х., Танака Ю. Т-клеточная иммунотерапия гаммадельта — обзор.Фармацевтические препараты (Базель). 2015; 8: 40–61.
CAS Статья Google Scholar
Накадзима Дж., Муракава Т., Фуками Т., Гото С., Канеко Т., Йошида Ю. и др. Исследование фазы I адоптивной иммунотерапии для пациентов с рецидивирующим немелкоклеточным раком легкого с аутологичными гаммадельта-Т-клетками. Eur J Cardiothorac Surg. 2010; 37: 1191–7.
PubMed Статья Google Scholar
Сакамото М., Накадзима Дж., Муракава Т., Фуками Т., Ёсида Ю., Мураяма Т. и др. Адоптивная иммунотерапия распространенного немелкоклеточного рака легкого с использованием увеличенных золедронатом гаммадельта Т-клеток: клиническое исследование фазы I. J Immunother. 2011; 34: 202–11.
CAS PubMed Статья Google Scholar
Ногучи А., Канеко Т., Камигаки Т., Фудзимото К., Одзава М., Сайто М. и др. Активированная золедронатом Т-клеточная терапия Vgamma9gammadelta возможна и восстанавливает повреждение T-клеток гаммадельта у пациентов с солидными опухолями.Цитотерапия. 2011; 13: 92–7.
CAS PubMed Статья Google Scholar
Ланг Дж. М., Кайкобад М. Р., Уоллес М., Стааб М. Дж., Хорват Д. Л., Уилдинг Г. и др. Пилотное испытание интерлейкина-2 и золедроновой кислоты для увеличения гаммадельта-Т-клеток в качестве лечения пациентов с рефрактерной почечно-клеточной карциномой. Cancer Immunol Immunother. 2011; 60: 1447–60.
CAS PubMed PubMed Central Статья Google Scholar
Кобаяши Х., Танака Й., Яги Дж., Минато Н., Танабе К. Исследование фазы I / II адоптивного переноса Т-клеток гаммадельта в комбинации с золедроновой кислотой и ИЛ-2 пациентам с прогрессирующей почечно-клеточной карциномой. Cancer Immunol Immunother. 2011; 60: 1075–84.
CAS PubMed Статья Google Scholar
Идзуми Т., Кондо М., Такахаши Т., Фуджиэда Н., Кондо А., Тамура Н. и др. Ex vivo характеристика репертуара Т-клеток гаммадельта у пациентов после адоптивного переноса Т-клеток Vgamma9 Vdelta2, экспрессирующих бета-цепь рецептора интерлейкина-2 и общую гамма-цепь.Цитотерапия. 2013; 15: 481–91.
CAS PubMed Статья Google Scholar
Какими К., Мацусита Х., Муракава Т., Накадзима Дж. Т-клеточная терапия гаммадельта для лечения немелкоклеточного рака легкого. Перевод Lung Cancer Res. 2014; 3: 23–33.
CAS PubMed PubMed Central Google Scholar
Беннуна Дж., Бомпас Э., Нейдхардт Э.М., Роллан Ф., Филип И., Галеа С. и др.Фаза I исследования Innacell gammadelta, аутологичного продукта клеточной терапии, высокообогащенного Т-лимфоцитами gamma9delta2, в комбинации с ИЛ-2, у пациентов с метастатической почечно-клеточной карциномой. Cancer Immunol Immunother. 2008; 57: 1599–609.
CAS PubMed Статья Google Scholar
Вильгельм М., Сметак М., Шефер-Эккарт К., Киммель Б., Биркманн Дж., Эйнселе Н. и др. Успешный адоптивный перенос и экспансия in vivo гаплоидентичных Т-клеток гаммадельта.J Transl Med. 2014; 12:45.
PubMed PubMed Central Статья CAS Google Scholar
Fournié J-J, Sicard H, Poupot M, Bezombes C, Blanc A, Romagné F, et al. Какие уроки можно извлечь из испытаний иммунотерапии рака на основе γδ Т-лимфоцитов? Cell Mol Immunol. 2013; 10: 35–41.
PubMed Статья CAS Google Scholar
Märten A, von Lilienfeld-Toal M, Büchler MW, Schmidt J.Растворимая МПК повышена в сыворотке пациентов с карциномой поджелудочной железы, что снижает цитотоксичность Т-лимфоцитов. Int J Cancer. 2006; 119: 2359–65.
PubMed Статья CAS Google Scholar
Lo Presti E, Toia F, Oieni S, Buccheri S, Turdo A, Mangiapane LR, et al. Плоскоклеточные опухоли привлекают Т-клетки гаммадельта, продуцирующие IL17 или IFNgamma, в зависимости от стадии опухоли. Cancer Immunol Res. 2017; 5: 397–407.
CAS PubMed Статья Google Scholar
Kunzmann V, Smetak M, Kimmel B, Weigang-Koehler K, Goebeler M, Birkmann J, et al. Роль гаммадельта-Т-клеток в опухолевой иммунотерапии против опухолевой антагонизма: результаты проспективного исследования фазы I / II. J Immunother. 2012; 35: 205–13.
CAS PubMed Статья Google Scholar
Du SH, Li Z, Chen C, Tan WK, Chi Z, Kwang TW, et al. Совместная экспансия индуцированных цитокинами клеток-киллеров и Т-клеток Vgamma9Vdelta2 для терапии CAR Т-клетками.PLoS ONE. 2016; 11: e0161820.
PubMed PubMed Central Статья CAS Google Scholar
Chen HC, Joalland N, Bridgeman JS, Alchami FS, Jarry U, Khan MWA и др. Синергетическое нацеливание на стволовые клетки рака молочной железы человеческими гаммадельта-Т-клетками и CD8 + Т-клетками. Immunol Cell Biol. 2017. https://doi.org/10.1038/icb.2017.21.
PubMed Central Статья PubMed Google Scholar
Хуанг Y, Мацумура Y, Hatano S, Noguchi N, Murakami T., Iwakura Y, et al. IL-21 подавляет ответ Т-клеток гаммадельта, продуцирующих IL-17A, после инфицирования Bacillus Calmette-Guerin посредством индукции апоптоза. Врожденный иммунитет. 2016; 22: 588–97.
CAS PubMed Статья Google Scholar
Wang X, Zhao X, Feng C, Weinstein A, Xia R, Wen W, et al. IL-36gamma трансформирует микроокружение опухоли и способствует противоопухолевым иммунным ответам, опосредованным лимфоцитами 1 типа.Раковая клетка. 2015; 28: 296–306.
CAS PubMed PubMed Central Статья Google Scholar
Ли В., Кубо С., Окуда А., Ямамото Х., Уэда Х., Танака Т. и др. Влияние ИЛ-18 на рост Т-клеток гаммадельта, стимулированного золедронатом и ИЛ-2. J Immunother. 2010. 33: 287–96.
CAS PubMed Статья Google Scholar
Ли В., Ямамото Х., Кубо С., Окамура Х.Модуляция врожденного иммунитета IL-18. J Reprod Immunol. 2009; 83: 101–5.
CAS PubMed Статья Google Scholar
Van Acker HH, Anguille S, Willemen Y, Van den Bergh JM, Berneman ZN, Lion E, et al. Интерлейкин-15 усиливает пролиферацию, стимулирующий фенотип и противоопухолевые эффекторные функции гамма-дельта-Т-клеток человека. J Hematol Oncol. 2016; 9: 101.
PubMed PubMed Central Статья CAS Google Scholar
Prendergast GC, Smith C, Thomas S, Mandik-Nayak L, Laury-Kleintop L, Metz R, et al. Индолеамин-2,3-диоксигеназные пути патогенного воспаления и иммунного спасения при раке. Cancer Immunol Immunother. 2014; 63: 721–35.
CAS PubMed PubMed Central Статья Google Scholar
Gomes AQ, Correia DV, Grosso AR, Lanca T, Ferreira C, Lacerda JF, et al. Идентификация панели из десяти антигенов белков клеточной поверхности, связанных с иммунным нацеливанием лейкозов и лимфом на гаммадельта-Т-клетки периферической крови.Haematologica. 2010; 95: 1397–404.
CAS PubMed PubMed Central Статья Google Scholar
Mondragon L, Kroemer G, Galluzzi L. Иммуносупрессивные Т-клетки гаммадельта способствуют канцерогенезу поджелудочной железы. Онкоиммунология. 2016; 5: e1237328.
PubMed PubMed Central Статья Google Scholar
Li Z, Li N, Li F, Zhou Z, Sang J, Chen Y и др.Белки иммунной контрольной точки PD-1 и TIM-3 оба высоко экспрессируются в тканях печени и коррелируют с их полиморфизмом генов у пациентов с гепатоцеллюлярной карциномой, связанной с HBV. Медицина. 2016; 95: e5749.
CAS PubMed PubMed Central Статья Google Scholar
He Y, Yu H, Rozeboom L, Rivard CJ, Ellison K, Dziadziuszko R, et al. Экспрессия белка LAG-3 при немелкоклеточном раке легкого и ее связь с PD-1 / PD-L1 и лимфоцитами, инфильтрирующими опухоль.J Thorac Oncol. 2017; 12: 814–23.
PubMed Статья Google Scholar
Lines JL, Pantazi E, Mak J, Sempere LF, Wang L, O’Connell S, et al. VISTA — это молекула иммунной контрольной точки для человеческих Т-клеток. Cancer Res. 2014; 74: 1924–32.
CAS PubMed PubMed Central Статья Google Scholar
Дельта — дельты и деятельность человека — отложения, земля, уровень и вода
Дельты были важными центрами человеческой деятельности на протяжении всей истории, отчасти из-за плодородия земли и легкого доступа к транспорту.Многие ранние человеческие цивилизации развивались на дельтах. Например, дельта реки Нил является домом для египетских культур более семи тысяч лет.
Дельты содержат большие площади водно-болотных угодий, где быстро накапливается органическое вещество. Следовательно, дельтовые буровые растворы очень богаты органическими веществами и являются хорошими источниками углеводородов горных пород при заглублении на соответствующие глубины. Неудивительно, что дельтовые отложения содержат обширные запасы угля, нефти и газа.Дельтовые песчаные тела также являются отличными коллекторами для подвижных углеводородов. Такое сочетание факторов делает дельты, возможно, наиболее важной углеводородной средой на Земле, . Благодаря этому экономическому процветанию современные и древние дельты, вероятно, были изучены более тщательно, чем любая другая осадочная среда .
Дельты очень низкого рельефа; большинство районов редко превышают несколько футов над уровнем моря. Следовательно, они содержат пресноводные, солоноватоводные и соленые бассейны с соответственно разнообразной и сложной экологией.Незначительные изменения высоты поверхности дельты могут затопить районы водой с гораздо большей или меньшей соленостью, поэтому на экологию дельты легко повлияет деятельность человека. Как указано выше, люди значительно изменили дельты и будут продолжать делать это в надежде сдержать наводнение , наводнение . В результате мы продолжим видеть ускоренное отступление дельты и разрушение водно-болотных угодий, если люди не разработают новые технологии борьбы с наводнениями или новые методы защиты водно-болотных угодий.
Книги
Лидер, Майк. Седиментология и осадочные бассейны: от Турбулентность к тектонике. Лондон: Blackwell Science. 1999.
Селби, М.Дж. Изменяющаяся поверхность Земли. Лондон: Oxford University Press, 1985.
Скиннер, Брайан Дж. И Стивен С. Портер. Динамическая Земля: Введение в физическую геологию. 4-е изд. Джон Вили и сыновья, 2000.
Турман, Гарольд В. и Алан П. Трухильо. Основы Океанография. 7-е изд. Englewood Cliffs, NJ: Prentice Hall, 2001.
| Функциональный ключ | Позиция (я) | Описание Действия | Графическое изображение | Длина |
|---|---|---|---|---|
В этом подразделе раздела «Последовательность» описываются природные варианты белковой последовательности. Естественный вариант i VAR_083465 | 18 | Q → QIGGQ в NEDBAS; неизвестное патологическое значение. Ручное утверждение, основанное на эксперименте i
| 1 | |
| Естественный вариант i VAR_083466 | 19-723 | Отсутствует в NEDBAS. Ручное утверждение, основанное на эксперименте i
| 705 | |
| Естественный вариант i VAR_083467 | 77-723 | Отсутствует в NEDBAS. Ручное утверждение, основанное на эксперименте i
| 647 | |
| Естественный вариант i VAR_083468 | 179 | C → F в NEDBAS. Ручное утверждение, основанное на эксперименте i
| 1 | |
| Естественный вариант i VAR_083469 | 498-723 | Отсутствует в NEDBAS. Ручное утверждение, основанное на эксперименте i
| 226 | |
| Естественный вариант i VAR_083470 | 509-723 | Отсутствует в NEDBAS; неизвестное патологическое значение. Ручное утверждение, основанное на эксперименте i
| 215 |
Повышенная мезиотемпоральная дельта-активность характеризует виртуальную навигацию у людей Этюдник с проходом техники в брюшном дыхании, а также в хирургии на спине и дыхании, легко и практично.
Nous avons étudié rétrospectivement 30 cas de NLPC consécutifs dans un seul center par un seul chirurgien.Les huit premiers пациентов ont été opérés en décubitus ventral и 22 suivants en décubitus dorsal, selon la техника де Вальдивия. Les données ont été analysées par test de Student. Результаты исследований по значению для p <0,05.
Лечебные хирургические осложнения, переливание крови и другие сопоставимые инфекции, связанные с двумя группами и другими медицинскими препаратами. Le taux de réussite en décubitus dorsal était de 72% и 63% en décubitus ventral ( p > 0,05).Cela correpondait aux résultats publiés dans la littérature. Существенная статистика различий, en faveur du groupe décubitus dorsal, Concernait la durée opératoire ( p = 0,02).
Notre étude a mis en évidence que la néphrolithotomie en décubitus dorsal était une альтернативная система и эффективность в классической технике en décubitus ventral. La Courbe d’apprentissage a été rapide.
Для оценки безопасности и эффективности чрескожной нефролитотомии, выполняемой в положении лежа на спине.Проверить, легко ли изменить операционную технику между положением лежа на животе и положением на спине.
Тридцать пациентов, перенесших чрескожную нефролитотомию одним хирургом в одной больнице, были изучены ретроспективно. Первые восемь случаев были выполнены в положении лежа, а следующие 22 пациента были прооперированы в положении лежа на спине в соответствии с операционной техникой Вальдивии. Данные были проанализированы с помощью теста t . Мы считали P <0,05 значимым.
Частота хирургических осложнений, частота переливаний и частота лихорадки были одинаковыми в обеих группах. Частота успеха составила 72% в группе положения лежа на спине и 63% в группе положения лежа ( P > 0,05). Эти результаты сопоставимы с данными литературы. Только время операции было короче в группе положения лежа на спине со значительной статистической разницей ( P = 0,02).
По нашему опыту, чрескожная нефролитотомия в положении лежа на спине была безопасной и эффективной, а результаты были аналогичны таковым в положении лежа.Процесс обучения был легким и быстрым.
Антропогенный авульсия в дельте Рейн-Маас перед исторической набережной (Нидерланды) | Геология
Результаты показывают, что антропогенные воздействия — оседание торфяников и увеличение нагрузки взвешенных наносов — на реку, эстуарий и торфяники сделали нижнюю дельту более склонной к отрыву. Чувствительность системы к воздействию человека в значительной степени может быть объяснена тем, что торф склонен к уплотнению.Отрывы были успешными благодаря взаимодействующим петлям обратной связи на стадии предварительного кондиционирования и после соединения, которые типичны для этой среды торфяника (рис. 3). Как в верхнем, так и в нижнем течении, усиленное проседание торфа создало больше условий для паводковых вод, что привело к более крупным трещинам и каналам ручьев. Это способствовало дополнительному переносу наносов и их отложению на торфяниках, вызывая дальнейшее проседание. Выше по течению эта петля положительной обратной связи была инициирована расслаиванием (загрузкой наносов), тогда как ниже по течению эта петля была инициирована проседанием торфяника (рис.3). В результате приливные каналы расширились на 15–25 км в торфяники, в результате чего точка, где река соединялась с уровнем морской базы, оказалась дальше вглубь суши (рис. 2C). Более того, за тысячелетие созревания с постоянно увеличивающимся отложением над берегом материнского канала вверх по течению его дамбы поднялись на ~ 1 м (рис. 2C). Это увеличило градиент водосбора с ∼4 см / км до ∼14 см / км, что дало небольшое преимущество по уклону наклона материнского канала (Старый Рейн) в 8–12 см / км вверх по течению от Утрехта (рис.2С). Это преимущество топографического градиента, улучшенное проникновение приливов к суше и уменьшение шероховатости растительности вместе вызвали значительное увеличение эффективного градиента энергии, особенно во время отлива в сочетании с высоким расходом реки и уровнем воды (рис. 2C). Как только соединение было установлено, каналы созрели и образовались естественные дамбы, устойчивые к боковой эрозии. Кроме того, масса дамбы вызвала дальнейшее сжатие торфа, тем самым еще больше затруднив развитие боковых каналов и приведя систему каналов к равновесному размеру (синие стрелки на рис.3). Положительной обратной связью приливно-речной связи была доставка большего количества наносов в эстуарий, что еще больше ускорило оседание торфа и проникновение ручьев, которые еще не были соединены с рекой (помечены звездочкой на рис. 3). Таким образом, один отрыв мог помочь развиться следующему отрыву — начиная до нашей эры (рис. 2A) вдоль южного края болота, затем следуют HIJ и, наконец, Lek.
Компартмент врожденных Vδ1 + γδ Т-клеток в груди человека связан с ремиссией при тройном отрицательном раке груди
γδ Т-клетки: охраняют и защищают ткани
Известно, что специфические подмножества γδ Т-клеток заселяют такие ткани, как кожа и кишка.Чтобы установить участие γδ Т-клеток в надзоре за раком груди, Wu et al. изолированных клеток из сеточных культур ткани груди человека. Они наблюдали цитолитические врожденные Т-клетки Vδ1 + в здоровой ткани груди человека, а также в ткани опухоли. Анализ небольшой группы женщин с трижды отрицательным раком молочной железы показал, что эти Vδ1 + Т-клетки были связаны с ремиссией и общей выживаемостью. Эти результаты демонстрируют, что резидентные в тканях человека γδ Т-клетки могут влиять на прогрессирование рака груди; эти клетки могут быть ключом к существующим или будущим иммунотерапевтическим вмешательствам.
Abstract
Врожденные тканевые резидентные компартменты γδ Т-клеток, способные защищать от канцерогенеза, хорошо известны у мышей. И наоборот, степень, в которой они существуют у людей, их потенциальные свойства и их вклад в пользу хозяина, по большей части не решены. Здесь мы демонстрируем, что здоровая человеческая грудь содержит отдельный компартмент Т-клеток γδ, в первую очередь экспрессирующий цепи Vδ1 рецептора Т-клеток (TCR), по сравнению с цепями Vδ2, которые преобладают в периферической крови.Резидентные в груди клетки Vδ1 + были функционально смещены в сторону цитолиза и продукции IFN-γ, но не IL-17, который был связан с воспалительными патологиями. Резидентные в груди клетки Vδ1 + могут быть активированы изначально через рецептор NKG2D, тогда как соседние CD8 + αβ Т-клетки нуждаются в передаче сигналов TCR. Сравнимая популяция клеток Vδ1 + была обнаружена в опухолях груди человека, и когда были проанализированы парные опухолевые и незлокачественные образцы от 11 пациентов с трижды отрицательным раком молочной железы, общая выживаемость без прогрессирования и общая выживаемость коррелировали с представлением клеток Vδ1 + , но не с общими γδ Т-клетками или Vδ2 + Т-клетками.Как и ожидалось, выживаемость без прогрессирования также коррелировала с αβ TCR. Однако, в то время как в большинстве случаев репертуар TCRαβ сфокусирован, что типично для антиген-специфических ответов, этого не наблюдалось для клеток Vδ1 + , что согласуется с их врожденной реактивностью. Таким образом, максимальная польза для пациента может быть получена за счет взаимодействия врожденных реакций, создаваемых резидентными тканями Vδ1 + компартментов, и адаптивных ответов, создаваемых αβ Т-клетками.
ВВЕДЕНИЕ
γδ Т-клетки представляют собой высококонсервативную третью линию лимфоцитов, которая использует соматическую перестройку гена для кодирования определяющего рецептора антигена ( 1 , 2 ).Хотя это является признаком адаптивного иммунитета, подмножества мышиных γδ Т-клеток также проявляют врожденную активность, проявляющуюся в быстрых ответах на самокодируемые «стрессовые антигены», такие как лиганды рецептора NKG2D ( 3 — 6 ) . Это известно как наблюдение за лимфоидным стрессом ( 7 ).
Учитывая, что лиганды NKG2D активируются сверхактивностью передачи сигналов рецептора эпидермального фактора роста (EGFR) и повреждением ДНК ( 8 , 9 ), естественно, что наблюдение за лимфоидным стрессом может способствовать иммунному надзору за раком ( 10 ).Мыши с дефицитом γδ Т-лимфоцитов демонстрируют значительно повышенную восприимчивость к раку в нескольких системах ( 4 , 11 — 13 ), и предпринимаются многочисленные попытки использовать их активность в клинических условиях ( 14 ). Такие подходы могут повысить эффективность существующих иммунотерапевтических методов, таких как блокада контрольных точек, и, в частности, Т-клетки химерного антигенного рецептора (CAR), которые показали ограниченный успех в лечении солидных опухолей. Более того, способность некоторых субпопуляций γδ Т-клеток секретировать хемокины и цитокины и / или представлять антиген убедительно свидетельствует об их способности способствовать терапевтическому потенциалу других типов клеток ( 12 , 15 — 17 ).
У мышей компартменты Т-клеток сигнатуры γδ связаны с отдельными тканями, такими как эпидермис, дерма, легкие, матка и эпителий кишечника ( 18 — 25 ), что, по-видимому, обеспечивает оптимальную способность обнаруживать злокачественную трансформацию и реагировать на нее. соседние клетки. Соответственно, мыши с дефицитом γδ Т-лимфоцитов обладают повышенной восприимчивостью к кожным канцерогенам из-за отсутствия дендритных эпидермальных γδ Т-клеток ( 5 ). Неизвестно, заселяются ли локальные компартменты Т-лимфоцитов все ткани.Тем не менее, перспектива ассоциированного с грудью компартмента мыши была подтверждена тем фактом, что представительство, хотя и непостоянное, γδ Т-клеток в лактирующих молочных железах было по крайней мере в четыре раза выше, чем в дренирующих лимфатических узлах. Более того, эти клетки использовали множество Т-клеточных рецепторов γδ (TCRγδs), что отличало их от резидентных γδ Т-клеточных компартментов кожи и кишечника ( 26 ).
Уже давно существует интерес к тому, в какой степени ассоциированные с тканью компартменты γδ Т-клеток могут сохраняться у людей и вносят ли они вклад в иммунный надзор за раком.С одной стороны, у людей нет очевидных аналогов дендритных эпидермальных γδ Т-клеток; с другой стороны, у бесчелюстных позвоночных есть резидентные в коже и кишечнике интраэпителиальные клетки со многими параллелями с γδ Т-клетками, что позволяет предположить, что такие компартменты сохранялись более полумиллиарда лет ( 27 ). Поэтому мы предположили, что неоптимальные методы обнаружения и / или извлечения Т-клеток из тканей человека могли затруднить попытки идентифицировать и охарактеризовать консервативные экстралимфоидные компартменты Т-клеток γδ.Эта гипотеза согласуется с неэффективностью и ошибками, описанными как для извлечения TCRαβ + резидентных в ткани клеток памяти T (T RM ) ( 28 ), так и для визуализации инфильтрирующих опухоль лимфоцитов (TIL) in situ ( 29 ). и подтверждается нашей недавней характеристикой большого интраэпителиального компартмента γδ Т-клеток в кишечнике человека ( 30 ).
В этом отношении уход за женщинами в рамках практики наблюдения и лечения большого риска рака молочной железы предоставил редкую возможность проанализировать статус γδ Т-клеток в здоровой ткани, полученной в результате: редукционной маммопластики или мастэктомии с уменьшенным риском; злокачественная ткань после широкой местной резекции; и парные злокачественные и незлокачественные ткани после терапевтических мастэктомий.Кроме того, важность исследования возможного существования местных γδ Т-клеток была подчеркнута свидетельствами того, что плотности TIL были положительными прогностическими индикаторами при некоторых типах рака груди ( 31 , 32 ). Несмотря на это, потенциал иммунотерапии при раке груди остается неясным из-за неутешительной реакции на современные иммунотерапевтические методы, такие как блокада контрольных точек ( 33 , 34 ).
РЕЗУЛЬТАТЫ
Т-клетки Vδ1 составляют основную подмножество резидентных γδ человеческой груди и смещены в сторону цитолиза.
Наша первоначальная цель состояла в том, чтобы оценить статус резидентных Т-клеток в здоровой груди человека.Однако получение и определение характеристик лимфоцитов из здоровых тканей человека обычно затрудняется низким и невоспроизводимым выходом и низкой жизнеспособностью клеток. Чтобы решить эту проблему, Кларк и его коллеги ( 35 — 38 ) разработали «сеточную» систему культивирования эксплантатов, которая позволила восстановить и охарактеризовать большое количество здоровых резидентных в коже и легких Т-клеток без существенного изменения их фенотипа. .
Поэтому мы применили сетки для дезагрегирования тканей груди 29 здоровых субъектов, перенесших редукционную маммопластику или мастэктомию со снижением риска.По сравнению с ограниченным и непостоянным восстановлением и низкой жизнеспособностью лимфоцитов, исследованных непосредственно ex vivo, сетки способствовали восстановлению и поддержанию CD3 — естественных киллеров (NK) / врожденных лимфоидных клеток (ILC), γδ Т-клеток и CD4 + и CD8 + αβ Т-клетки в каждом случае, хотя было некоторое обогащение NK / ILC и γδ Т-лимфоцитами (таблица S1 и фиг. 1A).
Рис. 1 Здоровая ткань груди человека содержит резидентные в ткани Vδ1 + , T c 1-скошенные γδ Т-клетки.( A ) Типичные графики проточной цитометрии, показывающие стратегию стробирования для идентификации лимфоцитов, включая субпопуляции γδ Т-клеток, выделенные из ткани груди человека и после культивирования в сетке. Лимфоциты были заблокированы по размеру и разбросу (1) с последующим исключением живых мертвецов (2), синглетным гейтом (3) и CD45 (4) перед подгруппой (от 5 до 8). ( B ) Сводные точечные диаграммы, показывающие клетки TCRγδ + , выделенные из здоровой ткани груди человека, выраженные как процент восстановленных клеток CD3 + ( n = 29) (указана медиана).В подмножестве из них использование цепи Vδ определяли количественно и выражали как процент пан-TCRγδ + ( n = 18) (указаны медианы). ( C ) Экспрессия маркеров клеточной поверхности NKG2D, CD28, PD-1, CD103 и CD69 на Т-клетках Vδ1 + ( n = от 9 до 11) (указаны медианы). ( D ) Функциональный фенотип резидентных в ткани Vδ1 + Т-клеток. Точечные диаграммы, показывающие окрашивание внутриклеточных цитокинов на IFN-γ ( n = 12), IL-13 ( n = 8), IL-17A ( n = 9), TNF ( n = 10) и экспрессия CD107a на клеточной поверхности ( n = 5) после стимуляции in vitro массовых культур CD3 + PMA и иономицином (4 часа) (указаны медианы).( E ) Сводные данные, показывающие процент резидентных в груди Vδ1 + или CD4 αβ Т-клеток, окрашенных внутриклеточно на IL-17A. Клетки выделяли путем культивирования эксплантатов, а затем выращивали в двух формах среды, исключающей IL-17, с последующей активацией in vitro ФМА и иономицином (4 часа) ( n = 3, за исключением клеток Vδ1 + , выращенных в TGF -β – содержащая среда, где n = 2) (среднее значение с указанием SEM).
Как это обычно бывает для γδ Т-клеток периферической крови (PB) ( 39 ), представление γδ Т-клеток в груди показало значительные индивидуальные различия (рис.1Б). Однако клетки, экстрагируемые молочной железой, четко отличались от Т-клеток PB γδ по использованию цепи TCRVδ. В то время как большинство PB-клеток экспрессируют Vδ2 в паре с Vγ9, большинство γδ Т-клеток донорской груди были преимущественно Vδ1 + (медиана, 60,5% γδ Т-клеток), как и большинство резидентных в коже и кишечнике γδ Т-клеток человека (рис. 1B) ( 40 ). Во всех случаях было некоторое представление Vδ2 + Т-клеток (медиана, 13,6% от γδ Т-клеток) и клеток, не экспрессирующих ни Vδ1, ни Vδ2 (медиана, 20.45% γδ Т-клеток), которые в некоторых случаях были почти исключительно Vδ3 + (рис. S1A).
Фенотипическая последовательность γδ Т-клеток в сеточных культурах и аналогичных клетках, исследованных непосредственно ex vivo, была очевидна из паттернов экспрессии нескольких биологически важных поверхностных маркеров, хотя CD69 экспрессировался большим количеством клеток в сеточной культуре (рис. S1, B и C). , два верхних ряда). Путем многопараметрического анализа подмножества доноров мы могли вывести консенсусный фенотип Т-клеток Vδ1 + , который напоминал фенотип экстралимфоидных γδ Т-клеток в других тканях мышей и людей ( 4 , 41 — 44 ) , а именно, однородная положительность для активирующего рецептора NK-клеток NKG2D и для CD69, и в значительной степени отсутствие костимулятора лимфоидных Т-клеток, CD28 (рис.1С и рис. S1C, нижний ряд). В среднем ~ 20% Т-клеток груди Vδ1 + экспрессировали PD-1, тогда как несколько больше экспрессировали интегрин эпителиального взаимодействия, CD103 (α E β7), хотя и с высокой индивидуальной вариабельностью (рис. 1C и рис. S1C). , Нижний ряд).
Для оценки функционального потенциала клеток их инкубировали с форбол 12-миристат 13-ацетатом (PMA) и иономицином, которые совместно имитируют передачу сигналов TCR, и анализировали на продукцию внутриклеточных цитокинов и на поверхностную экспрессию CD107a, маркера дегрануляции. и экзоцитоз цитолитических медиаторов, таких как гранзимы и перфорин.Связанные с грудью Т-клетки Vδ1 + сочетают экспрессию CD107a с продукцией фактора некроза опухоли (TNF) и интерферона-γ (IFN-γ) (рис. 1D), фенотипа Т-цитолитического типа 1 (T c 1), который среди CD8 + TCRαβ + TIL считается очень полезным для пациентов ( 45 — 48 ). Клетки некоторых доноров экспрессировали интерлейкин-13 (IL-13) (в среднем 5,2% γδ Т-клеток), что недавно было связано с наблюдением за опухолями Т-клеток ( 49 ), но не было продукции IL-17A, т.е. эффекторный цитокин, обычно продуцируемый γδ Т-клетками мыши, у которых он был связан с промотированием опухоли ( 50 — 52 ).
Поскольку производство IL-17 человеческими γδ Т-клетками, как сообщается, трудно наблюдать ( 53 ), мы проверили, будут ли резидентные γδ Т-клетки в культуре реагировать в культуре на условия перекоса IL-17, а именно, IL-1β, ИЛ-6, ИЛ-23, ИЛ-2 ± трансформирующий фактор роста – β (TGF-β) ( 54 ). Однако, в то время как резидентные CD4 + αβ Т-клетки, экстрагированные и поддерживаемые вместе с Vδ1 + Т-клетками в идентичных культурах эксплантатов молочной железы (таблица S1), продуцировали IL-17A [медиана, 16.5% (с использованием ИЛ-2) и 12,6% (с использованием ИЛ-2 + ИЛ-15) CD4 + Т-клеток] и заметно увеличенное производство в условиях перекоса ИЛ-17 (медианы 41,5 и 35,5%, соответственно) (Рис. 1E), груди Vδ1 + Т-клетки продуцировали незначительное количество IL-17A при всех обстоятельствах (Рис. 1E).
Чтобы поместить фенотипы Т-клеток груди Vδ1 + в контекст, аналогичный анализ был проведен на совместно экстрагированных Т-клетках Vδ2 + и Vδ1 — Vδ2 — γδ, на CD4 + и CD8 + αβ Т-клетки и на CD3 — лимфоцитах, которые будут включать NK и ILC.Наибольшее сходство с Т-клетками Vδ1 + продемонстрировали клетки Vδ2 + и Vδ1 — , Vδ2 — γδ и CD8 + αβ Т-клетки, хотя некоторые такие клетки экспрессировали CD28 и CD8 + . αβ Т-клетки были более равномерно CD103 + (фиг. S1D). NK и ILC также были подобны Т-клеткам Vδ1 + , за исключением того, что в них всегда отсутствовал PD-1. Наконец, как и ожидалось, CD4 + αβ Т-клетки лишены экспрессии NKG2D и в основном представлены CD28 + и CD103 — (рис.S1D), что согласуется с недавно описанным тканевым резидентным фенотипом CD4 + ( 55 ).
После активации ассоциированные с грудью CD8 + αβ Т-клетки были функционально наиболее похожи на Т-клетки Vδ1 + по их фенотипу T c 1, показывая еще большую частоту клеток, продуцирующих IFN-γ (рис. S1E). Аналогично, эксплантированные в грудь CD4 + αβ Т-клетки включали больше продуцентов IL-13, чем Vδ1 + Т-клетки. В общем, здоровая грудь содержит сложную лимфоидную экосистему из нескольких типов клеток со связанными, но разными фенотипами.
Человеческая грудь Vδ1
+ Т-клетки подобны врожденнымРезидентные в коже мыши γδ Т-клетки могут отвечать in vivo на активацию лиганда NKG2D без явной стимуляции TCR ( 5 ). В отношении рака это потенциально важно, потому что лиганды NKG2D активируются повреждением ДНК ( 8 ) и повышенной активностью EGFR ( 9 ). Поэтому мы исследовали, будут ли эксплантированные груди Vδ1 + Т-клетки отвечать на связанный с планшетом рекомбинантный белок MICA, лиганд NKG2D, обычно экспрессируемый опухолями.MICA спровоцировал подмножество Т-клеток Vδ1 + , чтобы продуцировать TNF и IFN-γ и активировать CD107a в ответ, ингибируемый анти-NKG2D (фиг. 2A). Напротив, CD8 + αβ Т-клетки в идентичных клеточных культурах не давали значительных ответов на MICA по сравнению с контролями (фиг. 2A и фиг. S2A), тогда как оба типа клеток демонстрировали повышенные ответы, когда MICA применялся в качестве костимулятора против антибиотиков. -CD3 (рис. 2B) ( 56 , 57 ). Врожденную чувствительность других γδ Т-клеток груди было сложно исследовать, потому что даже в тех случаях, когда они составляли большую долю тканевых γδ Т-клеток, не-Vδ1 + клеток обычно было слишком мало для надежного анализа (рис.S2B). Несмотря на это, мы наблюдали некоторые NKG2D-зависимые, врожденные ответы среди Vδ1 — Vδ2 — T-клеток, но не среди Vδ2 + T-клеток груди (рис. S2B).
Рис. 2. Резидентные Т-клетки Vδ1 + в ткани груди подобны врожденным.( A ) Сводные данные, показывающие внутриклеточное окрашивание для IFN-γ ( n = 19 для Vδ1 + и n = 15 для CD8 + ), TNF ( n = 17 для Vδ1 + и n = 13 для CD8 + ) и экспрессия CD107a на клеточной поверхности ( n = 10 для Vδ1 + и n = 5 для CD8 + ) после активации молочной железы in vitro. резидентные клетки Vδ1 + или Vδ1 + и CD8 + αβ Т-клеток из одних и тех же культур, подвергнутые воздействию рекомбинантного MICA (10 мкг / мл) на планшете в присутствии брефельдина A (BFA).Отображается в процентах от родительского Vδ1 + или CD8 + гейта. ( B ) Сводные данные, показывающие внутриклеточное окрашивание для IFN-γ ( n = 6 для Vδ1 + и n = 6 для CD8 + ) и TNF ( n = 5 для Vδ1 + и n = 5 для CD8 + ) после активации in vitro резидентных в ткани груди Vδ1 + и CD8 + αβ Т-клеток низкими дозами связанных с планшетом анти-CD3-антител (50 нг / мл) с рекомбинантным MICA (10 мкг / мл) в присутствии BFA или без него.Там, где указано, стимулированные MICA клетки предварительно обрабатывали антителом против человеческого NKG2D (нанесено на график как процент от родительского Vδ1 + или CD8 + gate). ( C ) Сводные данные для резидентных в груди Vδ1 + Т-клеток, демонстрирующие внутриклеточную продукцию IFN-γ после активации in vitro с помощью IL-12 ( n = 3) или IL-18 ( n = 3) или IL-12 + IL-18 ( n = 9) и только со средой ( n = 9). Для всех панелей указано среднее значение с SEM.** P ≤ 0,01, *** P ≤ 0,001 и **** P ≤ 0,0001, Kruskal-Wallis с апостериорным тестом Данна, скорректированным для множественных испытаний.
Также сообщалось, что врожденные γδ Т-клетки дают сильные, независимые от TCR ответы на комбинации STAT (преобразователь сигнала и активатор транскрипции) — сигнальный цитокин и член семейства IL-1 ( 58 ). Эксплантированные грудью Т-клетки Vδ1 + продуцировали IFN-γ в ответ на IL-12 + IL-18, но не на любой из них по отдельности, тогда как CD4 + и CD8 + αβ Т-клетки значительно хуже реагировали на IL-12. + Ил-18 (рис.S2C). В общем, здоровая грудь содержала смесь врожденных Vδ1 + Т-клеток и в первую очередь адаптивных αβ Т-клеток.
Врожденные γδ Т-клетки при раке молочной железы человека
Идентификация врожденных γδ Т-клеток в здоровой человеческой груди послужила предпосылкой для исследования ассоциированного с тканями лимфоидного компартмента у субъектов с раком груди. Хотя рак груди различается по лимфоидным инфильтратам ( 32 , 59 ), γδ Т-клетки неизменно восстанавливались и были в значительной степени сопоставимы с таковыми из здоровых тканей с точки зрения использования TCR: Vδ1 был преобладающим, хотя образцы опухолей включали некоторые примеры, где преобладали либо Vδ2 + клетки, либо Vδ1 — Vδ2 — клетки (рис.S3A). Как и γδ Т-клетки из здоровой груди, клетки, выделенные из опухолей с использованием сеток, были фенотипически сопоставимы с теми, которые исследовались непосредственно ex vivo, хотя клетки сетки снова показали более высокую экспрессию CD69 и, в некоторой степени, NKG2D (рис. S3B). Сходство использования TCR и экспрессии поверхностных маркеров для γδ Т-клеток из опухолевой и незлокачественной ткани было особенно очевидно в парных образцах от 26 субъектов (фиг. 3A и фиг. S3C). Более того, эта сопоставимость распространялась на другие субпопуляции лимфоцитов, одновременно взятые из парных образцов (рис.S3D).
Рис. 3 Врожденно-подобные клетки Vδ1 + T c 1 являются основным подмножеством TIL молочной железы.( A ) γδ Т-клеток последовательно выделяли из образцов опухоли груди человека, полученных после мастэктомий после культивирования в сетке. Точечные графики, показывающие количество клеток TCRγδ + , выделенных из парной доброкачественной ткани молочной железы (•) или опухоли (ο), выраженное в процентах от CD3 + Т-клеток. Т-клетки γδ были дополнительно фенотипированы для Vδ1 + , Vδ2 + и Vδ1 —, Vδ2 — использования TCR, выраженного в процентах от TCRγδ + Т-клеток ( n = 25) (указаны медианы) .( B ) Vδ1 + Т-клетки, инфильтрирующие опухоль груди, функционально искажены. Точечные диаграммы, показывающие окрашивание внутриклеточных цитокинов на IFN-γ ( n = 12), IL-13 ( n = 11), IL-17A ( n = 4), TNF ( n = 9) и экспрессия CD107a на клеточной поверхности ( n = 4) после активации in vitro ФМА (10 нг / мл) и иономицином (1 мкг / мл) в присутствии BFA (указаны медианы). ( C ) Сводные данные, показывающие внутриклеточное окрашивание на IFN-γ ( n = от 9 до 12, в зависимости от условий активации), TNF ( n = 7) и CD107a на поверхности клетки ( n = от 4 до 7 в зависимости от условий активации) экспрессии после активации in vitro Vδ1 + и CD8 + αβ Т-клеток с помощью рекомбинантной MICA, связанной с планшетом (10 мкг / мл) в присутствии BFA или активации in vitro с помощью IL-12 ( 100 нг / мл) и IL-18 (100 нг / мл).Там, где указано, стимулированные MICA клетки предварительно обрабатывали блокирующим антителом против человеческого NKG2D (среднее с указанным SEM). ( D ) Линии опухолевых клеток, MCF7 и HCC1954, инкубировали с отрицательно отсортированными γδ Т-клетками (γδ), полученными из незлокачественной ткани молочной железы или опухоли груди при E: T, равном 5: 1, в присутствии (γδ + αNKG2D) или отсутствие (γδ) блокирующего антитела против NKG2D в течение 48 часов. Линии клеток без эффекторных γδ Т-клеток использовали в качестве отрицательного контроля (контроль). На точечных графиках показаны концентрации расщепленного каспазой цитокератина 18 (cCK18).Каждая точка данных представляет собой среднее значение двух технических повторов; средние значения для этих точек данных показаны горизонтальной линией (обратите внимание, что было только три донора для анализа уничтожения клеток HCC1954 опухолевыми γδ Т-клетками в присутствии анти-NKG2D). △ и ○ — два донора, у которых была пара доброкачественной ткани молочной железы и опухоль молочной железы. * P <0,05, ** P ≤ 0,01, *** P ≤ 0,001 и **** P ≤ 0,0001, Краскела-Уоллиса с апостериорным тестом Данна, скорректированным для множественных испытаний.
Аналогичным образом, функциональный потенциал TILs Vδ1 + был сопоставим с функциональным потенциалом Т-лимфоцитов Vδ1 + незлокачественной молочной железы, поскольку они были искажены по T c -1 и дефицитным по IL-17 (рис. 3B). TIL Vδ1 + снова реагировали на лиганды NKG2D и IL-12 + IL-18 в отсутствие явной передачи сигналов γδ TCR, тогда как совместно изолированные TIL CD8 + αβ не проявляли значимых ответов на MICA по сравнению с контролями, хотя они действительно лучше реагировали на IL-12 + IL-18, чем их аналоги из здоровой груди (рис.3С).
Учитывая их сильную цитолитическую чувствительность, мы проверили способность полученных из груди γδ Т-клеток убивать две линии опухолевых клеток груди, MCF7 и HCC1954, для которых лизис опухолевых клеток отличался от гибели лимфоцитов путем количественной оценки высвобождения цитокератина 18 ( 60 ). Обратите внимание, что, поскольку γδ Т-клетки не ограничены MHC (основным комплексом гистосовместимости), было возможно оценить их функциональные ответы на неавтологичные опухолевые линии. γδ Т-клетки от здоровых доноров груди ( n = 4) воспроизводимо убивали клетки MCF7 при соотношении эффектор: мишень (E: T) 5: 1 (рис.S3E), и, используя это соотношение, мы обнаружили, что клетки из здоровой груди и из образцов опухоли ( n = 4) показали сопоставимую способность убивать MCF7 и HCC1954 (рис. 3D). Однако, в то время как блокада рецептора NKG2D уменьшала гибель γδ Т-клетками из здоровой груди, гибель TCRγδ + TILs подвергалась меньшему влиянию (рис. 3D). Мы также наблюдали уничтожение первичных опухолевых клеток аутологичными TCRγδ + TIL у одного пациента, у которого клетки первичной опухоли могли расти и стабильно сохраняться (рис.S3F). В целом, первичные γδ Т-клетки, полученные от рака груди, были функционально компетентными, могли отвечать врожденным путем посредством вовлечения NKG2D и могли лизировать клетки опухоли груди, хотя это не было явно зависимым от NKG2D.
Vδ1
+ TIL и стойкая ремиссияУчитывая функциональный сдвиг TIL c 1 γδ TIL, мы хотели изучить их статус по отношению к клиническому исходу в агрессивной подгруппе рака молочной железы, когда время до событий относительно короткое. . С этой целью мы искали пациентов с тройным отрицательным раком молочной железы (TNBC), у которых мы могли получить доступ к достаточному количеству парных опухолевых и доброкачественных тканей и для которых были доступны точные данные клинического наблюдения.Этим критериям соответствовали 11 пациентов, пролеченных в больницах Гая и Сент-Томаса в Лондоне (исследование BTBC, номер REC: 13 / LO / 1248), для которых были взяты биобанки, фиксированные формалином и залитые парафином (FFPE) образцы. В остальном выбор пациентов не производился. У всех пациентов был локализованный или локорегиональный TNBC [Американский объединенный комитет по раку (AJCC) стадии с I по III], и им была проведена операция с лечебной целью. Пять из 11 остались в полной ремиссии при последнем наблюдении (диапазон от 48 до 63 месяцев; таблица S2; выделены синим цветом на следующих рисунках), тогда как у шести произошел рецидив отдаленного метастатического заболевания в течение 18 месяцев (диапазон от 6 до 18 месяцев; таблица S2; выделены красным на следующих рисунках).
блоков FFPE препарировали иглой, чтобы очертить опухоль и незлокачественные ткани для выделения геномной ДНК. Учитывая сложность иммуногистохимических подходов к обнаружению γδ Т-клеток в FFPE ( 61 ), мы использовали количественное секвенирование геномной ДНК реаранжированных генов цепей TCRα и TCRδ для определения абсолютного количества αβ и γδ Т-клеток. Было показано, что этот подход более чувствителен, чем иммуногистохимия, для обнаружения TIL ( 29 ) и используется в клинической практике, например, для оценки минимальной остаточной болезни.
Мы обнаружили, что как αβ, так и γδ Т-клетки были значительно более многочисленными на микрограмм ДНК, экстрагированной из опухолевой ткани, по сравнению с парными доброкачественными тканями (рис. 4A и рис. S4A). Однако примечательно, что в случаях ремиссии количество ДНК TCRα, TCRγδ, TCRVδ1 и TCRVδ2 неизменно было обогащено парными злокачественными тканями по сравнению с доброкачественными, тогда как картина в рецидивирующих случаях была по существу случайной (рис. S4A). Более того, помимо обогащения по сравнению со здоровой тканью, было заметно больше TCRα + и Vδ1 + TCR на микрограмм общей ДНК опухоли в случаях ремиссии по сравнению с рецидивом (рис.4Б). Таким образом, указывая на положительный клинический результат, динамика небольшого числа Т-клеток Vδ1 + была столь же сильной, как и гораздо более крупномасштабная динамика αβ Т-клеток, определенные подмножества которых, как было показано, предсказывают выживаемость в TNBC ( 62 ). Напротив, это не было так ни для полных TCRγδ, ни для Vδ2 TCRs (Fig. 4B), последние, возможно, отражают инфильтрацию клеток из PB.
Рис. 4 Vδ1 + Т-клетки в TNBC являются прогностическими факторами выживаемости без болезней и общей выживаемости.( A ) Общий ландшафт субпопуляций Т-клеток в незлокачественной ткани молочной железы («Ткань») и подобранной опухолевой ткани («Опухоль»), определенный путем количественного секвенирования реаранжированных генов TCR от пациентов, перенесших мастэктомию по поводу TNBC. Абсолютные копии TCR (заменитель количества Т-клеток) нанесены на микрограмм входной ДНК. График отдельных пациентов: красный — пациенты с рецидивом заболевания; синий — пациенты в стадии ремиссии (средняя полоса нанесена на график). * P <0,05 и ** P ≤ 0.01, знаковый ранговый критерий Уилкоксона для пар соответствий. ( B ) Внутриопухолевое присутствие αβ Т-клеток и γδ Т-клеток у пациентов с последующим рецидивом заболевания по сравнению с пациентами, оставшимися в ремиссии. * P <0,05, критерий Колмогорова-Смирнова. ( C ) ВБП (месяцы после операции) разделены на медианные субпопуляции Т-клеток, обнаруженные в 11 опухолях TNBC. * P <0,05 и ** P ≤ 0,01, тест Гехана-Бреслоу-Уилкоксона. ( D ) ОС (месяцы после операции) разделены на медианные субпопуляции Т-клеток, обнаруженные в 11 опухолях TNBC.* P <0,05 и ** P ≤ 0,01, тест Гехана-Бреслоу-Уилкоксона.
Заметные проявления корреляций были очевидны на графиках Каплана-Мейера выживаемости без прогрессирования (PFS), где ограниченное представление (ниже медианы значений) внутриопухолевых TCRα и Vδ1 TCRs было предиктором плохой PFS, тогда как ни общий TCRγδ, ни Vδ2 + TCRs предсказали исход (рис. 4C). Кроме того, наблюдалась положительная и значимая корреляция внутриопухолевого TCRα с TCR Vδ1 (Spearman r = 0.75, P ≤ 0,01) (рис. S4B), тогда как ни общие TCRγδ, ни Vδ2 TCR не коррелировали с TCRα: Более того, корреляция TCRα-TCRδ не существовала в доброкачественной ткани (рис. S4B). Кроме того, TCR Vδ1 предсказывали общую выживаемость (OS), хотя TCRα — нет (рис. 4D).
In situ доказательства наличия врожденных Vδ1
+ TILУстановив значимые положительные корреляты TCRαβ + T-клеток с PFS и Vδ1 + T-клеток с PFS и OS, мы оценили их репертуары TCR.При представлении в виде круговых древовидных графиков репертуары Vδ1 + в опухолях по сравнению с парными доброкачественными тканями не показали четкой общей фокусировки (примеры показаны на фиг. 5A), что определяется количественно D50 (наименьшее количество клонов, составляющих 50% от общего числа клонов). общее количество наблюдаемых последовательностей) из парных доброкачественных и опухолевых тканей (таблица S3). Фокусировка могла бы предложить адаптивный ответ, управляемый TCR, как мы наблюдали для репертуаров TCRα большинства опухолей по сравнению с парными нормальными тканями (примеры, показанные на рис.5B), что соответствует предыдущим отчетам ( 62 , 63 ).
Рис. 5 Отсутствие фокусировки опухоли Vδ1 + TCR относительно фокусировки последовательностей TCRα.( A ) Примеры круговых древовидных графиков репертуара Vδ1 + в парной доброкачественной ткани («ткань») и опухолевой ткани («опухоль»), где каждый круг представляет собой уникальный клонотип TCR, а размер круга пропорционально представлению указанного клона.Графики были созданы из общего количества Vδ1 + TCR. ( B ) Примеры круговых древовидных графиков репертуара TCRα в парной доброкачественной ткани («ткань») и опухолевой ткани («опухоль»). Графики были созданы для общего TCRα + TCR. ( C ) Последовательности Vδ1 + и TCRα + в доброкачественной и опухолевой ткани были подвергнуты пониженной выборке (внутри каждого пациента) до эквивалентных чисел для расчета показателей разнообразия. Степень фокусировки репертуара оценивалась по дельте коэффициента Джини и дельте D50 последовательностей цепей Vδ1 и верхних 10% по изобилию последовательностей TCRα в опухоли по сравнению с парными доброкачественными репертуарами.Чтобы проверить, различалась ли степень фокусировки репертуара между компартментами Vδ1 + и TCRα + у отдельных пациентов, для сравнения ΔGini и ΔD50 использовался критерий знакового ранга согласованных пар Уилкоксона. Все последовательности были проанализированы на основе аминокислотной последовательности. n = 11.
Для дальнейшего анализа данных мы также рассчитали коэффициент Джини (статистическая мера распределения, где 0 является полностью поликлональным, а 1 — моноклональным) для TCR Vδ1 из парных доброкачественных и опухолевых тканей (таблица S3).Обратите внимание, что TCR опухоли были подвергнуты пониженной выборке, так что эквивалентное количество TCR сравнивалось у каждого пациента. Затем такое же лечение применяли к TCRα, хотя мы рассматривали только 10% наиболее распространенных TCR TCRα, учитывая недавние данные о том, что наиболее релевантные антиген-реактивные αβ Т-клетки обычно находятся в этой фракции ( 64 ). Затем для каждого пациента мы рассчитали дельту коэффициента Джини парных опухолевых тканей и доброкачественных тканей как для Vδ1, так и для TCRα: обратите внимание, что фокусировка опухоли будет отражаться положительным коэффициентом ΔGini (рис.5С). Затем мы аналогичным образом рассчитали дельту значений D50 для парных опухолевых и незлокачественных тканей как для Vδ1, так и для TCRαβ: обратите внимание, что фокусировка опухоли будет отражаться отрицательным ΔD50 (рис. 5C). Эти анализы количественно подтвердили, что фокусировка репертуара TCRα происходила во всех образцах опухолей, кроме двух (KCL-059 и KCL-202), тогда как репертуар Vδ1 не показал смещения ни в сторону фокусировки, ни в сторону диверсификации (рис. 5C).
В качестве дополнительного подхода мы также применили показатели репертуара к показаниям TCR без понижающей дискретизации (необработанным), используя нормализованные меры клональности (нормализованная энтропия Шеннона, коэффициент Джини и D50), как ранее использовалось другими ( 29 , 63 ).Эти методы также предполагали фокусировку репертуара опухолей для TCRα (значимость была достигнута для D50), тогда как для Vδ1 таких результатов не было (рис. S5 и таблица S4). В совокупности эти данные убедительно свидетельствуют о том, что клеточные ответы Vδ1 + не управляются клонотипическими антигенами.
Учитывая, что среди реаранжировок генов TCR, TCRδ обладает наибольшим потенциалом для разнообразия соединений ( 65 ), неудивительно, что не наблюдались общедоступные последовательности Vδ1 в опухолях разных доноров (рис.6А). Однако было некоторое перекрытие последовательностей Vδ1 между опухолями и тканями от одного и того же донора (рис. S6). Хотя размер выборки был небольшим, отсутствие общедоступных последовательностей также согласуется с TCR-агностической, врожденной регуляцией резидентных в ткани Т-клеток Vδ1 + . Напротив, оценка образца сопоставимого размера показала, что некоторые Vδ2 TCRs были общими для нескольких доноров (рис. 6B). Большинство общих последовательностей отражают TCR, реактивные к фосфо-антигенам, которые могут быть активированы в опухолях ( 66 ), с консервативным гидрофобным остатком в положении 97 (таблица S5), специфически связанным с фосфо-антиген-опосредованной селекцией Vδ2 + Репертуар ( 67 , 68 ).
Рис. 6. Отсутствуют обнаруживаемые общедоступные внутриопухолевые клонотипы Vδ1 + .( A ) Пересечения клонотипов Vδ1 + между 11 образцами опухолей пациентов. Вертикальные столбцы представляют количество уникальных TCR, а точечная матрица представляет совместное использование TCR пациентами. Общий или общедоступный клонотип будет представлен как минимум двумя красными точками (разделяемыми двумя пациентами), соединенными вертикальной красной линией. Частные последовательности представлены несвязанной единственной красной точкой.( B ) Пересечения клонотипов Vδ2 + между 11 образцами опухолей пациентов. Все последовательности были проанализированы на основе аминокислотной последовательности.
ОБСУЖДЕНИЕ
За последнее десятилетие в иммунологии рака произошли огромные изменения, когда стало ясно, что опухоли часто являются антигенными и что опухолевые Т-клетки могут обеспечивать полезные для пациента реакции, особенно если подавление подавления блокадой контрольных точек ( 69 , 70 ). Следовательно, существует значительный интерес к иммунной экологии опухолей.За тот же период стало ясно, что несколько экстралимфоидных тканей, в которых обычно образуются опухоли, обычно содержат большие миелоидные и лимфоцитарные компартменты, включая γδ Т-клетки, которые становятся резидентными в тканях во время развития клеток, и системные αβ Т-клетки, которые становятся клетками T RM . после прайминга во вторичных лимфоидных органах ( 71 , 72 ).
Хотя положительные клинические исходы TNBC были связаны с клетками CD8 + TCRαβ + T RM ( 62 ), мало исследовалось, существует ли компартмент Т-клеток γδ, резидентных в груди человека, который мог бы влияют на исходы рака груди ( 73 ).Это исследование обращается к этому моменту, сначала устанавливая резидентный в ткани компартмент γδ Т-клеток в здоровой ткани молочной железы человека. Это может быть эволюционно консервативным, поскольку лимфоциты TCRγδ + были обнаружены в эпителии альвеолярной молочной железы лактирующих коров ( 74 ) и были связаны с лактирующими молочными железами у мышей ( 26 ). Чтобы охарактеризовать резидентные в груди γδ Т-клетки человека в достаточном количестве, мы использовали сеточные культуры, ранее использованные для выяснения ключевых характеристик Т-клеток кожи и легких человека ( 35 — 38 ).Хотя это ограничение, наблюдалась сильная фенотипическая согласованность с γδ Т-клетками груди, исследованными непосредственно ex vivo. Это позволило нам описать компартмент как в основном Vδ1 + , NKG2D + , CD69 + , частично CD103 + и с фенотипом T c 1 без продукции IL-17. Это очень отличается от Т-клеток γδ PB, но имеет общие черты с γδ Т-клетками кишечного эпителия человека ( 40 ).
Кроме того, γδ Т-клетки молочной железы человека по своей природе были чувствительны к активаторам NKG2D, тогда как совместно размещенные αβ Т-клетки требовали одновременной стимуляции TCR.Таким образом, здоровые клетки груди человека Vδ1 + обладают внутренним потенциалом для обнаружения и быстрого реагирования на локальные клетки на пути к злокачественному новообразованию. Некоторые из этих сигнатурных свойств могут быть общими с локальными Vδ1 — δ2 — Т-клетками, которые часто являются Vδ3 + , хотя эти клетки чаще всего присутствовали в очень небольшом количестве.
В опухолях молочной железы Vδ1 + Т-лимфоцитов и αβ Т-клеток часто было больше, чем в парных здоровых тканях, особенно у пациентов в стадии ремиссии.Это, вероятно, отражает точку перегиба, в которой произошел активирующий иммунный ответ на опухоли. Кроме того, при извлечении из опухолей груди сравнительно увеличенные популяции Vδ1 + и αβ Т-клеток были функционально компетентными, сохраняя врожденную чувствительность и потенциал T c 1 клеток из здоровой груди. Эти наблюдения выявили доказательства того, что TIL рака молочной железы PD-1 + функционально отвечали на рестимуляцию ( 75 ) и что их присутствие могло быть связано с благоприятным исходом ( 62 ).Хотя только несколько пациентов были доступны для углубленного анализа в этом исследовании, их было достаточно, чтобы показать значительную положительную корреляцию опухолевых Vδ1 + и αβ Т-клеток с клиническим исходом, при этом TCR Vδ1 коррелировали как с PFS, так и с OS. Поэтому привлекательно выдвинуть гипотезу о том, что максимальная польза для пациента достигается за счет взаимодействия врожденной отзывчивости локальных клеток Vδ1 + с антиген-специфическим образом действия αβ Т-клеток, в частности CD8 + TCRαβ + T RM ячеек ( 62 ).
Можно представить по крайней мере два благоприятных для пациента аспекта сотрудничества между Т-клетками Vδ1 + и TCRαβ + . Во-первых, благодаря распознаванию опухолей с помощью врожденных стимулов, ответы Т-клеток Vδ1 + не могут быть ограничены ни количеством соматических мутаций, генерирующих неоантиген, ни иммунным подавлением презентации пептидного антигена ( 47 , 76 — 79 ). Врожденные стимулы могут включать лиганды для нескольких рецепторов NK ( 43 ), включая, но не ограничиваясь этим, NKG2D.
Во-вторых, цитолитический фенотип T c 1 клеток Vδ1 + может быть усилен способностью клеток способствовать тканевой иммуногенности посредством секреции хемокинов и, возможно, посредством прямой презентации антигена ( 16 , 17 , 80 ). Решающая роль резидентных в ткани клеток Vδ1 + может заключаться в организации многокомпонентных местных иммунных ответов на определенные проблемы, оставаясь при этом толерантными к другим. В этом контексте иммуногенность ткани / опухоли может быть эффективно повышена в клинике с помощью агентов, стимулирующих активность локальных Т-клеток Vδ1 + , или путем адоптивного переноса клеток в сочетании с активацией / дерепрессией адаптивных Т-клеток.Поскольку γδ Т-клетки не ограничены MHC, они могут быть адоптивно перенесены от гетерологичных доноров, а поскольку они естественным образом резидентны в тканях, локальные клетки Vδ1 + могут справляться с гипоксической средой, которая оказывается враждебной системным лимфоцитам ( 81 , 82 ).
Такие соображения могут относиться к другим тканям человека, несущим локальные компартменты γδ Т-клеток, таким как кишечник, и могут поддерживать, как сообщается, сильную корреляцию γδ Т-клеток с благоприятными клиническими исходами в широком спектре опухолей человека ( 83 ).Тем не менее, в этом исследовании ни общие γδ TCR, ни Vδ2 + TCR не коррелировали с клиническим исходом. Это подчеркивает тот факт, что γδ Т-клетки включают биологически различные подмножества, как в случае αβ Т-клеток или ILC. Даже среди внутриопухолевых Т-клеток CD8 + , которые традиционно ассоциируются с пользой для пациента, наибольшая польза в TNBC была приписана дискретному подмножеству местных клеток CD8 + T RM ( 62 ). Сообщалось, что у мышей функционально разные субпопуляции γδ Т-клеток либо опосредуют, либо подавляют иммунный надзор за опухолью ( 84 , 85 ).Чаще всего IL-17 участвует в промотировании опухоли γδ Т-клетками ( 50 — 52 ), тогда как клетки, продуцирующие IL-17, по-видимому, редко встречаются у людей, где преобладающим фенотипом является цитолитический и TNF / IFN- γ продуцирует ( 86 ), как описано здесь.
У мышей врожденная реактивность γδ Т-клеток и их подавление продукции IL-17 были индуцированы в процессе развития субпопуляциями, тканеспецифичными элементами отбора из семейства бутирофилин-подобных (Btnl / BTNL), члены которого также могут регулируют γδ Т-клетки человека ( 30 , 40 , 58 , 87 ).Следовательно, возможно, что такие элементы действуют локально, чтобы выбрать и регулировать резидентные в груди человека Vδ1 + Т-клетки, в этом отношении молочная железа является одной из немногих тканей, экспрессирующих BTNL9 ( 88 ).
Наше исследование не фокусировалось на взаимодействии Vδ1 + Т-клеток груди с другими резидентными иммунными клетками груди, включая В-клетки ( 32 ). Точно так же не определялись пространственные отношения между γδ Т-клетками груди и ассоциированными с опухолью третичными лимфоидными структурами ( 31 , 32 ).На практике клинические исследования показали, что рак груди человека, включая TNBC, может быть уязвим для иммунной атаки ( 33 , 34 ), однако эффективность иммунотерапии при этом показании была относительно низкой. Мы твердо верим, что это можно исправить, сместив методы лечения с их уникальной ориентации на обычные, адаптивные Т-клеточные ответы и изучив естественную экологию локального компартмента Т-клеток груди. В частности, мы считаем, что это может способствовать иммуногенности опухолевых тканей, которая стимулирует и поддерживает полезные для пациента адаптивные ответы.
МАТЕРИАЛЫ И МЕТОДЫ
Дизайн исследования
Целью этого исследования было выяснить, может ли человеческая грудь содержать резидентный в ткани компартмент Т-клеток γδ, и определить, может ли это быть защитным при раке груди. Это исследование было предпринято сначала для демонстрации воспроизводимого присутствия γδ Т-клеток в здоровой ткани груди с особым акцентом на использовании Vδ цепи. Затем мы определили их функциональный потенциал и совместимость с защитным иммунным надзором за опухолью.Эти подходы впоследствии были применены к γδ Т-клеткам в опухолях молочной железы. Колосированные αβ Т-клетки, выделенные с использованием того же протокола и поддерживаемые в тех же условиях культивирования, использовали в качестве контроля. Установив присутствие γδ Т-клеток в ткани груди и опухолях человека и их врожденную способность иммунного надзора in vitro, мы исследовали присутствие этих клеток in situ (через их перестройки ДНК) и сопоставили это с прогнозом в клинических образцах. Мы также искали доказательства in situ, в частности клональность репертуара TCR, для клеток, функционирующих с подобной врожденной способностью иммунного надзора, как было установлено ex vivo.Подробный план исследования, размеры выборки, повторения и критерии включения / исключения представлены в подписях к рисункам или в материалах и методах. Размер выборки и количество повторений экспериментов были достаточными для проведения строгого статистического анализа, как описано в подписях к рисункам и в материалах и методах. Все антитела и ключевые реагенты перечислены в таблице S6. Первичные данные представлены в файле данных S1.
Клинический материал
Образцы груди человека были получены от взрослых пациентов женского пола, подвергшихся уменьшению груди или уменьшающей риск мастэктомии (29 пациентов) или резекции опухоли груди (90 пациентов) после информированного согласия в рамках клинического исследования без вмешательства (исследование BTBC REC no. .: 13 / LO / 1248, IRAS ID 131133; главный исследователь: А.Т .; название исследования: «Анализ функциональной стромы иммунных клеток и взаимодействий злокачественных клеток при раке груди с целью обнаружения и разработки методов диагностики и лечения подтипов рака груди»). Это исследование было одобрено местным комитетом по этике исследований и проводилось в соответствии с принципами Хельсинкской декларации. Образцы были собраны после операции в стерильный физиологический раствор и немедленно отправлены на разделку. Гистопатолог или квалифицированный специалист по патологии идентифицировал и собирал опухолевый материал и ипсилатеральную несмежную нормальную ткань молочной железы из хирургических образцов, из которых впоследствии были выделены лимфоциты, как описано ниже.Демографические данные пациентов представлены в таблице S2. В дополнение к пациентам, указанным выше, для экспериментов по секвенированию TCR опухоль и парная ДНК незлокачественной ткани была извлечена из блоков FFPE из биобанка 11 пациентов с TNBC стадии I-III AJCC, у которых были мастэктомии, для которых мы могли получить доступ к достаточному материалу и точному клиническому наблюдению. данные вверх. Никаких других критериев не применялось. Эти 11 случаев также были частью клинического исследования BTBC, описанного выше.
Первичная экстракция и культивирование лимфоцитов
Для прямого выделения ex vivo свежую опухоль или ткань молочной железы грубо измельчали скальпелями, а затем диссоциировали с помощью набора для диссоциации опухолей человека MACS на диссоциаторе softMACS в соответствии с инструкциями производителя (Miltenyi Biotec).Образцы дважды промывали стерильным RPMI 1640 и сразу использовали для последующих анализов. Лимфоциты также собирали с использованием системы эксплантов с сеткой, адаптированной из протокола, впервые описанного Кларком и его коллегами ( 35 ). Вкратце, свежую опухоль или ткань груди измельчали с помощью скальпелей и помещали на покрытые коллагеном хвоста крысы (100 мкг / мл; BD Biosciences) решетки Cellfoam (Cytomatrix Pty Ltd.). Каждую сетку помещали в отдельную лунку 24-луночного планшета для тканевых культур и культивировали в полной среде [среда Дульбекко, модифицированная Исков (Life Technologies), 10% инактивированная нагреванием фетальная бычья сыворотка (FBS; Life Technologies), l-глутамин ( 292 мкг / мл; Life Technologies), пенициллин (100 Ед / мл; Life Technologies), стрептомицин (100 мкг / мл; Life Technologies) и 2-меркаптоэтанол (3.5 мкл / литр; Life Technologies)] с добавлением рекомбинантного человеческого IL-2 (rhIL-2) (100 МЕ / мл; Proleukin; Novartis Pharmaceuticals UK Ltd.) и rhIL-15 (10 нг / мл; BioLegend). Сетки выдерживали в течение 3 недель в культуре при 37 ° C / 5% CO 2 , и лимфоциты собирали, промывая лунки / сетки 0,01 мМ сбалансированным солевым раствором Hepes / Hanks.
Проточная цитометрия и сортировка клеток с активацией флуоресценции
Клетки промывали стерильным фосфатно-солевым буфером (PBS) для удаления следов сыворотки и окрашивали в течение 20 минут при комнатной температуре с помощью LIVE / DEAD Fixable Aqua (Thermo Fisher Scientific) в PBS .Последующее окрашивание поверхности проводили в буфере для сортировки клеток с активацией флуоресценции (FACS) (PBS, 2% эмбриональной телячьей сыворотки и 1 мМ EDTA) в течение 20 минут при 4 ° C (см. Таблицу S6) перед двукратной промывкой буфером FACS и фиксацией. с CellFIX (BD) в течение 10 мин при комнатной температуре. Для внутриклеточного окрашивания цитокинов фиксированные клетки дважды промывали промывочным буфером для повышения проницаемости для внутриклеточного окрашивания (BioLegend) и окрашивали в течение 20 мин при комнатной температуре перед двумя последующими промываниями промывочным буфером для повышения проницаемости для внутриклеточного окрашивания.Образцы собирали на BD FACSCanto II или BD LSRFortessa и анализировали с помощью программного обеспечения FlowJo (FlowJo LLC). Для FACS клетки не фиксировали и не сортировали на BD FACSAria II, как подробно описано ниже. Антитела перечислены в таблице S6 и использовались при разведении 1:50, если не указано иное.
Анализы активации лимфоцитов in vitro
Прямые выделенные и выделенные из эксплантата лимфоциты стимулировали PMA (10 нг / мл; Sigma) и иономицином (1 мкг / мл; Sigma) в присутствии брефельдина A (BFA; 20 мкг) / мл; Sigma) в течение 4 часов при 37 ° C / 5% CO 2 перед окрашиванием поверхностных маркеров и внутриклеточных цитокинов и сбором данных на BD FACSCanto или Fortessa.Для анализа связанного с планшетом лиганда NKG2D лимфоциты собирали из культур эксплантатов за 24 часа до активации и помещали в полную среду без добавления цитокинов. После отдыха лимфоциты переносили в 96-луночные планшеты с плоским дном для культивирования клеток (Corning), покрытые rhMICA (10 мкг / мл; R&D Systems), анти-CD3 (50 нг / мл; BioLegend), анти-CD3 (50 нг). / мл; BioLegend) и rhMICA (10 мкг / мл; R&D Systems) или мышиный иммуноглобулин G2a (IgG2a) (50 нг / мл; BioLegend) в количестве 100000 клеток на лунку в 100 мкл полной среды.Планшеты инкубировали в течение 6 часов при 37 ° C / 5% CO 2 в присутствии IL-15 (10 нг / мл; BioLegend) и BFA (20 мкг / мл; Sigma). Когда CD107a использовался в качестве функционального считывающего устройства, антитело против человеческого CD107a (конечная концентрация 1: 400; BioLegend) также добавляли в начале анализа вместе с монензином в количестве 1 × (BioLegend). Для условий блокирования NKG2D антитело против NKG2D (10 мкг / мл; клон 1D11; BioLegend) добавляли к лимфоцитам непосредственно перед посевом.
Для анализа активации цитокинов лимфоциты инкубировали с IL-12 (100 нг / мл; PeproTech) и / или IL-18 (100 нг / мл; медицинские и биологические лаборатории) в течение 24 часов при 37 ° C. 5% CO 2 с BFA (20 мкг / мл; Sigma), добавленным в течение последних 4 часов перед окрашиванием поверхностных и внутриклеточных цитокинов для проточной цитометрии.Для анализов перекоса IL-17 эксплантаты ткани молочной железы культивировали в полной среде и в условиях, описанных выше, с добавлением rhIL-2 ± rhIL-15, rhIL-1β, rhIL-6, rhIL-23 и rhTGF-β. в течение 3 дней. Эти клетки затем активировали PMA и иономицином в присутствии BFA (20 мкг / мл) в течение 4 часов при 37 ° C / 5% CO. 2 перед окрашиванием поверхностных и внутриклеточных цитокинов для проточной цитометрии.
Анализы цитотоксичности
Полученные из сеточного эксплантата γδ Т-клетки из тканей и опухолей груди были выделены с помощью FACS посредством истощения клеток TCRαβ + и NKp46 + .Клетки-мишени, MCF7 и HCC1954 [Cancer Research UK (CRUK) / Служба клеток института Фрэнсиса Крика], за 24 часа до этого высевали из расчета 10 000 клеток на лунку в 96-луночные планшеты с плоским дном (Corning). Пятьдесят тысяч отрицательно отсортированных γδ Т-клеток добавляли к клеткам-мишеням в присутствии или в отсутствие блокирующего антитела NKG2D (10 мкг / мл; BioLegend). Клетки инкубировали при 37 ° C в течение 48 часов, после чего супернатанты собирали и хранили при -20 ° C до дальнейшего анализа. Апоптоз клеток-мишеней измеряли с использованием иммуноферментного анализа (ELISA) для цитокератина 18, расщепленного каспазой, специфичного для эпителиальных клеток (Diapharma), в соответствии с инструкциями производителя.
Клеточные линии и условия культивирования
Целевые клеточные линии MCF7 и HCC1954 были получены от CRUK Cell Services (Клэр Холл, Лондон) и содержались в среде Игла, модифицированной Дульбекко (Life Technologies), с добавлением 10% инактивированного нагреванием FBS (Life Technologies ), пенициллин (100 Ед / мл; Life Technologies) и стрептомицин (100 мкг / мл; Life Technologies) при 37 ° C / 5% CO 2 . Клетки HCC1954 поддерживали в RPMI 1640 (Gibco) с добавлением 10% инактивированного нагреванием FBS (Life Technologies), пенициллина (100 Ед / мл; Life Technologies) и стрептомицина (100 мкг / мл; Life Technologies) при 37 ° C / 5% CO 2 .
Экстракция ДНК
ДНК была экстрагирована из FFPE парной опухолевой ткани и блоков нормальной ткани 11 пациентов с TNBC, которым была проведена мастэктомия в рамках исследования BTBC. Ткань опухоли подвергали микродиссекции иглой после разрезания. Набор QIAamp DNA FFPE Tissue (Qiagen) использовался в соответствии с инструкциями производителя для экстракции ДНК. ДНК количественно определяли с помощью флуорометра Qubit, и материал от пациентов с> 1 мкг ДНК как из опухоли, так и из нормальной ткани отправляли на количественное секвенирование локуса TCRα / δ компанией Adaptive Biotechnologies.
Секвенирование TCR
Секвенирование TCR было выполнено Adaptive Biotechnologies. Платформа Adaptive Biotechnologies использует геномную ДНК и может определять количество Т-клеток. Чтения были согласованы и аннотированы Adaptive Biotechnologies, а данные были выведены в виде файлов .csv (доступны по адресу https://osf.io/d4eu6/) для последующего анализа с помощью анализатора иммунного SEQ (https://clients.adaptivebiotech.com/login ). Выходные данные были отфильтрованы на CDR3 в кадре, а также на соединениях семейства TCRA и TCRA V-J для TCRαβ и соединениях семейства TCRD с TCRD V-J для TCRγδ Т-клеток.Мы нормализовали абсолютное количество TCR до 1 мкг входной ДНК для каждого образца, чтобы обеспечить нормализованное сравнение количества проникающих Т-клеток во всех образцах. Все анализы проводились с использованием аминокислотных последовательностей CDR3, а не нуклеотидных последовательностей.
Анализ клонального репертуара
Чтобы сравнить показатели клональности у каждого пациента между парной опухолью и доброкачественной тканью, мы провели пониженную выборку TCR из каждой пары образцов. Для Vδ1 выборка TCR из опухолевой и незлокачественной ткани была уменьшена до количества клонов в меньшей выборке с вероятностью получения клонотипа, равной его частоте в полной выборке.Даунсэмплинг с заменой проводился 200 раз. Для TCRαβ клонотипы были упорядочены по убыванию частоты, и верхние 10% общих TCR в каждой паре опухоли и доброкачественной ткани были использованы для выборки с понижением, как описано выше. Затем к данным с пониженной дискретизацией были применены показатели клональности и построены медианные значения. В качестве альтернативы мы также применили нормализованные меры клональности (нормализованная энтропия Шеннона, коэффициент Джини и D50) к необработанным данным. TreeMaps были созданы с помощью программы Macrofocus TreeMap (www.treemap.com). Мы визуализировали пациентов с общими клонотипами с помощью пакета UpSet R (https://ieeexplore.ieee.org/abstract/document/6876017 и https://vcg.github.io/upset).
Статистический анализ
Статистическая значимость была определена с помощью критерия Колмогорова-Смирнова, лог-рангового критерия, критерия парных знаков Уилкоксона или критерия Манна-Уитни, как указано в подписях к рисункам, с использованием программного обеспечения Prism 7 (GraphPad). Все результаты считались значимыми при пороговом значении P <0.05. Если показаны результаты статистического теста, * P <0,05, ** P ≤ 0,01, *** P ≤ 0,001 и **** P ≤ 0,0001, если не указано иное.
ДОПОЛНИТЕЛЬНЫЕ МАТЕРИАЛЫ
stm.sciencemag.org/cgi/content/full/11/513/eaax9364/DC1
Рис. S1. Культура эксплантата позволила выделить значительное количество лимфоцитов, резидентных в тканях человека.
Рис. S2. Vδ1 + Т-клетки проявляют врожденную чувствительность.
Рис. S3. αβ и γδ Т-клетки могут быть выделены из опухолей груди и фенотипически напоминать клетки из здоровой ткани.
Рис. S4. Как αβ, так и γδ Т-клетки обогащены опухолями по сравнению с парными доброкачественными тканями.
Рис. S5. Vδ1 + Т-клетки не обнаруживают признаков опухолевой клональной фокусировки в отличие от αβ Т-клеток.
Рис. S6. У пациентов наблюдается ограниченное перекрытие репертуара Vδ1 между опухолью и парной доброкачественной тканью.
Таблица S1.Подтипы лимфоцитов наблюдались ex vivo после ферментативного расщепления и в культурах эксплантатов сетки.
Таблица S2. Клинические особенности когорты KCL TNBC.
Таблица S3. Показатели клональности TCR с пониженной выборкой.
Таблица S4. Показатели клональности необработанных TCR.
Таблица S5. Общедоступные внутриопухолевые фосфо-антиген-реактивные последовательности Vδ2 CDR3 и образцы, в которых они использовались.
Таблица S6. Таблица антител и основных реагентов.
Файл данных S1. Первичные данные.
Благодарности: Мы благодарим А.Клиффорду, С. Иршаду и Т. Алагутураи за согласие и набор пациентов, а также нынешних и бывших коллег за полезные советы и обсуждения. Мы благодарим службы проточной цитометрии отделения иммунобиологии Питера Горера и Центр биомедицинских исследований больницы Гая и Святого Томаса (BRC) за выдающуюся поддержку. Финансирование: Работа была поддержана премией исследователей Wellcome Trust (WT) (106292 / Z / 14 / Z) для AH и Институтом Фрэнсиса Крика, который получает основное финансирование от CRUK (FC001093), Британского медицинского исследования. Совет (FC001093) и WT (FC001093).F.K.-C. финансировалась стипендией BRC. Ю.В. финансировалась стипендиатом Национального института исследований в области здравоохранения. R.T.W. финансировалась стипендией Совета медицинских исследований (Великобритания) и лекциями Национального института исследований в области здравоохранения. А.Г. и А.Т. были профинансированы фондом «Рак груди сейчас» Королевского колледжа Лондона. Работа проводилась в рамках Ускорителя иммунной терапии рака CRUK (CITA) и Главного центра Лондонского Сити CRUK. Вклад авторов: F.K.-C., Y.W., R.T.W. и A.Х. задумал исследование и спланировал эксперименты. F.K.-C. и Ю.В. проводил и анализировал эксперименты. C.N.-L., J.O., and P.G. обработали образцы пациентов и предоставили техническую помощь. А.Г., А.Л., Д.Б., П.В. помогли с анализом данных. В. набирали пациентов. Рукопись написали F.K.-C., Y.W. и A.H. В. и А.Х. руководили исследованием.