
молекула АТФ – что это такое Каково значение атф
АТФ или по полной расшифровке аденозинтрифосфорная кислота, является «аккумулятором» энергии в клетках организма. Ни одна биохимическая реакция не проходит без участия АТФ. Молекулы АТФ находятся в ДНК и РНК.
Состав АТФ
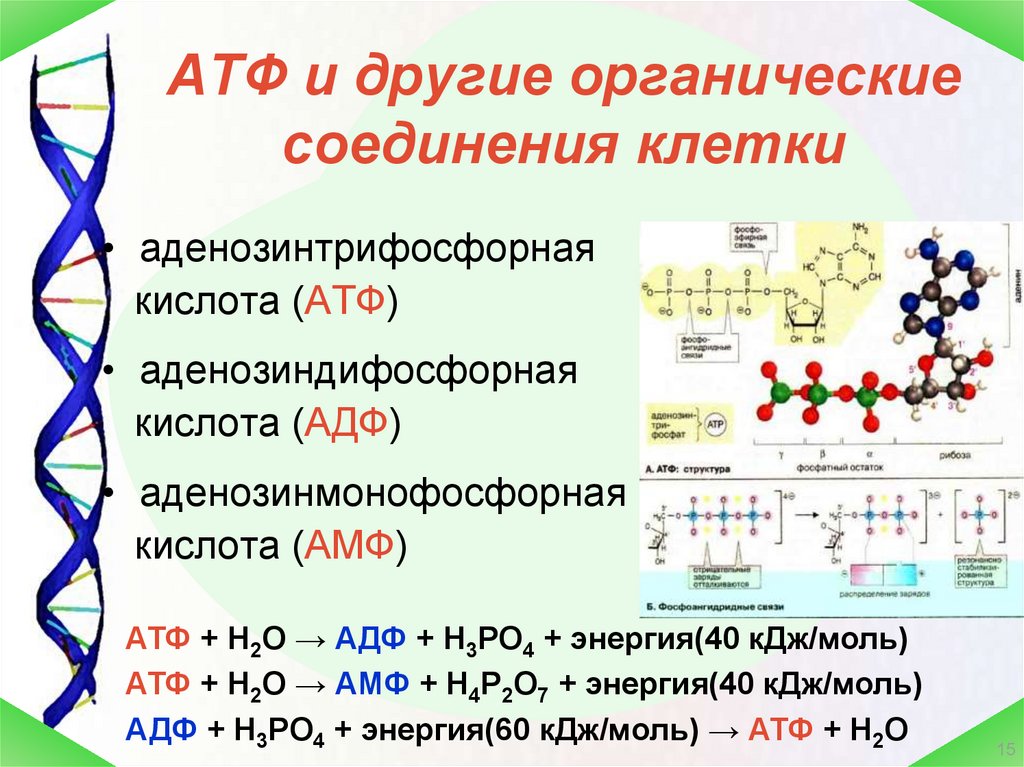
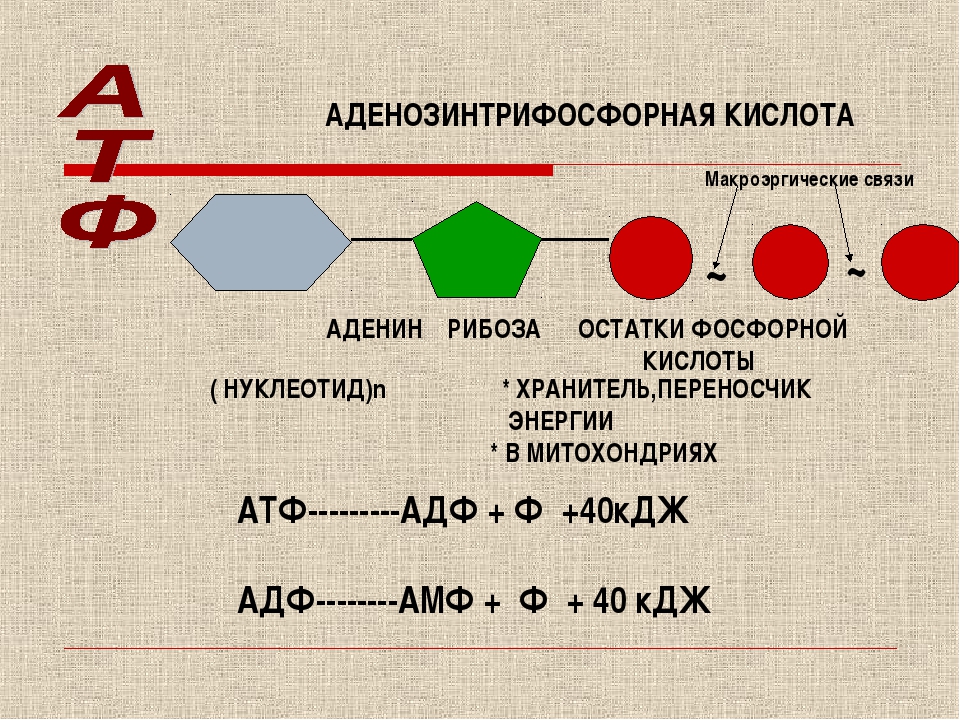
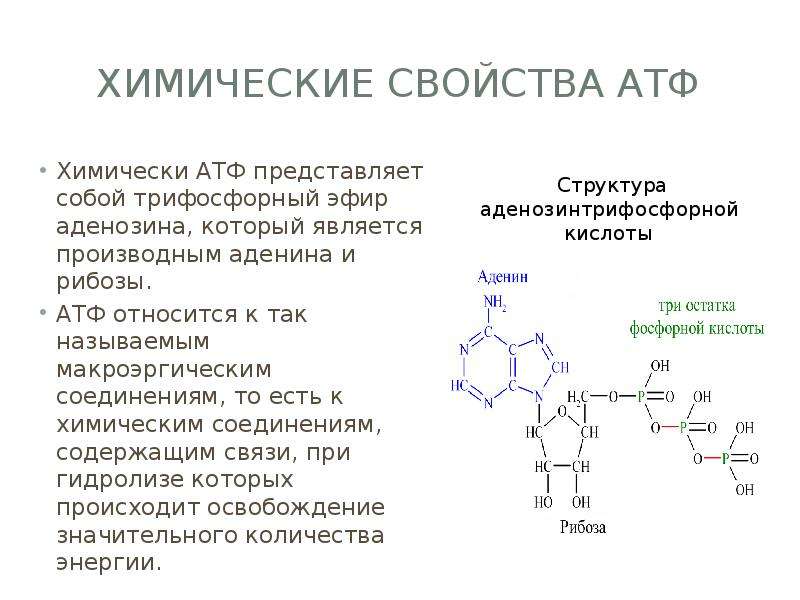
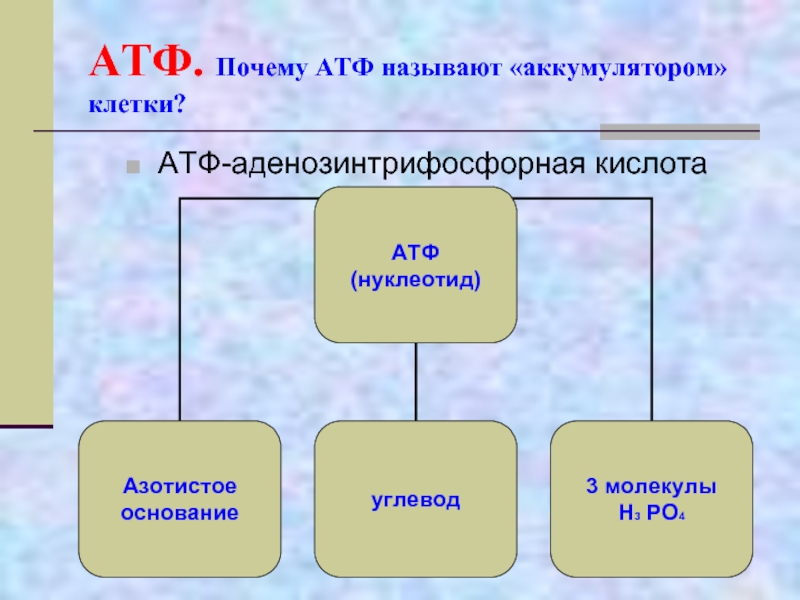
Молекула АТФ имеет три составляющих: три остатка фосфорной кислоты, аденин и рибоза. То есть, АТФ имеет строение нуклеотида и относится к нуклеиновым кислотам. Рибоза-это углевод,а аденин-азотистое основание. Остатки кислоты объединены друг с другом неустойчивыми энергетическими связями. Энергия появляется при отщеплении молекул кислоты. Отделение происходит благодаря биокатализаторам. После отъединения, молекула АТФ уже превращается в АДФ (если отщепилась одна молекула) или в АМФ (если отщепились две молекулы кислоты). При отделении одной молекулы фосфорной кислоты выходит 40 кДж энергии.
Роль в организме
АТФ играет не только энергетическую роль в организме,но и ряд других:
- является результатом синтезирования нуклеиновых кислот.

- регулирование многие биохимических процессов.
- сигнального вещества в других взаимодействиях клеток.
Синтез АТФ
Получение АТФ проходит в хлоропластах и митохондриях. Важнейший процесс в синтезировании молекул АТФ — это диссимиляции. Диссимиляция — это разрушение сложного до более простого.
Синтез АТФ проходит не в один этап, а в три этапа:
- Первый этап — подготовительный. Под действием ферментов в пищеварении происходит распад того, что мы поглотили. При этом жиры разлагаются до глицерина и жирных кислот, белки до аминокислот, а крахмал до глюкозы. То есть, всё подготавливается для дальнейшего использования. Выделяется тепловая энергия
- Второй этап — это гликолиз (безкислородный). Вновь происходит распад, но здесь распаду подвергается ещё и глюкоза. Так же участвуют ферменты. Но 40 % энергии остаются в АТФ, а остальное расходуется в тепло.
- Третий этап — гидролиз (кислородный). Он происходит уже в самих митохондриях.
 Здесь участие принимает и кислород, который мы вдыхаем, и ферменты. После полной диссимиляции выделяется энергия для образования АТФ.
Здесь участие принимает и кислород, который мы вдыхаем, и ферменты. После полной диссимиляции выделяется энергия для образования АТФ.
АТФ и другие соединения клетки (витамины)
Особо важную роль в биоэнергетике клетки играет адениловый нуклеотид, к которому присоединены два остатка фосфорной кислоты. Такое вещество называют аденозинтрифосфорной кислотой (АТФ).
В химических связях между остатками фосфорной кислоты молекулы АТФ запасена энергия, которая освобождается при отщеплении органического фосфата: АТФ = АДФ + Ф + Е, где Ф — фермент, Е — освобождающаяся энергия. В этой реакции образуется аденозиндифосфорная кислота (АДФ) — остаток молекулы АТФ и органический фосфат.
Энергию АТФ все клетки используют для процессов биосинтеза, движения, производства тепла, нервных импульсов, свечений (например, у люминесцентных бактерий), т.е. для всех процессов жизнедеятельности.
АТФ — универсальный биологический аккумулятор энергии, который синтезируется в митохондриях (внутриклеточных органоидах).
Митохондрия, таким образом, исполняет в клетке роль «энергетической станции». Принцип образования АТФ в хлоропластах клеток растений в общем тот же — использование протонного градиента и преобразование энергии электрохимического градиента в энергию химических связей.
Световая энергия Солнца и энергия, заключенная в потребляемой пище, запасается в молекулах АТФ. Запас АТФ в клетке невелик. Так, в мышце запаса АТФ хватает на 20-30 сокращений. При усиленной, но кратковременной работе мышцы работают исключительно за счет расщепления содержащейся в них АТФ. После окончания работы человек усиленно дышит — в этот период происходит расщепление углеводов и других веществ (происходит накопление энергии) и запас АТФ в клетках восстанавливается протонов. Протоны проходят через этот канал под действием движущей силы электрохимического градиента. Энергия этого процесса используется ферментом, содержащимся в тех же самых белковых комплексах и способным присоединить фосфатную группу к аденозиндифосфату (АДФ), что и приводит к синтезу АТФ.
Витамины: Vita — жизнь.
Витамины — биологически активные вещества, синтезирующиеся в организме или поступающие с пищей, которые в малых количествах необходимы для нормального обмена веществ и жизнедеятельности организма.
В 1911г. Польский химик К. Функ выделил из рисовых отрубей вещество, излечивающее параличи голубей, питавшихся только полированным рисом. Химический анализ этого вещества показал, что в его состав входит азот.
Открытое им вещество Функ назвал витамином (от слов «вита»- жизнь и «амин»- содержащий азот.
Биологическая роль витаминов заключается в их регулярном действии на обмен веществ. Витамины обладают каталитическими свойствами, то есть способностью стимулировать химические реакции, протекающие в организме, а также активно участвуют в образовании и функции ферментов. Витамины влияют на усвоение организмом питательных веществ, способствуют нормальному росту клеток и развитию всего организма. Являясь составной частью ферментов, витамины определяют их нормальную функцию и активность. Таким образом, недостаток в организме какого-либо витамина ведет к нарушению процессов обмена веществ.
Являясь составной частью ферментов, витамины определяют их нормальную функцию и активность. Таким образом, недостаток в организме какого-либо витамина ведет к нарушению процессов обмена веществ.
Группы витаминов:
СУТОЧНАЯ ПОТРЕБНОСТЬ В ВИТАМИНАХ
С — аскорбиновая кислота: 70 — 100 мг.
В — тиамин: 1,5 — 2,6 мг.
В — рибофлавин: 1,8 — 3 мг.
А — ретинол: 1,5 мг.
D — кальциферол: для детей и взрослых 100 МЕ,
до 3 лет 400 МЕ.
Е — токоферол: 15 — 20 мг.
В биологии АТФ — это источник энергии и основа жизни. АТФ — аденозинтрифосфат — участвует в процессах метаболизма и регулирует биохимические реакции в организме.
Что это?
Понять, что такое АТФ, поможет химия. Химическая формула молекулы АТФ — C10h26N5O13P3. Запомнить полное название несложно, если разбить его на составные части. Аденозинтрифосфат или аденозинтрифосфорная кислота — нуклеотид, состоящий из трёх частей:
- аденина — пуринового азотистого основания;
- рибозы — моносахарида, относящегося к пентозам;
- трёх остатков фосфорной кислоты.

Рис. 1. Строение молекулы АТФ.
Более подробная расшифровка АТФ представлена в таблице.
АТФ впервые обнаружили гарвардские биохимики Суббарао, Ломан, Фиске в 1929 году. В 1941 году немецкий биохимик Фриц Липман установил, что АТФ является источником энергии живого организма.
Образование энергии
Фосфатные группы соединены между собой высокоэнергетическими связями, которые легко разрушаются. При гидролизе (взаимодействии с водой) связи фосфатной группы распадаются, высвобождая большое количество энергии, а АТФ превращается в АДФ (аденозиндифосфорную кислоту).
Условно химическая реакция выглядит следующим образом:
ТОП-4 статьи которые читают вместе с этой
АТФ + Н2О → АДФ + Н3РО4 + энергия
Рис. 2. Гидролиз АТФ.
Часть высвободившейся энергии (около 40 кДж/моль) участвует в анаболизме (ассимиляции, пластическом обмене), часть — рассеивается в виде тепла для поддержания температуры тела.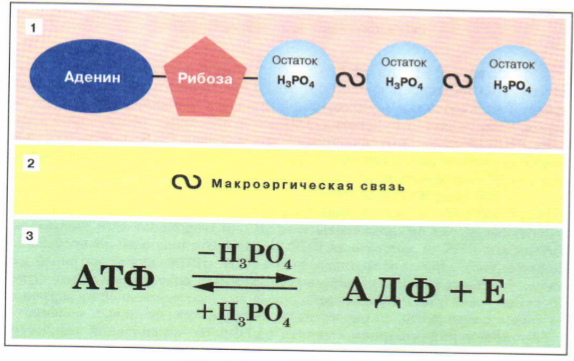
Синтез АТФ
АТФ располагается в цитоплазме, ядре, хлоропластах, в митохондриях. Синтез АТФ в животной клетке происходит в митохондриях, а в растительной — в митохондриях и хлоропластах.
АТФ образуется из АДФ и фосфата с затратой энергии. Такой процесс называется фосфорилированием:
АДФ + Н3РО4 + энергия → АТФ + Н2О
Рис. 3. Образование АТФ из АДФ.
В растительных клетках фосфорилирование происходит при фотосинтезе и называется фотофосфорилированием. У животных процесс протекает при дыхании и называется окислительным фосфорилированием.
В животных клетках синтез АТФ происходит в процессе катаболизма (диссимиляции, энергетического обмена) при расщеплении белков, жиров, углеводов.
Функции
Из определения АТФ понятно, что эта молекула способна давать энергию.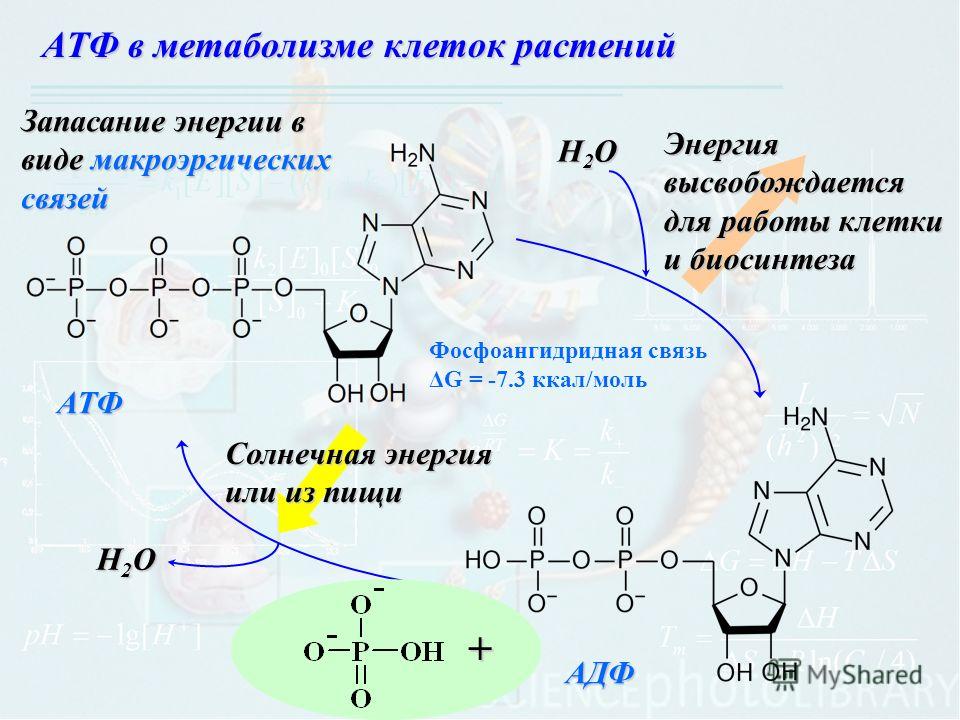
- является материалом для синтеза нуклеиновых кислот;
- является частью ферментов и регулирует химические процессы, ускоряя или замедляя их протекание;
- является медиатором — передаёт сигнал синапсам (местам контакта двух клеточных мембран).
Что мы узнали?
Из урока биологии 10 класса узнали о строении и функциях АТФ — аденозинтрифосфорной кислоты. АТФ состоит из аденина, рибозы и трёх остатков фосфорной кислоты. При гидролизе фосфатные связи разрушаются, что высвобождает энергию, необходимую для жизнедеятельности организмов.
Тест по теме
Оценка доклада
Средняя оценка: 4.6 . Всего получено оценок: 621.
АТФ — это сокращённое название Аденозин Три-Фосфорной кислоты. А также можно встретить название Аденозинтрифосфат. Это нуклеоид, который играет огромную роль в обмене энергией в организме. Аденозин Три-Фосфорная кислота — это универсальный источник энергии, участвующий во всех биохимических процессах организма.
Структура и формула АТФ
Если говорить об АТФ более подробно , то это молекула, которая даёт энергию всем процессам, происходящим в организме, в том числе она же даёт энергию для движения. При расщеплении молекулы АТФ происходит сокращение мышечного волокна, вследствие чего выделяется энергия, позволяющая произойти сокращению. Синтезируется Аденозинтрифосфат из инозина — в живом организме.
Для того чтобы дать организму энергию Аденозинтрифосфату необходимо пройти несколько этапов. Вначале отделяется один из фосфатов — с помощью специального коэнзима. Каждый из фосфатов даёт десять калорий. В процессе вырабатывается энергия и получается АДФ (аденозин дифосфат).
Если организму для действия нужно больше энергии , то отделяется ещё один фосфат. Тогда формируется АМФ (аденозин монофосфат). Главный источник для выработки Аденозинтрифосфата — это глюкоза, в клетке она расщепляется на пируват и цитозол. Аденозинтрифосфат насыщает энергией длинные волокна, которые содержат протеин — миозин. Именно он формирует мышечные клетки.
Аденозинтрифосфат насыщает энергией длинные волокна, которые содержат протеин — миозин. Именно он формирует мышечные клетки.
В моменты, когда организм отдыхает, цепочка идёт в обратную сторону, т. е. формируется Аденозин Три-Фосфорная кислота. Опять же в этих целях используется глюкоза. Созданные молекулы Аденозинтрифосфата будут вновь использоваться, как только это станет необходимо. Когда энергия не нужна, она сохраняется в организме и высвобождается как только это потребуется.
Молекула АТФ состоит из нескольких, а точнее, трёх компонентов:
- Рибоза — это пятиуглеродный сахар, такой же лежит в основе ДНК.
- Аденин — это объединённые атомы азота и углерода.
- Трифосфат.
В самом центре молекулы Аденозинтрифосфата находится молекула рибозы, а её край является основной для аденозина. С другой стороны рибозы расположена цепочка из трёх фосфатов.
Системы АТФ
При этом нужно понимать, что запасов АТФ будет достаточно только первые две или три секунды двигательной активности, после чего её уровень снижается. Но при этом работа мышц может осуществляться только с помощью АТФ. Благодаря специальным системам в организме постоянно синтезируются новые молекулы АТФ. Включение новых молекул происходит в зависимости от длительности нагрузки.
Но при этом работа мышц может осуществляться только с помощью АТФ. Благодаря специальным системам в организме постоянно синтезируются новые молекулы АТФ. Включение новых молекул происходит в зависимости от длительности нагрузки.
Молекулы АТФ синтезируют три основные биохимические системы:
- Фосфагенная система (креатин-фосфат).
- Система гликогена и молочной кислоты.
- Аэробное дыхание.
Рассмотрим каждую из них в отдельности.
Фосфагенная система — в случае если мышцы будут работать недолго, но крайне интенсивно (порядка 10 секунд), будет использоваться фосфагенная система. В этом случае АДФ связывается с креатин фосфатом. Благодаря этой системе происходит постоянная циркуляция небольшого количества Аденозинтрифосфата в мышечных клетках. Так как в самих мышечных клетках тоже имеется фосфат креатина, он используется, чтобы восстановить уровень АТФ после высокоинтенсивной короткой работы. Но уже секунд через десять уровень креатин фосфата начинает снижаться — такой энергии хватает на короткий забег или интенсивную силовую нагрузку в бодибилдинге.
Гликоген и молочная кислота — снабжает энергией организм медленнее, чем предыдущая. Она синтезирует АТФ, которой может хватить на полторы минуты интенсивной работы. В процессе глюкоза в мышечных клетках формируется в молочную кислоту за счёт анаэробного метаболизма .
Так как в анаэробном состоянии кислород организмом не используется, то данная система даёт энергию так же как и в аэробной системе, но время экономится. В анаэробном режиме мышцы сокращаются крайне мощно и быстро. Такая система может позволить пробежать четыреста метров спринта или более длительную интенсивную тренировку в зале. Но долгое время работать таким образом не позволит болезненность в мышцах, которая появляется из-за переизбытка молочной кислоты.
Аэробное дыхание — эта система включается, если тренировка продолжается более двух минут. Тогда мышцы начинают получать Аденозинтрифосфат из углеводов, жиров и протеинов. В этом случае АТФ синтезируется медленно, зато энергии хватает надолго — физическая активность может продолжаться несколько часов. Это происходит благодаря тому, что глюкоза распадается без препятствий, у неё нет никаких противодействий, препятствующих со стороны — как препятствует молочная кислота в анаэробном процессе.
Это происходит благодаря тому, что глюкоза распадается без препятствий, у неё нет никаких противодействий, препятствующих со стороны — как препятствует молочная кислота в анаэробном процессе.
Роль АТФ в организме
Из предыдущего описания понятно, что основная роль аденозинтрифосфата в организме — это обеспечение энергией всех многочисленных биохимических процессов и реакций в организме. Большинство энергозатратных процессов у живых существ происходят благодаря АТФ.
Но помимо этой главной функции, аденозинтрифосфат выполняет и другие:
Роль АТФ в организме и жизни человека хорошо известна не только учёным, но и многим спортсменам и бодибилдерам, так как её понимание помогает сделать тренировки более эффективными и правильно рассчитывать нагрузки. Для людей, которые занимаются силовыми тренировками в зале, спринтерскими забегами и другими видами спорта, очень важно понимать, какие упражнения требуется выполнять в тот или иной момент времени. Благодаря этому можно сформировать желаемое строение тела, проработать мышечную структуру, снизить излишний вес и добиться других желаемых результатов.
В любой клетке нашего организма протекают миллионы биохимических реакций. Они катализируются множеством ферментов, которые зачастую требуют затрат энергии. Где же клетка ее берет? На этот вопрос можно ответить, если рассмотреть строение молекулы АТФ — одного из основных источников энергии.
АТФ — универсальный источник энергии
АТФ расшифровывается как аденозинтрифосфат, или аденозинтрифосфорная кислота. Вещество является одним из двух наиболее важных источников энергии в любой клетке. Строение АТФ и биологическая роль тесно связаны. Большинство биохимических реакций может протекать только при участии молекул вещества, особенно это касается Однако АТФ редко непосредственно участвует в реакции: для протекания любого процесса нужна энергия, заключенная именно в аденозинтрифосфата.
Строение молекул вещества таково, что образующиеся связи между фосфатными группами несут огромное количество энергии. Поэтому такие связи также называются макроэргическими, или макроэнергетическими (макро=много, большое количество).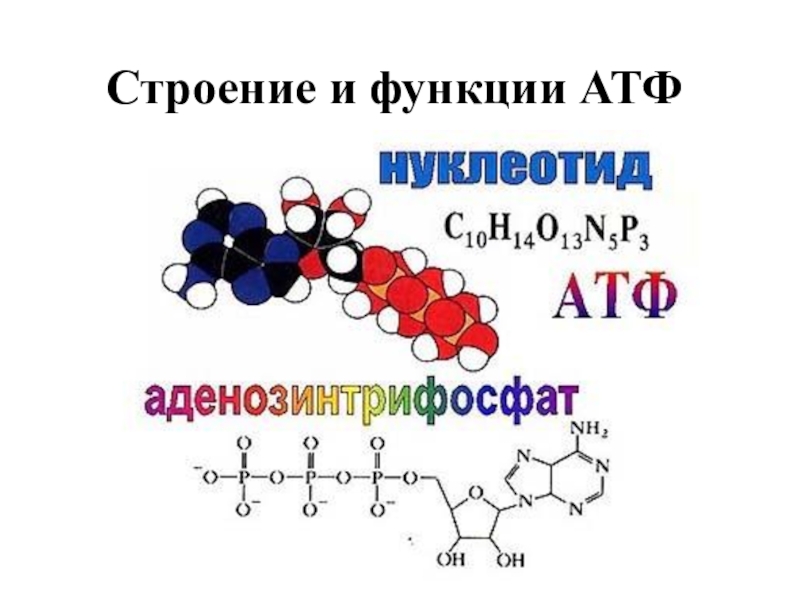 Термин впервые ввел ученый Ф. Липман, и он же предложил использовать значок ̴ для их обозначения.
Термин впервые ввел ученый Ф. Липман, и он же предложил использовать значок ̴ для их обозначения.
Очень важно для клетки поддерживать постоянный уровень содержания аденозинтрифосфата. Особенно это характерно для клеток мышечной ткани и нервных волокон, потому что они наиболее энергозависимы и для выполнения своих функций нуждаются в высоком содержании аденозинтрифосфата.
Строение молекулы АТФ
Аденозинтрифосфат состоит из трех элементов: рибозы, аденина и остатков
Рибоза — углевод, который относится к группе пентоз. Это значит, что в составе рибозы 5 атомов углерода, которые заключены в цикл. Рибоза соединяется с аденином β-N-гликозидной связь на 1-ом атоме углерода. Также к пентозе присоединяются остатки фосфорной кислоты на 5-ом атоме углерода.
Аденин — азотистое основание. В зависимости от того, какое азотистое основание присоединяется к рибозе, выделяют также ГТФ (гуанозинтрифосфат), ТТФ (тимидинтрифосфат), ЦТФ (цитидинтрифосфат) и УТФ (уридинтрифосфат). Все эти вещества схожи по строению с аденозинтрифосфатом и выполняют примерно такие же функции, однако они встречаются в клетке намного реже.
Все эти вещества схожи по строению с аденозинтрифосфатом и выполняют примерно такие же функции, однако они встречаются в клетке намного реже.
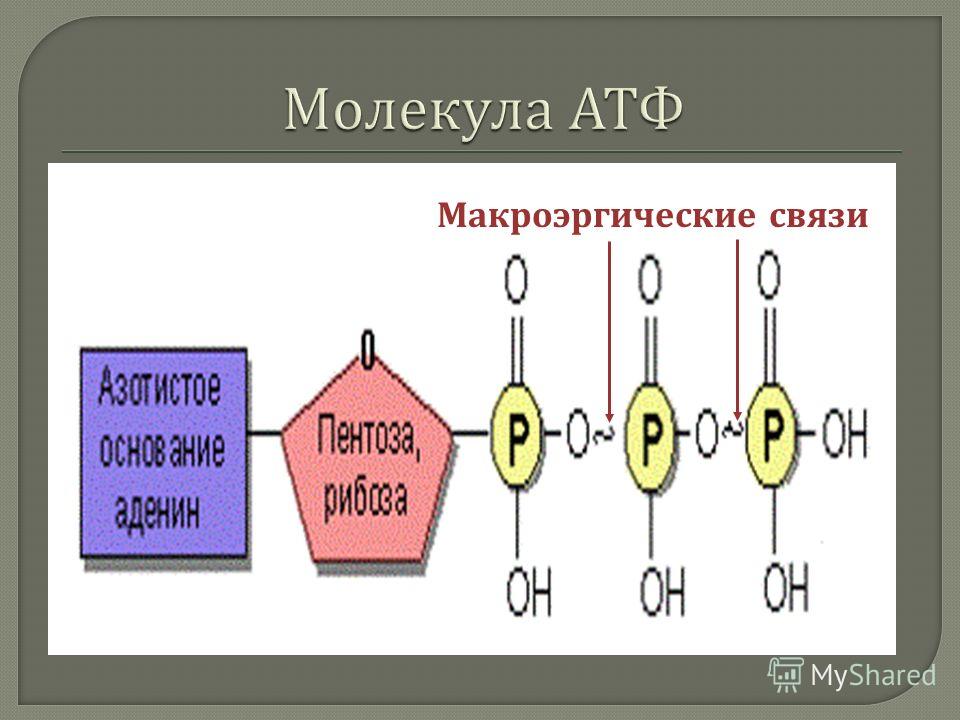
Остатки фосфорной кислоты . К рибозе может присоединиться максимально три остатка фосфорной кислоты. Если их два или только один, то соответственно вещество называется АДФ (дифосфат) или АМФ (монофосфат). Именно между фосфорными остатками заключены макроэнергетические связи, после разрыва которых высвобождается от 40 до 60 кДж энергии. Если разрываются две связи, выделяется 80, реже — 120 кДж энергии. При разрыве связи между рибозой и фосфорным остатком выделяется всего лишь 13,8 кДж, поэтому в молекуле трифосфата только две макроэргические связи (Р ̴ Р ̴ Р), а в молекуле АДФ — одна (Р ̴ Р).
Вот каковы особенности строения АТФ. По причине того, что между остатками фосфорной кислоты образуется макроэнергетическая связь, строение и функции АТФ связаны между собой.
Строение АТФ и биологическая роль молекулы. Дополнительные функции аденозинтрифосфата
Кроме энергетической, АТФ может выполнять множество других функций в клетке.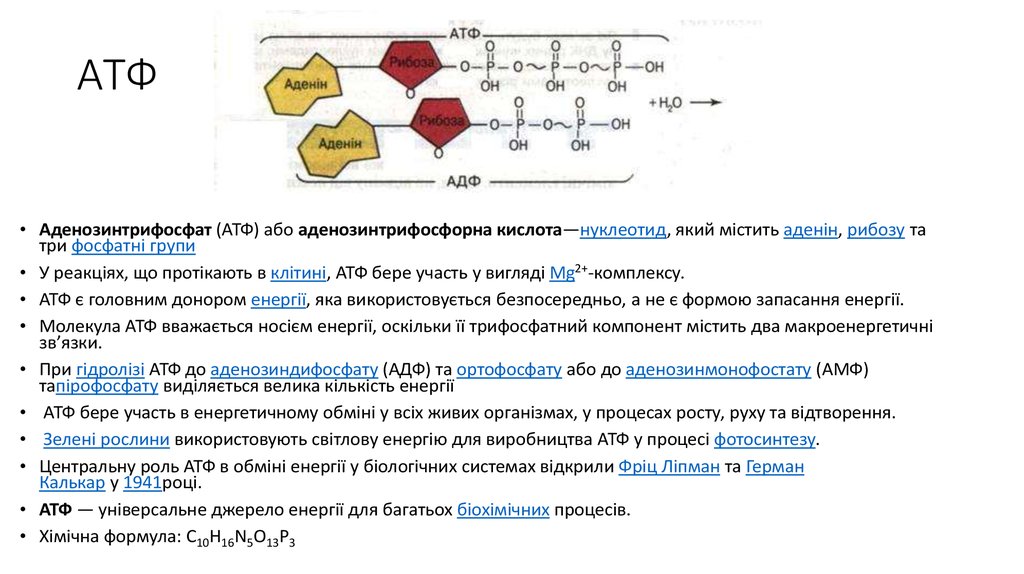 Наряду с другими нуклеотидтрифосфатами трифосфат участвует в построении нуклеиновый кислот. В этом случае АТФ, ГТФ, ТТФ, ЦТФ и УТФ являются поставщиками азотистых оснований. Это свойство используется в процессах и транскрипции.
Наряду с другими нуклеотидтрифосфатами трифосфат участвует в построении нуклеиновый кислот. В этом случае АТФ, ГТФ, ТТФ, ЦТФ и УТФ являются поставщиками азотистых оснований. Это свойство используется в процессах и транскрипции.
Также АТФ необходим для работы ионных каналов. Например, Na-K канал выкачивает 3 молекулы натрия из клетки и вкачивает 2 молекулы калия в клетку. Такой ток ионов нужен для поддержания положительного заряда на наружной поверхности мембраны, и только с помощью аденозинтрифосфата канал может функционировать. То же касается протонных и кальциевых каналов.
АТФ является предшественником вторичного мессенжера цАМФ (циклический аденозинмонофосфат) — цАМФ не только передает сигнал, полученный рецепторами мембраны клетки, но и является аллостерическим эффектором. Аллостерические эффекторы — это вещества, которые ускоряют или замедляют ферментативные реакции. Так, циклический аденозинтрифосфат ингибирует синтез фермента, который катализирует расщепление лактозы в клетках бактерии.
Сама молекула аденозинтрифосфата также может быть аллостерическим эффектором. Причем в подобных процессах антагонистом АТФ выступает АДФ: если трифосфат ускоряет реакцию, то дифосфат затормаживает, и наоборот. Таковы функции и строение АТФ.
Как образуется АТФ в клетке
Функции и строение АТФ таковы, что молекулы вещества быстро используются и разрушаются. Поэтому синтез трифосфата — это важный процесс образования энергии в клетке.
Выделяют три наиболее важных способа синтеза аденозинтрифосфата:
1. Субстратное фосфорилирование.
2. Окислительное фосфорилирование.
3. Фотофосфорилирование.
Субстратное фосфорилирование основано на множественных реакциях, протекающих в цитоплазме клетки. Эти реакции получили название гликолиза — анаэробный этап В результате 1 цикла гликолиза из 1 молекулы глюкозы синтезируется две молекулы которые дальше используются для получения энергии, и также синтезируются два АТФ.
- С 6 Н 12 О 6 + 2АДФ + 2Фн —> 2С 3 Н 4 O 3 + 2АТФ + 4Н.

Дыхание клетки
Окислительное фосфорилирование — это образование аденозинтрифосфата путем передачи электронов по электронно-транспортной цепи мембраны. В результате такой передачи формируется градиент протонов на одной из сторон мембраны и с помощью белкового интегрального комплекта АТФ-синтазы идет построение молекул. Процесс протекает на мембране митохондрий.
Последовательность стадий гликолиза и окислительного фосфорилирования в митохондриях составляет общий процесс под названием дыхание. После полного цикла из 1 молекулы глюкозы в клетке образуется 36 молекул АТФ.
Фотофосфорилирование
Процесс фотофосфорилирования — это то же окислительное фосфорилирование лишь с одним отличием: реакции фотофосфорилирования протекают в хлоропластах клетки под действием света. АТФ образуется во время световой стадии фотосинтеза — основного процесса получения энергии у зеленых растений, водорослей и некоторых бактерий.
В процессе фотосинтеза все по той же электронно-транспортной цепи проходят электроны, в результате чего формируется протонный градиент. Концентрация протонов на одной из сторон мембраны является источником синтеза АТФ. Сборка молекул осуществляется посредством фермента АТФ-синтазы.
Концентрация протонов на одной из сторон мембраны является источником синтеза АТФ. Сборка молекул осуществляется посредством фермента АТФ-синтазы.
В среднестатистической клетке содержится 0,04% аденозинтрифосфата от всей массы. Однако самое большое значение наблюдается в мышечных клетках: 0,2-0,5%.
В клетке около 1 млрд молекул АТФ.
Каждая молекула живет не больше 1 минуты.
Одна молекула аденозинтрифосфата обновляется в день 2000-3000 раз.
В сумме за сутки организм человека синтезирует 40 кг аденозинтрифосфата, и в каждый момент времени запас АТФ составляет 250 г.
Заключение
Строение АТФ и биологическая роль его молекул тесно связаны. Вещество играет ключевую роль в процессах жизнедеятельности, ведь в макроэргических связях между фосфатными остатками содержится огромное количество энергии. Аденозинтрифосфат выполняет множество функций в клетке, и поэтому важно поддерживать постоянную концентрацию вещества. Распад и синтез идут с большой скоростью, т.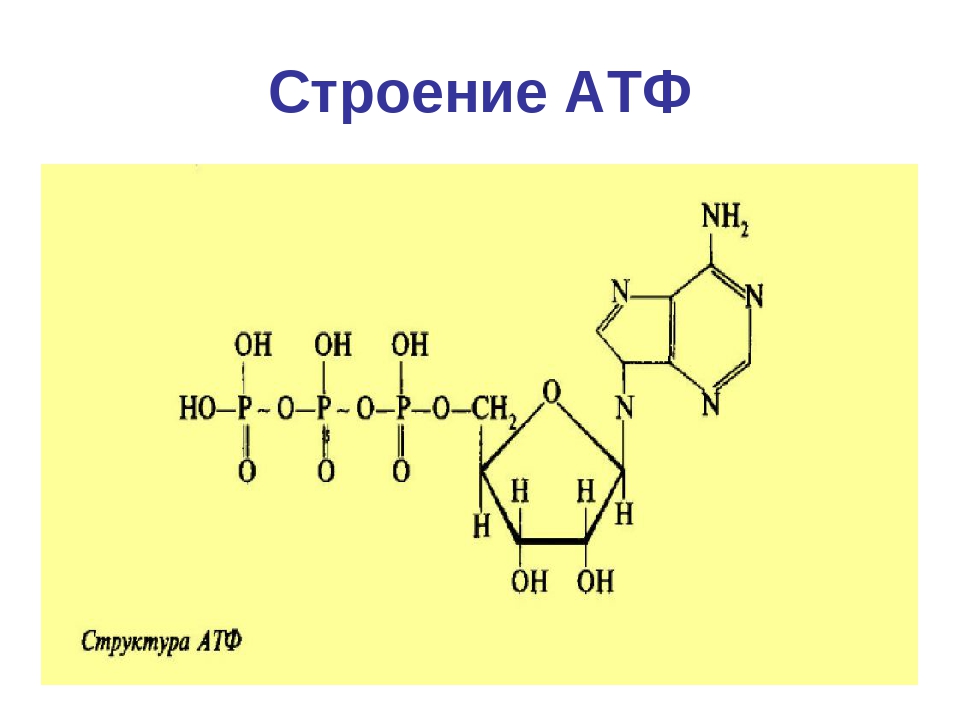 к. энергия связей постоянно используется в биохимических реакциях. Это незаменимое вещество любой клетки организма. Вот, пожалуй, и все, что можно сказать о том, какое строение имеет АТФ.
к. энергия связей постоянно используется в биохимических реакциях. Это незаменимое вещество любой клетки организма. Вот, пожалуй, и все, что можно сказать о том, какое строение имеет АТФ.
АТФ и ее роль в клетке
В цитоплазме каждой клетки, а также в митохондриях, хлоропластах и ядрах содержится аденозинтрифосфорная кислота (АТФ). Она поставляет энергию для большинства реакций, происходящих в клетке. С помощью АТФ клетка синтезирует новые молекулы белков, углеводов, жиров, избавляется от отходов, осуществляет активный транспорт веществ, биение жгутиков и ресничек и т. д.
Молекула АТФ представляет собой нуклеотид, образованный азотистым основанием аденином, пятиуглеродным сахаром рибозой и тремя остатками фосфорной кислоты. Фосфатные группы в молекуле АТФ соединены между собой высокоэнергетическими (макроэргическими) связями :
Связи между фосфатными группами не очень прочные, и при их разрыве выделяется большое количество энергии.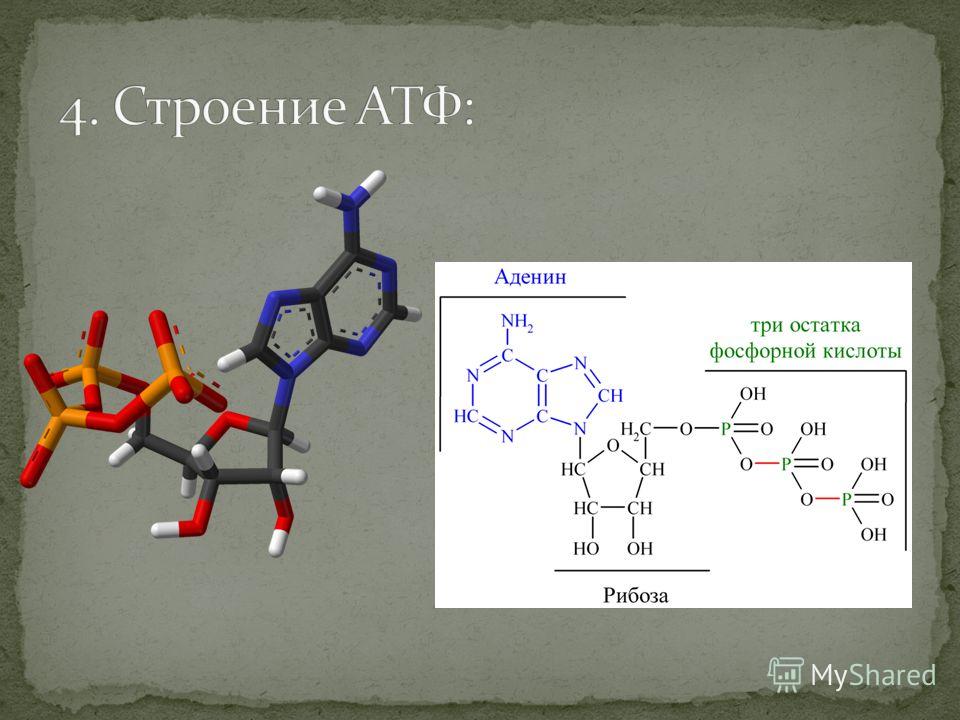 В результате гидролитического отщепления от АТФ фосфатной группы образуется аденозиндифосфорная кислота (АДФ) н высвобождается порция энергии:
В результате гидролитического отщепления от АТФ фосфатной группы образуется аденозиндифосфорная кислота (АДФ) н высвобождается порция энергии:
АДФ также может подвергаться дальнейшему гидролизу с отщеплением еще одной фосфатной группы и выделением второй порции энергии; при этом АДФ преобразуется в аденозин-монофосфат (АМФ), который далее не гидролизуется:
АТФ образуется из АДФ и неорганического фосфата за счет энергии, освобождающейся при окислении органических веществ и в процессе фотосинтеза. Этот процесс называется фосфорилированием. При этом должно быть затрачено не менее 40 кДж/моль энергии, которая аккумулируется в макроэргических связях:
Следовательно, основное значение процессов дыхания и фотосинтеза определяется тем, что они поставляют энергию для синтеза АТФ, с участием которой в клетке выполняется большая часть работы.
Таким образом, АТФ — это главный универсальный поставщик энергии в клетках всех живых организмов.
АТФ чрезвычайно быстро обновляется. У человека, например, каждая молекула АТФ расщепляется и вновь восстанавливается 2 400 раз в сутки, так что ее средняя продолжительность жизни менее 1 мин. Синтез АТФ осуществляется главным образом в митохондриях и хлоропластах (частично в цитоплазме). Образовавшаяся здесь АТФ направляется в те участки клетки, где возникает потребность в энергии.
Источник : Н.А. Лемеза Л.В.Камлюк Н.Д. Лисов «Пособие по биологии для поступающих в ВУЗы»
Креатинкиназа общая
Креатинкиназа – фермент, который стимулирует превращение креатина в креатинфосфат и обеспечивает энергией мышечное сокращение.
Синонимы русские
КК, креатинфосфокиназа (КФК).
Синонимы английские
Creatine Kinase (CK), Creatine Phosphokinase (CPK).
Метод исследования
УФ кинетический тест.
Единицы измерения
Ед/л (единица на литр).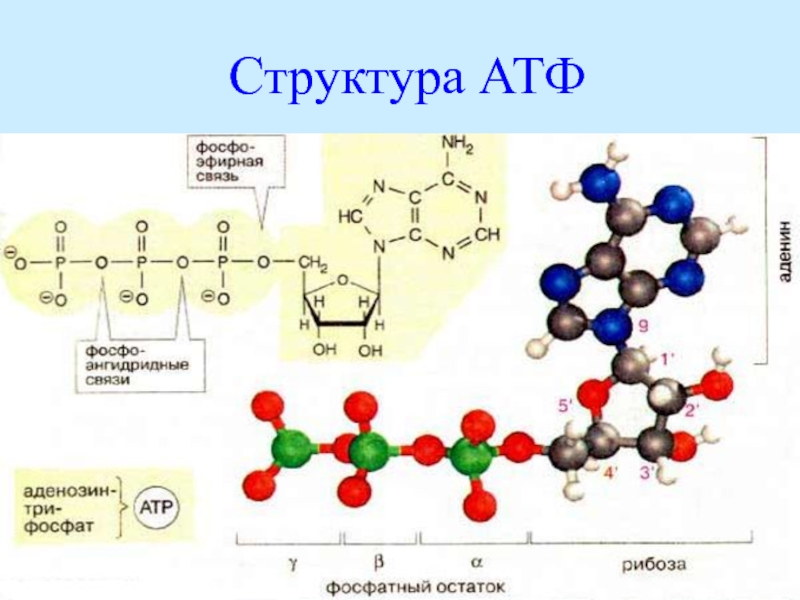
Какой биоматериал можно использовать для исследования?
Венозную кровь.
Как правильно подготовиться к исследованию?
- Не принимать пищу в течение 12 часов перед исследованием.
- Исключить физическое и эмоциональное перенапряжение в течение 30 минут до исследования.
- Не курить в течение 30 минут до исследования.
Общая информация об исследовании
Креатинкиназа – это фермент, который катализирует реакцию переноса фосфорильного остатка с АТФ на креатин с образованием креатинфосфата и АДФ. АТФ (аденозинтрифосфат) – молекула, являющаяся источником энергии в биохимических реакциях человеческого организма.
Реакция, катализируемая креатинкиназой, обеспечивает энергией мышечные сокращения. Различают креатинкиназу, содержащуюся в митохондриях и цитоплазме клеток.
Молекула креатинкиназы состоит из двух частей, которые могут быть представлены одной из двух субъединиц: М, от английского muscle – «мышца», и B, brain – «мозг». Таким образом, в организме человека креатинкиназа есть в виде трех изомеров: ММ, МВ, ВВ. ММ-изомер содержится в скелетной мускулатуре и миокарде, МВ – в основном в миокарде, ВВ – в тканях головного мозга, в небольшом количестве в любых клетках организма.
Таким образом, в организме человека креатинкиназа есть в виде трех изомеров: ММ, МВ, ВВ. ММ-изомер содержится в скелетной мускулатуре и миокарде, МВ – в основном в миокарде, ВВ – в тканях головного мозга, в небольшом количестве в любых клетках организма.
В крови здорового человека креатинкиназа присутствует в небольших количествах, в основном в виде ММ-изомера. Активность креатинкиназы зависит от возраста, пола, расы, мышечной массы и физической активности.
Поступление креатинкиназы в кровоток в больших количествах происходит при повреждении содержащих ее клеток. При этом по повышению активности определенных изомеров можно сделать вывод о том, какая ткань поражена: ММ-фракция – повреждение мышц и в меньшей степени поражение сердца, МВ-фракция – повреждение миокарда, ВВ-фракция – онкологические заболевания. Обычно делают анализы на общую креатинкиназу и ее МВ-фракции.
Таким образом, повышение креатинкиназы в крови позволяет сделать вывод об опухолевом процессе, поражении сердца или мышц, которое в свою очередь может развиться как при первичном повреждении данных органов (при ишемии, воспалении, травмах, дистрофических процессах), так и вследствие их поражения при других состояниях (из-за отравления, метаболических нарушений, интоксикаций).
Сердечные заболевания, при которых разрушаются клетки, – это инфаркт миокарда, миокардиты, миокардиодистрофии, токсическое поражение миокарда. Анализ на креатинкиназу имеет наибольшее значение для диагностики инфаркта миокарда, так как активность этого фермента повышается раньше других, уже через 2-4 часа после инфаркта, и достигает максимума через 1-2 суток, затем нормализуется. Чем раньше начато лечение инфаркта, тем лучше для пациента, поэтому так важна своевременная и точная диагностика.
Заболевания мышц, при которых разрушаются клетки, – это миозиты, миодистрофии, травмы, особенно при сдавливании, пролежни, опухоли, интенсивная работа мышц, в том числе происходящая при судорогах. Кроме того, отмечена обратная зависимость уровня гормонов щитовидной железы и креатинкиназы: при снижении T3 и T4 активность креатинкиназы повышается и наоборот.
Интересно, что впервые анализ на креатинкиназу был использован для выявления миопатии, однако в настоящее время его используют главным образом для диагностики инфаркта миокарда.
Для чего используется исследование?
- Для подтверждения диагноза «инфаркт миокарда», «миокардит», «миокардиодистрофия».
- Для подтверждения диагноза «полимиозит», «дерматомиозит», «миодистрофия».
- Чтобы проверить наличие заболеваний щитовидной железы.
- Чтобы убедиться в наличии опухолевого процесса и оценить его тяжесть.
- Чтобы оценить тяжесть течения полимиозита, дерматомиозита, миодистрофии, миопатии.
- Чтобы выявить носительство гена миопатии Дюшенна.
- Для диагностики и оценки тяжести поражения сердца и мышечной системы при интоксикации из-за инфекции, а также при отравлениях (угарным газом, ядом змеи, лекарственными средствами).
Когда назначается исследование?
- При симптомах ишемической болезни сердца.
- При симптомах инфаркта миокарда, в частности при стертой клинической картине, особенно при повторном инфаркте, атипичной локализации, болевом синдроме или ЭКГ-признаках, затруднении дифференциальной диагностики с другими формами ишемической болезни сердца.

- При гипотиреозе.
- При симптомах миозита, миодистрофии, миопатии.
- При планировании беременности женщиной, в семье которой были больные миопатией Дюшенна.
- При заболеваниях, которые могут привести к поражению сердца или мышечной системы.
Что означают результаты?
Референсные значения
|
Возраст, пол |
Референсные значения |
|
|
2 — 5 дней |
||
|
5 дней — 6 мес. |
||
|
6 — 12 мес. |
||
|
1 — 3 года |
||
|
3 — 6 лет |
||
|
6 — 12 лет |
женский |
|
|
мужской |
||
|
12 — 17 лет |
женский |
|
|
мужской |
||
|
> 17 лет |
женский |
|
|
мужской |
||
Результаты анализа говорят о наличии или отсутствии поражения миокарда, скелетной мускулатуры, опухолевого процесса, заболеваний щитовидной железы. Верная трактовка полученных показателей позволяет сделать вывод о форме поражения и степени его тяжести.
Верная трактовка полученных показателей позволяет сделать вывод о форме поражения и степени его тяжести.
Причины повышения активности креатинкиназы общей:
- инфаркт миокарда,
- миокардиты,
- миокардиодистрофии,
- полимиозит,
- дерматомиозит,
- мышечные дистрофии,
- травмы, ожоги,
- гипотиреоз,
- опухолевый процесс в организме,
- распад опухоли,
- прием дексаметазона, статинов, фибратов, амфотерицина В, обезболивающих, алкоголя, кокаина,
- интенсивная физическая нагрузка,
- судороги, эпилептический статус,
- оперативные вмешательства.
Причины понижения активности креатинкиназы общей:
- снижение мышечной массы,
- алкогольное поражение печени,
- коллагенозы (например, ревматоидный артрит),
- гипертиреоз,
- прием аскорбиновой кислоты, амикацина, аспирина,
- беременность.
Что может влиять на результат?
- Необходимо сообщать врачу точную информацию о принимаемых лекарствах, а также об имеющихся хронических заболеваниях.
- Оперативные вмешательства и в некоторых случаях внутривенные инъекции повышают активность креатинкиназы.
Важные замечания
Повышение активности креатинкиназы общей не является прямым указанием на какое-либо заболевание, так что оно должно трактоваться специалистом с учетом клинической картины и результатов дополнительного обследования.
Также рекомендуется
Кто назначает исследование?
Кардиолог, терапевт, невролог, педиатр, онколог, эндокринолог.
Литература
- Пархоменко А. Н., Иркин О. И., Лутай Я. М. – Роль биологических маркеров в неотложной кардиологии. – Отдел реанимации и интенсивной терапии, Национальный научно-исследовательский центр «Институт кардиологии им.
 акад. Н.Д. Стражеско», Киев.
акад. Н.Д. Стражеско», Киев. - B. Galarraga, D. Sinclair 1 , M. N. Fahie, F. C. McCrae, R. G. Hull and J. M. Ledingham. – A rare but important cause for a raised serum creatine kinase concentration: two case reports and a literature review.
- Ana L Huerta-Alardín, Joseph Varon and Paul E Marik. – Bench-to-bedside review: Rhabdomyolysis – an overview for clinicians.
- Archana Prakash, A. K. Lal, K. S. Negi. – Serum Creatine Kinase Activity in Thyroid Disorders.
- SourceClinical Methods: The History, Physical, and Laboratory Examinations. 3rd edition. Boston: Butterworths; 1990. Chapter 32.
Что такое митохондрия? — Мэлсмон
Клетка – это сложная система, живущая по своим законам и правилам. Организация клетки сложна и многогранна: для правильного функционирования органеллы – составляющие клетки – должны находиться в гармонии, при этом каждая должна выполнять свою уникальную функцию своевременно и полноценно. И только одна структура – митохондрия – имеет целый ряд специфических задач.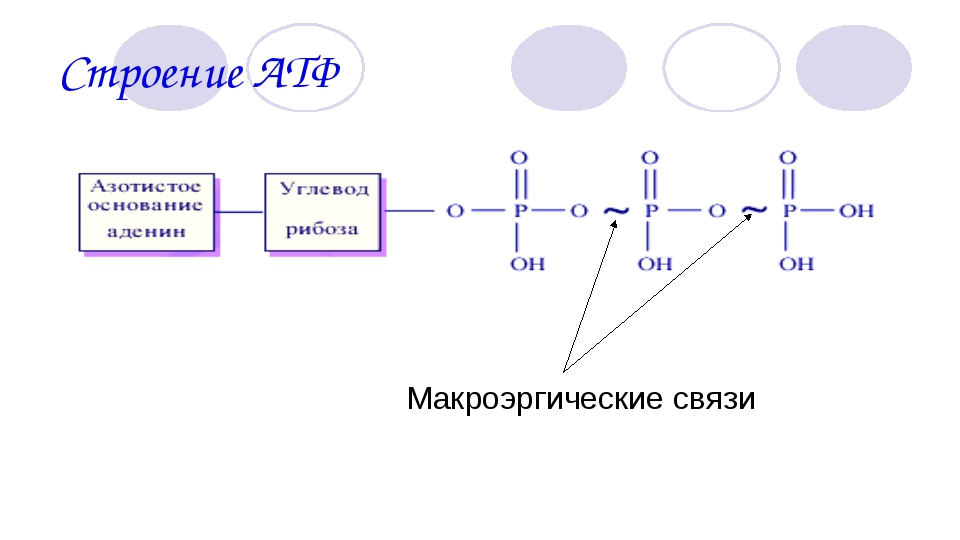 Многофункциональность и уникальность митохондрии стала причиной возникновения теории ее бактериального происхождения.
Многофункциональность и уникальность митохондрии стала причиной возникновения теории ее бактериального происхождения.
Митохондрия, о существовании которой узнали в середине XIX века, на протяжении практически 150 лет считалась исключительно энергетической станцией клетки. Действительно, основной функцией митохондрии является преобразование питательных веществ в полезную энергию с использованием молекул кислорода – синтез молекул АТФ. Этот процесс генерирования энергии внутри клетки известен как клеточное дыхание. Организм использует энергию АТФ, расходуя ее на проведение нервного сигнала, мышечное сокращение, образование тепла, синтез нужных клеточных компонентов, уничтожение ненужных веществ и пр.
Около 600 млн лет назад в клетку так называемых гетеротрофов внедряется бактерия, которая умеет утилизировать кислород. Есть точка зрения, что появление внутри клетки нового типа бактерий было вызвано постоянным увеличением в атмосфере Земли кислорода, начавшим поступать из мирового океана в атмосферу около 2,4 млрд лет назад.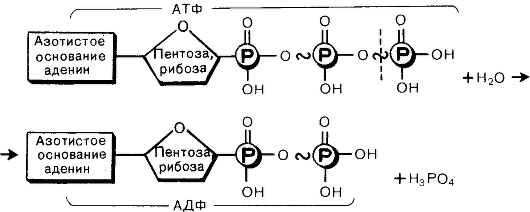 Высокая окислительная способность кислорода представляла опасность для внутриклеточных органических и неорганических элементов, и появляются бактерии, уничтожающие кислород в присутствии ионов водорода с образованием воды. Таким образом внутри клетки содержание кислорода уменьшается, а с ним и уменьшается вероятность нежелательного окисления клеточных компонентов, что, наверно, полезно для клетки.
Высокая окислительная способность кислорода представляла опасность для внутриклеточных органических и неорганических элементов, и появляются бактерии, уничтожающие кислород в присутствии ионов водорода с образованием воды. Таким образом внутри клетки содержание кислорода уменьшается, а с ним и уменьшается вероятность нежелательного окисления клеточных компонентов, что, наверно, полезно для клетки.
Хотя митохондрия была первоначально идентифицирована как ключевая органелла, по существу необходимая для выработки энергии и окислительного метаболизма, существует немало свидетельств того, что митохондрии тесно вовлечены в регуляцию жизненно важных клеточных процессов, таких как запрограммированная гибель и рост клеток. Открытие митохондриальных «формирующих белков» (Dynamin-related protein (Drp), mitofusins (Mfn) и т.д.) показало, что митохондрии являются высокодинамичными органеллами, постоянно меняющими морфологию в результате процессов деления и слияния.
Еще одной уникальной особенностью митохондрии является наличие собственной ДНК (митохондриальная ДНК), причем наследуется она только от матери.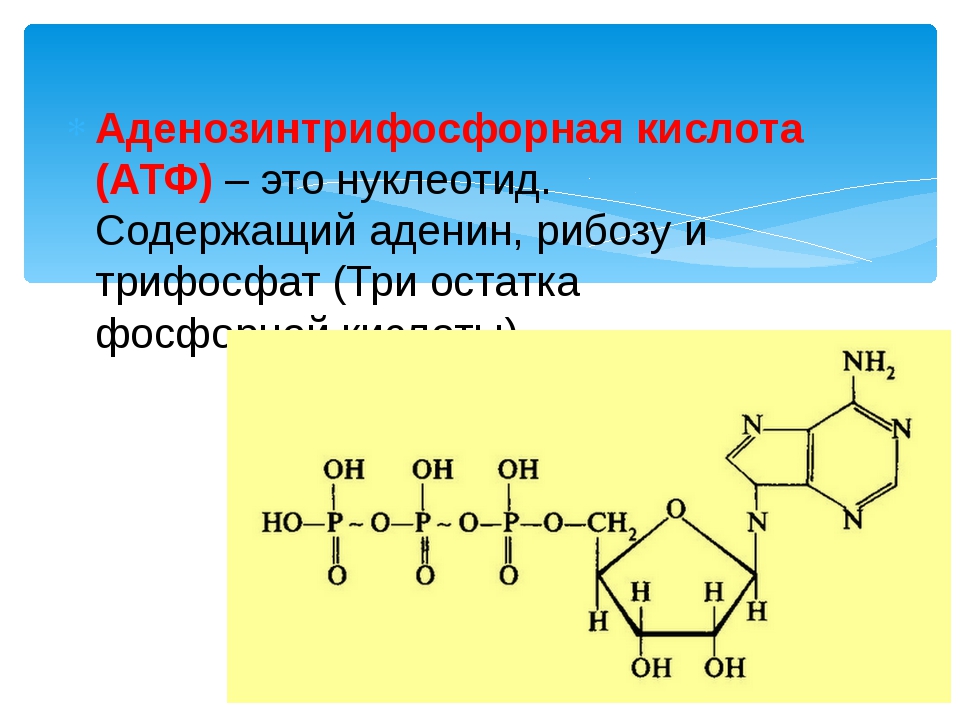
В зависимости от энергетической потребности каждой клетки, содержание в ней митохондрий может варьироваться от одной-двух до нескольких тысяч штук. Как известно, самыми энергопотребляемыми клетками являются клетки мышц, сердца, печени, почек и мозга. Соответственно, содержание в них митохондрий является максимальным.
На основе статьи: Митохондрии помнят, что они были бактериями. Журнал «Коммерсантъ Наука» №5 от 28.07.2017, стр. 25. Дмитрий Зоров, доктор биологических наук, МГУ им. М.В. Ломоносова
Что влияет на функционирование митохондрии?С возрастом функции митохондрий прогрессивно ухудшаются. Митохондрии теряют свою дыхательную активность, накапливаются повреждения в их митохондриальной ДНК, и образуется чрезмерное количество активных форм кислорода (свободные радикалы). Между 30 и 70 годами, в среднем, функция митохондрий ухудшается на 25-30%.
Что же является причиной нарушений функционирования митохондрий? Ответ, как всегда, лежит на поверхности: митохондриальная дисфункция является причиной самой жизни.
Наследственность, солнечная активность, загрязненный воздух больших городов, неправильное и несбалансированное питание, вредные привычки, гиподинамия, стресс – все это в совокупности и по-отдельности приводит к тому, что страдают самые маленькие, но при это столь важные, составляющие нашего тела – митохондрии.
Безусловно, старение неизбежно. Но наш биологический возраст может сильно отличаться от хронологического, причем митохондрии имеют много общего с биологическим старением. По данным недавних исследований, темпы биологического старения могут сильно отличаться у разных людей. Исследователи измерили более десятка различных биомаркеров, таких как длина теломера, повреждение ДНК, холестерин ЛПНП, метаболизм глюкозы и чувствительность к инсулину, в трех точках жизни людей: в возрасте 22, 32 и 38 лет.
«Мы обнаружили, что кто-то в возрасте 38 лет биологически мог выглядеть на 10 лет моложе или старше, судя по биологическим маркерам. Несмотря на одинаковый возраст, биологическое старение происходит совершенно разными темпами.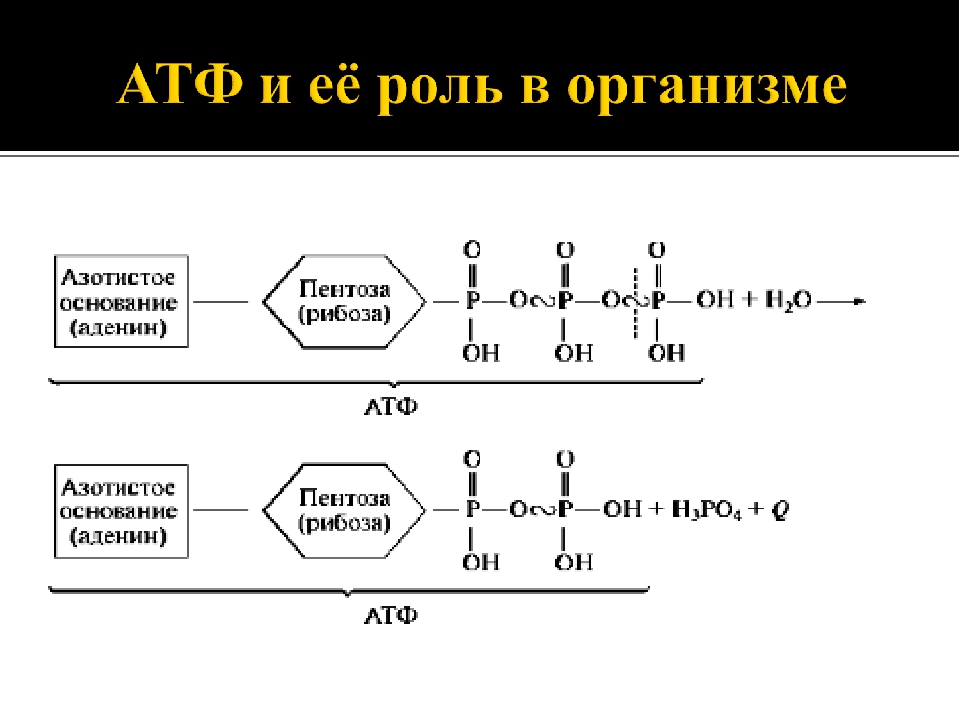 Интересно, что когда этих людей сфотографировали и показали их фотографии прохожим с просьбой угадать хронологический возраст изображенных людей, то люди угадывали биологический, а не хронологической возраст».
Интересно, что когда этих людей сфотографировали и показали их фотографии прохожим с просьбой угадать хронологический возраст изображенных людей, то люди угадывали биологический, а не хронологической возраст».
Таким образом, возраст, на который нас воспринимают окружающие, напрямую соответствует биологическим биомаркерам, которые в значительной степени обусловлены здоровьем митохондрий. Поэтому, хотя старения и не избежать, поддержание собственных митохондрий в здоровом состоянии позволяет в значительной степени управлять процессом старения.
Насколько важна клеточная энергия и функция митохондрий?
Низкий уровень энергии = Возможные генетические ошибки (Болезнь клетки)
Нормальный уровень энергии = Правильный синтез генов и белков (Здоровая клетка)
Митохондриальная дисфункция вовлечена практически во все заболеванияВ последний десятилетия во всем мире активно изучается вопрос связи митохондриальной активности и возрастассоциированных заболеваний. Как оказалось, митохондрии оказывают непосредственное влияние на работу всего организма в целом:
Как оказалось, митохондрии оказывают непосредственное влияние на работу всего организма в целом:
- Нейродегенартивные заболевания
- Исследователи клиники Майо, использующие генетические мышиные модели, обнаружили, что митохондрии в клетках головного мозга перестают нормально функционировать уже на ранних стадиях заболевания Альцгеймера (1). Помимо этого, растущее количество исследований подтверждает связь между болезнью Альцгеймера и диабетом II типа. Пациенты с сахарным диабетом II типа имеют значительно повышенный риск развития болезни Альцгеймера и наоборот (2). Недавние результаты генетических исследований показывают, что дефектный контроль качества митохондрий может играть важную роль в развитии болезни Паркинсона (10).
- Развитие онкологических заболеваний
- Митохондрии играют важную роль в питании клеток, генерируя АТФ. Они также являются основным источником генерирования активных форм кислорода (АФК), которые играют регулирующую роль в процессе жизни и естественной гибели клеток.
 Мутации в митохондриальной ДНК и нарушение регуляции митохондриального метаболизма часто описываются в опухолях человека. Хотя роль окислительного стресса как следствия мутаций митохондриальной ДНК была доказана достаточно давно, причинная роль митохондрий в прогрессировании опухоли была продемонстрирована только недавно. (3)
Мутации в митохондриальной ДНК и нарушение регуляции митохондриального метаболизма часто описываются в опухолях человека. Хотя роль окислительного стресса как следствия мутаций митохондриальной ДНК была доказана достаточно давно, причинная роль митохондрий в прогрессировании опухоли была продемонстрирована только недавно. (3)
- Митохондрии играют важную роль в питании клеток, генерируя АТФ. Они также являются основным источником генерирования активных форм кислорода (АФК), которые играют регулирующую роль в процессе жизни и естественной гибели клеток.
- Синдром хронической усталости
- Синдром хронической усталости (или миалгический энцефаломиелит) является физиологическим состоянием, при котором пациент чувствует высокий уровень усталости без очевидной причины. По международным данным это состояние характерно для одного из 400 людей. Многочисленные исследования показывают, что у пациентов, страдающих синдромом хронической усталости, наблюдается очевидная митохондриальная недостаточность (4, 5, 6, 7)
- Диабет
- Исследования предоставили доказательства митохондриальной дисфункции в скелетных мышцах пациентов с диабетом II и преддиабетического типа, главным образом из-за более низкого содержания митохондрий (митохондриальный биогенез) (2, 8).

- Исследования предоставили доказательства митохондриальной дисфункции в скелетных мышцах пациентов с диабетом II и преддиабетического типа, главным образом из-за более низкого содержания митохондрий (митохондриальный биогенез) (2, 8).
- Заболевания сердца и сосудистой системы
- Сердечная недостаточность является одной из главных проблем общественного здравоохранения, и в настоящее время нет профилактического лечения. Существующая терапия обеспечивает симптоматическое облегчение, но не может обратить вспять молекулярные изменения, которые происходят в кардиомиоцитах. Механизмы сердечной недостаточности являются сложными и множественными, но одним из критических факторов развития этого заболевания является митохондриальная дисфункция. (9)
- Mitochondrial dysfunction is a trigger of Alzheimer’s disease pathophysiology. Paula I. Moreira, Cristina Carvalho, Xiongwei Zhu, Mark A. Smith, George Perry; Volume 1802, Issue 1, January 2010, Pages 2-10
- Inflammation, defective insulin signaling, and mitochondrial dysfunction as common molecular denominators connecting type 2 diabetes to Alzheimer disease. De Felice FG, Ferreira ST; Diabetes. 2014 Jul;63(7):2262-72.
 doi: 10.2337/db13-1954. Epub 2014 Jun 15.
doi: 10.2337/db13-1954. Epub 2014 Jun 15. - Mitochondrial dysfunction and cancer metastasis. Chen EI. J Bioenerg Biomembr. 2012 Dec;44(6):619-22. doi: 10.1007/s10863-012-9465-9.
- Mitochondrial dysfunction and the pathophysiology of Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS). Booth NE, Myhill S, McLaren-Howard J. Int J Clin Exp Med. 2012;5(3):208-20. Epub 2012 Jun 15.
- Targeting mitochondrial dysfunction in the treatment of Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS) — a clinical audit. Myhill S, Booth NE, McLaren-Howard J. Int J Clin Exp Med. 2013;6(1):1-15. Epub 2012 Nov 20.
- Patients with chronic fatigue syndrome performed worse than controls in a controlled repeated exercise study despite a normal oxidative phosphorylation capacity. Vermeulen RC, Kurk RM, Visser FC, Sluiter W, Scholte HR. J Transl Med. 2010 Oct 11;8:93. doi: 10.1186/1479-5876-8-93.
- Chronic fatigue syndrome and mitochondrial dysfunction.
 Sarah Myhill, Norman E. Booth, John McLaren-Howard. Int J Clin Exp Med. 2009; 2(1): 1–16. Published online 2009 Jan 15.
Sarah Myhill, Norman E. Booth, John McLaren-Howard. Int J Clin Exp Med. 2009; 2(1): 1–16. Published online 2009 Jan 15. - Mitochondrial dysfunction in type 2 diabetes and obesity. Højlund K, Mogensen M, Sahlin K, Beck-Nielsen H. Endocrinol Metab Clin North Am. 2008 Sep;37(3):713-31, x. doi: 10.1016/j.ecl.2008.06.006.
- Mitochondria as a therapeutic target in heart failure. Bayeva M, Gheorghiade M, Ardehali H. J Am Coll Cardiol. 2013 Feb 12;61(6):599-610. doi: 10.1016/j.jacc.2012.08.1021. Epub 2012 Dec 5.
- Mitochondrial quality control and Parkinson’s disease: a pathway unfolds. de Castro IP, Martins LM, Loh SH. Mol Neurobiol. 2011 Apr;43(2):80-6. doi: 10.1007/s12035-010-8150-4. Epub 2010 Dec 1.
Учитывая то, что здоровье митохондрии в конечном счете сказывается на здоровье всего организма в целом, первые признаки нарушения работы митохондрий могут быть весьма разнообразны.
Общее состояние
Дефицит энергии в первую очередь отражается на нашем общем самочувствии. Это быстрая утомляемость – нам чаще нужна передышка при выполнении привычных дел. Непроходящее чувство слабости даже после выпитого кофе. Повышенная тревога и раздражительность, нарушенный сон. Последствия всего этого отражаются на нашем внешнем виде. Кожа становится уставшей, тусклой и истончённая. Лицо приобретает сероватый цвет, видны круги под глазами, появляются раздражения кожи. Постепенно ухудшается здоровье, мы чаще начинаем болеть, особенно в хронической форме. Всё заметнее признаки старения – тело стареет быстрее.
Здоровый образ жизни
Синдром энергодефицита отражается на нашей физической активности. Посещения спортивного зала становится сопряжено с сильным переутомлением. Подходы к снарядам занимают больше времени и требуют больше усилий. Восстановление энергии происходит медленно, порой не ощущается вовсе.
Клеточный энергодефицит также приводит к низкой усвояемости питательных веществ. Это драматически влияет на эффективность выбранной диеты. Неспособность эффективно усваивать необходимые химические элементы приводит к сложным последствиям для нашего организма.
Это драматически влияет на эффективность выбранной диеты. Неспособность эффективно усваивать необходимые химические элементы приводит к сложным последствиям для нашего организма.
Интимная жизнь
Клеточный энергодефицит приводит к снижению либидо – ослабевает сексуальное влечение. Мы уделяем меньше внимания партнёру, получаем меньше удовольствия от интимной близости. У женщин, хронический энергодефицит способствует преждевременному наступлению менопаузы и климакса с соответствующим симптомокомплексом.
Поскольку митохондриальная функция лежит в основе всего, что происходит в организме, то оптимизация митохондриальной функции, и предотвращение нарушения функции митохондрий путем получения всех необходимых питательных веществ и прекурсоров, необходимых митохондриям, чрезвычайно важна для здоровья и профилактики заболеваний.
Мэлсмон – метаболическая таргетированная терапия (МТТ)Более 50 лет назад большинство научных и медицинских терапевтических подходов были сосредоточены на клеточном метаболизме. С появлением генетики на терапевтической стадии доминировал сдвиг в сторону геномики, а затем и протеомики (белковых профилей).
С появлением генетики на терапевтической стадии доминировал сдвиг в сторону геномики, а затем и протеомики (белковых профилей).
Подход к применению продуктов или соединений, которые увеличивают или поддерживают митохондриальную функцию, тем самым борясь с метаболической дисфункцией, получил название метаболическая таргетированная терапия (МТТ).
Плацентарный препарат Мэлсмон – гордость японской фармацевтики. Он эффективно борется с митохондриальной дисфункцией и восстанавливает клеточный запас энергии. Разработанный ещё в 50-х годах прошлого столетия он включён в национальную программу обязательного медицинского обслуживания Японии и используется в составе множества современных протоколов лечения. Сегодня Мэлсмон активно применяется в программах долголетия, где помогает сохранить здоровье, молодость и красоту.
Мэлсмон – плацентарный препарат системного действия, направленного на нормализацию работы митохондрии. Восстановление энергетического запаса клетки позволяет эффективно бороться со всеми признаками синдрома клеточного энергодефицита.
Достоверно установлено, что Мэлсмон положительно влияет на общее самочувствие:
- снижает утомляемость, придаёт сил, повышает работоспособность
- стабилизирует эмоциональное состояние, понижая уровень тревоги и раздражительности
- улучшает качество сна.
Безусловно, это положительно отражается и на внешнем виде: лицо приобретает здоровый цвет, исчезают признаки усталости, снижается воспаление, повышается упругость, кожа «сияет» изнутри и выглядит моложе.
К уникальным особенностям препарата Мэлсмон относятся:
- Безопасность класса А. Проверка биоматериала осуществляется тремя независимыми лабораториями.
- Высокий уровень очистки. Не содержит гормонов, витаминов и факторов роста.
- 100% биодоступность. Препарат мгновенно включаются в метаболические процессы.
Безопасность Мэлсмон
Отсутствие в препарате биогенных стимуляторов обуславливает его безопасность, возможность длительного применения, возможность применения у пациентов с сопутствующей патологией.
Мэлсмон не вступает в лекарственное взаимодействие
Может назначаться в сочетании с другими препаратами, в том числе в сочетании с менопаузальной гормональной терапией. По результатам многолетней практики применения, МЭЛСМОН не дает побочных эффектов на фоне своей высокой эффективности.
Факты
Доказанная повышение митохондриальной активности
Динамика митохондриальной активности лимфоцитов при инкубировании крови с препаратом Мэлсмон. Измайлова Т.Д., Персонализированные протоколы метаболической коррекции как основа anti-age программ, Инъекционные методы в косметологии, №1, 2016, стр. 24-37
Оригинальность лекарственного препарата Мэлсмон
Комапния Melsmon Pharmaceutical Co., Ltd гарантирует безопасность и качество только оригинального лекарственного препарата МЭЛСМОН.
Использование поддельного или контрафактного препарата гидролизата плаценты человека для подкожного применения опасно для здоровья и жизни человека. Влечёт за собой риски: отсутствие ожидаемых эффектов терапии, непредсказуемые побочные эффекты, риск инфицирования вирусами.
Убедитесь в подлинности
Для России препарат МЭЛСМОН производится в оригинальной упаковке с защитной пломбой. Информация на упаковке и ампулах, инструкция – только на русском языке, без иероглифов. Упаковки по 10 и 50 ампул.
Каждая ампула имеет отрывной стикер с информацией о серии и сроке годности препарата.
Этапы энергетического обмена – Российский учебник
Живой организм построен из клеток. А некоторые, например, амеба или инфузория, состоят из одной клетки. Сложные многоклеточные особи представлены сочетаниями клеток, которые образуют различные системы организма: дыхательную или пищеварительную. Весь организм пронизан клетками нервной ткани, которые обеспечивают регуляцию и управление макроорганизмом.
Живая клетка от неживой отличается тем, что в ней постоянно и непрерывно протекает два разнонаправленных процесса:
- синтез, или строительство новых органелл (пластический обмен или ассимиляция)
- катаболизм, или разрушение питательных элементов с образованием энергии (энергетический обмен или диссимиляция)
В живых особях баланс между ассимиляцией и диссимиляцией поддерживается не всегда. Если наблюдать за жизнью организма, можно заметить, что сначала тело становится больше по размерам, крепче и выносливее. Чем старше становится организм, тем меньше заметен рост, а в старости начинают преобладать процессы распада, организм не успевает восстанавливаться и погибает.
Чтобы машина ехала, ей нужен бензин. А бензин делают из нефти. Чтобы макроорганизм существовал, ему нужна энергия. В учебниках биологии часто можно встретить фразу глюкоза — ресурс энергии для клетки. Это так. Но глюкоза — как нефть для автомобиля. Поэтому сначала глюкозу нужно превратить в бензин. И таким бензином для клетки будет молекула АТФ.
Молекула АТФ известна миру довольно давно. Через десять лет, в 2029 году отпразднует сто лет со дня открытия вещество, благодаря которому в живой клетке поддерживается упорядоченность структур и она может противостоять хаосу и растущей энтропии (почему так происходит, подробно рассказано на странице 30 учебника «Естествознание 11 класс» под редакцией Титова С.А.).
В 1929 году, практически одновременно, сотрудники Института биологии кайзера Вильгельма в Берлине Карл Ломан и Отто Мейергоф и ученые Гарвардской Медицинской школы Сайрус Фиске и Йеллапрагад Суббарао в Гарвардской Медицинской школе опубликовали работы, в которых описали молекулу АТФ.
В 30 годах ХХ века в лаборатории Мейергофа появился молодой ученый, Фриц Липман, который заинтересовался энергетическими аспектами клеточного метаболизма, и в 1941 году талантливый биохимик доказал, что АТФ – основной двигатель энергетических реакций в живой клетке. А в 1953 году его вклад в физиологию совместно с Х. Кребсом был удостоен Нобелевской премии.
АТФ — аденозинтрифосфат, нуклеотид, глобальный ресурс энергии для сложных реакций обмена веществ, происходящих в клетках организма. Схематично молекула АТФ представлена на странице 99 учебника «Естествознание 11 класс» под редакцией Титова С.А..
Вся суть энергетического обмена сводится к решению задачи, как энергию из сложных молекул пищи превратить в молекулу АТФ. В процессе эволюции эта задача была решена.
Так как же булочка из Макдональдса превращается в энергию макроэргических связей АТФ?
В энергетическом обмене выделяют несколько процессов, разделенных не только временем, но и протекающих в различных частях клетки:
- Подготовительный этап
- Гликолиз
- Кислородное окисление
Естествознание. 10 класс. Учебник
Учебник отличаются качественным современным оформлением, в нём приводятся многочисленные слайды и микрофотографии. Выполняя проблемные, поисковые и исследовательские задания, школьники не только активно усваивают материал, но и учатся мыслить, искать и анализировать информацию из разных источников, в том числе из интернета. Особое внимание уделяется практическим заданиям: ученикам предлагается проводить опыты, конструировать модели, разрабатывать проекты.
КупитьПодготовительный этап у человека и других многоклеточных макроорганизмов начинается в ротовой полости, продолжается в желудочно-кишечном тракте и представляет собой многоступенчатый процесс распада сложных полимеров белков, жиров и углеводов пищи до мономеров.
Из курса химии помним, что во время разрыва связей элементов выделяется энергия. Для образования аденозинтрифосфата этой энергии недостаточно, и она вся уходит во внешнюю среду.
Что же происходит у простых одноклеточных организмов, у которых нет ротиков и животиков? Пища, захваченная одноклеточным организмом, попадает в пищеварительную вакуоль или лизосому, где при помощи ферментов-катализаторов, способствующих пищеварению, протекает начальный этап диссимиляции.
Подготовленные во время предварительного этапа вещества переходят во второй бескислородный этап энергетического обмена, который называется гликолиз.
Два греческих слова (гликос – «сладкий» и лизис – «расщеплять») дали название второй бескислородной фазе энергетического обмена — глико́лизу.
Гликолиз является цепочкой из 10 биохимических превращений, итогом которых является трансформация стабильной молекулы глюкозы в трехуглеродные молекулы пирувата, – или пировиноградной кислоты.
Что ещё почитать?
«Подождите, – могут сказать дотошные ученики, – глюкоза у нас находится в пищеварительной системе. А процессы метаболизма идут в клетках по всему организму, например, на кончике носа или задней лапе. Как же попадает глюкоза в цитоплазму клетки во всем организме?».Глюкоза всасывается в желудочно-кишечном тракте, попадает в кровоток, током крови разносится ко всем тканям и клеткам организма, и благодаря белку-переносчику инсулину попадает в клетку.
Цитоплазма отдельной клетки – место осуществления реакций гликолиза. Энергии при этом выделяется совсем немного. Ее хватает лишь на формирование 2-х молекул АТФ. Казалось бы, энергия получена, и процесс может остановиться. Так и происходит у некоторых бактерий. Но никакому нормальному многоклеточному организму таких запасов АТФ не хватит. В пировиноградной кислоте остался еще достаточный запас энергии, которую тоже хотелось бы использовать макроорганизму.
Естествознание. 11 класс. Учебник
Учебник отличаются качественным современным оформлением, в нём приводятся многочисленные слайды и микрофотографии. Выполняя проблемные, поисковые и исследовательские задания, школьники не только активно усваивают материал, но и учатся мыслить, искать и анализировать информацию из разных источников, в том числе из интернета. Особое внимание уделяется практическим заданиям: ученикам предлагается проводить опыты, конструировать модели, разрабатывать проекты.
КупитьУ многоклеточных тел пируват переходит в третью фазу диссимиляции — клеточное дыхание в митохондриях. Дыханием процесс называется, поскольку в ходе химических реакций в митохондриях происходит потребление кислорода и выделение углекислого газа в цитоплазму клетки, а дальше, с помощью кровообращения и дыхания, – во внешнюю среду.
Клеточное дыхание представлено двумя этапами:
- цикл Кребса, протекающий в матриксе митохондрий
- окислительное фосфорилирование, протекающее на кристах митохондрий при участии ферментов дыхательной цепи
Итогом кислородного этапа энергетического обмена является выделение количества энергии, достаточного для образования 36 молекул АТФ, воды и СО2. При этом нужно помнить, что аденозинтрифосфат содержит три остатка фосфата, а макроэргических связей образуется только две. Суммарное уравнение биохимических реакций, протекающих в третьей фазе диссимиляции, можно записать так:
2C3H4O3+6O2+36H3PO4+36АДФ=6CO2+42H2O+36АТФ
В итоге этих реакций происходит накопление огромного количества энергии — 36 молекул аденозинтрифосфата против 2-х, что запасаются в процессе гликолиза. Однако поскольку эта фаза требует кислорода для своих реакций, в бескислородной среде процесс протекать не может.
При дефиците кислорода пируват окисляется до лактата. Именно ему принадлежит ощущение приятной боли после хорошей тренировки. У хорошо тренированных людей с активным кровоснабжением и хорошо развитой сетью капилляров нужно затратить большую физическую нагрузку перед тем, как начнет накапливаться молочная кислота.
Вспомним, что еще 2 молекулы аденозинтрифосфата накапливаются на этапе гликолиза. Таким образом, при распаде одной молекулы глюкозы образуется 38 молекул АТФ.
На портале LECTA В параграфе 22 учебника «Естествознание 11 класс» под редакцией Титова С.А. внимательные ученики найдут ответ на вопрос, почему цианистый калий – любимое средство убийц в детективных романах.
ТестБиология 6 класс что такое АТФ. НУЖНО СРОЧНО ПОМОГИТЕ!!!
конспект на тему Вирус — это бесклеточная жизнь (Кто знает то помогите пожалуйста)
ПОМОГИТЕ ПОЖАЛУЙСТА ПОМОГИТЕ ПОЖАЛУЙСТА
К беспозвоночным не относится * гидра планария ланцетник острица аскарида
Чим клітина грибів відрізняється від клітини тварин, рослин та бактерій? СРОЧНО ДАЮ 25 БАЛОВ
Найдите в тексте предложения с биологическими ошибками, укажите номера этих предложений: 1.Насекомые относятся к типу Членистоногие. 2. Наиболее часто … встречающиеся насекомые это комары, мухи, пауки, бабочки. 3. Тело пауков состоит и головогруди, брюшка и 4х пар конечностей. 4. Пауки-хищные животные, для некоторых пауков характерно наружнее пищеварение. 5.Развитие пауков происходит в несколько стадий: яйцо-личинка-куколка-взрослое насекомое. 6. У пауков часто наблюдается половой диморфизм.
ПРОШУ ПОМОГИТЕ ПОЖАЛУЙСТА ЭТО ОЧЕНЬ СРОЧНО!1. Выбери правильный вариант.2. Ответь на вопрос. 1. Вороной конь —качественное прилагательноеотносительно … е прилагательноепритяжательное прилагательное 2. Имеет ли данное прилагательное краткую форму?ДаНет 1. Круговая порука —качественное прилагательноеотносительное прилагательноепритяжательное прилагательное 2. Имеет ли данное прилагательное краткую форму?ДаНет 1. Сиреневый цветок (цветок сирени) —качественное прилагательноеотносительное прилагательноепритяжательное прилагательное 2. Это прилагательное называет признак по отношению к материалу, месту, действию, отвлеченному понятию?НетДа
В результате мейоза образуются:гаметыклетки кожиклетки древесины дубанервные клетки
Задача 1 : Угол АОС – прямой. Если ОС биссектриса угла АОК то чему равна градусная мера угла АОК.
13. Виберіть із наведеного переліку фукці, притаманні внутрішнім жіночим органам:а) утвореня гормонін;б) утворення статевих клітин; в) орган, в якому … відбуваєтьсязапліднення; г) орган, в якому відбувається розвиток зародка і плоду.Завдання з німи и малюнком
доделайте прошу. куча баллов
Где синтезируется атф в животной клетке. АТФ в биологии – определение и расшифровка (10 класс)
Основным источником энергии для клетки являются питательные вещества: углеводы, жиры и белки, которые окисляются с помощью кислорода. Практически все углеводы, прежде чем достичь клеток организма, благодаря работе желудочно-кишечного тракта и печени превращаются в глюкозу. Наряду с углеводами расщепляются также белки — до аминокислот и липиды — до жирных кислот.В клетке питательные вещества окисляются под действием кислорода и при участии ферментов, контролирующих реакции высвобождения энергии и ее утилизацию.
Почти все окислительные реакции происходят в митохондриях, а высвобождаемая энергия запасается в виде макроэргического соединения — АТФ. В дальнейшем для обеспечения внутриклеточных метаболических процессов энергией используется именно АТФ, а не питательные вещества.
Молекула АТФ содержит: (1) азотистое основание аденин; (2) пентозный углевод рибозу, (3) три остатка фосфорной кислоты. Два последних фосфата соединены друг с другом и с остальной частью молекулы макроэргическими фосфатными связями, обозначенными на формуле АТФ символом ~. При соблюдении характерных для организма физических и химических условий энергия каждой такой связи составляет 12000 калорий на 1 моль АТФ, что во много раз превышает энергию обычной химической связи, поэтому фосфатные связи и называют макроэргическими. Более того, эти связи легко разрушаются, обеспечивая внутриклеточные процессы энергией сразу, как только в этом возникает необходимость.
При высвобождении энергии АТФ отдает фосфатную группу и превращается в аденозиндифосфат. Выделившаяся энергия используется практически для всех клеточных процессов, например в реакциях биосинтеза и при мышечном сокращении.
Схема образования аденозинтрифосфата в клетке, показывающая ключевую роль митохондрий в этом процессе.GI — глюкоза; FA — жирные кислоты; АА — аминокислота.
Восполнение запасов АТФ происходит путем воссоединения АДФ с остатком фосфорной кислоты за счет энергии питательных веществ. Этот процесс повторяется вновь и вновь. АТФ постоянно расходуется и накапливается, поэтому она получила название энергетической валюты клетки. Время оборота АТФ составляет всего несколько минут.
Роль митохондрий в химических реакциях образования АТФ . При попадании внутрь клетки глюкоза под действием ферментов цитоплазмы превращается в пировиноградную кислоту (этот процесс называют гликолизом). Энергия, высвобождаемая в этом процессе, затрачивается на превращение небольшого количества АДФ в АТФ, составляющего менее 5% общих запасов энергии.
На 95% осуществляется в митохондриях. Пировиноградная кислота, жирные кислоты и аминокислоты, образующиеся соответственно из углеводов, жиров и белков, в матриксе митохондрий в итоге превращаются в соединение под названием «ацетил-КоА». Это соединение, в свою очередь, вступает в серию ферментативных реакций под общим названием «цикл трикарбоновых кислот» или «цикл Кребса», чтобы отдать свою энергию.
В цикле трикарбоновых кислот ацетил-КоА расщепляется до атомов водорода и молекул углекислого газа. Углекислый газ удаляется из митохондрий, затем — из клетки путем диффузии и выводится из организма через легкие.
Атомы водорода химически очень активны и поэтому сразу вступают в реакцию с кислородом, диффундирующим в митохондрии. Большое количество энергии, выделяющейся в этой реакции, используется для превращения множества молекул АДФ в АТФ. Эти реакции достаточно сложны и требуют участия огромного числа ферментов, входящих в состав крист митохондрий. На начальном этапе от атома водорода отщепляется электрон, и атом превращается в ион водорода. Процесс заканчивается присоединением ионов водорода к кислороду. В результате этой реакции образуются вода и большое количество энергии, необходимой для работы АТФ-синтетазы — крупного глобулярного белка, выступающего в виде бугорков на поверхности крист митохондрий. Под действием этого фермента, использующего энергию ионов водорода, АДФ превращается в АТФ. Новые молекулы АТФ направляются из митохондрий ко всем отделам клетки, включая ядро, где энергия этого соединения используется для обеспечения самых разных функций.
Данный процесс синтеза АТФ в целом называют хемиосмотическим механизмом образования АТФ.
Использование аденозинтрифосфата митохондрий для реализации трех важных функций клетки:
мембранного транспорта, синтеза белка и мышечного сокращения.
метаболиты гликолиза (1,3-дифосфоглицерат ,фосфоенолпируват ),
метаболиты цикла трикарбоновых кислот (сукцинил-КоА ) и
креатинфосфат .
Основным способом получения АТФ в клетке является окислительное фосфорилирование , протекающее в структурах внутренней мембраны митохондрий. При этом энергия атомов водорода молекул НАДН и ФАДН 2 , образованных в гликолизе, ЦТК, окислении жирных кислот,в ходе окислительно-восстановительных процессов преобразуется в энергию связей АТФ.
Однако также есть другой способ фосфорилирования АДФ до АТФ – субстратное фосфорилирование . Этот способ связан спередачей энергии макроэргической связи какого-либо вещества (субстрата) на АДФ. К таким веществам относятся:
Пируват окисляется до ацетил-КоА.
Пировиноградная кислота (ПК, пируват) является продуктом окисления глюкозы и некоторых аминокислот. Ее судьба различна в зависимости от доступности кислорода в клетке. Ванаэробных условиях она восстанавливается домолочной кислоты . Ваэробных условиях пируват симпортом с ионами Н + , движущимися по протонному градиенту, проникает в митохондрии. Здесь происходит его превращение в ацетил-коэнзим А (ацетил-КоА ) с помощьюпируватдегидрогеназного мульферментного комплекса.
Пируватдегидрогеназный мульферментный комплекс
Суммарное уравнение окисления пировиноградной кислоты
Пируватдегидрогеназный мульферментный комплекс расположен в матриксе митохондрий эукариотов. Состоит у человека из96 субъединиц , организовавнных в три функциональных белка. Гигантское образование, имеет50 нм в диаметре, что впять раз!!! больше, чемрибосома .
Процесс проходит пять последовательных реакций, в которых принмает участие 5 коферментов:
Пируватдегидрогеназа (Е 1 , ПК-дегидрогеназа), коферментом служиттиаминдифосфат (ТДФ), катализирует 1-ю реакцию.
Дигидролипоил трансацетилаза (в русскоязычной литературе встречаются названия —дигидролипоат-ацетилтрансфераза илипоамид редуктаза трансацетилаза (Е 2), кофермент —липоевая кислота , катализирует 2-ю и 3-ю реакции.
Дигидролипоил дегидрогеназа (дигидролипоат-дегидрогеназа) (Е 3), кофермент –ФАД , катализирует 4-ю и 5-ю реакции.
Помимо указанных коферментов, которые прочно связаны с соответствующими ферментами, в работе комплекса принимают участие коэнзим А иНАД .
Суть первых трех реакций сводится к декарбоксилированию пирувата (катализируется пируватдегидрогеназой, Е 1), окислению пирувата до ацетила и переносу ацетила на коэнзим А (катализируетсядигидролипоил трансацетилазой , Е 2).
Реакции синтеза ацетил-sКоА
Оставшиеся 2 реакции необходимы для окисления дигидролипоата обратно в липоат с образованием ФАДН 2 и восстановления НАДН (катализируютсядигидролипоил дегидрогеназой , Е 3).
Реакции образования надн Регуляция пируватдегидрогеназного комплекса
Регулируемым ферментом ПВК-дегидрогеназного комплекса является первый фермент – пируватдегидрогеназа (Е 1). Этому служат два вспомогательных фермента –киназа ифосфатаза, обеспечивая еефосфорилирования идефосфорилирования .
Киназа активируется при избытке конечного продукта биологического окисленияАТФ и продуктов ПВК-дегидрогеназного комплекса –НАДН иацетил-КоА . Активная киназа фосфорилирует пируватдегидрогеназу и инактивирует ее.
Фермент фосфатаза , активируясь ионамикальция или гормономинсулином , дефосфорилирует и активирует пируватдегидрогеназу.
Любой организм может существовать до тех пор, пока происходит поступление питательных веществ из внешней среды и пока продукты его жизнедеятельности выделяются в эту среду. Внутри клетки происходит непрерывный очень сложный комплекс химических превращений, благодаря которым из питательных веществ образуются компоненты тела клетки. Совокупность процессов превращения материи в живом организме, сопровождающихся постоянным ее обновлением, и называется обменом веществ.
Часть общего обмена, которая состоит в поглощении, усвоении питательных веществ и создании за их счет структурных компонентов клетки, называется ассимиляцией — это конструктивный обмен. Вторую часть общего обмена составляют процессы диссимиляции, т.е. процессы разложения и окисления органических веществ, в результате которых клетка получает энергию, — это энергетический обмен. Конструктивный и энергетический обмен составляют единое целое.
В процессе конструктивного обмена клетка из довольно ограниченного числа низкомолекулярных соединений синтезирует биополимеры своего тела. Биосинтетические реакции протекают при участии разнообразных ферментов и требуют затрат энергии.
Живые организмы могут использовать только химически связанную энергию. Каждое вещество обладает определенным запасом потенциальной энергии. Главными материальными носителями ее являются химические связи, разрыв или преобразование которых приводит к освобождению энергии. Энергетический уровень одних связей имеет величину 8-10 кДж — эти связи называются нормальными. В других связях заключена значительно большая энергия — 25-40 кДж — это так называемые макроэргические связи. Почти все известные соединения, обладающие такими связями, имеют в своем составе атомы фосфора или серы, по месту которых в молекуле и локализованы эти связи. Одним из соединений, играющих важнейшую роль в жизнедеятельности клетки, является аденозинтрифосфорная кислота (АТФ).
Аденозинтрифосфорная кислота (АТФ) состоит из органического основания аденина (I), углевода рибозы (II) и трех остатков фосфорной кислоты (III). Соединение аденина и рибозы называется аденозином. Пирофосфатные группы имеют макроэргические связи, обозначенные значком ~. Разложение одной молекулы АТФ с участием воды сопровождается отщеплением одной молекулы фосфорной кислоты и выделением свободной энергии, которая равна 33-42 кДж/моль. Все реакции с участием АТФ регулируются ферментными системами.
Рис.1. Аденозинтрифосфорная кислота (АТФ)
Энергетический обмен в клетке. Синтез АТФ
Синтез АТФ происходит в мембранах митохондрий в процессе дыхания, поэтому все ферменты и кофакторы дыхательной цепи, все ферменты окислительного фосфорилирования локализованы в данных органеллах.
Синтез АТФ происходит таким образом, что два иона Н + отщепляются от АДФ и фосфата (Р) с правой стороны мембраны, компенсируя потерю двух Н + при восстановлении вещества В. Один из кислородных атомов фосфата переносится на другую сторону мембраны и, присоединив два иона Н + из левого отсека, образует Н 2 О. Остаток фосфорила присоединяется к АДФ, образуя АТФ.
Рис.2. Схема окисления и синтеза АТФ в митохондриальных мембранах
В клетках организмов изучено много биосинтетических реакций, использующих энергию, заключенную в АТФ, в ходе которых происходят процессы карбоксилирования и декарбоксилирования, синтеза амидных связей, образования макроэргических соединений, способных переносить энергию от АТФ к анаболическим реакциям синтеза веществ. Эти реакции играют важную роль в процессах обмена веществ растительных организмов.
С участием АТФ и других макроэргических нуклеозидполифосфатов (ГТФ, ЦТФ, УГФ) может происходить активирование молекул моносахаридов, аминокислот, азотистых оснований, ацилглицеринов путем синтеза активных промежуточных соединений, являющихся производными нуклеотидов. Так, например, в процессе синтеза крахмала с участием фермента АДФ-глюкозо-пирофосфорилазы образуется активированная форма глюкозы — аденозиндифосфатглюкоза, которая легко становится донором глюкозных остатков при формировании структуры молекул этого полисахарида.
Синтез АТФ происходит в клетках всех организмов в процессе фосфорилирования, т.е. присоединения неорганического фосфата к АДФ. Энергия для фосфорилирования АДФ образуется в ходе энергетического обмена. Энергетический обмен, или диссимиляция, представляет собой совокупность реакций расщепления органических веществ, сопровождающихся выделением энергии. В зависимости от среды обитания диссимиляция может протекать в два или три этапа.
У большинства живых организмов — аэробов, живущих в кислородной среде, — в ходе диссимиляции осуществляется три этапа: подготовительный, бескислородный и кислородный, в процессе которых органические вещества распадаются до неорганических соединений. У анаэробов, обитающих в среде, лишенной кислорода, или у аэробов при его недостатке диссимиляция протекает лишь в два первых этапа с образованием промежуточных органических соединений, еще богатых энергией.
Первый этап — подготовительный — заключается в ферментативном расщеплении сложных органических соединений на более простые (белков — на аминокислоты, жиров — на глицерин и жирные кислоты, полисахаридов — на моносахариды, нуклеиновых кислот — на нуклеотиды). Распад органических субстратов пищи осуществляется на разных уровнях желудочно-кишечного тракта многоклеточных организмов. Внутриклеточное расщепление органических веществ происходит под действием гидролитических ферментов лизосом. Высвобождающаяся при этом энергия рассеивается в виде теплоты, а образующиеся малые органические молекулы могут подвергнуться дальнейшему расщеплению или использоваться клеткой как «строительный материал» для синтеза собственных органических соединений.
Второй этап — неполное окисление (бескислородный) — осуществляется непосредственно в цитоплазме клетки, в присутствии кислорода не нуждается и заключается в дальнейшем расщеплении органических субстратов. Главным источником энергии в клетке является глюкоза. Бескислородное, неполное расщепление глюкозы называют гликолизом.
Гликолиз — многоступенчатый ферментативный процесс превращения шестиуглеродной глюкозы в две трехуглеродные молекулы пировиноградной кислоты (пирувата, ПВК) С3Н4О3. В ходе реакций гликолиза выделяется большое количество энергии — 200 кДж/моль. Часть этой энергии (60%) рассеивается в виде теплоты, остальное (40%) используется на синтез АТФ.
В результате гликолиза одной молекулы глюкозы образуется по две молекулы ПВК, АТФ и воды, а также атомы водорода, которые запасаются клеткой в форме НАД Н, т.е. в составе специфического переносчика — никотинамидадениндинуклеотида. Дальнейшая судьба продуктов гликолиза — пирувата и водорода в форме НАД Н — может складываться по-разному. У дрожжей или в клетках растений при недостатке кислорода происходит спиртовое брожение — ПВК восстанавливается до этилового спирта:
В клетках животных, испытывающих временный недостаток кислорода, например в мышечных клетках человека при чрезмерной физической нагрузке, а также у некоторых бактерий происходит молочнокислое брожение, при котором пируват восстанавливается до молочной кислоты. При наличии в среде кислорода продукты гликолиза претерпевают дальнейшее расщепление до конечных продуктов.
Третий этап — полное окисление (дыхание) — протекает при обязательном участии кислорода. Аэробное дыхание представляет собой цепь реакций, контролируемых ферментами внутренней мембраны и матрикса митохондрии. Попав в митохондрию, ПВК взаимодействует с ферментами матрикса и образует: диоксид углерода, который выводится из клетки; атомы водорода, которые в составе переносчиков направляются к внутренней мембране; ацетилкофермент А (ацетил-КоА), который вовлекается в цикл трикарбоновых кислот (цикл Кребса). Цикл Кребса — это цепь последовательных реакций, в ходе которых из одной молекулы ацетил-КоА образуются две молекулы СО2, молекула АТФ и четыре пары атомов водорода, передаваемые на молекулы-переносчики — НАД и ФАД (флавинадениндинуклеотид). Суммарную реакцию гликолиза и цикла Кребса можно представить в следующем виде:
Итак, в результате бескислородного этапа диссимиляции и цикла Кребса молекула глюкозы расщепляется до неорганического диоксида углерода (СО2), а высвободившаяся при этом энергия частично расходуется на синтез АТФ, но в основном сберегается в нагруженных электронами переносчиках НАД Н2 и ФАД Н2. Белки-переносчики транспортируют атомы водорода к внутренней мембране митохондрий, где передают их по цепи встроенных в мембрану белков. Транспорт частиц по цепи переноса осуществляется таким образом, что протоны остаются на внешней стороне мембраны и накапливаются в межмембранном пространстве, превращая его в Н+-резервуар, а электроны передаются на внутреннюю поверхность внутренней митохондриальной мембраны, где соединяются в конечном итоге с кислородом.
В результате деятельности ферментов цепи переноса электронов внутренняя мембрана митохондрий изнутри заряжается отрицательно, а снаружи — положительно (за счет Н), так что между ее поверхностями создается разность потенциалов. Известно, что во внутреннюю мембрану митохондрий встроены молекулы фермента АТФ-синтетазы, обладающие ионным каналом. Когда разность потенциалов на мембране достигает критического уровня (200 мВ), положительно заряженные частицы Н+ силой электрического поля начинают проталкиваться через канал АТФазы и, оказавшись на внутренней поверхности мембраны, взаимодействуют с кислородом, образуя воду.
Нормальное протекание метаболических реакций на молекулярном уровне обусловлено гармоничным сочетанием процессов катаболизма и анаболизма. При нарушении катаболических процессов прежде всего возникают энергетические трудности, нарушаются регенерация АТФ, а также поступление необходимых для биосинтетических процессов исходных субстратов анаболизма. В свою очередь, первичное или связанное с изменениями процессов катаболизма повреждение анаболических процессов ведет к нарушению воспроизведения функционально важных соединений — ферментов, гормонов и др.
Нарушение различных звеньев метаболических цепей неравнозначно по своим последствиям. Наиболее существенные, глубокие патологические изменения катаболизма происходят при повреждении системы биологического окисления при блокаде ферментов тканевого дыхания, гипоксии и др. или повреждении механизмов сопряжения тканевого дыхания и окислительного фосфорилирования (например, разобщение тканевого дыхания и окислительного фосфорилирования при тиреотоксикозе). В этих случаях клетки лишаются основного источника энергии, почти все окислительные реакции катаболизма блокируются или теряют способность аккумулировать освобождающуюся энергию в молекулах АТФ. При ингибировании реакций цикла трикарбоновых кислот выработка энергии в процессе катаболизма сокращается примерно на две трети.
Рассказы о биоэнергетике Скулачев Владимир Петрович
Где и как образуется АТФ?
Первой системой, для которой выяснили механизм образования АТФ, оказался гликолиз — вспомогательный тип энергообеспечения, включающийся в условиях нехватки кислорода. При гликолизе молекула глюкозы расщепляется пополам и полученные обломки окисляются до молочной кислоты.
Такое окисление сопряжено с присоединением фосфорной кислоты к каждому из фрагментов молекулы глюкозы, то есть с их фосфорилированием. Последующий перенос фосфатных остатков с фрагментов глюкзы на АДФ дает АТФ.
Механизм образования АТФ при внутриклеточном дыхании и фотосинтезе долгое время оставался совершенно неясным. Было известно только, что ферменты, катализирующие эти процессы, встроены в биологические мембраны — тончайшие пленки (толщиной около одной миллионной доли сантиметра), состоящие из белков и фосфорилированных жироподобных веществ — фосфолипидов.
Мембраны — важнейший структурный компонент любой живой клетки. Внешняя мембрана клетки отделяет протоплазму от окружающей клетку среды. Клеточное ядро окружено двумя мембранами, которые образуют ядерную оболочку — преграду между внутренним содержимым ядра (нуклеоплазмой) и остальной частью клетки (цитоплазмой). Кроме ядра, в клетках животных и растений находят еще несколько структур, окруженных мембранами. Это эндоплазматическая сеть — система мельчайших трубочек и плоских цистерн, стенки которых образованы мембранами. Это, наконец, митохондрии — шарообразные или вытянутые пузырьки размером мельче ядра, но крупнее компонентов эндоплазматической сети. Диаметр митохондрии обычно около микрона, хотя иногда митохондрии образуют ветвящиеся и сетчатые структуры протяженностью в десятки микрон.
В клетках зеленых растений, помимо ядра, эндоплазматической сети и митохондрий, находят еще и хлоропласты — мембранные пузырьки более крупные, чем митохондрии.
Каждая из этих структур выполняет свою, специфическую биологическую функцию. Так, ядро — вместилище ДНК. Здесь происходят процессы, лежащие в основе генетической функции клетки, и начинается сложная цепь процессов, приводящая в конечном итоге к синтезу белка. Этот синтез завершается в мельчайших гранулах — рибосомах, большая часть которых связана с эндоплазматической сетью. В митохондриях происходят окислительные реакции, совокупность которых называется внутриклеточным дыханием. Хлоропласты отвечают за фотосинтез.
Клетки бактерий устроены проще. Обычно они имеют только две мембраны — внешнюю и внутреннюю. Бактерия — это как бы мешок в мешке, а точнее, очень мелкий пузырек с двойной стенкой. Здесь нет ни ядра, ни митохондрий, ни хлоропластов.
Существует гипотеза, что митохондрии и хлоропласты произошли из бактерий, захваченных клеткой более крупного и высокоорганизованного существа. Действительно, биохимия митохондрий и хлоропластов во многом напоминает бактериальную. Морфологически митохондрии и хлоропласты тоже в известном смысле подобны бактериям: они окружены двумя мембранами. Во всех трех случаях: в бактериях, митохондриях и хлоропластах — синтез АТФ происходит во внутренней мембране.
Долгое время считалось, что образование АТФ при дыхании и фотосинтезе протекает аналогично уже известному превращению энергии при гликолизе (фосфорилирование расщепляемого вещества, его окисление и перенос остатка фосфорной кислоты на АДФ). Однако все попытки экспериментально доказать эту схему оканчивались неудачей.
Аденозинтрифосфорная кислота-АТФ — обязательный энергетический компонент любой живой клетки. АТФ также нуклеотид, состоящий из азотистого основания аденина, сахара рибозы и трех остатков молекулы фосфорной кислоты. Это неустойчивая структура. В обменных процессах от нее последовательно отщепляются остатки фосфорной кислоты путем разрыва богатой энергией, но непрочной связи между вторым и третьим остатками фосфорной кислоты. Отрыв одной молекулы фосфорной кислоты сопровождается выделением около 40 кДж энергии. В этом случае АТФ переходит в аденозиндифосфорную кислоту (АДФ), а при дальнейшем отщеплении остатка фосфорной кислоты от АДФ образуется аденозинмонофосфорная кислота (АМФ).
Схема строения АТФ и превращения ее в АДФ (Т.А. Козлова, В.С. Кучменко. Биология в таблицах. М.,2000)
Следовательно, АТФ — своеобразный аккумулятор энергии в клетке, который «разряжается» при ее расщеплении. Распад АТФ происходит в процессе реакций синтеза белков, жиров, углеводов и любых других жизненных функций клеток. Эти реакции идут с поглощением энергии, которая извлекается в ходе расщепления веществ.
АТФ синтезируется в митохондриях в несколько этапов. Первый из них — подготовительный — протекает ступенчато, с вовлечением на каждой ступени специфических ферментов. При этом сложные органические соединения расщепляются до мономеров: белки — до аминокислот, углеводы — до глюкозы, нуклеиновые кислоты — до нуклеотидов и т. д. Разрыв связей в этих веществах сопровождается выделением небольшого количества энергии. Образовавшиеся мономеры под действием других ферментов могут претерпеть дальнейший распад с образованием более простых веществ вплоть до диоксида углерода и воды.
Схема Синтез АТФ в мвтохондрии клетки
ПОЯСНЕНИЯ К СХЕМЕ ПРЕВРАЩЕНИЕ ВЕЩЕСТВ И ЭНЕРГИИ В ПРОЦЕССЕ ДИССИМИЛЯЦИИ
I этап — подготовительный: сложные органические вещества под действием пищеварительных ферментов распадаются на простые, при этом выделяется только тепловая энергия.
Белки ->аминокислоты
Жиры-> глицерин и жирные кислоты
Крахмал ->глюкоза
II этап-гликолиз (бескислородный): осуществляется в гиалоплазме, с мембранами не связан; в нем участвуют ферменты; расщеплению подвергается глюкоза:
У дрожжевых грибов молекула глюкозы без участия кислорода превращается в этиловый спирт и диоксид углерода (спиртовое брожение):
У других микроорганизмов гликолиз может завершаться образованием ацетона, уксусной кислоты и т, д. Во всех случаях распад одной молекулы глюкозы сопровождается образованием двух молекул АТФ. В ходе бескислородного расщепления глюкозы в виде химической связи в молекуле АТФ сохраняется 40% анергии, а остальная рассеивается в виде теплоты.
III этап-гидролиз (кислородный): осуществляется в митохондриях, связан с матриксом митохондрий и внутренней мембраной, в нем участвуют ферменты, расщеплению подвергается молочная кислота: СзН6Оз+ЗН20 —>3СО2+ 12Н. С02 (диоксид углерода) выделяется из митохондрий в окружающую среду. Атом водорода включается в цепь реакций, конечный результат которых — синтез АТФ. Эти реакции идут в такой последовательности:
1. Атом водорода Н с помощью ферментов-переносчиков поступает во внутреннюю мембрану митохондрий, образующую кристы, где он окисляется: Н-е—>H+
2. Протон водорода H+ (катион) выносится переносчиками на наружную поверхность мембраны крист. Для протонов эта мембрана непроницаема, поэтому они накапливаются в межмембранном пространстве, образуя протонный резервуар.
3. Электроны водорода e переносятся на внутреннюю поверхность мембраны крист и тут же присоединяются к кислороду с помощью фермента оксидазы, образуя отрицательно заряженный активный кислород (анион): O2 + е—>O2-
4. Катионы и анионы по обе стороны мембраны создают разноименно заряженное электрическое поле, и когда разность потенциалов достигнет 200 мВ, начинает действовать протонный канал. Он возникает в молекулах ферментов АТФ-синтетаз, которые встроены во внутреннюю мембрану, образующую кристы.
5. Через протонный канал протоны водородаH+ устремляются внутрь митохондрий, создавая высокий уровень энергии, большая часть которой идет на синтез АТФ из АДФ и Ф (АДФ+Ф—>АТФ), а протоны H+ взаимодействуют с активным кислородом, образуя воду и молекулярный 02:
(4Н++202- —>2Н20+02)
Таким образом, О2, поступающий в митохондрии в процессе дыхания организма, необходим для присоединения протонов водорода Н. При его отсутствии весь процесс в митохондриях прекращается, так как электронно-транспортная цепь перестает функционировать. Общая реакция III этапа:
(2СзНбОз + 6Oз + 36АДФ + 36Ф —> 6С02 + 36АТФ + +42Н20)
В результате расщепления одной молекулы глюкозы образуются 38 молекул АТФ: на II этапе — 2 АТФ и на III этапе — 36 АТФ. Образовавшиеся молекулы АТФ выходят за пределы митохондрии и участвуют во всех процессах клетки, где необходима энергия. Расщепляясь, АТФ отдает энергию (одна фосфатная связь заключает 40 кДж) и в виде АДФ и Ф (фосфата) возвращается в митохондрии.
Расшифровка происхождения миофибробластов при фиброзе почек человека
Даффилд, Дж. С. Клеточные и молекулярные механизмы фиброза почек. J. Clin. Инвестируйте . 124 , 2299–2306 (2014).
CAS PubMed PubMed Central Google Scholar
Kramann, R., DiRocco, D. P. & Humphreys, B. D. Понимание происхождения, активации и регуляции матрикс-продуцирующих миофибробластов для лечения фиброзной болезни. Дж. Патол . 231 , 273–289 (2013).
CAS PubMed Google Scholar
Фридман, С. Л., Шеппард, Д., Даффилд, Дж. С. и Виолетт, С. Терапия фиброзных заболеваний: приближается к исходной черте. Sci. Пер. Med . 5 , 167ср1 (2013).
PubMed Google Scholar
Фальке, Л. Л., Голизаде, С., Goldschmeding, R., Kok, R.J. & Nguyen, T.Q. Различное происхождение миофибробластов — последствия для фиброза почек. Nat. Преподобный Нефрол . 11 , 233–244 (2015).
CAS PubMed Google Scholar
Young, M. D. et al. Одноклеточные транскриптомы почек человека раскрывают клеточную идентичность почечных опухолей. Наука 361 , 594–599 (2018).
ADS CAS PubMed PubMed Central Google Scholar
Канг, Х. М. и др. Дефектное окисление жирных кислот в эпителиальных клетках почечных канальцев играет ключевую роль в развитии фиброза почек. Nat. Med . 21 , 37–46 (2015).
CAS PubMed Google Scholar
Naba, A. et al. Внеклеточный матрикс: инструменты и идеи для эпохи «омиков». Матрикс Биол . 49 , 10–24 (2016).
CAS PubMed Google Scholar
Fan, Y. et al. Сравнение транскриптомных профилей почек при ранней и поздней стадии диабетической нефропатии выявляет новые потенциальные механизмы прогрессирования заболевания. Диабет 68 , 2301–2314 (2019).
CAS PubMed PubMed Central Google Scholar
Криз В., Кайсслинг Б. и Ле Хир М. Эпителиально-мезенхимальный переход (ЭМП) при фиброзе почек: факт или фантазия? J. Clin. Инвестируйте . 121 , 468–474 (2011).
CAS PubMed PubMed Central Google Scholar
Huang, S. & Susztak, K. Эпителиальная пластичность по сравнению с EMT при фиброзе почек. Trends Mol. Med . 22 , 4–6 (2016).
CAS PubMed Google Scholar
Elices, M. J. et al. VCAM-1 на активированном эндотелии взаимодействует с интегрином лейкоцитов VLA-4 в сайте, отличном от сайта связывания VLA-4 / фибронектина. Cell 60 , 577–584 (1990).
CAS PubMed Google Scholar
Канг, Х. М. и др. Sox9-позитивные клетки-предшественники играют ключевую роль в регенерации эпителия почечных канальцев у мышей. Cell Rep . 14 , 861–871 (2016).
CAS PubMed PubMed Central Google Scholar
Ramachandran, P. et al.Разрешение фиброзной ниши цирроза печени человека на одноклеточном уровне. Природа 575 , 512–518 (2019).
ADS CAS PubMed PubMed Central Google Scholar
Ван, Ю.-Й. и другие. Переход макрофагов в миофибробласты способствует интерстициальному фиброзу при хроническом повреждении почечного аллотрансплантата. J. Am. Soc. Нефрол . 28 , 2053–2067 (2017).
CAS PubMed PubMed Central Google Scholar
Henderson, N.C. et al. Нацеливание на интегрин αv определяет основной молекулярный путь, который регулирует фиброз в нескольких органах. Nat. Med . 19 , 1617–1624 (2013).
CAS PubMed Google Scholar
Wernig, G. et al. Объединяющий механизм при различных фиброзных заболеваниях. Proc. Natl Acad. Sci. США 114 , 4757–4762 (2017).
CAS PubMed Google Scholar
Venkatachalam, M.A., Weinberg, J.M., Kriz, W. & Bidani, A.K. Неудачное восстановление канальцев, переход AKI-CKD и прогрессирование заболевания почек. J. Am. Soc. Нефрол . 26 , 1765–1776 (2015).
CAS PubMed PubMed Central Google Scholar
Kramann, R. et al. Парабиоз и секвенирование одноклеточной РНК выявляют ограниченный вклад моноцитов в миофибробласты при фиброзе почек. JCI Insight 3 , e99561 (2018).
PubMed Central Google Scholar
Gerstein, M. B. et al. Архитектура регулирующей сети человека, полученная на основе данных ENCODE. Природа 489 , 91–100 (2012).
ADS CAS PubMed PubMed Central Google Scholar
Буэнростро, Дж. Д., Гиреси, П.Г., Заба, Л. С., Чанг, Х. Ю. и Гринлиф, В. Дж. Транспозиция нативного хроматина для быстрого и чувствительного эпигеномного профилирования открытого хроматина, ДНК-связывающих белков и положения нуклеосом. Nat. Методы 10 , 1213–1218 (2013).
CAS PubMed PubMed Central Google Scholar
Palumbo-Zerr, K. et al. Орфанный ядерный рецептор NR4A1 регулирует передачу сигналов трансформирующего фактора роста-β и фиброз. Nat. Med . 21 , 150–158 (2015).
CAS PubMed Google Scholar
Schubert, M. et al. Гены реакции на возмущение обнаруживают сигнальные следы в экспрессии раковых генов. Nat. Коммуна . 9 , 20 (2018).
ADS PubMed PubMed Central Google Scholar
Zhao, S. et al. NKD2, негативный регулятор передачи сигналов Wnt, подавляет рост опухоли и метастазирование в остеосаркоме. Онкоген 34 , 5069–5079 (2015).
CAS PubMed PubMed Central Google Scholar
Li, C. et al. Миристоилированный Naked2 сопровождает трансформирующий фактор роста α к базолатеральной плазматической мембране поляризованных эпителиальных клеток. Proc. Natl Acad. Sci. США 101 , 5571–5576 (2004).
ADS CAS PubMed Google Scholar
Moerman, T. et al. GRNBoost2 и Arboreto: эффективный и масштабируемый вывод сетей регуляции генов. Биоинформатика 35 , 2159–2161 (2019).
CAS Google Scholar
Lemos, D. R. et al. Интерлейкин-1β активирует MYC-зависимый метаболический переключатель в стромальных клетках почек, необходимый для прогрессирующего тубулоинтерстициального фиброза. J. Am. Soc. Нефрол . 29 , 1690–1705 (2018).
CAS PubMed PubMed Central Google Scholar
Tsukui, T. et al. Атлас клеток легких, продуцирующих коллаген, идентифицирует несколько подгрупп с разной локализацией и релевантностью для фиброза. Nat. Коммуна . 11 , 1920 (2020).
ADS CAS PubMed PubMed Central Google Scholar
Adams, T. S. et al. Одноклеточная последовательность РНК выявляет эктопические и аберрантные популяции резидентных клеток легких при идиопатическом фиброзе легких. Sci. Adv. 6 , eaba1983 (2020).
CAS PubMed PubMed Central Google Scholar
Kramann, R. et al. Периваскулярные предшественники Gli1 + вносят ключевой вклад в индуцированный повреждением фиброз органа. Cell Stem Cell 16 , 51–66 (2015).
CAS PubMed Google Scholar
Такасото, М.и другие. Органоиды почек из iPS-клеток человека содержат множество клонов и моделируют нефрогенез человека. Природа 526 , 564–568 (2015).
ADS Google Scholar
Corces, M. R. et al. Усовершенствованный протокол ATAC-seq снижает фон и позволяет исследовать замороженные ткани. Nat. Методы 14 , 959–962 (2017).
CAS PubMed PubMed Central Google Scholar
Шривастава А., Малик Л., Смит Т., Садбери И. и Патро Р. Алевин эффективно оценивают точную численность генов на основе данных dscRNA-seq. Биология генома . 20 , 65 (2019).
PubMed PubMed Central Google Scholar
Patro, R., Duggal, G., Love, M. I., Irizarry, R. A. & Kingsford, C. Salmon обеспечивает быструю и достоверную количественную оценку экспрессии транскрипта. Nat. Методы 14 , 417–419 (2017).
CAS PubMed PubMed Central Google Scholar
Frankish, A. et al. Справочная аннотация GENCODE для геномов человека и мыши. Nucleic Acids Res . 47 (D1), D766 – D773 (2019).
CAS PubMed Google Scholar
Lun, A. T. L., McCarthy, D. J. & Marioni, J. C. Пошаговый рабочий процесс для низкоуровневого анализа данных РНК-секвенирования отдельных клеток с помощью Bioconductor. F1000 Рез. . 5 , 2122 (2016).
Google Scholar
Scrucca, L., Fop, M., Murphy, T. B. & Raftery, A. E. mclust 5: кластеризация, классификация и оценка плотности с использованием гауссовских моделей конечной смеси. Р Дж . 8 , 289–317 (2016).
PubMed PubMed Central Google Scholar
Фортунато, С.И Бартелеми, М. Предел разрешения при обнаружении сообщества. Proc. Natl Acad. Sci. США 104 , 36–41 (2007).
ADS CAS PubMed Google Scholar
Zeisel, A. et al. Молекулярная архитектура нервной системы мыши. Ячейка 174 , 999–1014.e22 (2018).
CAS PubMed PubMed Central Google Scholar
Lake, B. B. et al. Одноядерный конвейер секвенирования РНК для расшифровки молекулярной анатомии и патофизиологии почек человека. Nat. Коммуна . 10 , 2832 (2019).
ADS PubMed PubMed Central Google Scholar
Clark, J. Z. et al. Представление и относительное количество селективных маркеров клеточного типа в данных по РНК-Seq цельной почки. Почки Инт . 95 , 787–796 (2019).
CAS PubMed PubMed Central Google Scholar
Wu, C. et al. BioGPS: расширяемый и настраиваемый портал для запросов и организации ресурсов аннотации генов. Биология генома . 10 , R130 (2009).
PubMed PubMed Central Google Scholar
Durinck, S. et al. BioMart и Bioconductor: мощная связь между биологическими базами данных и анализом данных микрочипов. Биоинформатика 21 , 3439–3440 (2005).
CAS PubMed Google Scholar
Durinck, S., Spellman, P. T., Birney, E. & Huber, W. Отображение идентификаторов для интеграции наборов геномных данных с пакетом R / Bioconductor biomaRt. Nat. Protoc. 4 , 1184–1191 (2009).
CAS PubMed PubMed Central Google Scholar
Stuart, T. et al. Всесторонняя интеграция одноклеточных данных. Cell 177 , 1888–1902.e21 (2019).
CAS PubMed PubMed Central Google Scholar
Сюй, С. и Су, З. Идентификация типов клеток из одноклеточных транскриптомов с использованием нового метода кластеризации. Биоинформатика 31 , 1974–1980 (2015).
CAS PubMed PubMed Central Google Scholar
Macosko, E. Z. et al. Профилирование экспрессии отдельных клеток с высокой степенью параллельности генома с использованием нанолитровых капель. Cell 161 , 1202–1214 (2015).
CAS PubMed PubMed Central Google Scholar
Альтер О., Браун П. О. и Ботштейн Д. Разложение по сингулярным значениям для обработки и моделирования данных экспрессии в масштабе всего генома. Proc. Natl Acad. Sci. США 97 , 10101–10106 (2000).
ADS CAS PubMed Google Scholar
Росвалл, М. и Бергстром, К. Т. Карты случайных блужданий в сложных сетях показывают структуру сообщества. Proc. Natl Acad. Sci. США 105 , 1118–1123 (2008).
ADS CAS PubMed Google Scholar
Dahlin, J. S. et al. Одноклеточный гематопоэтический ландшафт разрешает 8 клональных траекторий и дефекты у Kit мутантных мышей. Кровь 131 , e1 – e11 (2018).
CAS PubMed PubMed Central Google Scholar
Ибрагим, М. М. и Краманн, Р. genesorteR: ранжирование характеристик в сгруппированных данных по отдельным ячейкам. Препринт на https://doi.org/10.1101/676379 (2019).
Haghverdi, L., Lun, A. T. L., Morgan, M. D. и Marioni, J. C. Пакетные эффекты в данных секвенирования РНК одной клетки корректируются путем сопоставления общих ближайших соседей. Nat. Биотехнология . 36 , 421–427 (2018).
CAS PubMed PubMed Central Google Scholar
Блондель В. Д., Гийом Ж.-Л., Ламбьотт Р. и Лефевр Э. Быстрое развертывание сообществ в большие сети. J. Stat. Мех . 2008 , P10008 (2008).
Google Scholar
Лун А.Т.Л., Бах К.И Мариони, Дж. С. Объединение клеток для нормализации данных секвенирования одноклеточной РНК со многими нулевыми счетчиками. Биология генома . 17 , 75 (2016).
PubMed Google Scholar
Рэнд, В. М. Объективные критерии оценки методов кластеризации. J. Am. Стат. Assoc . 66 , 846–850 (1971).
Google Scholar
Янг, Л., Лю, Дж., Лу, К., Риггс, А. Д. и Ву, X. SAIC: итеративный подход кластеризации для анализа данных последовательности РНК отдельных клеток. BMC Genomics 18 (Приложение 6), 689 (2017).
PubMed PubMed Central Google Scholar
Караискос Н. и др. Эмбрион Drosophila при разрешении одноклеточного транскриптома. Наука 358 , 194–199 (2017).
ADS CAS PubMed Google Scholar
Макиннес, Л., Хили, Дж. И Мелвилл, Дж. UMAP: приближение и проекция однородного многообразия для уменьшения размерности. Препринт на https://arxiv.org/abs/1802.03426 (2018).
Street, K. et al. Slingshot: происхождение клеток и псевдодинамический вывод для транскриптомики одиночных клеток. BMC Genomics 19 , 477 (2018).
PubMed PubMed Central Google Scholar
Субраманян, А.и другие. Анализ обогащения набора генов: основанный на знаниях подход к интерпретации профилей экспрессии в масштабе всего генома. Proc. Natl Acad. Sci. США 102 , 15545–15550 (2005).
ADS CAS Google Scholar
Liberzon, A. et al. База данных молекулярных сигнатур (MSigDB) 3.0. Биоинформатика 27 , 1739–1740 (2011).
CAS PubMed PubMed Central Google Scholar
Yu, G., Wang, L.-G., Han, Y. & He, Q.-Y. clusterProfiler: пакет R для сравнения биологических тем среди генных кластеров. OMICS 16 , 284–287 (2012).
CAS PubMed PubMed Central Google Scholar
Консорциум генных онтологий. Генная онтология: инструмент для объединения биологии. Nat. Genet . 25 , 25–29 (2000).
PubMed Central Google Scholar
Holland, C.H. et al. Надежность и применимость инструментов анализа факторов транскрипции и путей к данным одноклеточной РНК-seq. Биология генома . 21 , 36 (2020).
CAS PubMed PubMed Central Google Scholar
Macosko, E.Z. et al. Профилирование экспрессии отдельных клеток с высокой степенью параллельности генома с использованием нанолитровых капель. Cell 161 , 1202–1214 (2015).
CAS PubMed PubMed Central Google Scholar
Yang, J. et al. Транскриптомика одиночных клеток выявляет неожиданные особенности ранних гемопоэтических предшественников. Nucleic Acids Res . 45 , 1281–1296 (2017).
CAS PubMed Google Scholar
Гу, З., Эйлс, Р. и Шлеснер, М. Сложные тепловые карты выявляют закономерности и корреляции в многомерных геномных данных. Биоинформатика 32 , 2847–2849 (2016).
CAS PubMed PubMed Central Google Scholar
Bushnell, B. BBMap: быстрый, точный, совместимый со сваркой выравниватель https://www.osti.gov/biblio/1241166 (2014).
Dobin, A. et al. STAR: сверхбыстрый универсальный выравниватель RNA-seq. Биоинформатика 29 , 15–21 (2013).
CAS PubMed Google Scholar
Li, H. et al. Формат Sequence Alignment / Map и SAMtools. Биоинформатика 25 , 2078–2079 (2009).
PubMed PubMed Central Google Scholar
Quinlan, A.R.BEDTools: швейцарский армейский инструмент для анализа свойств генома. Curr. Protoc. Биоинформатика 47 , 11.12.1–11.12.34 (2014).
Google Scholar
Adey, A. et al. Быстрое конструирование библиотек фрагментов дробовика с малым вводом и смещением путем транспозиции in vitro с высокой плотностью. Биология генома . 11 , R119 (2010).
CAS PubMed PubMed Central Google Scholar
Ибрагим, М. М., Лакади, С. А. и Олер, У. JAMM: искатель пиков для совместного анализа реплик NGS. Биоинформатика 31 , 48–55 (2015).
CAS PubMed Google Scholar
Luehr, S., Hartmann, H. & Söding, J. Веб-сервер XXmotif для eXhaustive, основанного на весовом матриксе обнаружения мотивов в нуклеотидных последовательностях. Nucleic Acids Res . 40 , W104 – W109 (2012).
CAS PubMed PubMed Central Google Scholar
Грант, К. Э., Бейли, Т. Л. и Ноубл, У. С. FIMO: сканирование на наличие вхождений данного мотива. Биоинформатика 27 , 1017–1018 (2011).
CAS PubMed PubMed Central Google Scholar
Ramírez, F. et al. deepTools2: веб-сервер нового поколения для глубокого анализа данных. Nucleic Acids Res . 44 (W1), W160 – W165 (2016).
PubMed PubMed Central Google Scholar
Robinson, J. T. et al. Программа просмотра интегративной геномики. Nat. Биотехнология . 29 , 24–26 (2011).
CAS PubMed PubMed Central Google Scholar
Aibar, S. et al. SCENIC: вывод и кластеризация одноячеечной регуляторной сети. Nat. Методы 14 , 1083–1086 (2017).
CAS PubMed PubMed Central Google Scholar
Шахт, Т., Освальд, М., Эйлс, Р., Эйхмюллер, С.B. & König, R. Оценка активности факторов транскрипции по влиянию на их гены-мишени. Биоинформатика 30 , i401 – i407 (2014).
CAS PubMed PubMed Central Google Scholar
Alvarez, M. J. et al. Функциональная характеристика соматических мутаций при раке с использованием сетевого вывода об активности белков. Nat. Genet . 48 , 838–847 (2016).
CAS PubMed PubMed Central Google Scholar
Гарсия-Алонсо, Л., Холланд, К. Х., Ибрагим, М. М., Турей, Д. и Саез-Родригес, Дж. Контрольный показатель и интеграция ресурсов для оценки активности факторов транскрипции человека. Genome Res . 29 , 1363–1375 (2019).
CAS PubMed PubMed Central Google Scholar
de Kanter, J. K., Lijnzaad, P., Candelli, T., Margaritis, T. & Holstege, F. C. P. CHETAH: метод селективной иерархической идентификации типов клеток для секвенирования одноклеточной РНК. Nucleic Acids Res . 47 , e95 (2019).
PubMed PubMed Central Google Scholar
Ефремова М., Венто-Тормо М., Тейхманн С. А. и Венто-Тормо Р. CellPhoneDB: определение межклеточной коммуникации на основе комбинированной экспрессии мультисубъединичных комплексов лиганд-рецептор. Nat. Protoc. 15 , 1484–1506 (2020).
CAS PubMed Google Scholar
Soneson, C., Love, M. I. & Robinson, M. D. Дифференциальный анализ RNA-seq: оценки на уровне транскриптов улучшают выводы на уровне генов. F1000 Рез. . 4 , 1521 (2015).
Google Scholar
Ritchie, M. E. et al. limma обеспечивает анализ дифференциальной экспрессии для секвенирования РНК и исследований на микрочипах. Nucleic Acids Res . 43 , e47 (2015).
PubMed PubMed Central Google Scholar
Короткевич Г., Сухов В., Сергушичев А. Анализ быстрого пополнения набора генов. Препринт на https://doi.org/10.1101/060012 (2019).
Произошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно.Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом.Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу.Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файлах cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта.Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
(PDF) Расшифровка функции и регуляции 12-часового цикла млекопитающих
наиболее ограниченного фазового распределения в
различных тканях павиана с пиком от
ZT12 до ZT16, среди всех ключевых циркадных
часовых TF (Mure et al., 2018). Если взаимосвязь между иерархией
и распределением фаз
в контроле циркадных часов
может быть экстраполирована на регулировку 12-часовых часов
, то семейство ETS из
TF, вероятно, будет из Крайне важно, так как их фазы в основном совпадают с
в четырех тканях с обильным 12-часовым транскриптомом (печень, надпочечники,
,железы, MUS и BAT) (рис. 3C). С другой стороны,
, фазы TF bZIP Atf6,
Creb3 и Creb3l2, а также Nfyb, на
несопоставимы в четырех тканях, причем
в BAT и MUS в противофазе от тех
. в печени и надпочечниках (рис. 3C).
Интересно, что фазы этих TF
в значительной степени соответствуют фазовому распределению
12-часового транскриптома в
этих четырех тканях (рис. 1C), предполагая, что
TFs bZIP и NFY могут вносят еще
в регуляцию 12-часовых выходных генов
.
В итоге, здесь я предлагаю, чтобы TF, принадлежащие
к семействам ETS, bZIP и NFY, были основными транскрипционными медиаторами
малийских 12-часовых часов mam-
со сложными положительными
прямой и отрицательной петли обратной связи
и, соединяющие их (рис. 3А).Кроме того, стоит отметить, что
не исключает возможности того, что другие TF также могут быть задействованы в регуляции транскрипции
12-часовых часов млекопитающих в более чем
тканях. специфическим образом (например, различные члены семейства KLF
также демонстрировали обогащение мотива ДНК
при 12-часовом цикле промоторов генов
и 12-часовых ритмах экспрессии генов в ограниченном количестве тканей
). В качестве последнего примечания:
, по-видимому, точно так же, как циркадный ритм,
12-часовые часы млекопитающих также подчиняются посттранскрипционному и пост-
трансляционному контролю
(Zhu et al., 2017; Пан
и др., 2020). Из-за ограниченного количества страниц я не буду обсуждать эти темы здесь.
Заключение
В этой краткой перспективе я обсудил
наши текущие знания о распространенности, функции
и регуляции 12-часовых ритмов у млекопитающих
. Из-за очень ранней стадии
этой области, многие из свидетельств
, представленных в этой статье, все еще являются предварительными
, а некоторые выводы
, следовательно, несколько спекулятивны и подлежат изменению. с новыми экспериментальными данными
.Я искренне надеюсь, что эта короткая статья
привлечет больше ученых в зарождающуюся область
12-часовых ритмов млекопитающих,
и еще много нерешенных вопросов, требующих ответа!
[Это исследование было поддержано Американской
Диабетической ассоциацией, младший факультет
Премия развития 1-18-JDF-025 до BZ]
Под редакцией Джиаруи Ву
Ссылки
Agrawal Singh, S., Lerdrup, M ., Гомеш А.R., et al.
(2019). PLZF нацелен на энхансеры развития для активации
во время остеогенной дифференцировки мезенхимальных стволовых клеток человека hu-
. eLife 8, e40364.
Анафи Р.С., Франси Л.Дж., Хогенеш Дж. Б. и др.
(2017). CYCLOPS выявляет человеческие транскрипционные ритмы
в состоянии здоровья и болезни. Proc.
Natl Acad. Sci. USA 114,5312–5317.
Antoulas, A.C., Zhu, B., Zhang, Q., et al. (2018). Новый математический метод
для выявления колебаний в транскрипции генов
: сравнительное исследование.PLoS One 13, e0198503.
Cretenet, G., Le Clech, M., and Gachon, F. (2010).
Скоординированный по суточным часам 12-часовой период
ритмическая активация пути IRE1apathway контролирует метаболизм липидов в печени мышей. Cell
Metab. 11,47–57.
Hughes, M.E., DiTacchio, L., Hayes, K.R., et al.
(2009). Гармоники транскрипции циркадного гена-
у млекопитающих. PLoS Genet. 5, e1000442.
Hughes, M.E., Hong, H.K., Chong, J.L., et al.
(2012). Специфическое для мозга спасение Clock выявило
системных транскрипционных ритмов в риферической ткани pe-
. PLoS Genet. 8, e1002835.
Кофф К. и Фивейски М.М. (1975). Гармония
из Науки Акустики. Studio City, CA:
Koff Music Co.
Koike, N., Yoo, S.H., Huang, H.C., et al. (2012).
Транскрипционная архитектура и хроматин
Пейзаж основных циркадных часов у млекопитающих
малах.Наука 338,349–354.
Mavrothalassitis, G., and Ghysdael, J. (2000).
Белки семейства ETS с транскрипционной репрессорной активностью
. Онкоген 19,6524–6532.
Макмастер, C.R. (2001). Липидный обмен и пузырьки
перемещение: больше, чем просто смазка транспортных средств
машин. Биохим. Cell Biol. 79 681–692.
Менет, Дж. С., Пескаторе, С., Росбаш, М.
(2014). ЧАСЫ: BMAL1 — это пионерский транскрипционный фактор
.Genes Dev. 28,8–13.
Mure, L.S., Le, H.D., Benegiamo, G., et al. (2018).
Атлас суточного транскриптома приматов
через основные нервные и периферические ткани.
Наука 359, eaao0318.
Oldfield, A.J., Yang, P., Conway, A.E., et al.
(2014). Белок гистонового складчатого домена NF-Y pro-
доступность хроматина для клеток
типоспецифических основных факторов транскрипции. Мол.
Ячейка 55,708–722.
Пан, Ю., Ballance, H., Meng, H., et al. (2020). 12-h
часовая регуляция потока генетической информации
XBP1. PLoS Biol. 18, e3000580.
Раз, Г. (2012). Возникновение даосизма:
Создание традиции (1-е изд.). Лондон:
Рутледж.
Санчес-Гурмачеш, Дж., И Гертин, Д.А.
(2014). Линии адипоцитов: прослеживание происхождения
жира. Биохим. Биофиз. Acta 1842,
340–351.
Сени, М.Л., Кэхилл, К., Enwright, J.F., 3rd, et al.
(2019). Суточные ритмы экспрессии генов в префронтальной коре
при шизофрении. Nat.
Общ. 10,3355.
Takahashi, J.S. (2017). Транскрипционная архитектура циркадных часов млекопитающих. Nat.
Ред. Genet. 18,164–179.
Табен П.Ф. и Вестермарк П.О. (2014).
Обнаружение ритмов во временных рядах с RAIN. J.
Biol. Ритмы 29,391–400.
Туэрауф, Д.J., Marcinko, M., Belmont, P.J., et al.
(2007). Эффекты специфических для изоформ характеристик
ATF6a и ATF6bon эндоплазматической
экспрессии гена стрессового ответа ретикулума
и жизнеспособности клеток. J. Biol. Chem. 282,
22865–22878.
Вестермарк, П.О., и Герцель, Х. (2013).
Механизм генерации 12-часового ритма
по циркадным часам. Cell Rep. 3,
1228–1238.
Уилкоксон, Д., и Чжан, Л.(2008). Circatidal
часов. Curr. Биол. 18, R753 – R755.
Йошида, Х., Оку, М., Судзуки, М., и др. (2006).
pXBP1 (U), кодируемый в пре-мРНК XBP1 nega-
, активно регулирует ответ развернутого белка
активатор pXBP1 (S) в ответ на стресс ER млекопитающих
sponse. J. Cell Biol. 172,565–575.
Zhang, R., Lahens, N.F., Ballance, H.I., et al.
(2014). Атлас экспрессии циркадных генов у
млекопитающих: значение для биологии и медицины
кино.Proc. Natl Acad. Sci. USA 111,
16219–16224.
Чжу, Б., Даксо, С.С., и О’Мэлли, Б.В. (2018).
Открытие «Musica Universalis» клетки:
краткая история биологических 12-часовых ритмов. J.
Endocr. Soc. 2,727–752.
Чжу Б., Гейтс Л.А., Сташи Э. и др. (2015).
Коактиватор-зависимая осцилляция хрома
Доступность олова определяет амплитуда циркадных генов
при загрузке REV-ERB. Мол.Cell 60,
769–783.
Zhu, B., Zhang, Q., Pan, Y., et al. (2017). 12-часовая координата
автономных клеток млекопитающих
определяет метаболические ритмы и ритмы стресса. Cell
Metab. 25,1305–1319.e9.
Zhu, X., Wang, Y., Pi, W., et al. (2012). NF-Y
привлекает как активатор транскрипции, так и репрессор
для модуляции тканевой и развивающейся экспрессии гена c-глобина
человека. PLoS One 7, e47175.
Расшифровка 12-часовых часов млекопитающих | 7
Загружено с https://academic.oup.com/jmcb/advance-article/doi/10.1093/jmcb/mjaa021/5834720 гостем 2 декабря 2020 г.
% PDF-1.4 % % ABCpdf 11302 92 0 объект > эндобдж xref 92 22 0000000032 00000 н. 0000001891 00000 н. 0000001966 00000 н. 0000002163 00000 н. 0000002492 00000 н. 0000002765 00000 н. 0000002898 00000 н. 0000002943 00000 н. 0000003097 00000 н. 0000003561 00000 н. 0000003810 00000 н. 0000004193 00000 п. 0000004343 00000 п. 0000005074 00000 н. 0000005318 00000 н. 0000005727 00000 н. 0000005917 00000 н. 0000007060 00000 н. 0000007337 00000 н. 0000012937 00000 п. 0000015815 00000 п. 0000041884 00000 п. трейлер ] / Инфо 90 0 R / Назад 1296394 / Корень 93 0 R / Размер 114 / Источник (WeJXFxNO4fJduyUMetTcP9 + oaONfINN4 + d61k8TlCWgP6nGmCGPlcM9oOUhvz215B9khgm8VtCFmyd8gIrwOjQRAIjPsWhM4vgMCV \ 8KvVF / K8lfv6PtDYZLeSUqQVGsKdKV1FtlhqhUnGEg =) >> startxref 0 %% EOF 93 0 объект > эндобдж 94 0 объект > эндобдж 95 0 объект > поток 㙧 5i ~ AtI ‘# + G {N + C X | lR; j
Канал связывания мРНК в области ES6S трансляции 48S-PIC способствует раскручиванию и сканированию РНК
Сводка приемки:
В статье дается четкое понимание механизма, объясняющего обратную корреляцию между содержанием мРНК 5’UTR G + C / ΔG и эффективностью трансляции мРНК.Это подтверждает модель функции eIF4A в раскручивании вторичной структуры 5 ‘мРНК для облегчения рекрутирования рибосом. Они документально подтвердили роль рибосомной области ES6S в контроле сканирования рибосом. Они предоставляют доказательства того, что мРНК проникает в область ES6S пре-инициаторного комплекса 48S (PIC) и разворачивается с помощью РНК-геликазы eIF4AI, делая 48S PIC более процессивным, особенно на мРНК с длинными и структурированными 5´UTR. В будущем будет важно различать активности eIF4AI и eIF4AII, поскольку в некоторых исследованиях предполагалось, что функции eIF4AI и eIF4AII не эквивалентны.
Письмо-решение после экспертной оценки:
Благодарим вас за отправку вашей статьи «Канал передачи мРНК в области ES6S трансляции 48S-PIC способствует раскручиванию и сканированию РНК» на рассмотрение eLife . Ваша статья была рецензирована тремя рецензентами, в том числе Наумом Соненбергом в качестве редактора-рецензента и рецензентом №1, а оценка проводилась под наблюдением редактора-рецензента и Джеймса Мэнли в качестве старшего редактора.
Рецензенты обсудили рецензии друг с другом, и редактор-рецензент подготовил это решение, чтобы помочь вам подготовить исправленную заявку.
Резюме:
Это интересная рукопись, в которой исследуется роль определенной топологической области рибосомы в инициации трансляции. Авторы выполнили эксперименты по вмешательству в область ES6S 48S-PIC и сообщили, что это избирательно блокирует трансляцию мРНК со структурированными 5 ‘UTRs. Они интерпретируют свои данные, чтобы указать, что мРНК, проникающие в эту область, сканируются медленнее и более процессно, чем мРНК, которые обходят эту область.В целом это хорошее исследование, но есть некоторые недостатки, включая отсутствие элементов управления и интерпретацию экспериментов. Рецензенты подняли много важных моментов, которые вы можете увидеть в полных обзорах. Перечисленные ниже проблемы являются наиболее важными.
Существенных изменений:
1) В соответствии с предыдущим исследованием, авторы показывают, что перекрестное связывание 4-thio-U идентифицирует взаимодействие между неструктурированной и вирусной мРНК с eIF3g и eIF4A. Однако обнаружено, что только eIF3g перекрестно сшивается с этой областью на неструктурированной мРНК, что ставит вопрос о том, насколько общим может быть положение eIF4A на невирусных мРНК.
2) Основным недостатком исследования является то, что авторы не показали напрямую, действительно ли конъюгат олиго-флуорофор, комплементарный ES6Sd и EGFP-eS4, ингибирует взаимодействие мРНК и области ES6S. Все интерпретации экспериментов предполагают это, но это должно быть непосредственно проверено с помощью эксперимента по сшиванию 4-тио-U в присутствии олиго и EGFP-eS4.
3) Авторы отслеживают скорость трансляции люциферазы в присутствии олиго и гиппуристанола на фиг. 7.Задержка появления белка LUC и синергетическое ингибирование интерпретируются как означающие, что скорость сканирования ниже. Это кажется сильным утверждением для анализа, который отслеживает только скорость трансляции люциферазы. Без дополнительных доказательств, которые более строго проверяют скорость «сканирования», авторы должны значительно смягчить интерпретацию этого эксперимента (то, что мРНК проникает в область ES6S, делает сканирование медленнее, но более процессным).
Доказательства в пользу «пронизывающей» модели мРНК в этом исследовании не очень убедительны.Утверждение о том, что eIF4A может функционировать в области ES6S для раскручивания дуплексных областей, следует квалифицировать, поскольку перекрестная связь eIF4A изучается только в присутствии вирусных мРНК.
4) Подходящим элементом управления для рисунка 3 — дополнение к рисунку 2B будет VIC-oligo C.
5) Приводит ли GFP-мечение другого рибосомного белка к тем же результатам, что и eS4-EGFP?
6) Рисунок 4A. Как мРНК, включая IRES HCV, реагируют на VIC-oligo C (контроль)? Этот отрицательный контроль отсутствует.Последовательности 5 ‘UTR репортеров, используемых здесь, показаны в виде компьютерной распечатки в дополнительном файле 1 несколькими грубыми способами без попытки сделать этот файл удобным для чтения — это следует исправить. Пожалуйста, подтвердите, что ни в одном из использованных репортеров нет исходных AUG, так как это может затруднить интерпретацию (при моем визуальном сканировании их не было).
7) Авторы не уточняют, является ли антитело, которое они использовали для IP в экспериментах по перекрестному связыванию, анти-4AI или 4AII.Этого не было сделано и в их предыдущей статье (Toribio et al., 2018). Поскольку функции eIF4AI и eIF4AII не эквивалентны, важно показать, какой eIF4A изучается в этой статье, и некоторое обсуждение различных функций 4AI и II будет очень информативным для читателя.
Рецензент № 1:
В статье дается четкое понимание механизма обратной корреляции между содержанием мРНК 5’UTR G + C / ΔG и эффективностью трансляции мРНК.Это обеспечивает дополнительную поддержку функции eIF4A по раскручиванию вторичной структуры мРНК 5 ‘для облегчения рекрутирования рибосом. Однако удивительно, что авторы не использовали свою превосходную экспериментальную систему, чтобы различать активности eIF4AI и eIF4AII. Фактически, они не уточняют, является ли антитело, которое они использовали для IP в экспериментах по перекрестному связыванию, анти-4AI или 4AII. Этого не было сделано и в их предыдущей статье (Toribio et al., 2018). Поскольку функции eIF4AI и eIF4AII не эквивалентны, важно показать, какой eIF4A изучается в этой статье, и некоторое обсуждение различных функций 4AI и II будет очень информативным для читателя.
Рисунок 4B; Не сразу понятно, почему на трансляцию РНК EMCV и полиовируса не влияет VIC-Oligo4, поскольку трансляция обоих зависит от eIF4AI.
Рисунок 5A; Профиль рибосомы в области полисом выглядит ненормальным. Эксперимент, должно быть, повторялся, но я не мог найти, сколько раз повторялся этот эксперимент (и несколько других). Могут ли они показать другой профиль?
Рецензент № 2:
В этой рукописи Ventoso и его коллеги проводят эксперименты по вмешательству в область ES6S 48S-PIC и сообщают, что это избирательно блокирует трансляцию мРНК со структурированными 5 ‘UTRs.Они интерпретируют свои данные, чтобы указать, что мРНК, проникающие в эту область, сканируются медленнее и более процессно, чем мРНК, которые обходят эту область. Это интересная рукопись, в которой исследуется роль определенной топологической области рибосомы в инициации трансляции.
Существенных изменений:
— Я полагаю, что подходящим элементом управления для рисунка 3 — дополнения к рисунку 2B будет VIC-oligo C.
— Приводит ли GFP-мечение другого рибосомального белка к тем же результатам, что и eS4-EGFP?
— Разве не должно быть контроля длины для SL20 на рисунке 4A? Это также хорошая возможность для авторов проверить, как систематическое увеличение структуры отслеживает эффективность трансляции, используя этот анализ.Кроме того, здесь следует более систематически анализировать влияние длины на эффективность. В настоящее время авторы не в полной мере использовали возможности этого анализа для усиления своей аргументации.
— Рисунок 4A. Как эти мРНК, включая IRES HCV, реагируют на VIC-oligo C (контроль)? Этот отрицательный контроль отсутствует. Последовательности 5 ‘UTR репортеров, используемых здесь, показаны как некоторая компьютерная распечатка в дополнительном файле 1 в некоторой грубой форме, без каких-либо попыток сделать это удобным для чтения — это следует исправить.Пожалуйста, подтвердите, что ни в одном из использованных репортеров нет исходных AUG, так как это может затруднить интерпретацию (при моем визуальном сканировании их не было).
— Рисунок 4B — SFV. Панель установлена неправильно. Дорожка 4, кажется, была преждевременно обрезана справа, и верхняя панель не совмещена должным образом с нижним контролем, окрашенным этидием.
Рисунок 5E — Стандартизированы ли эти значения для внутреннего контроля? Как эти репортеры реагируют на VCI-oligo C (отрицательный контроль).
— Я уверен, что авторы знают, что вестерн-блоттинг, показанный на рисунке 6, не сообщает о трансляционном ответе. Пожалуйста, покажите распределение тестовых мРНК по градиентам полисома или выполните метаболическую метку с последующим IP, чтобы сообщить о продукции зарождающегося белка.
— Подраздел «Полногеномный анализ показывает роль области ES6S в трансляции мРНК с G-богатыми 5´UTR». Исследования Rubio и Wolfe с использованием рокаглатов, вероятно, не следует здесь цитировать, поскольку они вызывают закрепление 4A на РНК, следовательно, они не приводят только к потере активности 4A, поскольку сами зажатые комплексы могут мешать трансляции (см. PMID 27309803).
— Все ли конструкции дают одинаковые конечные уровни люциферазы в анализе FTT? Почему площадь под кривыми на рис. 7c между двумя условиями сильно различается? Отражает ли это большее / меньшее количество транслируемых мРНК? Где находится контроль FITC-oligo C?
— Я не уверен, как интерпретировать рисунок 7D. Авторы указывают, что они используют субоптимальные концентрации VIC-oligo-4 для индукции умеренного блока трансляции (в 2-3 раза). Поскольку для получения эффекта VIC-oligo4 вам нужен только один олиго, связанный с рибосомной мишенью, это означает, что существует пул рибосом, которые также не связаны этим олиго.Я не уверен, что именно означает «синергия, полученная с помощью hipp — означает ли это синергетический эффект с теми рибосомами, которые имеют связанный VIC-oligo-4, или это означает, что hipp действует на те рибосомы, которые не имеют связанного VIC-oligo 4, и оказывает эффект, не связанный с областью ES6S, и это «оценивается» как синергетический эффект. Где контроли с VIC-oligo C.
— Авторы называют синтетическую 128-нуклеотидную мРНК «плоской». Это несколько разговорный язык, и авторы могут захотеть изменить его на более приемлемый с научной точки зрения ярлык.
— Подраздел «Путь мРНК через область ES6S 48S PIC». Авторы пишут: «Согласно нашим данным, eIF4A может быть расположен немного дальше по потоку, вероятно, между спиралями ES6SA 179 и ES6SB 180 (Рисунок 2C)». Я не вижу на рисунке 2c, где указаны сайты ES6SA и ES6SB.
Рецензент № 3:
Авторы сделали два основных открытия, которые потенциально могут изменить наше понимание рекрутирования мРНК.
Во-первых, авт. Показали, что перекрестное связывание 4-thio-U области мРНК от +24 до +34 происходит вокруг области ES6S субъединицы 40S (внутри 48S PIC).Это достигается с помощью короткой неструктурированной мРНК и GMP-PNP. В соответствии с этим неспецифическое УФ-сшивание также выявляет взаимодействия между мРНК и eS4 / uS4, что в целом согласуется с областью взаимодействия ES6S. В соответствии с предыдущим исследованием авторы показывают, что перекрестное связывание 4-thio-U идентифицирует взаимодействие между неструктурированной и вирусной мРНК с eIF3g и eIF4A. Это перекрестное связывание происходит в той же области мРНК, которая перекрестно связывается с областью ES6S, что позволяет предположить, что eIF3g и eIF4A расположены в этой области в 48S PIC.Однако обнаружено, что только eIF3g перекрестно сшивается с этой областью на неструктурированной мРНК, что ставит вопрос о том, насколько общим может быть положение eIF4A на невирусных мРНК.
Второе важное открытие заключается в том, что нарушение области ES6S либо конъюгатом олиго-флуорофор, который комплементарен ES6Sd, либо слитым с EGFP белком eS4 (расположенный рядом с ES6S), приводит к снижению скорости трансляции. Это показано с использованием репортерных мРНК, а также в масштабе всего генома. Интересно, что ингибирование трансляции очевидно на мРНК, несущих высокоструктурированные и / или длинные 5’UTR (включая G-квадруплекс, содержащий 5’UTR).Специфичность ингибирования контролируется с помощью использования того же олиго без флуорофора или олиго, комплементарного другому участку рРНК.
Основная слабость исследования состоит в том, что авторы не показали прямо, действительно ли олигонуклеотид и EGFP-eS4 ингибируют взаимодействие мРНК и области ES6S. Все интерпретации экспериментов предполагают это, но это должно быть непосредственно проверено с помощью эксперимента по сшиванию 4-тио-U в присутствии олиго и EGFP-eS4.
Авторы отслеживают скорость трансляции люциферазы в присутствии олиго и гиппуристанола на фиг. 7. Задержка появления белка LUC и синергетическое ингибирование интерпретируются как означающие, что скорость сканирования ниже. Это кажется сильным утверждением для анализа, который отслеживает только скорость трансляции люциферазы. Без дополнительных доказательств, которые более строго проверяют скорость «сканирования», авторы должны значительно смягчить интерпретацию этого эксперимента (то, что мРНК проникает в область ES6S, делает сканирование медленнее, но более процессным).
Этот рецензент действительно не видит убедительных доказательств в пользу «многопоточности» модели мРНК из этого исследования. Утверждение о том, что eIF4A может функционировать в области ES6S, раскручивая дуплексные области, также кажется чрезмерно интерпретированным — насколько я могу судить, сшивка eIF4A, по-видимому, происходит только в присутствии вирусных мРНК.
В целом это интересное исследование, но некоторые серьезные недостатки в интерпретации экспериментов несколько снижают энтузиазм.
[Примечание редакции: до принятия были запрошены дополнительные исправления, как описано ниже.]
Благодарим вас за повторное представление вашей работы под названием «Канал связывания мРНК в области ES6S трансляции 48S-PIC способствует раскручиванию и сканированию РНК» для дальнейшего рассмотрения на eLife . Ваша исправленная статья была положительно оценена Джеймсом Мэнли (старшим редактором), редактором-рецензентом и одним рецензентом.
Рукопись была улучшена, но есть некоторые оставшиеся проблемы, которые необходимо решить до принятия, как указано ниже: Есть одна проблема, которая не была должным образом решена.Авторы заявляют в своем опровержении, что лабораторные антитела Святого Иоанна против eIF4AI, согласно данным компании.
«Антитело, используемое здесь и в предыдущих статьях для обнаружения eIF4A с помощью IP-экспериментов, было STJ27247 из лаборатории Сент-Джона. Согласно таблице данных, предоставленной поставщиком, антитело вырабатывалось у кроликов с использованием аминокислот 1-406 а.о. eIF4AI в качестве иммуноген, поэтому мы предполагаем, что он специфичен для eIF4AI.Одна полоса белка была обнаружена с помощью вестерн-блоттинга с использованием этого антитела, что позволяет предположить, что антитело не реагирует с eIF4AII.«
Однако авторам необходимо продемонстрировать, что антитело специфично к eIF4AI. Кроме того, эта информация не представлена в разделе материалов и методов, и они продолжают ссылаться на eIF4A на протяжении всей статьи.
Важно решить эту проблему, потому что существующая запутанная литература, в которой утверждается, что eIF4AII проявляет антагонистическую функцию по отношению к eIF4AI, сбивает с толку литературу. Авторы должны предупредить об этом читателей.
https://doi.org/10.7554/eLife.48246.sa1Какие виды судебной медицины?
Область применения судебной медицины широка: это больше, чем отпечатки пальцев и образцы ДНК. Для организации различных специальностей в этой области Американская академия судебных наук (AAFS) официально признает 11 различных дисциплин судебной медицины.
Магистр судебной медицины Национального университетаобеспечивает теоретическую и практическую подготовку в области криминалистики и методов расследования места преступления.Когда вы начинаете свою карьеру в качестве судебного эксперта, возможно, у вас еще нет определенной специальности. По мере того, как вы набираетесь опыта и продолжаете учиться на работе или проходя дополнительные курсы, вы можете приобрести опыт в одном из этих типов судебной медицины.
Антропология: реконструкция жизниАмериканский совет судебной антропологии (ABFA) определяет свою область как применение науки физической или биологической антропологии в судебном процессе.
Судебная антропология состоит из нескольких дисциплин: культурной антропологии, археологии, лингвистики и физической (биологической) антропологии. Вместе все эти области знаний могут, как отмечает AAFS в своем руководстве по карьере, «попытаться восстановить как можно больше о жизни и смерти человека». Для этого судебные антропологи исследуют останки скелетов, чтобы определить характеристики, причину смерти, травм или заболеваний, сколько времени прошло после смерти и другие выводы, которые могут привести к раскрытию преступления, идентификации неизвестной жертвы или даже помощи с раскопками. остается на раскопках археологии.
Судебные антропологи обычно имеют докторскую степень и обычно не работают в судебных экспертах на постоянной основе. Скорее, они могут работать в университете, музее, отделении вооруженных сил или медицинской организации и при необходимости предоставлять свои услуги местным криминалистическим лабораториям. Хотя для того, чтобы носить это звание, требуется ученая степень, технические специалисты могут также работать вместе со специалистами судебной антропологии, например, при сборе, подготовке, анализе и документировании образцов в рамках судебно-медицинского расследования.
Криминалистика: понимание доказательствКогда люди думают о том, чем занимается судебный эксперт каждый день, они, вероятно, представляют себе задачи, связанные с криминалистикой, такие как судебная баллистика. Многие предметы собраны с места преступления, но не все актуальны. Вот тут и появляются криминалисты.
Чаще всего эти специалисты размещаются в судебно-медицинских лабораториях и преследуют две основные цели: идентифицировать улики и связывать людей, объекты и места с помощью этих доказательств.Криминалисты иногда специализируются на конкретных областях вещественных доказательств, некоторые из которых требуют дополнительной подготовки, в том числе:
- Огнестрельное оружие (судебная баллистика).
- Метки инструментов.
- ДНК.
- Осколки пожара и взрыва.
- Контролируемые вещества.
- Следы.
- Дикая природа.
Эксперты в области криминалистики могут работать в различных организациях, включая государственные и федеральные криминалистические лаборатории, полицейские управления, федеральные агентства (например, ФБР, DEA или ATF), офисы медицинских экспертов и даже U.S. Почтовая служба.
Магистр судебных наук в Национальном университете предлагает специализацию по криминалистике, которая включает курсы по следовым доказательствам, передовой судебной токсикологии, передовой судебной серологии и судебной антропологии.
Сегодня правоохранительные органы и лаборатории занимаются преступлениями (и, следовательно, доказательствами), которых не существовало несколько десятилетий назад. Это означает, что область судебной экспертизы будет продолжать развиваться по мере изменения технологий, а область цифровых и мультимедийных наук является одной из таких постоянно меняющихся областей.
Согласно AAFS, судебные эксперты в этой области исследуют аппаратные средства, программные приложения и цифровые файлы (аудио, текст, изображения, видео и т. Д.), Чтобы найти и проанализировать доказательства. Ключевым словом здесь является «найти», потому что часто есть терабайты данных, часы видеоматериалов (скажем, с камер безопасности или дорожных камер), которые нужно отсортировать, прежде чем найти что-либо значимое или актуальное.
Вот лишь несколько примеров обязанностей специалиста по цифровой криминалистике:
- Определение того, было ли изменено цифровое изображение.
- Анализ акустики записи.
- Определение устройств, подключенных к системе.
- Определение, были ли файлы удалены с диска или устройства.
- Поиск удаленной системы или пользователя.
- Поиск жертвы или подозреваемого по данным.
Хорошая программа судебной медицины обеспечит обучение в этой области. Например, курс «Цифровые доказательства» Национального университета охватывает ряд процедур и методов цифрового расследования, связанных с такими преступлениями, как мошенничество, преследование и кража личных данных.
Как и другие виды криминалистики, существует профессиональная организация для специалистов по цифровым технологиям: Международная ассоциация специалистов по компьютерным расследованиям.
Инженерные и прикладные науки: стихийные бедствия и техногенные катастрофыAAFS утверждает, что эта дисциплина является домом для самой разнообразной группы судебно-медицинских специалистов: специалистов по решению проблем, имеющих опыт работы в одной из многих суб-дисциплин инженерии, химии или физики.Эти эксперты вызываются как по уголовным, так и по гражданским делам для расследования таких вещей, как:
- Автомобильные столкновения.
- Обрушивается здание.
- Крушение поезда.
- Взрывы.
- Неисправности продукта.
- Загрязнение окружающей среды.
Этот район идеально подходит для тех, кто имеет солидный опыт в науке и интересуется расследованием места преступления. Фактически, многие люди поступают в онлайн-программу магистратуры судебной экспертизы после получения степени бакалавра в области химии, биохимии или инженерии.Также полезно понимание математики, биологии и геологии.
Общее: Разнообразие судебной экспертизыAAFS использует общую категорию для проведения специальных исследований в области судебно-медицинской экспертизы, полевых исследований, клинической работы, а также образования и исследований, которые не подходят для других более крупных дисциплин. Они включают в себя диапазон фокусировки, в том числе:
- Бухгалтерский учет.
- Ветеринарные услуги.
- Сестринское дело.
- Консультации.
- Искусство и скульптура.
- Радиология.
- Менеджмент и администрирование.
В эту категорию также входят новые области, такие как судебная ветеринария и судебная медицина. Иногда специальная область разрастается настолько, чтобы стать отдельной дисциплиной, например, цифровые и мультимедийные науки; в других случаях больше нишевых специальностей останется в списке общих.
(Поскольку возможности судебной медицины настолько широки, этот пост «Каковы различные типы степеней расследования места преступления?» Может быть вам полезен.)
Юриспруденция: судебно-медицинская экспертиза«Юриспруденция» — это не то слово, которое вы часто слышите в повседневной беседе. Короче говоря, это философия права. Члены, практикующие эту дисциплину судебной медицины, также должны иметь лицензию на занятие юридической практикой.
Любого судебно-медицинского эксперта, от ДНК-аналитиков до антропологов, можно попросить дать показания по уголовным и гражданским делам; однако, что, если бы были юристы, которые также обладают опытом в области основ судебной медицины и расследований? Есть.Эти юристы, которых иногда называют «судебно-медицинскими экспертами», также прошли подготовку по проведению расследований на месте преступления. Они работают независимо или вместе с другими юристами для сопровождения судебных дел. Это может включать сбор и изучение доказательств для того, чтобы делать выводы, консультировать клиентов и формировать судебные стратегии.
Юристы с экспертными знаниями в области судебной медицины могут работать в юридических фирмах, судебных системах, федеральных агентствах, некоммерческих организациях и даже выступать в качестве частных консультантов.В качестве примера, Brooklyn Defender Services в 2019 году разместила вакансию «специального судебно-медицинского консультанта: отдел по расследованию убийств / серьезных уголовных преступлений».
Одонтология: поиск ответов зуб за зубОдонтология, более известная как судебная стоматология, может сыграть важную роль в расследовании преступлений, а также в реагировании на стихийные бедствия и восстановлении. Подобно судебным антропологам, эти специалисты помогают в идентификации человеческих останков. И, как и в области юриспруденции, одонтолог должен иметь профессиональную степень: в данном случае доктор стоматологической медицины, доктор стоматологической хирургии или эквивалентную степень.
Зубной эксперт может внести свой вклад в судебно-медицинское расследование разными способами, например:
- Сравнение стоматологических останков со стоматологическими записями.
- Сравнение стоматологических карт.
- Оценка следов укусов.
- Старение человека.
- Оценка стоматологических или других травм полости рта.
Американское общество судебной одонтологии предлагает обширную информацию об этой стоматологической специальности.
Патология / биология: криминалистика вплоть до наукиAAFS объединяет судебную биологию и судебную патологию в одну дисциплину, но каждая область имеет свои собственные направления.Патология — это исследование болезни, и судебно-медицинский патологоанатом использует эти навыки и знания для оказания помощи в юридических вопросах. Иногда это включает в себя вскрытие, которое позволяет патологу исследовать органы, ткани и жидкости, чтобы найти причину или обстоятельства смерти. В других случаях судебно-медицинский патологоанатом может работать в лаборатории, изучая более мелкие образцы.
Патологоанатомы, как правило, врачи, однако специалисты-криминалисты могут помочь в сборе и анализе этих медицинских доказательств.Магистр судебных наук Национального университета обеспечивает основу для этого на курсах «Судебная патология I и II».
Эксперты-криминалисты изучают организмы или клетки организмов, которые могут иметь отношение к преступлению. Эта область охватывает науки о жизни: энтомологию (насекомые), ботанику (растения), экологию, генетику и микробиологию. Примеры того, как эти специальности могут помочь в расследовании смерти, согласно AAFS, могут включать:
- Ученый-ботаник, определяющий содержимое желудка или находящий скрытые могилы.
- Ветеринар, обслуживающий животных.
- Энтомолог, помогающий определить временную шкалу (например, когда насекомые могут захватить человеческие останки).
- Генетик, помогающий подтвердить личность организма.
AAFS утверждает, что у большинства судебных биологов есть степень магистра или доктора философии. в области биологических наук и обычно работают в музеях или университетах.
Психиатрия и бихевиоризм: понимание природы человекаПсихиатры, психологи и другие специалисты по поведенческой науке могут внести свой вклад в область судебной экспертизы, оценивая людей или давая показания по уголовным или гражданским делам.
Психиатры — это врачи, которые, вероятно, прошли дополнительную подготовку по вопросам того, как их область знаний связана с законом. Фактически, некоторые выбирают программы медицинской ординатуры специально для этого типа обучения. Они могут помочь определить, годен ли кто-то для судебного разбирательства или оценки психического заболевания в связи с преступлением. Судебные психиатры также могут помочь в случаях домашнего насилия, жестокого обращения с детьми, усыновления, опеки и попечительства.
Когда вы думаете о судебной психологии, на ум может прийти криминальное профилирование.Это определенно одна из областей, в которой психологи могут помочь специалистам правоохранительных органов; однако они могут использовать свое специализированное обучение для гораздо большего.
Запрошенные документы: не только толкатели бумагиЭта отрасль судебной экспертизы фокусируется на изучении физических документов и включает в себя гораздо больше, чем анализ почерка. Этот опыт, согласно AAFS, может быть использован для:
- Ищите изменения в документе.
- Расшифровать стертые записи.
- Восстановите сгоревшие или поврежденные документы.
- Классифицируйте и идентифицируйте компьютерные принтеры и пишущие машинки.
- Выявить авторство подписи или другого письма.
Эксперты по опрошенным документам обычно имеют ученую степень в области судебной медицины, а затем участвуют в структурированной двухлетней программе обучения без отрыва от производства.
Токсикология: вещества и законТоксикология — это изучение неблагоприятного воздействия химических веществ (например, наркотиков, алкоголя) на организм человека.Таким образом, судебная токсикология рассматривает эти эффекты в контексте закона. Они анализируют различные образцы, включая кровь, мочу, ткани, волосы и жидкости (например, из глаза, печени или мозга), а затем интерпретируют результаты. В нашем сообщении в блоге «Что такое судебная серология» содержится более подробная информация о работе с биологическими образцами.
Судебно-токсикологи специализируются на одном из трех основных направлений:
- Посмертная токсикология: помогает определить причину или способ смерти.
- Токсикология возможностей человека: для определения нарушений во время преступления.
- Судебно-медицинская экспертиза: используется на рабочем месте, при занятиях спортом и на условно-досрочном освобождении.
Судебные токсикологи обычно приступают к работе со степенью бакалавра химии или наук о жизни. Программы бакалавриата и магистратуры по судебной медицине часто представляют собой введение в токсикологию. Лаборатория судебной экспертизы также часто предлагает дополнительное обучение на рабочем месте в этой области.
Узнайте больше о видах судебной медициныЭто 11 дисциплин судебной медицины, определенных AAFS для своих профессиональных членов.На практике вы обнаружите, что организационная структура — отделы, подразделения, должности, обязанности, политики и процедуры — варьируются от места к месту. Фактически, если вы рассмотрите множество видов судебной медицины, легко увидеть, насколько важным становится сотрудничество и как иногда роли пересекаются.
Судебная медицина, безусловно, представляет собой разнообразную область, в которой сочетаются наука, человеческое поведение, закон и правосудие. Магистр судебных наук Национального университета, одна из многих удобных онлайн-программ, представляет собой надежную программу, которая хорошо подготовит вас к различным возможностям в этой захватывающей области.Посетите страницу нашей программы, чтобы узнать больше или запросить информацию.
Расшифровка сигнатурной модели рекрутирования транскрипционного кофактора, продиктованной кардинальными цис-регуляторными элементами в проксимальных промоторных областях
Расшифровка сигнатурной модели рекрутирования транскрипционного кофактора, продиктованной кардинальными цис-регуляторными элементами в проксимальных промоторных областях МетаPLoS Genetics
Christopher BennerIvan Garcia-Bassets
Abstract
Полногеномные карты гиперчувствительных сайтов (DHS) к ДНКазе I показывают, что большинство промоторов человека содержат постоянно активные цис-регуляторные элементы от -150 до +50 пар оснований ( -150 / + 50 п.н.) относительно сайта начала транскрипции (TSS).Факторы транскрипции (TF) привлекают кофакторы (ремоделирующие хроматин, гистоны / белковые модифицирующие ферменты и скаффолд-белки) к этим элементам, чтобы организовать локальную структуру хроматина и координировать баланс посттрансляционных модификаций поблизости, способствуя общей регуляции транскрипция. Однако правила TF-опосредованного рекрутирования кофакторов в промоторные области -150 / + 50 п.н. остаются плохо изученными. Здесь мы приводим доказательства общей модели, в которой ряд цис-регуляторных элементов (здесь называемых « кардинальными » мотивами) предпочитают действовать индивидуально, а не в фиксированных комбинациях, в областях -150 / + 50 п.н. для рекрутирования TF, которые диктуют сигнатуры кофакторов, отличительные от конкретных подмножеств промоторов.Впоследствии промоторы человека можно разделить на подклассы на основе присутствия кардинальных элементов и связанных с ними сигнатур кофакторов. Кроме того, в этом исследовании мы сосредоточили внимание на промоторах, содержащих мотив ядерного респираторного фактора 1 (NRF1) в качестве кардинального цис -… Продолжить чтение
Ссылки
11 мая 1991 г. · Исследования нуклеиновых кислот · NC Andrews, Д.В. Фаллер
23 октября 1986 · Природа · OR Choi, JD Engel
1 января 1988 · Годовой обзор биохимии · DS Gross, WT Garrard
1 декабря 1995 · Журнал вирусологии · Q Zhou, DA Engel
8 апреля 1994 г. · Cell · R Tjian, T Maniatis
19 декабря 1996 г. · Природа · Эндрю Дж. Баннистер, Тони Кузаридес
16 апреля 1998 г. · Наука · CH YuhE H Davidson
24 августа 1999 г. · Нуклеиновые кислоты Исследования · I García-BassetsF Azorín
5 ноября 1999 г. · Журнал молекулярной биологии · H RothenederE Haidweger
28 ноября 2002 г. · Молекулярная клетка · Тацуя Накамура Эли Канаани
15 января 2003 г. · BMC Bioinformatics, Christopher D Bader WV Hogue
25 ноября 2003 г. · Исследование нуклеиновых кислот · Патрик Дж. Дэниэлс , Глен К. Эндрюс
3 декабря 2003 г. · Журнал биологической химии · Кристоф Крампс Вернер Лутц
4 июня 2004 г. · Молекулярная генетика человека · Карен Т. СмитДэниел Рейнс
17 июля 2004 г. · Исследование генома · Питер Фицджеральд Чарльз Винсон
29 декабря 2004 г. · Ячейка · Ю-Цзян Шиян Ши
1 марта 2005 г. · Природа · Сяохуэй СеМанолис Келлис
9 июня 2005 г. · Труды Национальной академии наук Соединенных Штатов Америки · Мэтью Дж. ГюнтерРичард А. Янг
13 июля 2005 г. · Ячейка · Джейсон С. КэрроллМайлс Браун
5 августа 2005 г. · Природа · Мин Гю Ли Рамин Шихаттар
6 сентября 2005 г. · Молекулярная клетка · Ю-Цзян Ши Ян Ши
15 октября 2005 г. · The Журнал биологической химии · Федерико Форнерис, Андреа Маттеви,
,, 14 декабря 2005 г. · Исследование генома · Сара Дж. Купер, Ричард М. Майерс,
,, 12 апреля 2006 г. 90 008
21 сентября 2006 · Nature Reviews.Генетика · Роберт Дж. Клозей Чжан
23 сентября 2006 г. · Журнал биологической химии · Эстель Николас Рамин Шихаттар
16 декабря 2006 г. · Клеточные и молекулярные науки о жизни: CMLS · Натаниэль Д. Хайнцман, Бинг Рен
21 декабря 2006 г. · Исследования генома · Пол Джи Гиреси Джейсон Д. Либ
16 января 2007 · Природные биотехнологии · Фатих Озсолак Дэвид Э. Фишер
6 февраля 2007 · Природная генетика · Натаниэль Д. Хайнцман Бинг Рен
10 февраля 2007 г. Розенфельд
16 марта 2007 г. · Труды Национальной академии наук Соединенных Штатов Америки · Янг-Су Квон Сян-Донг Фу
30 марта 2007 г. · Природа · Ван ЦзяньсуньМайкл Г. Розенфельд
31 марта 2007 г. · Журнал нейрохимии · Грегори Капатос Яннинг Ву
9 мая 2007 г. · Nature Reviews.Генетика · Альбин СанделинДэвид Эй Хьюм
22 мая 2007 г. · Клетка · Артем Барски Кеджи Чжао
15 июня 2007 г. · Геномные исследования · Хуалин Сижипин Вэн
21 августа 2007 г. · Молекулярная клетка · Ширин СалекСтюарт Оркин
2007 · PLoS Genetics · Hualin XiGregory E Crawford30 августа 2007 г. · PloS One · Yuval TabachEytan Domany
20 октября 2007 г. · Природные протоколы · Мелисса С. КлайнГэри Д. Бадер
1 декабря 2007 г. · Международный журнал исследований в области биоинформатики и Приложения · Билл Андреопулос Сяоганг Ван
25 марта 2008 г. · Cell · Матье Люпен Майлс Браун
26 апреля 2008 г. · Текущее мнение в области клеточной биологии · Тамар Ювен-Гершон Джеймс Т. Кадонага
29 апреля 2008 г. · Текущее мнение в области клеточной биологии · Фей Ланьянь Ши
11 июля 2008 · Ячейка · Дэвид Дж. ПаглиариниВамси К. Мутха
25 сентября 2008 · Биоинформатика · Цзин Гао Давид Дж. Стейтс
17 декабря 2008 · Анналы Нью-Йоркской академии наук · RC Scarpulla
апр. 3, 2009 · Журнал EMBO · Виллем-Ян Велборен Хендрик Дж. Стунненберг
2 июня 2009 г. · Исследования нуклеиновых кислот · Кен Дайгоро Йокояма Грегори Эй Врей
18 августа 2009 г. · Биология развития · Тамар Ювен-Гершон, Джеймс Т. Кадонага
августа , 2009 · Ячейка · Чжибин Ван Кеджи Чжао26 сентября 2009 г. · Журнал EMBO · Франческа Де Санта-Джоаккино Натоли
27 октября 2009 г. · Nature Medicine · Ю Лян Томас М. Кристи
13 января 2010 г. · Ячейка · Джейми С. Пэн Джоанна Высока
16 января 2010 г. · Природа · Диего ПазиниКристиан Хелин
4 февраля 2010 г. · Гены и развитие · Банда ЛиДэнни Рейнберг
20 марта 2010 г. · Молекулярная биотехнология · Мартина Бауманн Вольфганг Эрнст
16 апреля 2010 г. · Природа · Tae- Кюнг КимМайкл Э. Гринберг
21 апреля 2010 · Клеточный цикл · Лата Балакришнан Барри Милавец
27 апреля 2010 · Молекулярная клетка · Нил Блэкледж Роберт Дж. Клозе
8 мая 2010 · Отчеты EMBO · Sujit S NairRatna 9000 K Vadlami 27, 2010 · Gen ome Research · Джейн М. Ландолин, Ричард М. Майерс,
,, 2 июня 2010 г. · Молекулярная клетка, · Свен Хайнц, Кристофер К, Гласс,
,, 29 июня 2010 г. И Чанг
14 июля 2010 г. · Природа · Вэнь Лю Майкл Дж. Розенфельд
27 июля 2010 г. · Журнал биологической химии · Мелина М. МусриМарселина Парризас
16 ноября 2010 г. · Клетка развития · Майкл П. ВерзиРамеш А. Шивдасани
декабря 3, 2010 · Исследования нуклеиновых кислот · Кристофер П. Чайлдерс · Кристин Дж. Элсик21 декабря 2010 · PLoS Genetics · Эрик М. Менденхолл, Брэдли Э. Бернштейн
3 февраля 2011 г. 2011 г. · Текущее мнение в области генетики и развития · Цзянь Ли, Дэвид С. Гилмор
17 мая 2011 г. · Клетка развития · Марлоуз Р. ТейссенБертольд Гёттгенс
17 мая 2011 г. · Природа · Донг Ван Сян-Донг Фу
18 мая 2011 г. · Гены и Dev побег · Эме М. Дитон, Адриан Бёрд
24 мая 2011 · Природа клеточной биологии · Антонио Адамо Хуан Карлос Изписуа Бельмонте
25 октября 2011 · Журнал EMBO · Сандра У ШмитцКристиан Хелин
5 ноября 2011 · Критические обзоры в биохимии и молекулярная биология · Дилетта ДолфиниРоберто Мантовани
8 ноября 2011 г. · Гены и развитие · Кеннет С. Зарет, Джейсон С. Кэрролл
27 декабря 2011 г. · Клетка · Орен Рэм Брэдли Э. Бернштейн
3 февраля 2012 г. · Природа · Уоррен Эй Уайт Ричард A Young
7 февраля 2012 г. · Biochimica Et Biophysica Acta · Джаяша Шандилья, Стефан Г.Е. Робертс
17 марта 2012 г. · Протоколы природы · Надежда Т. 3, 2012 · Текущее мнение в области клеточной биологии · Мануэль Иримиа, Бенджамин Дж. Бленкоу
5 апреля 2012 г. · Nature Reviews.Генетика · Эрик Л. Грир, Ян Ши
5 апреля 2012 г. · Nature Reviews. Молекулярная клеточная биология · Сюзанна Мариье Куистра, Кристиан Хелин
18 апреля 2012 г. · Геномные исследования · Housheng Hansen HeX Ширли Лю
24 апреля 2012 г. · Молекулярная клетка · Сунил К. РагхавБарт Депланке
23 июня 2012 г. · Природа · Мэтью Ф. BarberKatrin F Chua
17 августа 2012 г. · Клеточный цикл · Иван Гарсия-Бассетс, Донг Ван
8 сентября 2012 г. · Природа · Роберт Э. Турман, Джон А. Стаматояннопулос
8 сентября 2012 г. · Природа · Шейн Неф Джон А. 8, 2012 · Наука · Мэтью Т. Морано, Джон А. Стаматояннопулос
3 октября 2012 г. · Молекулярная и клеточная биология · Венугопалан Д. НаирСтюарт С. Силфон
16 ноября 2012 г. · Природа · Скотт Б. Вафаи, Вамси К. · Молекулярная клетка · Джошуа К. Блэк · Джонатан Р. Ветстин
22 января 2013 · Клетка · Марко НардиниРоберто Мантовани
4 июня 2013 · Природа · Венбо ЛиМайкл Дж. Розенфельд
19 октября 2013 г. mKazuya Yamagata
17 февраля 2015 г. · Научные доклады · Hebing ChenWenjie Shu
Цитаты
11 июня 2014 г. · Nature Communications · Delphine DuteilRoland Schüle
21 февраля 2016 г. 25, 2015 · Молекулярная нейробиология · Евгения Ю. Попова · Колин Дж. Барнстейбл
26 мая 2017 · Исследование нуклеиновых кислот · Мелисса Д. Илсли, Эндрю С. Перкинс
4 апреля 2019 г. · PloS One · Шулан Тиансусан Л. Слагер
21 февраля 2019 г. Фармакология · Ruohong ChenYiqun Deng
13 февраля 2018 г. · Письма об онкологии · Wei GaoYajie Wang
1 октября 2020 г. · Обращение.Сердечная недостаточность · Энтони М. Гасита · Элизабет М. МакНалли
11 марта 2017 · Письма FEBS · Макико Ниси-Тацуми Хитоши Шимано
9 ноября 2017 · Журнал клеточной физиологии · Си Чен Сяолин Лю
27 июля 2018 · Клеточная онкология (Дордре 27 июля 2018 г.) · Каумуди Бхаве, Деодутта Рой
22 августа 2018 г. · Исследования и лечение рака груди · Хайро РамосДеодутта Рой
15 августа 2019 г. · Онкоген · Синь Ху Чжэ Ли
4 июля 2018 г. · Научные доклады · Лин Чжан Джин-Марк Ванакер
20 ноября, 2019 · Nature Genetics · Ангелы Альменар-КеральтИван Гарсиа-Бассетс
30 марта 2017 · Труды Национальной академии наук Соединенных Штатов Америки · Джули Карнесекки Жан-Марк Ванакер
11 октября 2017 · Научный Отчеты · Йоко ТакиучиАкифуми Такаори-Кондо
30 ноября 2018 г. · Клетки · Джаянта К. ДасДеодутта Рой
6 октября 2020 г. · Границы физиологии · Кодзи Такемура Рэйко Инаги
18 января 2021 г. · Исследование биологического следа BeltagyOla H El-Habit
6 февраля 2021 г. · Международный журнал молекулярных наук · Джозеф М. Коллинз, Дансин Ван
23 апреля 2020 г. · Отчеты по клеткам · Ци Майван Гарсия-Бассет
3 апреля 2021 г. Gymrek
6 мая 2021 г. · Cell Reports Medicine · Мел Лина Пинсах-Абуин Сара Паганс
ChIP-seq
MNase-seq
иммунопреципитация
ChIP
Коиммунопреципитация
000 Коиммунопреципитация
000 Хроматография
000 -seq
иммунопреципитации
ChIPs
Родственные концепции
Белок NRF1, человеческий
Хроматин
Стрептодорназа
Ранние промоторы, Генетические
Генетические нуклеотидные последовательности
Генетические нуклеотидные последовательности
Регуляторный фактор
Сайт инициации транскрипции
Ремоделирование хроматина
Ядерная респираторная система Фактор 1
Связанные статьи
Cell
Renato OstuniGioacchino Natoli
Genome Biology
Yonatan Bilu, Naama Barkai
Plant & Cell Physiology
Moichi Kodima LomaxGenome Informatics
Алексис Ванденбон, Кента Накаи
/ документы / модель транскрипции на основе декодирования сигнатур / 24244184
.