
2. Энергетический обмен (катаболизм, диссимиляция)
Универсальным источником энергии во всех клетках служит АТФ (аденозинтрифосфат, или аденозинтрифосфорная кислота).
Все энергетические затраты любой клетки обеспечиваются за счёт универсального энергетического вещества — АТФ.
АТФ синтезируется в результате реакции фосфорилирования, то есть присоединения одного остатка фосфорной кислоты к молекуле АДФ (аденозиндифосфата):
АДФ + h4PO4+ 40 кДж = АТФ + h3O.
Энергия запасается в форме энергии химических связей АТФ. Химические связи АТФ, при разрыве которых выделяется много энергии, называются макроэргическими.
При распаде АТФ до АДФ клетка за счёт разрыва макроэргической связи получит приблизительно \(40\) кДж энергии.
Энергия для синтеза АТФ из АДФ выделяется в процессе диссимиляции.
Энергетический обмен (диссимиляция, катаболизм) — это совокупность химических реакций постепенного распада органических соединений, сопровождающихся высвобождением энергии, часть которой расходуется на синтез АТФ.
В зависимости от среды обитания организма, диссимиляция может проходить в два или в три этапа.
Процессы расщепления органических соединений у аэробных организмов происходят в три этапа: подготовительный, бескислородный и кислородный.
В результате этого органические вещества распадаются до простейших неорганических соединений.
У анаэробных организмов, обитающих в бескислородной среде и не нуждающихся в кислороде (а также у аэробных организмов при недостатке кислорода), диссимиляция происходит в два этапа: подготовительный и бескислородный.
В двухэтапном энергетическом обмене энергии запасается гораздо меньше, чем в трёхэтапном.
Первый этап — подготовительный
Подготовительный этап заключается в распаде крупных органических молекул до более простых: полисахаридов — до моносахаридов, липидов — до глицерина и жирных кислот, белков — до аминокислот.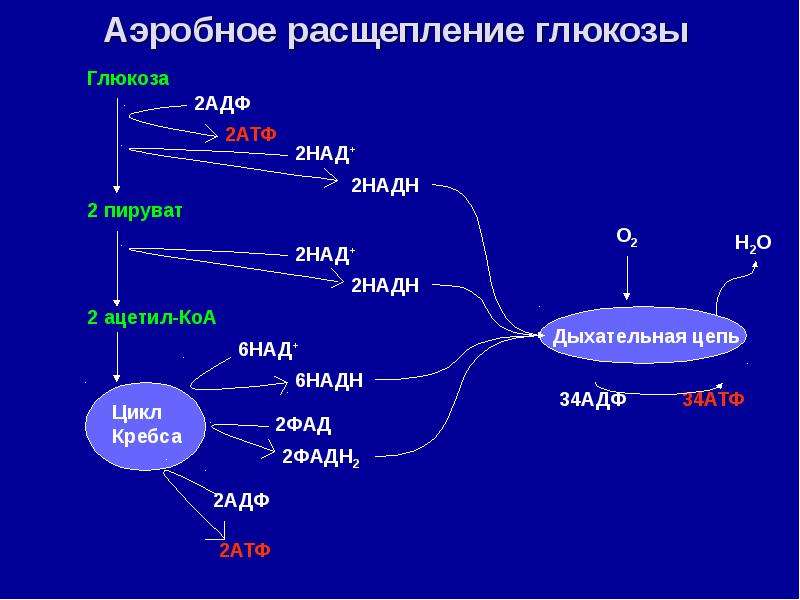
Этот процесс называется пищеварением. У многоклеточных организмов он осуществляется в желудочно-кишечном тракте с помощью пищеварительных ферментов. У одноклеточных организмов — происходит под действием ферментов лизосом.
В ходе биохимических реакций, происходящих на этом этапе, энергии выделяется мало, она рассеивается в виде тепла, и АТФ не образуется.
Второй этап — бескислородный (гликолиз)
Второй (бескислородный) этап заключается в ферментативном расщеплении органических веществ, которые были получены в ходе подготовительного этапа. Кислород в реакциях этого этапа не участвует.
Биологический смысл второго этапа заключается в начале постепенного расщепления и окисления глюкозы с накоплением энергии в виде \(2\) молекул АТФ.
Процесс бескислородного расщепления глюкозы называется гликолиз.
Гликолиз происходит в цитоплазме клеток.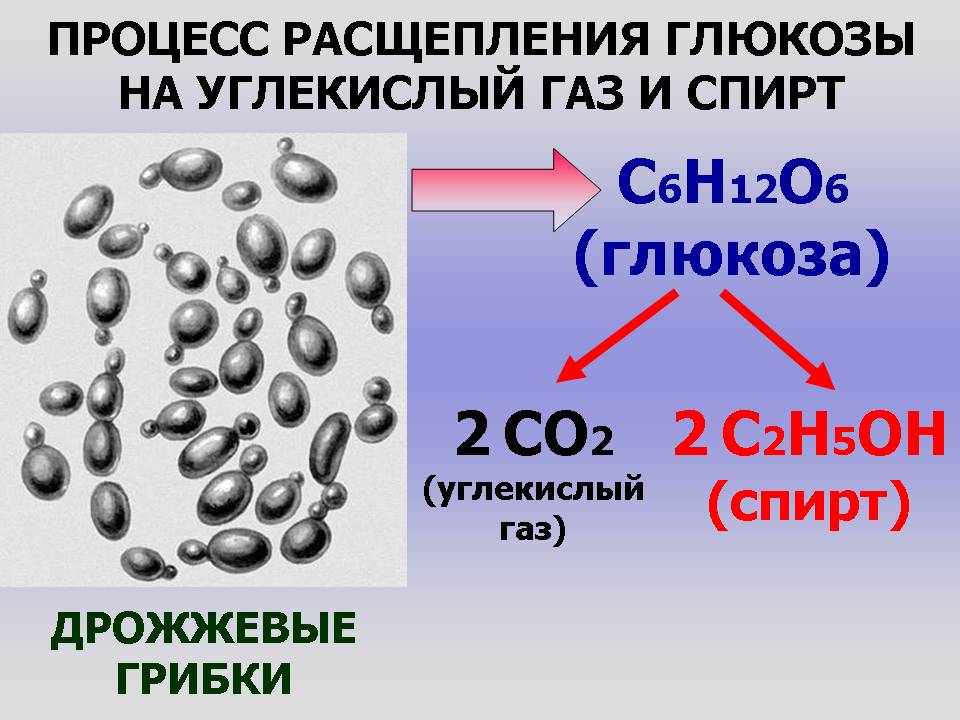
Он состоит из нескольких последовательных реакций превращения молекулы глюкозы C6h22O6 в две молекулы пировиноградной кислоты — ПВК C3h5O3 и две молекулы АТФ (в виде которой запасается примерно \(40\) % энергии, выделившейся при гликолизе). Остальная энергия (около \(60\) %) рассеивается в виде тепла.
C6h22O6+2h4PO4+2АДФ=2C3h5O3+2АТФ +2h3O.
Получившаяся пировиноградная кислота при недостатке кислорода в клетках животных, а также клетках многих грибов и микроорганизмов, превращается в молочную кислоту C3H6O3.
HOOC−CO−Ch4пировиноградная кислота→НАД⋅H+H+лактатдегидрогеназаHOOC−CHOH−Ch4молочная кислота.
В мышцах человека при больших нагрузках и нехватке кислорода образуется молочная кислота и появляется боль. У нетренированных людей это происходит быстрее, чем у людей тренированных.
При недостатке кислорода в клетках растений, а также в клетках некоторых грибов (например, дрожжей), вместо гликолиза происходит спиртовое брожение: пировиноградная кислота распадается на этиловый спирт C2H5OH и углекислый газ CO2:
C6h22O6+2h4PO4+2АДФ=2C2H5OH+2CO2+2АТФ+2h3O.
Третий этап — кислородный
В результате гликолиза глюкоза распадается не до конечных продуктов (CO2 и h3O), а до богатых энергией соединений (молочная кислота, этиловый спирт) которые, окисляясь дальше, могут дать её в больших количествах. Поэтому у аэробных организмов после гликолиза (или спиртового брожения) следует третий, завершающий этап энергетического обмена — полное кислородное расщепление, или клеточное дыхание.
Этот этап происходит на кристах митохондрий.
Третий этап, так же как и гликолиз, является многостадийным и состоит из двух последовательных процессов — цикла Кребса и окислительного фосфорилирования.
Третий (кислородный) этап заключается в том, что при кислородном дыхании ПВК окисляется до окончательных продуктов — углекислого газа и воды, а энергия, выделяющаяся при окислении, запасается в виде \(36\) молекул АТФ (\(2\) молекулы в цикле Кребса и \(34\) молекулы в ходе окислительного фосфорилирования).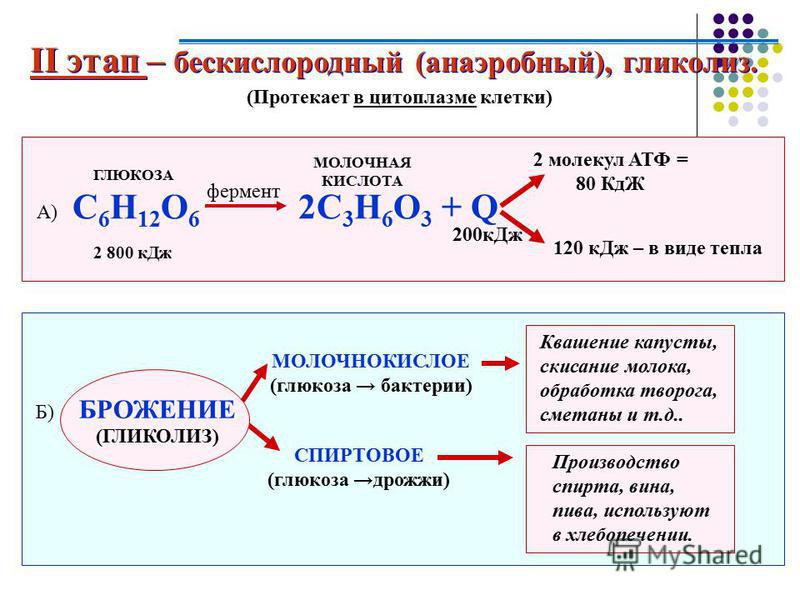
Этот этап можно представить себе в следующем виде:
2C3h5O3+6O2+36h4PO4+36АДФ=6CO2+42h3O+36АТФ.
Вспомним, что ещё две молекулы АТФ запасаются в ходе бескислородного расщепления каждой молекулы глюкозы (на втором, бескислородном, этапе). Значит, суммарный результат полного окисления молекулы глюкозы составляет \(38\) молекул АТФ.
Суммарное уравнение энергетического обмена:
C6h22O6+6O2=6CO2+6h3O+38АТФ.
В реакциях энергетического обмена используется не только глюкоза, но и липиды, белки. Но главным источником энергии в большинстве клеток являются углеводы.
ГЛИКОЛИЗ • Большая российская энциклопедия
В книжной версии
Том 7.
 Москва, 2007, стр. 231-232
Москва, 2007, стр. 231-232Скопировать библиографическую ссылку:
Авторы: А. Е. Медведев
ГЛИКО́ЛИЗ (от греч. γλυϰύς – сладкий и …лиз), ферментативный процесс анаэробного (протекающего без участия кислорода) расщепления глюкозы, сопровождающийся накоплением энергии, запасаемой в виде аденозинтрифосфата (АТФ). Широко распространён в природе, играет важную роль в метаболизме живых организмов. Г. лежит в основе разл. видов брожения. У облигатных анаэробных микроорганизмов, а также в некоторых клетках животных и человека (напр., зрелые эритроциты млекопитающих) Г. – единственный процесс, поставляющий энергию.
Полная детальная расшифровка отд. реакций Г. была осуществлена гл. обр. благодаря работам немецких биохимиков Г. Эмбдена, О. Мейергофа, О. Варбурга, К. Нойберга, а также Я. О. Парнаса и К. Кори (1930–37). Иногда Г. называют «путём Эмбдена – Мейергофа – Парнаса».
Условно в процессе Г. выделяют две стадии. На первой (подготовительной) сначала происходит фосфорилирование глюкозы (реакция 1), катализируемое ферментом гексокиназой (иногда глюкокиназой) и требующее затраты энергии молекулы АТФ (см.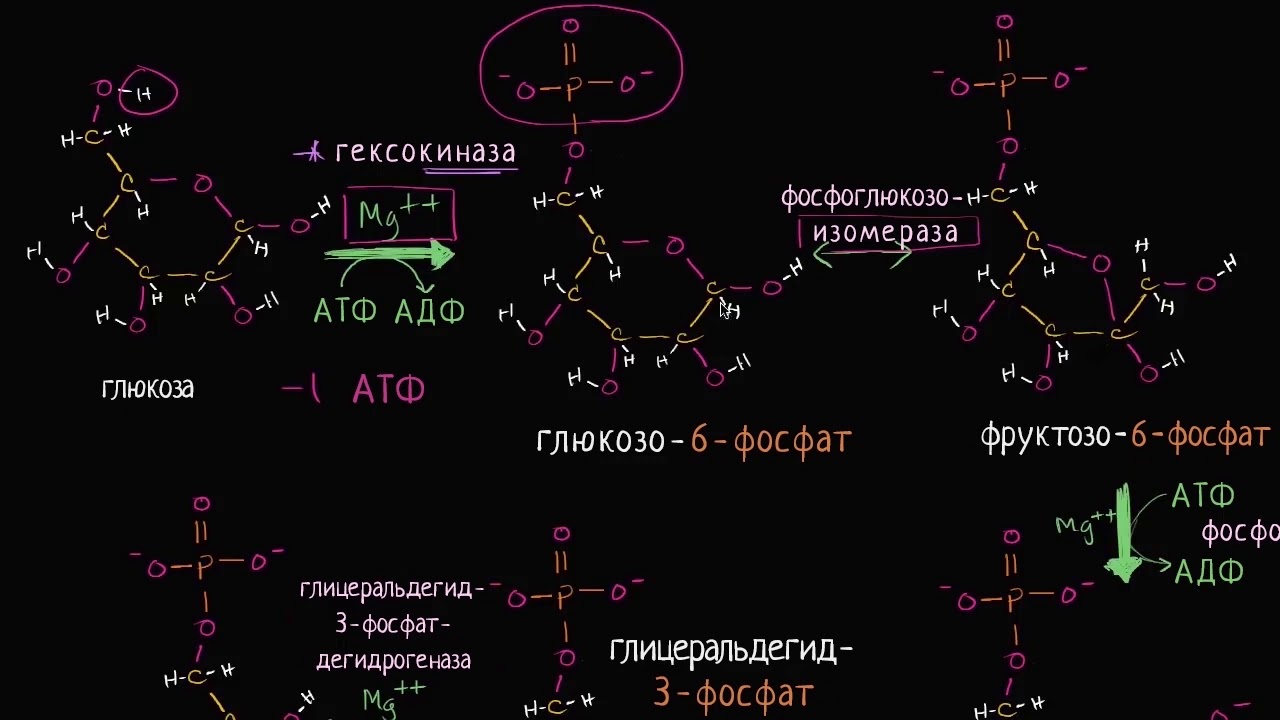 схему). Образовавшийся глюкозо-6-фосфат при участии фермента фосфоглюкоизомеразы превращается (2) во фруктозо-6-фосфат, который в свою очередь фосфорилируется (для этого требуется ещё одна молекула АТФ) с помощью фосфофруктокиназы-1 (3) с образованием фруктозо-1,6-дифосфата. Последний с помощью фруктозо-1,6-дифосфатальдолазы (4) расщепляется до двух фосфотриоз: глицеральдегид-3-фосфата и дигидроксиацетонфосфата. Последний легко изомеризуется под действием триозофосфатизомеразы в глицеральдегид-3-фосфат (5). В дальнейшем используется только первое соединение. Т. о., из одной молекулы глюкозы на первой стадии Г. образуются две молекулы глицеральдегид-3-фосфата и при этом используется энергия двух молекул АТФ. На второй – осн. стадии Г. – глицеральдегид-3-фосфат претерпевает серию последовательных превращений, приводящих к образованию пировиноградной кислоты (ионизированная формула – пируват) и выделению энергии, аккумулируемой в виде АТФ.
схему). Образовавшийся глюкозо-6-фосфат при участии фермента фосфоглюкоизомеразы превращается (2) во фруктозо-6-фосфат, который в свою очередь фосфорилируется (для этого требуется ещё одна молекула АТФ) с помощью фосфофруктокиназы-1 (3) с образованием фруктозо-1,6-дифосфата. Последний с помощью фруктозо-1,6-дифосфатальдолазы (4) расщепляется до двух фосфотриоз: глицеральдегид-3-фосфата и дигидроксиацетонфосфата. Последний легко изомеризуется под действием триозофосфатизомеразы в глицеральдегид-3-фосфат (5). В дальнейшем используется только первое соединение. Т. о., из одной молекулы глюкозы на первой стадии Г. образуются две молекулы глицеральдегид-3-фосфата и при этом используется энергия двух молекул АТФ. На второй – осн. стадии Г. – глицеральдегид-3-фосфат претерпевает серию последовательных превращений, приводящих к образованию пировиноградной кислоты (ионизированная формула – пируват) и выделению энергии, аккумулируемой в виде АТФ.

Схема гликолиза.
Дальнейшая судьба образовавшегося пирувата и НАДН определяется особенностями метаболизма конкретных типов клеток и их обеспеченностью кислородом. В аэробных условиях пируват и восстановленные эквиваленты от НАДН (гидрид-ион) поступают в митохондрии, где происходит превращение пирувата в ацетил-КоА (см. Кофермент А), который поступает в трикарбоновых кислот цикл. В отсутствии кислорода НАДН используется на восстановление пирувата в лактат (ионизированная форма молочной кислоты). Это необходимо для получения НАД, без которого невозможно протекание глицеральдегид-3-фосфатдегидрогеназной реакции, а значит и всего Г.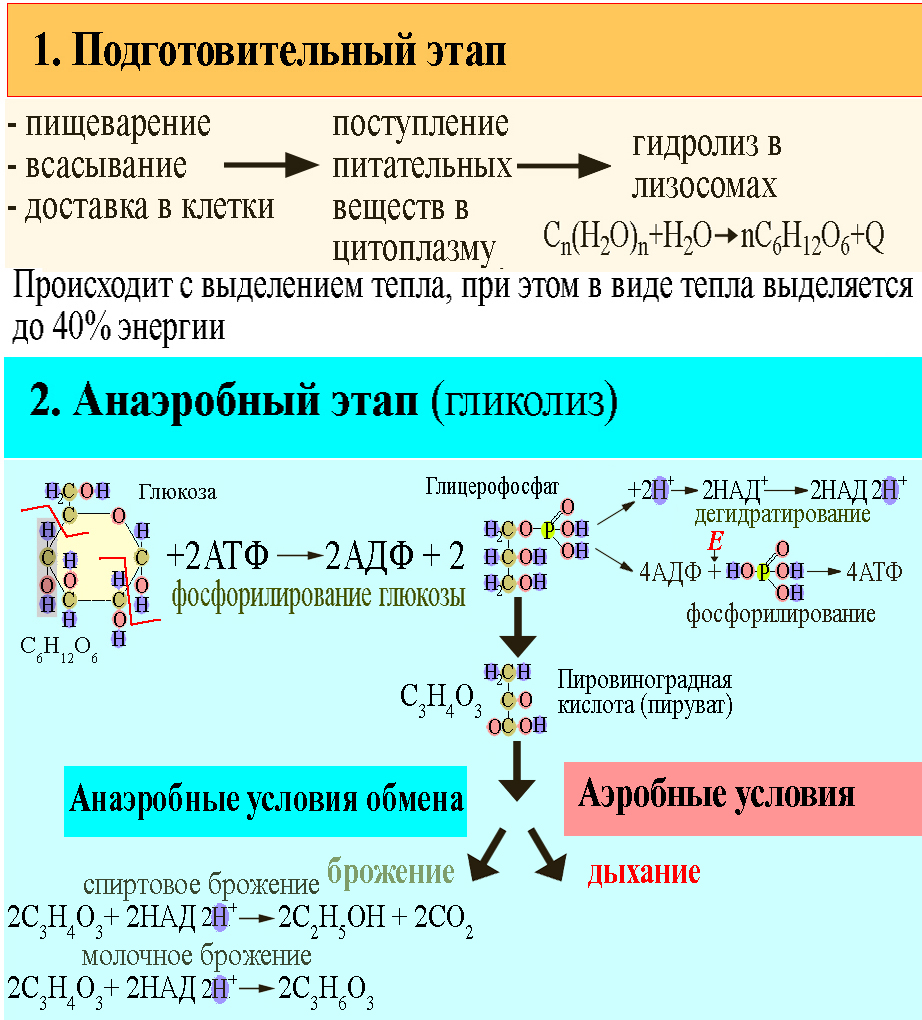

Все реакции Г., за исключением 1, 3 и 10-й, обратимы. Ферменты, катализирующие эти три реакции, являются объектами сложной регуляции. Скорость Г. лимитирует фосфофруктокиназа-1. Этот фермент активируют фруктозо-2,6-дифосфат, АДФ и аденозинмонофосфат (АМФ), а ингибируют АТФ и цитрат (ионизированная форма лимонной кислоты). АТФ подавляет также активность гексокиназы и пируваткиназы. Образование мощного активатора Г. – фруктозо-2,6-дифосфата – из фруктозо-6-фосфата осуществляет особый фермент фосфофруктокиназа-2. Гормоны адреналин и глюкагон через систему внутриклеточной сигнализации, включающую образование циклического 3’,5’-аденозинмонофосфата (цАМФ) и активацию фермента протеинкиназы, вызывают фосфорилирование и инактивацию пируваткиназы в печени, но не в мышцах.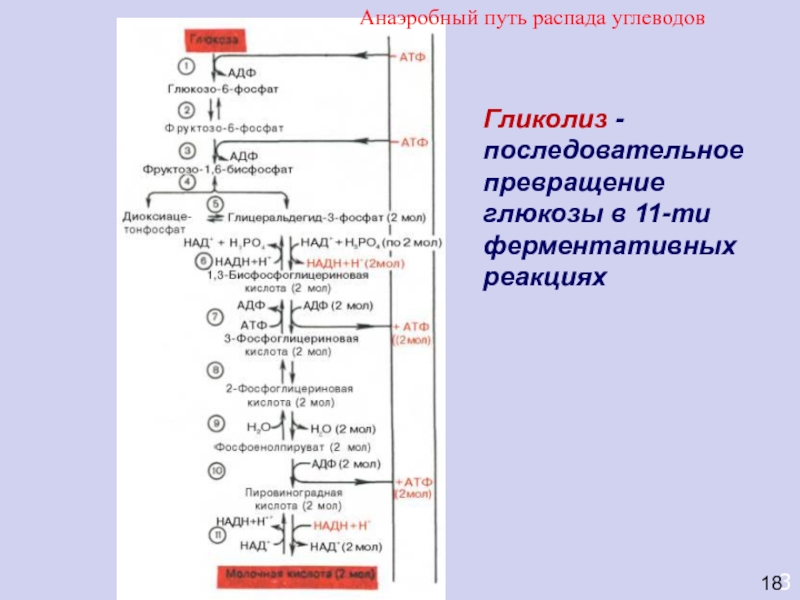
Установлено, что некоторым ферментам Г. свойствен ряд функций, не связанных с этим процессом. Напр., глицеральдегид-3-фосфатдегидрогеназа может переноситься в ядро и участвовать в индукции апоптоза, активируя процессы транскрипции.
ГЛИКОЛИЗ. «БИОЛОГИЧЕСКАЯ ХИМИЯ», Березов Т.Т., Коровкин Б.Ф.
Гликолиз (от греч. glycys – сладкий и lysis – растворение, распад) – это последовательность ферментативных реакций, приводящих к превращению глюкозы в пируват с одновременным образованием АТФ.
При аэробных
условиях пируват проникает в митохондрии, где полностью окисляется до СО2
и Н2О. Если содержание кислорода недостаточно, как это может иметь
место в активно сокращающейся мышце, пируват превращается в лактат.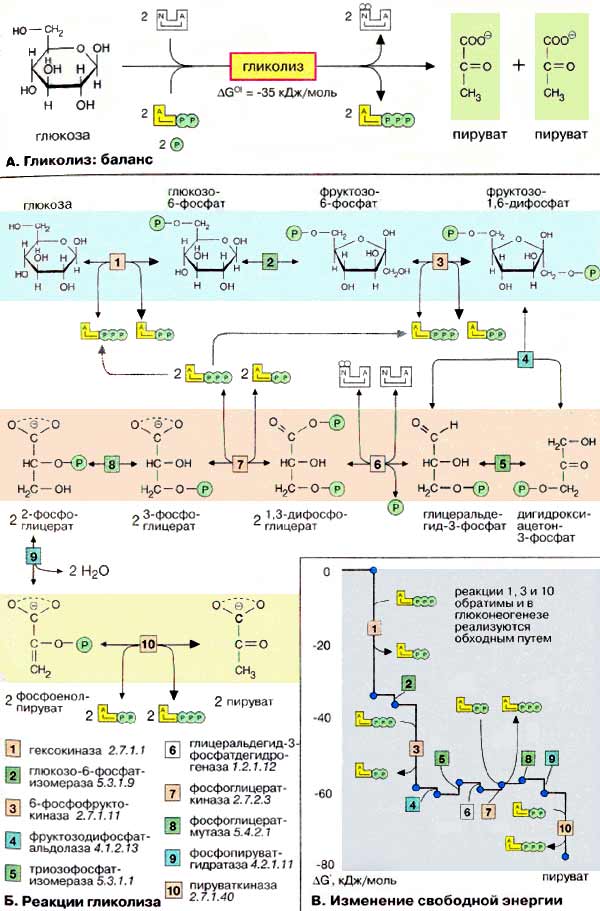
Итак, гликолиз – не только главный путь утилизации глюкозы в клетках, но и уникальный путь, поскольку он может использовать кислород, если
последний доступен (аэробные условия), но может протекать и в отсутствие кислорода (анаэробные условия).
Анаэробный гликолиз – сложный ферментативный процесс распада глюкозы, протекающий в тканях человека и животных без потребления кислорода. Конечным продуктом гликолиза является молочная кислота. В процессе гликолиза образуется АТФ. Суммарное уравнение гликолиза можно представить следующим образом:
В анаэробных условиях гликолиз – единственный процесс в животном организме, поставляющий энергию. Именно благодаря гликолизу организм человека и животных определенный период может осуществлять ряд физиологических функций в условиях недостаточности кислорода. В тех случаях, когда гликолиз протекает в присутствии кислорода, говорят об аэробном гликолизе .
Последовательность
реакций анаэробного гликолиза, так же как и их промежуточные продукты, хорошо
изучена.
Первой ферментативной реакцией гликолиза является фосфорилирование, т.е. перенос остатка ортофосфата на глюкозу за счет АТФ. Реакция катализируется ферментом гексокиназой:
Образование глюкозо-6-фосфата в гексокиназной реакции сопровождается освобождением значительного количества свободной энергии системы и может считаться практически необратимым процессом.
Наиболее важным свойством гексокиназы является ее ингибирование глюкозо-6-фосфатом, т.е. последний служит одновременно и продуктом реакции, и аллостерическим ингибитором.
Фермент
гексокиназа способен катализировать фосфорилирование не только D-глюкозы, но и
других гексоз, в частности D-фруктозы, D-маннозы и т. д. В печени, кроме
гексокиназы, существует фермент глюкокиназа, который катализирует фосфорилирование
только D-глюкозы. В мышечной ткани этот фермент отсутствует (подробнее см.
главу 16).
д. В печени, кроме
гексокиназы, существует фермент глюкокиназа, который катализирует фосфорилирование
только D-глюкозы. В мышечной ткани этот фермент отсутствует (подробнее см.
главу 16).
Второй реакцией гликолиза является превращение глюкозо-6-фос-фата под действием фермента глюкозо-6-фосфатизомеразы во фруктозо-6-фосфат:
Эта реакция протекает легко в обоих направлениях, и для нее не требуется каких-либо кофакторов.
Третья реакция катализируется ферментом фосфофруктокиназой; образовавшийся фруктозо-6-фосфат вновь фосфорилируется за счет второй молекулы АТФ:
Данная реакция аналогично гексокиназной практически необратима, протекает в присутствии ионов магния и является наиболее медленно текущей реакцией гликолиза. Фактически эта реакция определяет скорость гликолиза в целом.
Фосфофруктокиназа
относится к числу аллостерических ферментов. Она ингибируется АТФ и
стимулируется АМФ .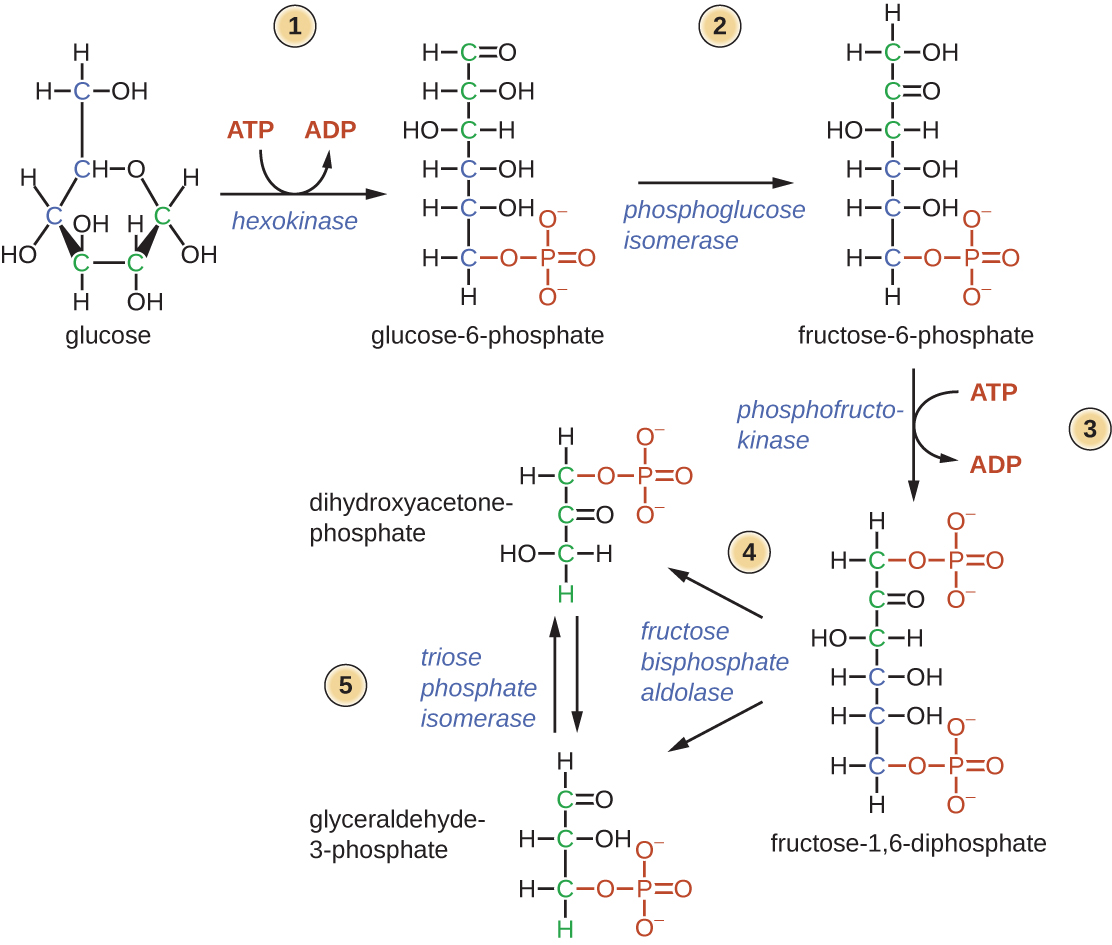 При значительных величинах отношения АТФ/АМФ активность
фосфофруктокиназы угнетается и гликолиз замедляется. Напротив, при снижении
этого коэффициента интенсивность гликолиза повышается. Так, в неработающей
мышце активность фосфофруктокиназы низкая, а концентрация АТФ относительно
высокая. Во время работы мышцы происходит интенсивное потребление АТФ и
активность фосфофруктокиназы повышается, что приводит к усилению процесса
гликолиза.
При значительных величинах отношения АТФ/АМФ активность
фосфофруктокиназы угнетается и гликолиз замедляется. Напротив, при снижении
этого коэффициента интенсивность гликолиза повышается. Так, в неработающей
мышце активность фосфофруктокиназы низкая, а концентрация АТФ относительно
высокая. Во время работы мышцы происходит интенсивное потребление АТФ и
активность фосфофруктокиназы повышается, что приводит к усилению процесса
гликолиза.
Четвертую реакцию гликолиза катализирует фермент альдолаза. Под влиянием этого фермента фруктозо-1,6-бисфосфат расщепляется на две фосфотриозы:
Эта реакция обратима. В зависимости от температуры равновесие устанавливается на различном уровне. При повышении температуры реакция сдвигается в сторону большего образования триозофосфатов (дигидро-ксиацетонфосфата и глицеральдегид-3-фосфата).
Пятая реакция – это реакция изомеризации триозофосфатов. Катализируется ферментом триозофосфатизомеразой:
Равновесие
данной изомеразной реакции сдвинуто в сторону дигид-роксиацетонфосфата: 95%
дигидроксиацетонфосфата и около 5% глице-ральдегид-3-фосфата.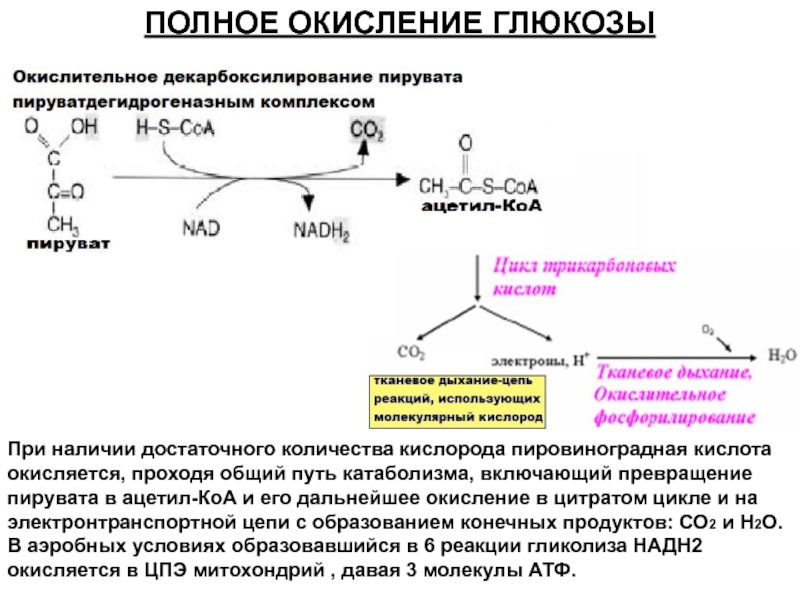 В последующие
реакции гликолиза может непосредственно включаться только один из двух
образующихся триозофосфатов, а именно глицеральдегид-3-фосфат. Вследствие этого
по мере потребления в ходе дальнейших превращений альдегидной формы фосфотриозы
ди-гидроксиацетонфосфат превращается в глицеральдегид-3-фосфат.
В последующие
реакции гликолиза может непосредственно включаться только один из двух
образующихся триозофосфатов, а именно глицеральдегид-3-фосфат. Вследствие этого
по мере потребления в ходе дальнейших превращений альдегидной формы фосфотриозы
ди-гидроксиацетонфосфат превращается в глицеральдегид-3-фосфат.
Образованием глицеральдегид-3-фосфата как бы завершается первая стадия гликолиза. Вторая стадия – наиболее сложная и важная. Она включает окислительно-восстановительную реакцию (реакция гликолитической оксидоредукции), сопряженную с субстратным фосфорилированием, в процессе которого образуется АТФ.
В результате шестой реакции глицеральдегид-3-фосфат в присутствии фермента глицеральдегидфосфатдегидрогеназы, кофермента НАД и неорганического фосфата подвергается своеобразному окислению с образованием 1,3-бисфосфоглицериновой кислоты и восстановленной формы НАД (НАДН). Эта реакция блокируется йод- или бромацетатом, протекает в несколько этапов:
1,3-Бисфосфоглицерат
представляет собой высокоэнергетическое соединение (макроэргическая связь
условно обозначена знаком «тильда» ~). Механизм действия
глицеральдегидфосфатдегидрогеназы сводится к следующему: в присутствии
неорганического фосфата НАД+ выступает как акцептор водорода,
отщепляющегося от глицеральдегид-3-фосфата. В процессе образования НАДН
глицеральдегид-3-фосфат связывается с молекулой фермента за счет SH-групп
последнего. Образовавшаяся связь богата энергией, но она непрочная и
расщепляется под влиянием неорганического фосфата, при этом образуется
1,3-бисфосфоглицериновая кислота.
Механизм действия
глицеральдегидфосфатдегидрогеназы сводится к следующему: в присутствии
неорганического фосфата НАД+ выступает как акцептор водорода,
отщепляющегося от глицеральдегид-3-фосфата. В процессе образования НАДН
глицеральдегид-3-фосфат связывается с молекулой фермента за счет SH-групп
последнего. Образовавшаяся связь богата энергией, но она непрочная и
расщепляется под влиянием неорганического фосфата, при этом образуется
1,3-бисфосфоглицериновая кислота.
Седьмая реакция катализируется фосфоглицераткиназой, при этом происходит передача богатого энергией фосфатного остатка (фосфатной группы в положении 1) на АДФ с образованием АТФ и 3-фосфогли-цериновой кислоты (3-фосфоглицерат):
Таким образом, благодаря действию двух ферментов (глицеральде-гидфосфатдегидрогеназы и фосфоглицераткиназы) энергия, высвобождающаяся при окислении альдегидной группы глицеральдегид-3-фосфата до карбоксильной группы, запасается в форме энергии АТФ. В отличие от окислительного фосфорилирования образование АТФ из высокоэнергетических соединений называется субстратным фосфорилированием.
Восьмая реакция сопровождается внутримолекулярным переносом оставшейся фосфатной группы, и 3-фосфоглицериновая кислота превращается в 2-фосфоглицериновую кислоту (2-фосфоглицерат).
Реакция легкообратима, протекает в присутствии ионов Mg2+. Кофактором фермента является также 2,3-бисфосфоглицериновая кислота аналогично тому, как в фосфоглюкомутазной реакции роль кофактора выполняет глюкозо-1,6-бисфосфат:
Девятая реакция катализируется ферментом енолазой, при этом 2-фосфоглицериновая кислота в результате отщепления молекулы воды переходит в фосфоенолпировиноградную кислоту (фосфоенолпируват), а фосфатная связь в положении 2 становится высокоэргической:
Енолаза активируется двухвалентными катионами Mg2+или Мn2+ и ингибируется фторидом.
Десятая реакция характеризуется разрывом высокоэргической связи и переносом фосфатного остатка от фосфоенолпирувата на АДФ (субстратное фосфорилирование). Катализируется ферментом пируваткиназой:
Для действия пируваткиназы необходимы ионы Mg2+, а также одновалентные катионы щелочных металлов (К+ или др.). Внутри клетки реакция является практически необратимой.
В результате одиннадцатой реакции происходит восстановление пировиноградной кислоты и образуется молочная кислота. Реакция протекает при участии фермента лактатдегидрогеназы и кофермента НАДН, образовавшегося в шестой реакции:
Последовательность протекающих при гликолизе реакций представлена на рис. 10.3.
Рис. 10.3. Последовательность реакций гликолиза.
1 - гексокиназа; 2 — фосфоглюкоизоме-раза; 3 — фосфофруктокиназа; 4 — альдо-лаза; 5 — триозофосфатизомераза; 6 — гли-церальдегидфосфатдегидрогеназа; 7 -фосфоглицераткиназа; 8 — фосфоглицеромутаза; 9 — енолаза; 10 - пируватки-наза; 11 - лактатдегидрогеназа.
Реакция восстановления пирувата завершает внутренний окислительно-восстановительный цикл гликолиза. НАД+ при этом играет роль промежуточного переносчика водорода от глицеральдегид-3-фосфата (6-я реакция) на пировиноградную кислоту (11-я реакция), при этом сам он регенерируется и вновь может участвовать в циклическом процессе, получившем название гликолитический оксидоредукции.
Биологическое значение процесса гликолиза заключается прежде всего в образовании богатых энергией фосфорных соединений. На первых стадиях гликолиза затрачиваются 2 молекулы АТФ (гексокиназная и фосфофрук-токиназная реакции). На последующих образуются 4 молекулы АТФ (фосфоглицераткиназная и пируваткиназная реакции). Таким образом, энергетическая эффективность гликолиза в анаэробных условиях составляет 2 молекулы АТФ на одну молекулу глюкозы.
Как отмечалось, основной реакцией, лимитирующей скорость гликолиза, является фосфофруктокиназная. Вторая реакция, лимитирующая скорость и регулирующая гликолиз – гексокиназная реакция. Кроме того, контроль гликолиза осуществляется также ЛДГ и ее изоферментами.
В тканях с аэробным метаболизмом (ткани сердца, почек и др.) преобладают изоферменты ЛДГ1 и ЛДГ2 (см. главу 4). Эти изоферменты инги-бируются даже небольшими концентрациями пирувата, что препятствует образованию молочной кислоты и способствует более полному окислению пирувата (точнее, ацетил-КоА) в цикле трикарбоновых кислот.
В тканях человека, в значительной степени использующих энергию гликолиза (например, скелетные мышцы), главными изоферментами являются ЛДГ5 и ЛДГ4. Активность ЛДГ5 максимальна при тех концентрациях пирувата, которые ингибируют ЛДГ1. Преобладание изоферментов ЛДГ4 и ЛДГ5 обусловливает интенсивный анаэробный гликолиз с быстрым превращением пирувата в молочную кислоту.
Как отмечалось, процесс анаэробного распада гликогена получил название гликогенолиза. Вовлечение D-глюкозных единиц гликогена в процесс гликолиза происходит при участии 2 ферментов – фосфорилазы а и фосфо-глюкомутазы. Образовавшийся в результате фосфоглюкомутазной реакции глюкозо-6-фосфат может включаться в процесс гликолиза. После образования глюкозо-6-фосфата дальнейшие пути гликолиза и гликогенолиза полностью совпадают:
В процессе гликогенолиза в виде макроэргических соединений накапливаются не две, а три молекулы АТФ (АТФ не тратится на образование глюкозо-6-фосфата). Кажется, что энергетическая эффективность глико-генолиза выглядит несколько более высокой по сравнению с процессом гликолиза, но эта эффективность реализуется только при наличии активной фосфорилазы а. Следует иметь в виду, что в процессе активации фосфо-рилазы b расходуется АТФ (см. рис. 10.2).
Предыдущая страница | Следующая страница
СОДЕРЖАНИЕ
Еще по теме:
Образование молочной кислоты при гликолизе
Молочная кислота образуется в мышцах в анаэробных условиях и является конечным продуктом гликолиза. Количество образовавшейся молочной кислоты эквивалентно количеству распавшейся глюкозы. Установлено, что содержание молочной кислоты в крови человека и животных повышается после мышечной работы. Особенно резкое увеличение количества молочной кислоты наблюдается после усиленных мышечных упражнений. Однако уровень молочной кислоты в крови быстро снижается, так как она поглощается печенью и превращается там в гликоген. Ресинтез гликогена из молочной кислоты не может протекать самопроизвольно и осуществляется только при условии сопряжения его с окислительными процессами, дающими энергию. По данным Пастера и Мейергофа, ресинтез гликогена сопряжен с окислением некоторой части молочной кислоты до углекислого газа и воды. Основная масса молочной кислоты при этом превращается в гликоген. В настоящее время установлено, что в аэробных условиях при достаточном притоке кислорода гликогек и глюкоза окисляются через стадию пировиноградной кислоты до СОг и Н2О, минуя образование молочной кислоты (см. стр. 172). [c.254]Во второй пробе (где происходил гликолиз) развивается интенсивное красное окрашивание, указывающее на образование молочной кислоты. [c.155]
Важнейшими из них, как в настоящее время установлено, являются 1) гликогенолиз или гликолиз (расщепление гликогена или глюкозы с образованием молочной кислоты, стр. 257) 2) тканевое дыхание (окисление до 02 и НгО тех или иных субстратов дыхания, главным образом углеводов) 3) перенос фосфатной группы с фосфокреатина на АДФ. Фосфорилирование самого креатина в мышечной ткани с образованием фосфокреатина в конечном счете осуществляется за счет энергии двух первых процессов. [c.426]
О-глюкоза — ОСНОВНОЙ источник энергии живых организмов. При гликолизе 1 г/моля глюкозы выделяется 196,3 кДж. Ферментативное расщепление глюкозы в живой клетке протекает до образования молочной кислоты, сопряженной с образованием аденозинтрифосфорной кислоты (АТФ). [c.102]
С другой стороны, в присутствии кислорода пировиноградная кислота полностью окисляется в СО2 и Н2О, причем этот процесс несравненно более выгоден с энергетической точки зрения, чем образование молочной кислоты. (Впрочем, затрата, сделанная в анаэробном процессе, в котором топливо используется неудовлетворительно, компенсируется регенерацией гликогена из молочной кислоты приведенным выше образом.) Мышечные экстракты содержат исключительно ферменты анаэробного гликолиза, а не дыхательные ферменты. Эти ферменты тесно связаны с некоторыми структурными элементами клеток, называемыми митохондриями. Поэтому окислительные процессы изучались со срезами органов или измельченными тканями. [c.254]
Все жизненные процессы сопровождаются гликолизом — биологическим расщеплением гликогена, приводящим к образованию молочной кислоты для животных организмов гликоген является одним из важнейших источников энергии. Он содержится во всех клетках животного организма. Наиболее богаты гликогеном печень (у упитанных животных до 10—20% гликогена) и мышцы (до 4%)- Он содержится также в некоторых низших растениях, например в дрожжах и грибах крахмал некоторых высших растений по свойствам близок к гликогену. [c.711]
Объектом для изучения анаэробного гликолиза уже с давних пор, наряду с дрожжевыми клетками, служит мышца. Оказалось, что гликолиз в мышцах и спиртовое брожение в дрожжевых клетках происходят по одному и тому же пути, следовательно, с образованием одинаковых промежуточных продуктов. Различия имеются только лишь на этапе превра-н1,ения пировиноградной кислоты, которая в мышцах при анаэробном гликолизе не подвергается декарбоксилированию, а восстанавливается с образованием молочной кислоты. Отсюда конечным продуктом анаэробного гликолиза в мышцах является молочная кислота, в то время как в дрожжевых клетках — этиловый спирт и углекислый газ. Необходимо отметить, что анаэробный распад углеводов с выделением молочной кислоты специфичен не только для мышц. Установлено, что подобный процесс происходит и в других тканях организма человека и животных. Он имеет место также у микроорганизмов (бактерий молочнокислого брожения), у которых анаэробный распад углеводов заканчивается образованием молочной кислоты. [c.288]
Анаэробное превращение углеводов, начинающееся с гликогена или глюкозы и заканчивающееся образованием молочной кислоты, получило название гликоген о лиза или соответственно гликолиза. Термин гликогенолиз употребляется в тех случаях, когда исходным субстратом превращения является гликоген, а термин гликолиз — когда таковым является глюкоза. [c.249]
Из схем видно, что основное отличие механизма гликолиза (анаэробного расщепления сахара с образованием молочной кислоты) от механизма окислительного распада углеводов сводится по существу к следующему при гликолизе пировиноградная кислота восстанавливается и превращается в молочную кислоту — конечный продукт анаэробного обмена, при дыхании образующаяся пировиноградная кислота подвергается дальнейшему окислению с образованием в конечном счете воды и СОз. [c.258]
Насколько резко может усиливаться обмен веществ в мышцах при их сокращении, показывают следуют,ие цифры покоящиеся мышцы человека поглощают около 1,7 мл кислорода на 1 кг ткани в минуту при очень напряженной работе мышечная ткань потребляет за то же время около 180 мл кислорода на 1 кг веса, т. е. окислительный обмен при работе мышцы усиливается примерно в 100 раз. Еще больше увеличивается при работе мышцы в анаэробных условиях образование молочной кислоты. Так, например, в мышцах лягушки на 1 кг ткани в состоянии покоя образуется 0,2 мг молочной кислоты в течение часа, при тетаническом же сокращении — до 180 мг, т. е. интенсивность анаэробного гликолиза при работе возрастает почти в 1000 раз. [c.413]
Работа 135. Образование молочной кислоты при гликолизе [c.180]
Насколько резко может усиливаться обмен веществ в мышцах при их сокращении, показывают следующие цифры покоящиеся мышцы человека поглощают около 1,7 мл кислорода на 1 кг ткани в минуту при очень напряженной работе мышечная ткань потребляет за то же время около 180 мл кислорода на 1 кг веса, т. е. окислительный обмен при работе мышцы усиливается примерно в 100 раз. Еще больше увеличивается при работе мышцы в анаэробных условиях образование молочной кислоты. Интенсивность анаэробного гликолиза при работе может возрастать почти в 1000 раз. [c.437]
Важнейшими из них, как в настоящее время установлено, являются 1) тканевое дыхание (окисление до СО2 и HgO тех или иных субстратов дыхания, главным образом углеводов) 2) гликогенолиз или гликолиз (расщепление гликогена или глюкозы с образованием молочной кислоты, стр. 265) 3) перенос фосфатной группы с фосфокреатина на АДФ [c.450]
Можно также отметить, что представлению о необходимости образования молочной кислоты для сокращения мышц противоречат данные Эмбдена, показавшего, что молочная кислота быстро образуется в мышце и продолжает накапливаться в течение нескольких первых секунд после расслабления. В охлажденной мышце гликолиз начинается вообще лишь через 1—2 сек после начала сокращения и заканчивается через 30 сек — 5 мин после его окончания. [c.451]
В результате анаэробной фазы обмена глюкозы (гликолиз), заканчивающейся образованием молочной кислоты, образуется 3 молекулы АТФ на каждую глюкозную единицу мышечного гликогена. [c.388]
Добавление глюкозы к взвеси пластинок приводит к ее потреблению, особенно отчетливо выраженному в анаэробных условиях, однако соответствующего нарастания содержания молочной кислоты при этом не наблюдалось. По-видимому, это можно объяснить не отсутствием гликолиза, поскольку потребление сахара имело место, но остановкой этого процесса на более ранних стадиях гликоли-тических превращений углеводов до образования молочной кислоты. [c.133]
Нельзя с уверенностью сказать, прекращается ли гликолиз при замораживании взвеси пластинок или же этот процесс останавливается на более ранних стадиях расщепления углеводов, до образования молочной кислоты. Последнее предположение кажется вероятным потому, что в пробах пластинок с добавлением глюкозы, хранившихся при —20°, сохраняется способность к подавлению потребления кислорода в течение всего срока исследования. [c.135]
В раковой ткани, в отличие от нормальной, гликогенолиз с образованием молочной кислоты идет также в аэробных условиях одновременно с дыханием. Раньше предполагали, что такой аэробный гликолиз идет теми же [c.502]
Гликолиз представляет собой необратимый процесс, равновесие которого полностью смещено в сторону образования молочной кислоты, что можно объяснить значительным уменьшением свободной энергии (АС =—135,9 кДж/моль). В то же время большая часть этапов этого процесса имеет небольшие значения изменений свободной энергии. [c.419]
Образование молочной кислоты в процессе гликолиза в зависимости от продолжительности упражнения [c.314]
Сходство путей метаболизма в различных видах — один из основных принципов биохимии. Классические исследования, посвященные спиртовой ферментации дрожжей и образованию молочной кислоты в тканях млекопитающих, показали, что эти два процесса по существу протекают одинаково и отличаются лишь конечными стадиями, когда в дрожжах происходит анаэробное декарбоксилирование пирувата, а в мышечной ткани — нет. И в том, и в другом процессе НАД восстанавливается, а энергия накапливается в виде АТФ. Последние исследования других биологических механизмов образования, накопления и передачи энергии выявили некоторые интересные различия между видами, например наличие нескольких путей диссимиляции сахаров в бактериях, но все же наблюдается удивительное сходство этих механизмов. Многие промежуточные соединения одинаковы для всех видов. В живых клетках в качестве аккумулятора энергии всегда используется АТФ. Никотииамиднуклео-тиды участвуют во многих реакциях с переносом электрона триозофосфаты всегда участвуют в гликолизе. Белки, являющиеся основой живых организмов, во всех исследованных видах состоят приблизительно из 20 аминокислот. Эти аминокислоты, по-видимому,. в целом ряде организмов синтезируются одинаково, хотя точно установлено наличие двух путей в случае лизина. При этом высшие растения и бактерии используют различные пути, а грибы — оба. Это интересно при прослеживании эволюционных линий по био- [c.234]
Следует подчеркнуть, что с энергетической точки зрения гликогенолиз, как и гликолиз, мало эффективны. Из всей потенциальной энергии глюкозного остатка молекулы гликогена только лишь часть становится доступной для использования клеткой при образовании из этого остатка двух молекул молочной кислоты. Остальное количество энергии (19 из 20 частей) остается в молекулах молочной кислоты. Однако, несмотря на малую энергетическую эффективность гликогенолиза, физиологическое значение его велико, особенно в тех случаях, когда в тканях организма ощущается недостаток кислорода. Так, например, в начальной фазе мышечной работы, когда доставка кислорода к мышце не соответствует потребности в нем, наблюдается усиленный распад гликогена с образованием молочной кислоты. [c.291]
В большинстве тканей животных углеводы при аэробных условиях полностью окисляются, превращаясь в воду и углекислый газ, в то время как в отсутствие кислорода образуется молочная кислота. Л. Пастер впервые обратил внимание на тот факт, что гликолиз тормозится кислородом. Это явление вошло в науку под названием реакции Пастера . Значительно позже О. Варбург показал, что в эмбриональных тканях и тканях злокачественных опухолей гликолиз не снижается и в присутствии кислорода. Образование молочной кислоты в присутствии кислорода получило название аэробного гликолиза . [c.298]
Исследованиями Палладина, его сотрудников и других изучены в головном мозге отдельные ферменты анаэробного гликолиза. Можно считать установленным, что распад углеводов с образованием молочной кислоты (анаэробный гликолиз) в нервной системе происходит по тому же пути, как и в других тканях. В ткани головного мозга имеет место также и аэробный гликолиз (образование молочной кислоты в присутствии кислорода). Следует, однако, отметить, что энергия углеводов в основном используется в результате их аэробного распада с образованием углекислого газа и воды. [c.564]
Расчеты, проведенные на основании многочисленных экспериментов с глюкозой, содержащей в различных положениях углеродной цепи, показывают, что около 85-90% глюкозы, потребляемой мозгом взрослого животного, полностью окисляется до СО2 и Н2О около 5% расходуется в реакциях гликолиза с образованием молочной кислоты и лишь 5-7% использует- [c.147]
Кривая связывания кислорода гемоглобином зависит от pH при данной величине р(Ог) сродство к кислороду уменьшается номере уменьшения pH (эффект Бора). Гликолиз представляет собой анаэробный процесс, приводящий к образованию молочной кислоты и диоксида углерода. Оба эти соединения имеют тенденцию к понижению pH и способствуют высвобождению кислорода из оксигемоглобина там, где в этом есть необходимость, В дезоксигемоглобине, напротив, содержатся немного более основные, чем у оксигемоглобина, группы (азот имидазола His-146 в р-цепях и His-122 в а-цепях, а также аминогрупп Val-1 в а-цепях), в силу чего дезоксигемоглобин связывает протон после высвобождения кислорода, что важно для обратного транспорта диоксида углерода к легким. Карбоангидраза катализирует образование бикарбоната в эритроцитах из диоксида углерода и воды, и ионы бикарбоната могут связываться с протонированными группами дезокси-гемоглобина. В легких дезоксигемоглобин перезаряжается кислородом, эффект Бора вызывает высвобождение бикарбоната, из которого под действием карбоангидразы образуется диоксид углерода, который затем выдыхается. Транспорт диоксида углерода дезоксигемоглобином приводит также к образованию производных карбаминовой кислоты с аминогруппами белка (схема (9) . Хотя оксигемоглобин также связывает диоксид углерода, у дезоксигемо-глобина эта способность выше ввиду большей доступности аминогрупп. [c.558]
Гликолиз. Понятие гликолиз означает расщепление глюкозы. Первоначально этим термином обозначали только анаэробное брожение, завершающееся образованием молочной кислоты (лактата) или этанола и СО,. В настоящее время понятие гликолиз используется более широко для описания распада глюкозы, проходящего через образование глю-козо-6-фосфата, фруктозобисфосфата и пирувата как в отсутствие, так и в присутствии кислорода. В последнем случае употребляют термин аэробный гликолиз в отлгиие от анаэробного гликолиза , завершающегося образованием молочной кислоты (лактата). [c.319]
Гликолизом называют анаэробный распад углеводов в тканях с образованием молочной кислоты. Процесс гликолиза подробно изучен и включает в себя ряд отдельных реакций. В мышцах главным субстратом гликолиза является гликоген, который подвергается сначала ф о с-форолизу (распаду с присоединением фосфорной кислоты) и далее, через фосфорные эфиры гексоз, триоз и через пировиноградную кислоту распадается до молочной кислоты. Вследствие этого процесс этот часто называют также г л и-когенолизом. [c.153]
Каскадный процесс, схематически показанный на рис. 25-11, в печени и скелетных мыпщах протекает одинаково вплоть до образования глюкозо-6-фос-фата. Но в мышцах нет глюкозо-6-фос-фатазы, и поэтому в них не образуется свободной глюкозы. Повышение концентрации глюкозо-6-фосфата здесь приводит к значительному увеличению скорости гликолиза с образованием молочной кислоты, в ходе которого вырабатывается АТР, доступный для использования в процессе сокращения. Как показали сравнительно недавние исследования, адреналин стимулирует распад гликогена в печени через еще один каскад усиления, параллельный тому, который показан на рис. 25-11. В этом втором каскадном процессе, который в опреде- [c.791]
Реакцию катализирует фермент фосфопируватгидратаза (енолаза, КФ» 4.2.1.11). Фермент ингибируется фторидом (конечная концентрация фторида 0,02 М) особенно в присутствии фосфата. Считают, что ингибирующее влияние оказывает Mg2+-фтopфo фaтнь[й комплекс. Таким образом, добавление в инкубационную среду фторида прерывает гликолиз на стадии превращения фосфоглицериновых кислот и приводит к накоплению 3-фосфоглицерата. В этих условиях образования скольких-либо заметных количеств молочной кислоты не происходит. [c.54]Эти процессы настолько тесно интегрированы, что при аноксии образование молочной кислоты прямо пропорционально работе мышечного сокращения, и мышца способна переносить такие количества лактата, какие никогда не встречаются в других тканях. Таким образом, последнее функциональное требование, которое предъявляет мышечный гликолиз, — это возможность накопления больших количеств лактата и его последующего метаболизироваиия. [c.51]
Механизм переноса Ог в полость пузыря связан со второй системой капилляров, находящейся уже в самом эпителии этого органа (рис. 110). Кровь попадает здесь в условия высокой кислотности, которую поддерживает весьма активная система аэробного гликолиза в эпителиальных клетках. Гликолитические ферменты этой ткани эффективно функционируют при высоких напряжениях Ог. Эффект Пастера (торможение гликолиза при высоком напряжении Ог) здесь отсутствует — либо благодаря особой форме фосфофруктокпназы, нечувствительной к ингибированию продуктами аэробного обмена, либо потому, что интенсивность аэробного обмена очень низка. Как бы то ни было, наблюдаемое закисление крови, поступающей в капилляры эпителия, вполне может быть отнесено за счет образования молочной кислоты. Кроме того, в эпителии имеется высокоактивная карбоангидраза, которая, по-видимому, способствует образованию нонов Н+. [c.355]
Углеводный обмен во всякой живой клетке (живом веществе) представляет единый процесс одновременно протекающих связанных между собой реакций распада и синтеза органических веществ. В центре углеводного обмена у животных стоят гликогенсз и гликогенолиз, т. е. процессы образования и распада гликогена. Они протекают главным образом в печени. Гликоген может образоваться как из углеводов, так и из неуглеводных источников, таких, например, как некоторые аминокислоты, глицерин, молочная, пировиноградная и пропионовая кислоты, а также и из многих других простых соединений. Термин гликогенолиз обозначает собственно расщепление гликогена до глюкозы. Но теперь часто под этим словом понимают всю сумму процессов, ведущих к гликолитическому образованию молочной кислоты в том случае, когда исходным субстратом является не глюкоза, а гликоген. Под гликолизом понимают вообще процессы распада углеводов от начала, т. е. от глюкозы или гликогена, безразлично, и до конечных продуктов. [c.376]
При сопротивлениях, составляющих более 50 % максимальной изометрической силы, кровоток через мышцу резко уменьшается, что сопровождается появлением локальной гипоксии. В этих условиях (при дефиците аэробной энергопродукции) значительно исчерпываются алактатные анаэробные резервы и в мышцах накапливается большое количество свободного креатина, заметно усиливается образование молочной кислоты в результате гликолиза. Из-за дефицита макроэргических соединений при выполнении большого объема работы происходит разрушение мышечных белков и накопление продуктов их распада (низкомолекулярных пептидов, аминокислот и т. п.). Продукты расщепления белков, как и свободный креатин, служат активаторами белкового синтеза в период отдыха после скоростно-силовой работы, когда восстанавливается нормальное снабжение тканей кислородом и усиливается доставка к ним питательных веществ. Накопление молочной кислоты при предельной работе и вызванное этим изменение внутримышечного осмотического давления способствуют задержанию в мышцах межклеточной жидкости, богатой питательными веществами. При систематическом повторении таких тренировок в мышцах существенно увеличивается содержание сократительных белков и возрастает общий объем мышечной массы. [c.387]
При спиртовом брожении в процессе расщепления одной молекулы глюкозы образуется четыре молекулы АТФ (50 ккал, или 210 кдж). Из них две расходуются на функциональную деятельность и синтез. По расчетам некоторых авторов, при гликолизе и гликогенолизе в богатых энергией фосфорных связях аккумулируется 35—40 /о всей освобождающейся свободной энергни, остальные 60—65% рассеиваются в виде теплоты. Коэффициент полезного действия клеток, органов, работающих в анаэробных условиях, не превышает 0,4 (в аэробных 0,5). Эти расчеты основаны главны.м образом на данных, полученных на мышечных экстрактах и дрожжевом соке. В условиях живого организма мышечные клетки, органы и ткани утилизируют энергию, вероятно, значительно больше. С физиологической точки зрения процесс гликогенолиза и гликолиза имеет исключительно важное значение, особенно когда жизненные процессы осуществляются в условиях недостатка кислорода. Папример, при энергичной работе мышц, особенно в первой фазе деятельности, всегда наблюдается разрыв между доставкой кислорода в мышцы и его потребностью. В этом случае начальные энергетические затраты покрываются в значительной степени за счет гликогенолиза. Аналогичные явления наблюдаются при различных патологических состоя иях (гипоксия мозгз, сердца и т. п.). Кроме того, потенциальная энергия, заключенная в молочной кислоте, в конечном счете не теряется для высокоорганизованного организма. Образующаяся молочная кислота быстро пере.ходит из мышц в кровь и далее доставляется в печень, где снова превращается в гликоген. Анаэробный распад углеводов с образованием молочной кислоты очень распространен в природе он наблюдается не только в мышцах, но и в других тканях животного организма. [c.334]
Единство и теснейшая связь процессов брожения и дыхания растений, микроорганизмов и животных вытекают из того факта, что почти у всех живых организмов имеются одинаковые ферменты и те же основные промежуточные продукты, которые образуются в процессе их жизнедеятельности. Начальные этапы распада углеводов при анаэробном и аэробно.м дыхании одинаковы и начинаются с образования фосфорных эфиров глюкозы, именно глюкозо-1-фосфата, глюкозо-6-фосфата и фруктозо-1,6-дифосфата. Фосфорилирование глюкозы является необходимым условием как при аэробном распаде углеводов до углекислого газа и воды во время дыхания, так и при распаде углеводов в анаэробных условиях с образованием молочной кислоты и спирта. Пути аэробного и анаэробного распада углеводов расходятся на стадии образования пировиноградной кислоты в животные тканях или соответственно уксусного альдегида в дрожжевых клетках. Пировиноградная кислота занимает центральное положение в обмене углеводов. Она образуется из глюкозы (после фосфорилирования) или из гликогена (после фосфоролиза) путем нормального гликолиза. В анаэробных условиях пировиноградная кислота либо распадается в результате прямого декарбоксилирования, как это наблюдается в дрожжах, либо восстанавливается водородом до молочной кислоты, как это имеет место в мышцах. Спирт и молочная кислота являются конечными продуктами анаэробного обмена. В аэробных условиях пи-роаиноградная кислота полностью окисляется до углекислого газа и воды, [c.339]
Между гликолизом и аэробным окислением углеводов существует тесная связь. Эта связь заключается прежде всего в том, что первые стадии гликолиза и аэробного окисления углеводов одинаковы. Расхождение путей анаэробного и аэробного распада начинается на стадии дальнейшего превращения пировиноградной кислоты, которая в отсутствие кислорода восстанавливается за счет водорода восстановлен1юй кодегидразы (К0Ш2) с образованием молочной кислоты в присутствии же кислорода она подвергается окислительному декарбоксилированию. При аэробном окислении углеводов по пентозному циклу расхождение путей начинается на стадии образования глюкозо-6-фосфорной кислоты. [c.298]
Что же касается механизма реакции Пастера, то он остается еще недостаточно выясненным, хотя для его объяснения существует ряд гипотез. Одна из этих гипотез указывает на то, что прекращение гликолиза при аэробных условиях является скорее кажущимся, чем действительным. В присутствии кислорода в некоторых тканях, например в мышечной, часть образующейся при гликолизе молочной кислоты окисляется до углекислого газа и воды с освобождением энергии, которая используется частично для ресинтеза из оставшейся части молочной кислоты гликогена. Следовательно, в этом случае в тканях образование молочной кислоты не прекраш.ается в присутствии кислорода. Сбережение запасов гликогена достигается тем, что некоторая, и при этом большая, часть образовавшейся молочной кислоты в присутствии кислорода снова превращается в гликоген. Другие гипотезы объяс 1яют реакцию Пастера тем, что кислород прекращает гликолиз, воздействуя на ферменты, катализирующие тот пли иной этап гликолиза, прекращая, или тормозя, их действие. Некоторые ферменты гликолиза содержат важные для проявления их действия сульфгидрильные группы (—5Н). Среди этих ферментов находится и дегидраза фосфоглицеринальдегида. Кислород окислением сульфгидрильных групп ферментов может приостановить гликолиз. [c.298]
Глюкоза в моче
Анализ выявляет в моче глюкозу, появляющуюся при патологии почек, эндокринной системы, побочном действии лекарств, отравлениях, осложненном течении беременности.
Синонимы русские
Тест на глюкозурию.
Синонимы английские
Urine glucose, urine glucose test, urine sugar test, glucosuria test.
Метод исследования
Ферментативный УФ метод (гексокиназный).
Единицы измерения
Ммоль/л (миллимоль на литр).
Какой биоматериал можно использовать для исследования?
Разовую порцию мочи.
Как правильно подготовиться к исследованию?
- Не употреблять алкоголь в течение 24 часов до исследования.
- Прекратить прием мочегонных препаратов в течение 48 часов до сдачи мочи (по согласованию с врачом).
- Исключить физическое и эмоциональное перенапряжение в течение 30 минут до исследования.
Общая информация об исследовании
Глюкоза относится к классу углеводов (сахаров) и служит в организме универсальным источником энергии. Ее название происходит от греческого слова glykys – «сладкий».
Глюкоза является мономером, в природе и у человека встречается как в составе ди- и полимеров, так и отдельно. Основная роль глюкозы в организме – энергетическая – участие в биохимических реакциях, обеспечивающих клетки организма энергией, – также она выполняет структурную функцию, входя в состав различных молекул.
Углеводы поступают в организм с едой. На их долю при сбалансированном питании приходится 75 % объема и 50 % калорийности суточного рациона. Углеводы пищи представлены крахмалом (полимер глюкозы, содержащийся в растительной пище), гликогеном (полимер глюкозы, содержащийся в животной пище), сахарозой (димер, состоящий из фруктозы и сахарозы), моносахарами (лактозы молока, фруктозы и глюкозы меда и фруктов). Углеводы могут образовываться из липидов и аминокислот, но этот процесс приводит к появлению кетоновых тел и азотистых продуктов, в больших количествах и при длительном действии неблагоприятно сказывающихся на состоянии организма. Также в печени содержится резерв гликогена. В ротовой полости и кишечнике сахара расщепляются, всасываются и далее поступают в кровь. Концентрация глюкозы в крови поддерживается на постоянном уровне взаимодействием гормонов. Как повышение, так и понижение глюкозы в крови опасно – возможна гипер- и гипогликемическая кома. В почках глюкоза попадает в первичную мочу, образующуюся при прохождении крови через корковое вещество почек (почечные клубочки). В мозговом веществе почки (канальцевой части) происходит практически полный переход глюкозы из первичной мочи в кровь (реабсорбция) при условии, что уровень глюкозы крови ниже определенного порога. В мочу глюкоза не попадает.
Существует два основных фактора, обуславливающих появление глюкозы в моче: повышение глюкозы крови выше почечного порога (гипергликемия) и нарушение реабсорбции глюкозы в почках в связи с их поражением. Возможно сочетание этих причин.
При заболеваниях почек, нарушающих работу нефрона, происходит неполная реабсорбция глюкозы и она появляется в моче. Встречается первичное поражение канальцев (тубулопатия) – редкое наследственное заболевание, при котором страдает способность к реабсорции некоторых веществ в почечных канальцах (синдром де Тони – Дебре – Фанкони, первичная глюкозурия), и вторичное поражение, когда нарушается функция почек в целом (гломерулонефриты, почечная недостаточность, отравления). Глюкозурия, связанная с работой почек, может обнаруживаться у беременных, особенно на поздних сроках. В этом случае для предотвращения осложнений обязательно наблюдение врача.
Увеличение содержания глюкозы в крови ведет к его увеличению в первичной моче, при превышении определенного порога даже здоровые почки не полностью реабсорбируют глюкозу, так что она попадает в мочу. Глюкоза крови может повышаться у здоровых людей при чрезмерном употреблении углеводов в пищу, стрессе, эпизодическом употреблении некоторых лекарств. В целом уровень глюкозы крови зависит от гормональной регуляции, и его колебания свидетельствуют о патологии эндокринной системы.
Гормоны, влияющие на обмен углеводов, принято делить на инсулин и контринсулярные. Действие инсулина направлено на снижение уровня глюкозы в крови: он способствует переходу глюкозы в ткани, стимулирует синтез гликогена и угнетает его расщепление до глюкозы, угнетает образование глюкозы из аминокислот и липидов. Инсулин синтезируется эндокринными клетками поджелудочной железы. При сахарном диабете нарушается синтез инсулина (инсулинозависимый сахарный диабет) или реакция на него клеток организма (инсулинонезависимый сахарный диабет), при удалении поджелудочной железы или ее значительном повреждении при панкреатите также развивается недостаточность инсулина. Это ведет к повышению глюкозы в крови и появлению глюкозы в моче. При лечении диабета добиваются стабильного уровня глюкозы крови, а глюкозурия может являться сигналом к коррекции терапии.
Контринсулярные гормоны – это глюкагон поджелудочной железы, кортизол коркового вещества надпочечников, адреналин мозгового вещества надочечников, соматотропин передней доли гипофиза, гормоны щитовидной железы. Их действие имеет ряд особенностей, но в целом в том, что касается метаболизма глюкозы, оно противоположно инсулину: стимуляция распада гликогена и инсулина, глюкоза синтезируется из липидов и аминокислот, ее уровень в крови повышается. Действие контринсулярных гормонов направлено на удовлетворение энергетических потребностей в период стресса, мышечного напряжения. К гипергликемии и появлению глюкозы в моче ведет повышение уровня контринсулярных гормонов. Это происходит при длительном приеме соответствующих препаратов (глюкокортикоидов, гормонов щитовидной железы, соматотропного гормона), при гормонпродуцирующих опухолях щитовидной железы, надпочечников, поджелудочной железы, гипофиза.
Для чего используется исследование?
- Для выявления сахарного диабета.
- Для контроля за течением сахарного диабета.
- Чтобы оценить эффективность лечения сахарного диабета.
- Для оценки функции почек.
- Чтобы оценить функционирование эндокринной системы (поджелудочной, щитовидной железы, гипофиза, надпочечников).
- Для контроля за состоянием беременной женщины.
Когда назначается исследование?
- При подозрении на сахарный диабет.
- Когда необходимо оценить течение сахарного диабета и эффективность его лечения.
- При подозрении на нарушение функции почек
- Если в семье были случаи тубулопатий.
- При подозрении на эндокринные нарушения в организме (гипертиреоз, синдром и болезнь Иценко – Кушинга, феохромоцитому, акромегалию).
- При ведении беременности, особенно на поздних сроках.
Что означают результаты?
Референсные значения: 0 — 0,8 ммоль/л.
Причины повышения уровня глюкозы в моче:
- сахарный диабет,
- гипертиреоз,
- болезнь и синдром Иценко – Кушинга,
- феохромоцитома,
- акромегалия,
- наследственная тубулопатия (синдром Фанкони),
- почечная недостаточность,
- беременность,
- прием кортикостероидов и АКТГ,
- прием некоторых лекарств (седативные и обезболивающие средства),
- прием большого количества богатой углеводами пищи,
- стресс, мышечное напряжение, в том числе судороги.
Причины понижения уровня глюкозы в моче
У здоровых людей моча содержит крайне незначительное количество глюкозы. Полное исчезновение глюкозы происходит при бактериальных инфекциях мочевыводящих путей, однако на практике этот факт не учитывается в диагностических целях.
Имеет значение снижение глюкозы на фоне предшествующего повышения, что означает нормализацию уровня глюкозы в крови или стабилизацию функции почек.
Что может влиять на результат?
- У здоровых людей возможно повышение уровня глюкозы в крови и в моче после употребления богатой углеводами пищи, при стрессе, мышечном напряжении.
- На результат анализа влияют принимаемые лекарства.
- Однократное повышение глюкозы в моче не является основанием для постановки диагноза. Решение принимает врач с учетом клинической картины и анамнеза.
Кто назначает исследование?
Эндокринолог, нефролог, акушер-гинеколог, терапевт, педиатр.
Литература
- Зайчик А. Ш., Чурилов Л. П. Основы патохимии. СПб., 2001.
- Дедов И. И., Мельниченко Г. А., Фадеев В. Ф. Эндокринология. М., 2000 г.
- Вуд Мари Э., Бани Пол А.. Секреты гематологии и онкологии. СПб: 2001.
- Glucosuria. Cowart SL, Stachura ME. Boston: Butterworths; 1990.
- Ulster Med J. 2003 May; 72(1): 48-49. Hyperglycaemia, glycosuria and ketonuria may not be diabetes. J. Gray, A. Bhatti, and J. M. O’Donohoe.
Урок 10. углеводы. глюкоза. олигоса- хариды. сахароза — Химия — 10 класс
Химия, 10 класс
Урок № 10. Углеводы. Глюкоза. Олигосахариды. Сахароза
Перечень вопросов, рассматриваемых в теме: урок посвящён изучению углеводов, особенностям их строения. Рассмотрено влияние функциональных групп на свойства углеводов. Даётся характеристика химических свойств глюкозы и сахарозы. Объяснена биологическая роль углеводов и области их применения.
Глоссарий
Алкилирование – реакция образования простых эфиров в результате замещения атома водорода углеводородным радикалом в гидроксогруппе.
Ацилирование – реакция образования сложных эфиров в результате взаимодействия спиртов, в том числе многоатомных, с кислотами или кислотными ангидридами.
Брожение маслянокислое – превращение глюкозы под действием маслянокислых бактерий в масляную кислоту. Сопровождается выделением углекислого газа и водорода.
Брожение молочнокислое – превращение глюкозы под действием молочнокислых бактерий в молочную кислоту.
Брожение спиртовое – разложение глюкозы под действием дрожжей с образованием этилового спирта и углекислого газа.
Глюкоза – моносахарид состава С6Н12О6, состоящий из 6 атомов углерода, 5 гидроксильных групп и альдегидной группы. Может существовать как в виде линейной, так и циклической молекул. Вступает в реакции окисления, восстановления, ацилирования, алкилирования, подвергается молочнокислому, спиртовому, маслянокислому брожению.
Крахмал – полисахарид, состоящий из остатков α-глюкозы.
Лактоза, или молочный сахар – дисахарид С12Н22О11, состоящий из остатков глюкозы и галактозы, подвергается гидролизу, может окисляться до сахариновых кислот.
Моносахариды – углеводы, не подвергающиеся гидролизу, состоят из 3–10 атомов углерода, могут образовывать циклические молекулы с одним циклом (глюкоза, фруктоза, рибоза).
Невосстанавливающие углеводы – углеводы, не содержащие альдегидной группы и не способные к реакциям восстановления (фруктоза, сахароза, крахмал).
Олигосахариды – углеводы, образующие при гидролизе от 2 до 10 молекул моносахаридов (сахароза, лактоза).
Полисахариды – углеводы, образующие при гидролизе от нескольких десятков до сотен тысяч молекул моносахаридов (целлюлоза, крахмал).
Рибоза— моносахарид, относится к пентозам. Линейная молекула содержит альдегидную группу. Образует пятичленный цикл. Входит в состав РНК.
Сахароза – дисахарид, состоящий из остатков α-глюкозы и β-фруктозы. Относится к невосстанавливающим углеводам, так как не содержит альдегидную группу и не может восстанавливать гидроксид меди (II) до одновалентного оксида меди и серебро из аммиачного раствора гидроксида серебра. Является многоатомным спиртом. Подвергается гидролизу.
Углеводы – кислородсодержащие органические соединения, содержащие карбонильную и несколько гидроксильных групп.
Фруктоза – моносахарид состава С6Н12О6, относится к кетозам. Может существовать как в виде линейной молекулы, так и образовывать пятичленный цикл.
Целлюлоза – полисахарид, состоящий из остатков β-глюкозы.
Основная литература: Рудзитис, Г. Е., Фельдман, Ф. Г. Химия. 10 класс. Базовый уровень; учебник/ Г. Е. Рудзитис, Ф. Г, Фельдман – М.: Просвещение, 2018. – 224 с.
Дополнительная литература:
1. Рябов, М.А. Сборник задач, упражнений и тестов по химии. К учебникам Г.Е. Рудзитис, Ф.Г. Фельдман «Химия. 10 класс» и «Химия. 11 класс»: учебное пособие / М.А. Рябов. – М.: Экзамен. – 2013. – 256 с.
2. Рудзитис, Г.Е. Химия. 10 класс : учебное пособие для общеобразовательных организаций. Углублённый уровень / Г.Е. Рудзитис, Ф.Г. Фельдман. – М. : Просвещение. – 2018. – 352 с.
Открытые электронные ресурсы:
- Единое окно доступа к информационным ресурсам [Электронный ресурс]. М. 2005 – 2018. URL: http://window.edu.ru/ (дата обращения: 01.06.2018).
ТЕОРЕТИЧЕСКИЙ МАТЕРИАЛ ДЛЯ САМОСТОЯТЕЛЬНОГО ИЗУЧЕНИЯ
Понятие об углеводах, их классификация
Углеводами называются кислородсодержащие органические соединения, содержащие карбонильную и несколько гидроксильных групп и обычно отвечающие общей формуле Сп(Н2О)т. К углеводам относятся глюкоза, фруктоза, рибоза, сахароза, лактоза, крахмал, целлюлоза и другие. Углеводы могут существовать как в виде линейных, так и циклических молекул. Углеводы, молекулы которых могут образовывать только один цикл, называют моносахаридами (глюкоза, фруктоза, рибоза). Если молекула углевода при гидролизе распадается на несколько (от двух до десяти моносахаридов), они называются олигосахаридами (сахароза, лактоза). Углеводы, образующие при гидролизе десятки, сотни и более моносахаридов, называются полисахаридами (крахмал, целлюлоза).
Моносахариды
В молекуле моносахарида может быть от двух до десяти атомов углерода. Все моносахариды имеют окончание -оза. В названии сначала указывается количество атомов углерода, а затем прибавляется окончание: триоза, тетроза, пентоза, гексоза.
Для живых организмов наиболее важны пентоза и гексоза. Моносахариды с альдегидной группой называют альдозами (например, глюкоза), а содержащие кетогруппу – кетозами (например, фруктоза). Нумерация атомов углерода в альдозах начинается с атома альдегидной группы, а в кетозах – с крайнего атома, наиболее близкого к карбонильной группе.
Глюкоза
Самым распространённым моносахаридом в природе является глюкоза. Она содержится в сладких ягодах и фруктах. Мёд также содержит много глюкозы.
Глюкоза относится к группе гексоз, так как содержит шесть атомов углерода. Молекулы глюкозы могут быть как линейными (D-глюкоза, альдоза), так и циклическими (α и β-глюкоза). Линейная молекула глюкозы содержит на конце альдегидную группу. Общей формулой С6Н12О6 можно обозначить как глюкозу, так и фруктозу.
Фруктоза относится к кетозам и образуется пятичленный цикл. Она является изомером глюкозы. Фруктоза, также, как и глюкоза, может существовать в виде линейных и циклических молекул, в зависимости от положения заместителей у второго атома углерода различают α- и β-фруктозу.
Глюкоза – бесцветное кристаллическое вещество. Она хорошо растворяется в воде, имеет сладкий вкус. Факт наличия в молекуле глюкозы альдегидной группы доказывает реакция «серебряного зеркала». С фруктозой эта реакция не идёт. Один моль глюкозы реагирует с пятью молями уксусной кислоты с образованием сложного эфира, что доказывает наличие в молекуле глюкозы пяти гидроксильных групп. Такая реакция называется ацилированием. Если к раствору глюкозы на холоде добавить растворы сульфата меди и щелочи, то вместо осадка образуется ярко-синее окрашивание. Эта реакция доказывает, что глюкоза – многоатомный спирт. Благодаря наличию в молекуле глюкозы альдегидной группы, она может не только вступать в реакцию «серебряного зеркала», но и восстанавливать гидроксид меди (II) до одновалентного оксида. Водород в присутствии никелевого катализатора восстанавливает глюкозу до сорбита – шестиатомного спирта. В реакциях с низшими спиртами в кислой среде или с йодистым метилом в щелочной среде гидроксильные группы участвуют в образовании простых эфиров – происходит реакция алкилирования.
Глюкоза, в зависимости от условий, вступает в реакции брожения с образованием различных продуктов. Под действием молочнокислых бактерий глюкоза превращается в молочную кислоту – этот процесс получил название «молочнокислое брожение». Он используется при изготовлении кисломолочных продуктов. В присутствии дрожжей глюкоза подвергается спиртовому брожению. Этот вид брожения используется при изготовлении алкогольных напитков, а также дрожжевого теста. В этом процессе, кроме спирта, образуется углекислый газ, который делает тесто пышным. Брожение глюкозы, в результате которого образуется масляная кислота, происходит под действием особых маслянокислых бактерий. Этот вид брожения применяют в производстве масляной кислоты, эфиры которой широко используют в парфюмерии. Но если маслянокислые бактерии попадут в пищевые продукты, они могут вызвать их гниение.
Одним из продуктов фотосинтеза, который идет с участием зеленых растений, является глюкоза. Для человека и животных глюкоза является основным источником энергии для осуществления обменных процессов. В организмах животных глюкоза накапливается в виде гликогена (полисахарида, образованного остатками глюкозы). В растениях глюкоза превращается в крахмал (полисахарид, состоящий из остатков α-глюкозы). Клеточные оболочки высших растений построены из целлюлозы (полисахарид, состоящий из остатков β-глюкозы).
В крови человека находится около 0,1% глюкозы. Этой концентрации достаточно для снабжения организма энергией. Но при заболевании, называемом «сахарный диабет», глюкоза не расщепляется, её концентрация в крови может достигать 12%, что приводит к серьёзным нарушениям в работе всего организма.
В лабораторных условиях глюкозу можно получить из формальдегида в присутствии гидроксида кальция. Впервые этот синтез осуществил Александр Михайлович Бутлеров в 1861 году. В промышленности глюкозу получают гидролизом крахмала под действием серной кислоты.
Сахароза
Самым распространенным дисахаридом является сахароза. В природе она в большом количестве находится в свёкле и в сахарном тростнике. Молекула сахарозы состоит из остатков α-глюкозы и β-фруктозы.
Сахароза – бесцветное кристаллическое вещество, хорошо растворимое в воде, в два раза слаще глюкозы. Температура плавления равна 160 оС. В результате реакции сахарозы с гидроксидом меди появляется ярко-синее окрашивание, что характерно для многоатомных спиртов, но при нагревании раствора красный осадок не образуется, что указывает на отсутствие альдегидной группы. В присутствии минеральных кислот при нагревании сахароза подвергается гидролизу, распадаясь на α-глюкозу и β-фруктозу. Если к суспензии известкового молока прилить раствор сахарозы, то осадок растворяется. Образуется растворимый в воде сахарат кальция. Эта реакция лежит в основе получения сахарозы из сахарной свеклы и сахарного тростника. Если через раствор сахарата кальция пропустить углекислый газ, то образуется осадок карбоната кальция и раствор сахарозы.
Сахарозу применяют в пищевой промышленности для изготовления кондитерских изделий, консервирования (джемы, варенья, компоты).
ПРИМЕРЫ И РАЗБОР РЕШЕНИЙ ЗАДАЧ ТРЕНИРОВОЧНОГО МОДУЛЯ
1. Расчет количества реагента, необходимого для реакции с глюкозой
Условие задачи: Для получения ацетоуксусного эфира глюкозы на 1 моль глюкозы необходимо 5 моль уксусной кислоты. Сколько граммов 35%-ного раствора уксусной кислоты требуется, чтобы полностью прореагировать с 10 г глюкозы, если выход продукта реакции равен 75%?
Ответ запишите в виде целого числа.
Шаг первый: найдём молярные массы глюкозы и уксусной кислоты.
М(С6Н12О6) = 6·12 + 12·1 + 6·16 = 180 (г/моль).
М(СН3СООН) = 2·12 + 1·16 + 4·1 = 60 (г/моль).
Шаг второй: Найдём массу уксусной кислоты, которая вступает в реакцию с 10 г глюкозы. Для этого составим пропорцию:
180 г глюкозы реагирует с 5·60 г уксусной кислоты;
10 г глюкозы реагирует с х1 г уксусной кислоты.
(г).
Шаг третий: Найдём массу уксусной кислоты с учетом выхода продукта реакции. Для этого составим пропорцию:
16,7 г уксусной кислоты прореагирует с 75% глюкозы;
х2 г уксусной кислоты прореагирует со 100% глюкозы.
(г).
Шаг четвёртый: Найдём массу 35%-ного раствора уксусной кислоты, в котором содержится 22,2 г кислоты. Для этого составим пропорцию:
В 100 г раствора содержится 35 г кислоты;
в х3 г раствора содержится 22,2 г кислоты.
(г)
Ответ: 63
2. Расчёт количества энергии, полученной организмом при расщеплении глюкозы.
Условие задачи: В процессе расщепления 1 моль глюкозы в организме человека выделяется 200 кДж энергии. В сутки старшекласснику необходимо 12500 кДж энергии. Какой процент от суточной потребности в энергии восполнит ученик, съевший 200 г винограда, если содержание глюкозы в винограде составляет 30%? Ответ запишите с точностью до десятых долей.
Шаг первый: Найдём молярную массу глюкозы:
М(С6Н12О6) = 6·12 + 12·1 + 6·16 = 180 (г/моль).
Шаг второй: Найдём массу глюкозы, которая содержится в 200 г винограда.
Для этого массу винограда умножим на 30% и разделим на 100%:
г.
Шаг третий: Найдём количество моль глюкозы, которое содержится в 60 г этого углевода.
Для этого массу глюкозы разделим на её молярную массу:
(моль).
Шаг четвёртый: Найдём количество энергии, которая выделится при расщеплении 0,33 моль глюкозы.
Для этого составим пропорцию:
При расщеплении 1 моль глюкозы выделяется 200 кДж энергии;
при расщеплении 0,33 моль глюкозы выделяется х1 кДж энергии.
(кДж).
Шаг пятый: Найдём, какой процент от суточной потребности составляет это количество энергии.
Для этого составим пропорцию:
12500 кДж составляет 100% суточной потребности;
66 кДж составляет х2% суточной потребности.
(%).
Ответ: 0,5.
Этапы энергетического обмена
☰
Энергетический обмен — это по-этапный распад сложных органических соединений, протекающий с выделением энергии, которая запасается в макроэргических связях молекул АТФ и используется потом в процессе жизнедеятельности клетки, в том числе на биосинтез, т.е. пластический обмен.
В аэробных организмах выделяют три последовательных этапа энергетического обмена:
- Подготовительный — расщепление биополимеров до мономеров.
- Бескислородный — гликолиз — расщепление глюкозы до пировиноградной кислоты.
- Кислородный — расщепление пировиноградной кислоты до углекислого газа и воды.
Подготовительный этап
На подготовительном этапе энергетического обмена происходит расщепление поступивших с пищей органических соединений на более простые, обычно мономеры. Так углеводы расщепляются до сахаров, в том числе глюкозы; белки — до аминокислот; жиры — до глицерина и жирных кислот.
Хотя при этом выделяется энергия, она не запасается в АТФ и, следовательно, не может быть использована впоследствии. Энергия рассеивается в виде тепла.
Расщепление полимеров у многоклеточных сложноорганизованных животных протекает в пищеварительном тракте под действием выделяющихся сюда железами ферментов. Затем образовавшиеся мономеры всасываются в кровь в основном через кишечник. Уже кровью питательные вещества разносятся по клеткам.
При этом не все вещества разлагаются до мономеров в пищеварительной системе. Расщепление многих происходит непосредственно в клетках, в их лизосомах. У одноклеточных организмов поглощенные вещества попадают в пищеварительные вакуоли, где и перевариваются.
Образовавшиеся мономеры могут использоваться как для энергетического, так и пластического обмена. В первом случае они расщепляются, во-втором – из них синтезируются компоненты самих клеток.
Бескислородный этап энергетического обмена
Бескислородный этап протекает в цитоплазме клеток и в случае аэробных организмов включает только гликолиз — ферментативное многоступенчатое окисление глюкозы и ее расщепление до пировиноградной кислоты, которую также называют пируватом.
Молекула глюкозы включает шесть атомов углерода. При гликолизе она расщепляется до двух молекул пирувата, который включает три атома углерода. При этом отщепляется часть атомов водорода, которые передаются на кофермент НАД, который, в свою очередь, потом будет участвовать в кислородном этапе.
Часть выделяющейся при гликолизе энергии запасается в молекулах АТФ. На одну молекулу глюкозы синтезируется всего две молекулы АТФ.
Энергия, оставшаяся в пирувате, запасенная в НАД, у аэробов далее будет извлечена на следующем этапе энергетического обмена.
В анаэробных условиях, когда кислородный этап клеточного дыхания отсутствует, пируват «обезвреживается» в молочную кислоту или подвергается брожению. При этом энергия не запасается. Таким образом, здесь полезный энергетический выход обеспечивается только малоэффектвным гликолизом.
Кислородный этап
Кислородный этап протекает в митохондриях. В нем выделяют два подэтапа: цикл Кребса и окислительное фосфорилирование. Поступающий в клетки кислород используется только на втором. В цикле Кребса происходит образование и выделение углекислого газа.
Цикл Кребса протекает в матриксе митохондрий, осуществляется множеством ферментов. В него поступает не сама молекула пировиноградной кислоты (или жирной кислоты, аминокислоты), а отделившаяся от нее с помощью кофермента-А ацетильная группа, включающая два атома углерода бывшего пирувата. За многоступенчатый цикл Кребса происходит расщепление ацетильной группы до двух молекул CO2 и атомов водорода. Водород соединяется с НАД и ФАД. Также происходит синтез молекулы ГДФ, приводящей к синтезу потом АТФ.
На одну молекулу глюкозы, из которой образуется два пирувата, приходится два цикла Кребса. Таким образом, образуется две молекулы АТФ. Если бы энергетический обмен заканчивался здесь, то суммарно расщепление молекулы глюкозы давало бы 4 молекулы АТФ (две от гликолиза).
Окислительное фосфорилирование протекает на кристах – выростах внутренней мембраны митохондрий. Его обеспечивает конвейер ферментов и коферментов, образующий так называемую дыхательную цепь, заканчивающуюся ферментом АТФ-синтетазой.
По дыхательной цепи происходит передача водорода и электронов, поступивших в нее от коферментов НАД и ФАД. Передача осуществляется таким образом, что протоны водорода накапливаются с внешней стороны внутренней мембраны митохондрий, а последние ферменты в цепи передают только электроны.
В конечном итоге электроны передаются молекулам кислорода, находящимся с внутренней стороны мембраны, в результате чего они заряжаются отрицательно. Возникает критический уровень градиента электрического потенциала, приводящий к перемещению протонов через каналы АТФ-синтетазы. Энергия движения протонов водорода используется для синтеза молекул АТФ, а сами протоны соединяются с анионами кислорода с образованием молекул воды.
Энергетический выход функционирования дыхательной цепи, выраженный в молекулах АТФ, велик и суммарно составляет от 32 до 34 молекул АТФ на одну исходную молекулу глюкозы.
Каковы четыре фазы полного распада глюкозы?
Глюкоза — это шестиуглеродный сахар, который можно проглатывать или вводить непосредственно в организм, но чаще он является побочным продуктом метаболизма сложных углеводов, белков или жиров. Глюкозу можно использовать для синтеза гликогена и других топлив для хранения или дальнейшего расщепления для обеспечения энергией метаболических процессов — серии реакций, которые в совокупности называются клеточным дыханием. Стадии распада глюкозы можно разделить на четыре отдельные фазы.
Гликолиз
Первоначальный распад глюкозы происходит в цитоплазме клетки. Это анаэробная реакция клеточного дыхания, означающая, что она не требует кислорода. Здесь, в серии из восьми отдельных реакций, шестиуглеродная молекула глюкозы метаболизируется с использованием двух молекул аденозинтрифосфата (АТФ) с образованием двух трехуглеродных молекул пирувата, двух молекул H 2 O (воды) и четырех молекул АТФ для чистый прирост двух молекул АТФ.АТФ является основным источником энергии в метаболизме человека.
Подготовительная реакция
Эта реакция происходит в матриксе или внутри митохондрий клеток. Здесь две молекулы пирувата от гликолиза объединяются с двумя молекулами кофермента A (CoA) с образованием двух молекул ацетил-CoA и двух молекул диоксида углерода (CO 2 ). Эта реакция протекает в одну стадию и, как и гликолиз, является анаэробной.
Цикл лимонной кислоты
Также называемый циклом трикарбоновой кислоты (ТСА) или циклом Кребса, эта серия анаэробных реакций, как и подготовительная реакция, протекает в матриксе митохондрий.Здесь две молекулы ацетил-КоА из подготовительной реакции объединяются с рядом фосфатных и нуклеотидных компонентов с образованием двух АТФ, четырех СО2 и ряда нуклеотидных посредников. Эти посредники имеют решающее значение для аэробного дыхания, которое происходит на следующей стадии расщепления глюкозы.
Цепь переноса электронов
На этом этапе, который происходит на внутренних мембранах митохондрий, наконец, в картину входит кислород. Транспортерами на этой схеме являются молекулы НАД и ФАД, нуклеотидных посредников, упомянутых выше.В присутствии шести молекул кислорода протоны передаются от НАД и ФАД к другим молекулам НАД и ФАД вниз по цепи, позволяя экстрагировать АТФ в различных точках. В результате получается 34 молекулы АТФ.
Обратите внимание, что после этой стадии общая химическая реакция гликолиза кажется завершенной:
C 6 H 12 O 6 + 6O 2 -> 6CO 2 + 6H 2 O + 38 ATP
Какой продукт распада глюкозы имеет больше всего энергии?
Очевидно, что с двумя АТФ от гликолиза, двумя из цикла лимонной кислоты и 34 из цепи переноса электронов на молекулу глюкозы, цепь переноса электронов на сегодняшний день является наиболее энергоемкой.Вот почему люди не могут быть лишены кислорода на долгое время и почему очень интенсивные (анаэробные) упражнения не могут продолжаться более нескольких минут: большинство физиологических функций зависит от постоянного использования цепи переноса электронов.
Энергия | Безграничная микробиология
Метаболизм углеводов
Организмы расщепляют углеводы для производства энергии для клеточных процессов, а фотосинтезирующие растения производят углеводы.
Задачи обучения
Анализировать важность углеводного обмена для производства энергии
Основные выводы
Ключевые моменты
- Распад глюкозы, которую живые организмы используют для производства энергии, описывается уравнением: [латекс] {\ text {C}} _ {6} {\ text {H}} _ {12} {\ text {O}} _ {6} +6 {\ text {O}} _ {2} \ rightarrow 6 {\ text {CO}} _ {2} +6 {\ text {H}} _ {2} \ text {O} + \ text {энергия} [/ латекс].
- Процесс фотосинтеза, который растения используют для синтеза глюкозы, описывается уравнением: [латекс] 6 \ text {CO} _ {2} +6 {\ text {H}} _ {2} \ text {O} + \ text { энергия} \ rightarrow {\ text {C}} _ {6} {\ text {H}} _ {12} {\ text {O}} _ {6} +6 \ text {O} _ {2} [/ латекс].
- Потребляемая глюкоза используется для производства энергии в виде АТФ, который используется для выполнения работы и химических реакций в клетке.
- Во время фотосинтеза растения превращают световую энергию в химическую энергию, которая используется для создания молекул глюкозы.
Ключевые термины
- аденозинтрифосфат : многофункциональный нуклеозидтрифосфат, используемый в клетках в качестве кофермента, часто называемый «молекулярной единицей энергетической валюты» при внутриклеточном переносе энергии
- глюкоза : простой моносахарид (сахар) с молекулярной формулой [латекс] {\ text {C}} _ {6} {\ text {H}} _ {12} {\ text {O}} _ { 6} [/ латекс] C6h22O6; это основной источник энергии для клеточного метаболизма
Метаболизм углеводов
Углеводы — одна из основных форм энергии для животных и растений.Растения вырабатывают углеводы, используя световую энергию солнца (в процессе фотосинтеза), в то время как животные едят растения или других животных для получения углеводов. Растения хранят углеводы в длинных полисахаридных цепях, называемых крахмалом, в то время как животные хранят углеводы в виде молекулы гликогена. Эти большие полисахариды содержат много химических связей и, следовательно, хранят много химической энергии. Когда эти молекулы расщепляются во время метаболизма, энергия химических связей высвобождается и может быть использована для клеточных процессов.
Все живые существа используют углеводы как форму энергии. : Растения, такие как дуб и желудь, используют энергию солнечного света для производства сахара и других органических молекул. И растения, и животные (например, эта белка) используют клеточное дыхание для получения энергии из органических молекул, изначально производимых растениями
Производство энергии из углеводов (клеточное дыхание)
Метаболизм любого моносахарида (простого сахара) может производить энергию для использования клеткой.Избыточные углеводы хранятся в виде крахмала в растениях и в виде гликогена у животных, готовые к метаболизму, если потребность организма в энергии внезапно возрастет. Когда эта потребность в энергии увеличивается, углеводы расщепляются на составляющие моносахариды, которые затем распределяются по всем живым клеткам организма. Глюкоза (C 6 H 12 O 6 ) является типичным примером моносахаридов, используемых для производства энергии.
Внутри клетки каждая молекула сахара расщепляется в ходе сложной серии химических реакций.Поскольку химическая энергия высвобождается из связей в моносахариде, она используется для синтеза высокоэнергетических молекул аденозинтрифосфата (АТФ). АТФ — это основная энергетическая валюта всех клеток. Точно так же, как доллар используется в качестве валюты для покупки товаров, клетки используют молекулы АТФ для немедленной работы и проведения химических реакций.
Распад глюкозы в процессе метаболизма, называемый клеточным дыханием, можно описать уравнением:
[латекс] {\ text {C}} _ {6} {\ text {H}} _ {12} {\ text {O}} _ {6} +6 {\ text {O}} _ {2} \ rightarrow 6 {\ text {CO}} _ {2} +6 {\ text {H}} _ {2} \ text {O} + \ text {energy} [/ latex]
Производство углеводов (фотосинтез)
Растения и некоторые другие виды организмов производят углеводы в процессе, называемом фотосинтезом.Во время фотосинтеза растения преобразуют световую энергию в химическую энергию, превращая молекулы углекислого газа (CO 2 ) в молекулы сахара, такие как глюкоза. Поскольку этот процесс включает в себя построение связей для синтеза большой молекулы, для продолжения требуется ввод энергии (света). Синтез глюкозы путем фотосинтеза описывается этим уравнением (обратите внимание, что оно является обратным предыдущему уравнению):
[латекс] 6 \ text {CO} _ {2} +6 {\ text {H}} _ {2} \ text {O} + \ text {energy} \ rightarrow {\ text {C}} _ {6 } {\ text {H}} _ {12} {\ text {O}} _ {6} +6 \ text {O} _ {2} [/ latex]
В рамках химических процессов растений молекулы глюкозы могут объединяться с другими типами сахаров и превращаться в них.В растениях глюкоза хранится в форме крахмала, который может снова расщепляться на глюкозу посредством клеточного дыхания, чтобы поставлять АТФ.
Изменения свободной энергии в химических реакциях
ΔG определяет направление и степень химического изменения.
Задачи обучения
Вспомните возможные изменения свободной энергии для химических реакций.
Основные выводы
Ключевые моменты
- Если свободная энергия реагентов больше, чем у продуктов, энтропия мира будет увеличиваться, когда реакция происходит, как написано, и поэтому реакция будет иметь тенденцию происходить спонтанно.
- Если свободная энергия продуктов превышает свободную энергию реагентов, то реакция не происходит.
- Важным следствием одностороннего нисходящего пути свободной энергии является то, что как только она достигает минимально возможного значения, чистое изменение прекращается.
- При спонтанном изменении энергия Гиббса всегда уменьшается и никогда не увеличивается.
Ключевые термины
- спонтанное изменение : Спонтанный процесс — это эволюция системы во времени, в которой она высвобождает свободную энергию (обычно в виде тепла) и переходит в более низкое, более термодинамически стабильное энергетическое состояние.
Направление и масштабы химических изменений
ΔG определяет направление и степень химического изменения. Помните, что ΔG имеет значение только для изменений, при которых температура и давление остаются постоянными. Это условия, при которых большинство реакций проводится в лаборатории. Система обычно открыта для атмосферы (постоянное давление), и процесс начинается и заканчивается при комнатной температуре (после того, как все тепло, которое было добавлено или высвободилось в результате реакции, рассеивается.)
Важность функции Гиббса трудно переоценить: она определяет, возможно ли данное химическое изменение термодинамически. Таким образом, если свободная энергия реагентов больше, чем у продуктов, энтропия мира будет увеличиваться, и реакция протекает самопроизвольно. И наоборот, если свободная энергия продуктов превышает свободную энергию реагентов, реакция не состоится.
При спонтанном изменении энергия Гиббса всегда уменьшается и никогда не увеличивается.Это, конечно, отражает тот факт, что энтропия мира ведет себя прямо противоположным образом (из-за отрицательного знака в члене TΔS). Вот пример:
[латекс] {\ text {H}} _ {2} \ text {O} (\ text {liquid}) \ rightarrow {\ text {H}} _ {2} \ text {O} (\ text {ice }) [/ латекс]
Вода с температурой ниже нуля градусов Цельсия испытывает уменьшение своей энтропии, но тепло, выделяемое в окружающую среду, более чем компенсирует это, поэтому энтропия мира увеличивается, свободная энергия H 2 O уменьшается, и процесс происходит самопроизвольно. .
Важным следствием одностороннего нисходящего пути свободной энергии является то, что как только она достигает минимально возможного значения, чистое изменение прекращается. Это, конечно, представляет собой состояние химического равновесия. Эти отношения резюмируются следующим образом:
- [латекс] \ Delta G <0 [/ латекс]: реакция будет происходить спонтанно справа.
- [латекс] \ Delta G> 0 [/ latex]: реакция будет происходить спонтанно слева.
- [латекс] \ Delta G = 0 [/ latex]: реакция находится в равновесии и не будет идти ни в одном направлении.
Условия самопроизвольного изменения
Напомним условие самопроизвольного изменения:
ΔG = ΔH — TΔS <0
, где ΔG = изменение свободной энергии Гиббса, ΔH = изменение энтальпии, T = абсолютная температура и ΔS = изменение энтропии
Очевидно, что температурная зависимость ΔG почти полностью зависит от изменения энтропии, связанного с процессом. (уместно сказать «почти», потому что значения ΔH и ΔS сами по себе немного зависят от температуры; оба постепенно увеличиваются с температурой).В частности, обратите внимание, что в приведенном выше уравнении знак изменения энтропии определяет, станет ли реакция более или менее спонтанной при повышении температуры.
Для любой данной реакции знак ΔH также может быть положительным или отрицательным. Это означает, что существует четыре возможности влияния температуры на спонтанность процесса:
Случай 1: ΔH
<0 и ΔS> 0В этих условиях оба члена ΔH и TΔS будут отрицательными, поэтому ΔG будет отрицательным независимо от температуры.Экзотермическая реакция, энтропия которой возрастает, будет спонтанной при всех температурах.
Случай 2: ΔH
<0 и ΔS <0Если реакция достаточно экзотермична, она может привести к отрицательному значению ΔG только при температурах, ниже которых | TΔS | <| ΔH |. Это означает, что существует температура, определяемая [латексом] T = \ frac {\ Delta H} {\ Delta S} [/ latex], при которой реакция находится в равновесии; реакция будет происходить самопроизвольно только ниже этой температуры. Замерзание жидкости или конденсация газа - наиболее распространенные примеры этого состояния.
Случай 3: ΔH> 0 и ΔS> 0
Это обратное предыдущему случаю; увеличение энтропии должно преодолеть недостаток эндотермического процесса, так что TΔS> ΔH. Поскольку влияние температуры должно «усилить» влияние положительного ΔS, процесс будет самопроизвольным при температурах выше [латекса] T = \ frac {\ Delta H} {\ Delta S} [/ latex]. (Подумайте о плавлении и кипении.)
Случай 4: ΔH> 0 и ΔS
<0Поскольку оба ΔH и ΔS работают против этого, этот вид процесса не будет происходить самопроизвольно при любой температуре.Вещество А всегда имеет большее количество доступных энергетических состояний и поэтому всегда является предпочтительной формой.
Внутренняя энергия и энтальпия
Энтальпия реакции измеряет тепло, выделяемое / поглощаемое реакцией, которая происходит при постоянном давлении.
Задачи обучения
Проверить энтальпию реакции
Основные выводы
Ключевые моменты
- При постоянном объеме теплота реакции равна изменению внутренней энергии системы.
- При постоянном давлении теплота реакции равна изменению энтальпии системы.
- Большинство химических реакций происходит при постоянном давлении, поэтому энтальпия чаще используется для измерения теплоты реакции, чем внутренняя энергия.
Ключевые термины
- энтальпия : В термодинамике, мера теплосодержания химической или физической системы.
- внутренняя энергия : свойство, характеризующее состояние термодинамической системы, изменение которого равно поглощенному теплу за вычетом работы, совершаемой системой.
- первый закон термодинамики : Тепло и работа являются формами передачи энергии; внутренняя энергия замкнутой системы изменяется по мере того, как тепло и работа передаются в нее или из нее.
В термодинамике работа ( W ) определяется как процесс передачи энергии от одной системы к другой. Первый закон термодинамики гласит, что энергия замкнутой системы равна количеству тепла, подводимого к системе, за вычетом количества работы, выполняемой системой над своим окружением.Количество энергии для замкнутой системы записывается следующим образом:
[латекс] \ Delta U = Q — W [/ латекс]
В этом уравнении U — это полная энергия системы, Q — тепло и Вт — работа. В химических системах наиболее распространенным типом работы является работа давления-объема ( PV ), при которой объем газа изменяется. Подставляя это для работы в приведенное выше уравнение, мы можем определить изменение внутренней энергии для химической системы:
[латекс] \ Delta U = Q-P \ Delta V [/ латекс]
Изменение внутренней энергии при постоянном объеме
Давайте исследуем изменение внутренней энергии [латекс] \ Delta U [/ латекс] при постоянном объеме.При постоянном объеме [латекс] \ Delta V = 0 [/ латекс] уравнение для изменения внутренней энергии сводится к следующему:
[латекс] \ Delta U = Q_V [/ латекс]
Нижний индекс V добавлен к Q , чтобы указать, что это теплопередача, связанная с химическим процессом при постоянном объеме. Однако эту внутреннюю энергию часто очень трудно вычислить в реальных условиях, потому что химики, как правило, проводят свои реакции в открытых колбах и стаканах, которые позволяют газам уходить в атмосферу.Следовательно, объем не поддерживается постоянным, и вычисление [латекс] \ Delta U [/ латекс] становится проблематичным. Чтобы исправить это, мы вводим понятие энтальпии , которое гораздо чаще используется химиками.
Стандартная энтальпия реакции
Энтальпия реакции определяется как внутренняя энергия реакционной системы, плюс произведение давления и объема. Выдается:
[латекс] H = U + PV [/ латекс]
Добавляя термин PV , становится возможным измерить изменение энергии в химической системе, даже если эта система действительно воздействует на окружающую среду.Чаще всего нас интересует изменение энтальпии данной реакции, которое можно выразить следующим образом:
[латекс] \ Delta H = \ Delta U + P \ Delta V [/ латекс]
Когда вы запускаете химическую реакцию в лаборатории, реакция происходит при постоянном давлении, потому что атмосферное давление вокруг нас относительно постоянно. Мы рассмотрим изменение энтальпии для реакции при постоянном давлении, чтобы понять, почему энтальпия является таким полезным понятием для химиков.
Энтальпия реакции при постоянном давлении
Давайте еще раз посмотрим на изменение энтальпии для данного химического процесса.Выдается следующим образом:
[латекс] \ Delta H = \ Delta U + P \ Delta V [/ латекс]
Однако мы также знаем, что:
[латекс] \ Delta U = Q-W = Q-P \ Delta V [/ латекс]
Подставляя для объединения этих двух уравнений, получаем:
[латекс] \ Delta H = Q-P \ Delta V + P \ Delta V = Q_P [/ latex]
Таким образом, при постоянном давлении изменение энтальпии просто равно количеству тепла, выделяемого / поглощаемого реакцией. Из-за этой связи изменение энтальпии часто называют просто «теплотой реакции».”
Энтальпия : Объяснение того, почему энтальпию можно рассматривать как «теплосодержание» в системе постоянного давления.
Каково химическое уравнение клеточного дыхания?
Важно знать, что приведенное выше уравнение является суммарным уравнением. Процесс клеточного дыхания включает в себя множество различных стадий (реакций) расщепления глюкозы с использованием кислорода для производства углекислого газа, воды и энергии в форме АТФ.
6 атомов углерода , присутствующие в молекуле глюкозы, позволяют образовывать 6 молекул диоксида углерода .
12 атомов водорода в глюкозе делают возможным образование 6 молекул воды .
Чтобы уравновесить атомы кислорода на стороне реагента, вам нужно сосчитать 6 атомов глюкозы. Чтобы образовать 6 молекул углекислого газа и 6 молекул воды, у вас будет 18 атомов кислорода на стороне продукта (6 * 2) + (6 * 1) = 18.Чтобы получить 18 атомов кислорода на стороне реагента, вам понадобятся дополнительные 12 атомов кислорода из кислорода # O_2 #, чтобы уравновесить числа.
Процесс клеточного дыхания будет производить 36 молекул АТФ у эукариот (растений / животных и т. Д.) На каждую молекулу глюкозы. Этот процесс будет производить 38 молекул АТФ на каждую глюкозу у прокариот (бактерий).
Причина, по которой эукариоты производят меньшее количество АТФ, заключается в том, что им нужно использовать энергию для перемещения пирувата (из гликолиза), необходимого для цикла Кребса, в митохондрии.
В этом видео дается краткий обзор дыхания и обсуждается лаборатория, в которой проверяется, как температура может изменять скорость дыхания дрожжей. В теплых условиях процесс протекает быстрее из-за большего движения частиц.
Это видео дает более подробное обсуждение темы клеточного дыхания.
Надеюсь, это поможет!
При окислении в организме в процессе, называемом метаболизмом, глюкоза производит углекислый газ, воду и некоторые соединения азота и в процессе дает энергию, которая может использоваться клетками. Выход энергии составляет около 686 килокалорий (2870 килоджоулей) на моль, которую можно использовать для работы или для поддержания тепла. Этот показатель энергии представляет собой изменение свободной энергии Гиббса ΔG в реакции, меру максимального количества работы, получаемой в результате реакции.В качестве основного источника энергии в организме он не требует переваривания и часто вводится внутривенно пациентам в больницах в качестве питательного вещества. Энергия глюкозы получается в результате реакции окисления C 6 H 12 O 6 + 6O 2 -> 6CO 2 + 6H 2 O, где моль глюкозы (около 180 грамм) реагирует с шестью молями O 2 с выходом энергии ΔG = 2870 кДж. Шесть молей кислорода в STP занимают 6 x 22.4л = 134 л. Энергетический выход глюкозы часто выражается как выход кислорода на литр, что составляет 5,1 ккал на литр или 21,4 кДж на литр. Этот выход энергии можно измерить, фактически сжигая глюкозу и измеряя выделяемую энергию в калориметре. Но в живых организмах окисление глюкозы способствует ряду сложных биохимических реакций, которые обеспечивают энергию, необходимую клеткам. Первым шагом в расщеплении глюкозы во всех клетках является гликолиз, производящий пируват, который является отправной точкой для всех других процессов клеточного дыхания.В клетках, где присутствует кислород (аэробное дыхание), эти процессы были смоделированы в TCA или цикле Кребса. Основная часть использования энергии окисления глюкозы — это преобразование АДФ в АТФ, при этом богатая энергией молекула АТФ впоследствии используется в качестве энергетической валюты клетки. Глюкоза производится растениями с помощью солнечной энергии в процессе, называемом фотосинтезом. Этот синтез осуществляется на небольших энергетических фабриках, называемых хлоропластами в листьях растений.Хлоропласты захватывают энергию света и производят молекулы глюкозы из углекислого газа из воздуха и воды из почвы.
| Индекс Биохимические концепции Химические концепции Ссылки Tillery, Enger and Ross Tuszynski and Dixon |
Ch 15
.2: Цикл лимонной кислоты
Результаты обучения
- Опишите цикл лимонной кислоты (цикл Кребса).
- Назовите продукты цикла лимонной кислоты.
- Определите молекулы энергоносителя, образующиеся в цикле лимонной кислоты.
- Опишите, что происходит с пируватом до того, как он войдет в цикл лимонной кислоты.
Аэробное дыхание
Заманчивые подсказки — вулканические газы, обширные отложения железной руды и пузырьки древнего воздуха, заключенные в янтарь, — предполагают драматические изменения в истории земной атмосферы.Сопоставление этих ключей с летописью окаменелостей приводит к двум важным выводам: ранняя жизнь развивалась в отсутствие кислорода, и что кислород впервые появился между 2 и 3 миллиардами лет назад (см. Рисунок ниже) из-за фотосинтеза сине-зелеными бактериями, цианобактериями. . Химия клеточного дыхания отражает эту историю. Его первая стадия — гликолиз — универсальна и не требует использования кислорода.
Абсолютно зависимый от газообразного кислорода, нам трудно представить, что его появление должно было иметь катастрофические последствия для анаэробных организмов, которые развивались в его отсутствие.Но кислород очень реактивен, и сначала его влияние на эволюцию было настолько отрицательным, что некоторые назвали этот период «кислородной катастрофой». Однако, когда кислород постепенно сформировал защитный озоновый слой , жизнь восстановилась. После того, как первые организмы научились использовать кислород в своих интересах, разнообразие аэробных организмов резко возросло. Согласно Теории эндосимбиоза, поглощение некоторых из этих аэробных бактерий привело к появлению эукариотических клеток с митохондриями, и многоклеточность, , последовала эволюция многоклеточных эукариотических организмов.Сегодня мы живем в атмосфере, состоящей из \ (21 \% \) кислорода, и большая часть жизни следует за гликолизом с двумя последними, аэробными стадиями клеточного дыхания.
Вспомните цель клеточного дыхания: высвобождение энергии из глюкозы для производства АТФ, универсальной молекулы энергии для клеточной работы. Следующее уравнение описывает весь процесс, хотя и суммирует многие отдельные химические реакции.
\ [\ ce {6O2} + \ underbrace {\ ce {C6h22O6}} _ {\ text {хранимое химическое вещество} \\ \ text {энергия, срок поставки}} + \ ce {398P_i} \ ce {-> [\ text {митохондии}]} \ ce {38 АТФ} + \ ce {6O2} + \ ce {6h3O} \]
И снова первая стадия этого процесса, гликолиз, является древней, универсальной и анаэробной.В цитоплазме большинства клеток гликолиз расщепляет каждую 6-углеродную молекулу глюкозы на две 3-углеродные молекулы пирувата . Химическая энергия, которая была сохранена в разорванных связях, передается 2 молекулам АТФ и 2 молекулам НАДН.
Судьба пирувата зависит от вида и наличия или отсутствия кислорода. Если кислород присутствует для запуска последующей реакции, пируват попадает в митохондрии, где цикл лимонной кислоты (также известный как цикл Кребса ) (стадия 2) и цепь переноса электронов (стадия 3) расщепляют его и окисляют. это полностью в \ (\ ce {CO_2} \) и \ (\ ce {H_2O} \).Выделяемая энергия строит намного больше молекул АТФ, хотя, конечно, часть теряется в виде тепла. Давайте подробно рассмотрим, как митохондрии используют кислород для производства большего количества АТФ из глюкозы путем аэробного дыхания.
Цикл лимонной кислоты: получение энергии из пирувата
Аэробное дыхание начинается с попадания в митохондрии продукта гликолиза — пирувата. На каждую исходную молекулу глюкозы в митохондрии попадут две молекулы пирувата. Пируват, однако, не входит в цикл лимонной кислоты.Перед тем, как войти в этот цикл, пируват должен быть преобразован в 2-углеродную ацетил-коэнзим А (ацетил-КоА) звено.
Рисунок \ (\ PageIndex {1} \): после гликолиза два 3-углеродных пирувата попадают в митохондрии, где они превращаются в две 2-углеродные молекулы ацетил-коэнзима А (КоА). Затем ацетил-КоА вступает в цикл Кребса. Обратите внимание, что удаленный углерод превращается в двуокись углерода, составляющую два из шести таких конечных продуктов окисления глюкозы. Энергия, выделяемая при этом распаде, переносится НАДН.Превращение пирувата в ацетил-КоА называется реакцией пируватдегидрогеназы. Это катализируется комплексом пируватдегидрогеназы. Этот процесс производит один электронный носитель НАДН, высвобождая молекулу \ (\ ce {CO_2} \). Эта стадия также известна как связующая реакция или стадия перехода, поскольку она связывает гликолиз и цикл лимонной кислоты. Конечно, поскольку два пирувата образуются в результате гликолиза, образуются два ацетил-КоА, а также 2 молекулы НАДН.
Рисунок \ (\ PageIndex {2} \): Цикл Кребса завершает расщепление глюкозы, начавшееся при гликолизе.+} \), один FADH \ (_ 2 \) и один ATP (из GTP) и выпускает 2 \ (\ ce {CO_2} \). Цикл Кребса также известен как цикл лимонной кислоты или цикл трикарбоновой кислоты (цикл TCA).- В митохондриях каждый пируват расщепляется и объединяется с коферментом, известным как КоА, с образованием 2-углеродной молекулы, ацетил-КоА, которая может вступать в цикл Кребса. Один атом углерода (на пируват) «теряется» в виде диоксида углерода. Энергия, выделяющаяся при этом пробое, улавливается двумя молекулами НАДН. См. Рисунок выше.Жирные кислоты также могут распадаться на ацетил-КоА. Таким образом, липиды, как и жиры, можно «сжечь», чтобы получить АТФ, используя цикл лимонной кислоты.
- Цикл Кребса (см. Рисунок выше) начинается с объединения каждого ацетил-КоА с четырехуглеродной молекулой-носителем с образованием 6-углеродной молекулы лимонной кислоты (или цитрата, его ионизированной формы).
- Цикл переносит лимонную кислоту через ряд химических реакций, которые постепенно высвобождают энергию и захватывают ее в нескольких молекулах-носителях.+ \) восстанавливаются до NADH, одна молекула FAD (другой временный носитель энергии) восстанавливается до \ ( FADH_2 \) , и образуется одна молекула ATP (фактически предшественник, GTP, гуанинтрифосфат). Изучите рисунок выше, чтобы найти каждое из этих событий захвата энергии.
- Обратите внимание, что происходит с атомами углерода (черные точки на рисунке выше). Для каждого 2-углеродного ацетил-КоА, который входит в цикл, высвобождаются две молекулы диоксида углерода, завершая распад исходной 6-углеродной молекулы глюкозы.На последнем этапе регенерируется исходная 4-углеродная молекула, которая начала цикл, так что другой ацетил-КоА может войти в цикл.
Таким образом, цикл лимонной кислоты завершает расщепление глюкозы, которое началось с гликолиза. Его химические реакции окисляют все шесть исходных атомов углерода до \ (\ ce {CO_2} \) и улавливают энергию, выделяемую в 2 АТФ, 6 НАДН и 2 ФАДН \ (_ 2 \). Эти энергоносители присоединяются к 2 АТФ и 2 НАДН, образующимся при гликолизе, и 2 НАДН, образующимся при превращении 2 пирувата в 2 молекулы ацетил-КоА.
В конце цикла лимонной кислоты глюкоза полностью расщепляется, но было произведено только четыре АТФ. Более того, хотя кислород необходим для запуска цикла лимонной кислоты, химические реакции цикла сами по себе не потребляют \ (\ ce {O_2} \). Завершение клеточного дыхания, стадия 3, производит большую часть АТФ.
Дополнительные ресурсы
- Цикл лимонной кислоты (также известный как цикл Кребса): virtuallabs.stanford.edu/other/biochem/TCA.swf
- Цикл Кребса (он же цикл лимонной кислоты): http: // johnkyrk.ru / krebs.html
Авторы и ссылки
Фонд CK-12 Шэрон Бьюик, Ричард Парсонс, Тереза Форсайт, Шонна Робинсон и Жан Дюпон.
Эллисон Султ, Ph.D. (Кафедра химии, Университет Кентукки)
Triose — обзор | Темы ScienceDirect
Фруктозные диеты
Хотя генетические факторы, такие как «экономные гены», могут играть важную роль в развитии метаболического синдрома, скорость нынешней эпидемии метаболического синдрома в Соединенных Штатах и во всем мире предполагает значительный вклад окружающей среды. факторы.Таким образом, взаимодействие между генами и недавние изменения в окружающей среде могли «разоблачить» патогенный потенциал этих генов. Например, недавняя экологическая оценка диабета 2 типа и изменений в структуре питания в Соединенных Штатах за последнее столетие выявила поразительную связь между повышенным потреблением рафинированных углеводов в форме кукурузного сиропа с высоким содержанием фруктозы (HFCS) и увеличением распространенности диабета этого типа. 2 сахарный диабет 29 . В период с 1970 года, когда на рынок был введен КСВСФ, по 2000 год потребление КСВСФ на душу населения в Соединенных Штатах увеличилось с 0.С 292 кг / год до 33,4 кг / год, что более чем в 100 раз больше 30 .
Фруктоза оказывает значительное влияние на метаболизм глюкозы и липидов. Метаболизм фруктозы в печени включает фосфорилирование ферментом фруктокиназой с последующим расщеплением образующегося продукта, фруктозо-1-фосфата, на глицеральдегид и дигидроксиацетонфосфат ферментом альдолазой B. Таким образом, молекулы фруктозы обходят основной этап, регулирующий скорость гликолиза. , 6-фосфофруктокиназа и метаболизируются непосредственно в триозофосфаты, которые могут подпитывать липогенез de novo .Потребление фруктозы в небольших (каталитических) количествах способствует усвоению глюкозы печенью — эффекту, опосредованному активацией глюкокиназы, вызванной фруктозой. Однако при приеме в больших количествах фруктоза продолжает вступать в гликолитический путь, приводя к избыточному липогенезу de novo и гипертриглицеридемии 31 . Кроме того, в отличие от глюкозы, фруктоза не стимулирует выработку инсулина или лептина и не подавляет грелин 32 . Поскольку эти гормоны участвуют в долгосрочном энергетическом гомеостазе, хроническое потребление продуктов с высоким содержанием фруктозы может привести к гипертриглицеридемии, ожирению и метаболическому синдрому.
Хотя в краткосрочной перспективе фруктоза не стимулирует секрецию инсулина, в долгосрочной перспективе увеличение массы тела и увеличение циркулирующих неэтерифицированных жирных кислот (СЖК) приводит к накоплению внутримиоцитов липидов и «липотоксичности» (см. Ниже) которые вызывают инсулинорезистентность и компенсаторную гиперинсулинемию. Следует также отметить, что, хотя основные механизмы не совсем понятны, диеты с высоким содержанием фруктозы также вызывают гипертензию у экспериментальных животных 33-36 .
Наконец, интересно, что генетический фон является важной детерминантой метаболических нарушений, вызванных диетами с высоким содержанием фруктозы. Например, дефицит мембранного белка FAT / CD36, продукта гена, участвующего в клеточном поглощении жирных кислот, защищает от резистентности к инсулину, вызванной диетой с высоким содержанием жиров, но предрасполагает к резистентности к инсулину, индуцированной диетой с высоким содержанием фруктозы у мышей 37 . Это в сочетании с относительно высокой частотой дефицита CD36 у людей от 0.От 3% до 18,5% в зависимости от субпопуляции 38 , 39 , подчеркивает важность генетических и экологических (диетических) взаимодействий как движущей силы в развитии метаболического синдрома. Аналогичным образом, недавно было показано, что кормление фруктозой определенных линий инбредных мышей (например, CBA / JN и C3H / He) индуцировало фенотип, характерный для MetS (например, постпрандиальную гипертриглицеридемию, гиперинсулинемию и накопление висцерального жира). Это было связано с большой индукцией ключевого липогенного фактора транскрипции, Sterol Regulatory Element-binding Protein –1c (SREBP1c), в печени этих животных.Напротив, другие штаммы (например, BDA / 2N и DBA / 1JN) не показали таких метаболических нарушений и индукции печеночного SREBP1c 40 . Другим интересным примером взаимодействий «среда-ген» является фенотип мышей с селективной делецией в жировой ткани ацил-КоА: диаглицерин-ацилтрансферазы (DGAT1). DGAT1 является ключевым ферментом в биосинтезе TG и хранении энергии у млекопитающих. Тем не менее, мыши, которым пересажена белая жировая ткань, лишенная гена DGAT1, устойчивы к ожирению, вызванному диетой, и непереносимости глюкозы 41 .Аналогичным образом устранение гена, кодирующего SCD-1, фермента, участвующего в дельта-9-цис-десатурации насыщенных жирных кислот для использования в биосинтезе триглицеридов и эфиров холестерина, приводит к повышению чувствительности к инсулину, снижению ожирения, устойчивости к диете. -индуцированное ожирение и индукция нескольких генов окисления липидов у мышей 42 . Еще одним недавним примером является роль киназы 1 mTOR / S6 в индуцированной диетой с высоким содержанием жиров инсулинорезистентности и ожирении. В частности, удаление гена S6 киназы 1 у мышей защищает их от ожирения, вызванного диетой с высоким содержанием жиров, и инсулинорезистентности из-за повышенного расхода энергии и функциональности митохондрий 43 .
Эти примеры согласуются с основной концепцией о том, что аллостатические системы, выбранные для поддержки гомеостаза и, в конечном итоге, для обеспечения организмов преимуществом выживания, возможно, стали «устаревшими» и что их присутствие может предрасполагать к аллостатической нагрузке в текущей среде избыточной энергии. избыток.
Например, как упоминалось выше, дефицит CD36 защищает от резистентности к инсулину, вызванной диетой с высоким содержанием жиров, функциональность SREBP1c защищает от метаболических нарушений, вызванных простыми углеводами, DGAT1 и SCD-1 защищают от дефицита накопления энергии, а система киназы 1 mTOR / S6 поддерживает гомеостатический регулирование распределения питательных веществ по путям выработки и хранения энергии.Однако эти же системы могут привести к аллостатической нагрузке и болезням. Таким образом, дефицит CD36 в контексте диеты с высоким содержанием простых углеводов может привести к инсулинорезистентности, хроническая активация SREBP1c может привести к избыточному липогенезу и вторичной гиперинсулинемии и инсулинорезистентности, активность DGAT1 и SCD-1 в среде изобилия энергии может привести к ожирение и инсулинорезистентность, а также конститутивная нутритивная активация киназной системы mTOR / S6 может привести к устойчивой инсулинорезистентности и ожирению.
Таким образом, может показаться, что хроническое потребление фруктозы и, в целом, чрезмерное потребление калорий на фоне набора «устаревших» генов, предназначенных для поддержки гомеостаза и повышения выживаемости, может привести к метаболическим конфликтам, которые вызывают метаболическую аллостатическую нагрузку и развитие МетС.
Сбалансированное химическое уравнение для клеточного дыхания: значение и функция
Клеточное дыхание — это процесс, с помощью которого клетки преобразуют питательные вещества в энергию, которая используется для обеспечения различных функций, таких как транспортировка, перемещение и синтез макромолекул.
Работа клеточного дыхания заключается в образовании аденозинтрифосфата, молекулы, используемой для получения энергии. Как происходит переход между питательными веществами и аденозинтрифосфатом или АТФ? Каковы этапы клеточного дыхания?
Когда вы просыпаетесь утром, подумайте о том, какая драгоценная привилегия — быть живым — дышать, думать, наслаждаться, любить. — Marcus Aurelius
Сбалансированное химическое уравнение (формула) для клеточного дыхания
Уравновешенное уравнение (формула), представляющее клеточное дыхание:
C 6 H 12 O 6 + 6O 2 → 6CO 2 + 6H 2 O + 38 ATP, или:
Эту формулу также можно читать как:
Глюкоза + кислород -> вода + углекислый газ + энергия
По сути, это означает что в клеточном дыхании глюкоза и кислород превращаются в воду, углекислый газ и энергию.Ваше тело использует кислород, которым вы вдыхаете, а также пищу, которую вы едите, для производства энергии.
| 4 стадии клеточного дыхания | |||||||||
| Гликолиз (распад глюкозы) | |||||||||
| Ссылочная реакция | |||||||||
| Цикл Кребса | |||||||||
| Цикл Кребса | |||||||||
| Цепь транспорта электронов | Кислород, которым вы вдыхаете, расщепляет сахара из пищи и производит тепловую энергию, как при сжигании древесины для высвобождения энергии.При клеточном дыхании кислород используется для расщепления сахара, высвобождается энергия сахара и в качестве побочного продукта образуется углекислый газ. Высвободившаяся энергия сохраняется в ваших клетках для дальнейшего использования.
Некоторое количество АТФ, используемого клетками, происходит непосредственно в результате реакций, вызвавших трансформацию глюкозы. Однако большое количество АТФ вырабатывается позже, во время фазы клеточного дыхания, называемой окислительным фосфорилированием.Процесс аэробного дыхания (аэробное означает, что он использует кислород) разделен на четыре отдельных этапа. Окислительное фосфорилирование можно рассматривать как заключительный этап процесса клеточного дыхания.
Стадии клеточного дыханияЧетырьмя стадиями клеточного дыхания являются окисление гликолиза и пируват. , цикл лимонной кислоты и окислительное фосфорилирование. Гликолиз — это этап, на котором глюкоза превращается в другие молекулы в результате ряда химических превращений. Он находится в цитозоле клеток и может функционировать как с кислородом, так и без него. Глюкоза — это сахар с шестью атомами углерода, и при аэробном дыхании глюкоза превращается в две молекулы пирувата. Когда молекулы пирувата окисляются, они производят два НАДН, которые помогают переносить электроны в другие реакции, а также две молекулы АТФ. Окисление пирувата происходит, когда пируват попадает в митохондриальный матрикс, который является самой внутренней частью митохондрий (структура клеток, производящая энергию).В митохондриальном матриксе пируват связывается с коферментом А и превращается в двухуглеродную молекулу. Эта новая структура известна как «ацетил-КоА». Этот процесс генерирует НАДН и высвобождает углекислый газ. Цикл лимонной кислоты (также известный как цикл Кребса или цикл трикарбоновой кислоты) — это цикл, в котором ацетил-КоА, полученный на последней стадии, объединяется с молекулой щавелевоуксусной кислоты. При этом образуется молекула лимонной кислоты, которая претерпевает сложный цикл реакций.На последнем этапе этого цикла создается молекула щавелевоуксусной кислоты, поэтому цикл может начаться снова. Во время цикла выделяется углекислый газ и производятся НАДН, ФАДх3 и АТФ. Электроны в NADH и FADh3 отправляются в цепь переноса электронов. Во время стадии окислительного фосфорилирования молекулы NADH и FADh3, которые были созданы во время других стадий, сбрасывают свои электроны в цепи переноса электронов. Это означает, что они больше не загружены электронами, сводя их к более простым формам, NAD + и FAD.Затем электроны перемещаются по транспортной цепи, которая высвобождает энергию. Этот процесс выталкивает протоны из митохондриального матрикса, который образует градиент. Протоны возвращаются в матрицу через фермент, называемый АТФ-синтазой, который, как это звучит, производит АТФ. Наконец, цепь переноса электронов заканчивается, когда кислород принимает электроны и связывается с протонами, образуя воду. Сколько производится АТФ?Сколько АТФ вырабатывается этим процессом в целом? Большая часть АТФ на самом деле генерируется за счет протонного градиента, который создается во внутреннем матриксе митохондрий.Окислительное фосфорилирование может генерировать около 26-28 единиц АТФ, а фосфорилирование субстрата может генерировать еще около 4-6 единиц. Однако подготовка к гликолизу также потребляет немного АТФ, а это означает, что на самом деле общий выход для процесса, вероятно, составит около 30 единиц АТФ. Анаэробное дыхание и ферментацияРисунок, представляющий клеточное дыхание в органеллах, называемых митохондриями. «Диаграмма дыхания» от Bio4lyfe через Wikimedia Commons, CC-BY-SA 3.0Все вышеперечисленные процессы предполагают наличие достаточного количества кислорода для процесса аэробного дыхания. Если кислород недоступен, что произойдет вместо этого — «анаэробное дыхание», формой которого является ферментация? Аэробное дыхание намного эффективнее анаэробного дыхания, производя примерно в 18 раз больше энергии, чем при ферментации. Напротив, процесс ферментации будет производить только небольшое количество АТФ (2 единицы), а иногда и молочную кислоту. Во время ферментации АТФ извлекается только путем гликолиза.Пируват, который образуется при гликолизе, не проходит остальную часть процесса, пропуская окисление и цикл лимонной кислоты. Он также не проходит через цепь переноса электронов. Из-за того, что цепь переноса электронов не функционирует, НАДН не может сбрасывать туда свои электроны и восстанавливаться до НАД +.
Вместо этого существует несколько дополнительных реакций ферментации, которые создают НАД + из НАДН, который был произведен посредством гликолиза.Это достигается за счет того, что НАДН использует органическую молекулу, такую как пируват, для отбрасывания переносимых ею электронов, а гарантированная подача НАД + означает, что гликолиз может продолжать функционировать. Некоторые клетки могут совершать так называемое «молочнокислое брожение». В этом процессе НАДН переносит любые электроны на молекулы пирувата, а это означает, что лактат образуется как побочный продукт. Виды бактерий, которые создают йогурт, осуществляют этот процесс молочнокислого брожения, но также и клетки вашего тела.Ваши эритроциты и мышечные клетки могут ферментировать молочную кислоту. В случае с вашими эритроцитами необходимо, чтобы они могли ферментировать молочную кислоту, потому что у них нет митохондрий и, следовательно, они не могут выполнять аэробное клеточное дыхание. Ваши мышечные клетки будут производить молочнокислое брожение только при напряжении после того, как они были использованы для очень тяжелых упражнений и кислород, который присутствовал, уже был израсходован. Молочная кислота, вырабатываемая мышечными клетками, затем с кровотоком переносится в печень.В печени он снова превращается в пируват для нормальной обработки посредством клеточного дыхания. Способность клеток дышать и создавать энергию — вот что позволяет им выполнять сложные задачи. Похожие записи |
