
Мышечная энергетика. Часть1
Тренируйтесь для повышения взрывной мощности
АТФ — главный поставщик энергии в организме человека. Он состоит из аденозина и трех фосфатных групп. Когда связь между фосфатной группой и аденозином разрывается, вырабатывается энергия, используемая в вышеуказанных случаях. Проблема в том, что организм человека не в состоянии накапливать АТФ. Он содержится в мышцах в количестве, достаточном лишь для работы длительностью в 1 секунду (5).
Что же происходит, когда вы продолжаете работать? Ведь большинство видов деятельности длятся дольше.
Человеческий организм использует несколько систем энергоснабжения, каждая из которых по-разному производит и использует АТФ. При выполнении какой-либо работы вы тренируете одну или несколько энергетических систем, а когда принимаете пищевые добавки — пытаетесь усилить их. Поэтому понимание принципа их работы позволит вам не только повысить результаты тренировок, но и более грамотно потреблять пищевые добавки.
Главными энергетическими системами являются фосфагенная (система быстрого реагирования), анаэробный гликолиз, аэробный гликолиз и окислительная энергетическая система, иногда называемая электронно-транспортной цепью. Аэробный гликолиз и окислительная система используются организмом при длительных, изнурительных нагрузках, и они неприменимы в бодибилдинге. Мы не будем на них подробно останавливаться. Наша статья — о фосфагенной энергетической системе и анаэробном гликолизе.
Как работает система быстрого реагирования
Когда организму для осуществления какого-либо движения срочно требуется энергия, он расщепляет имеющийся АТФ.
При отщеплении одной фосфатной группы от аденозина выделяется энергия. В результате остается молекула с двумя фосфатными группами или аденозина дифосфат (АДФ).
АТФ => АДФ + энергия
Помните ли вы, что запаса АТФ хватает только на очень короткое время? При длительных энергозатратах «горючего» требуется больше. Тут вам поможет вещество, хорошо знакомое культуристам — это креатина фосфат (КФ).
Он состоит из молекулы креатина и молекулы фосфата. Когда вам нужно больше АТФ, фосфат креатина входит в реакцию с АДФ, в ходе которой восполняет недостающую фосфатную группу аденозину.
АДФ + КФ => АТФ + К
То есть, КФ способствует ресинтезу АТФ.
Понятно, что количество АТФ — это не всецело определяющий фактор. Конечно, это макроэргическое вещество весьма важно для организма. Это одна из тех вещей, которых никогда не бывает слишком много. Но гораздо важнее, сколько КФ вы имеете перед началом тренировки. Чем больше, тем дольше вы можете упражняться и поддерживать интенсивность.
Как используется эта система
По данным исследователей, система быстрого энергоснабжения покрывает энергозатраты в течение 6-10 секунд. Она предназначена для обеспечения короткой, высокоинтенсивной работы — такой, как первые несколько повторений при работе с отягощениями, короткие спринтерские забеги, прыжки и т.д.
Существуют специальные тесты, проверяющие работу именно этой системы. Некоторые из них очень популярны среди тренеров и наставников. Это одно максимальное повторение, вертикальный прыжок, прыжок с места, спринт на 30 м. Одно максимальное повторение показывает, какой вес вы в состоянии поднять один раз. Оно демонстрирует вашу способность мобилизовать необходимые мышечные группы для развития максимального усилия. Другие тесты показывают, как быстро вы можете это делать.
Тот, кто стремится к силе и мощности, должен иметь хорошо тренированную фосфагенную энергетическую систему. Пауэрлифтеры, тяжелоатлеты, метатели, спринтеры, прыгуны и футболисты — все входят в эту категорию. Так как же тренировать эту систему?
Тренировка фосфагенной энергетической системы
Суть тренировки — в увеличении количества креатина фосфата, который необходим для ресинтеза АТФ во время высокоинтенсивных упражнений.
Существует три основных способа: работа с отягощениями, плиометрические упражнения и спринт. В первом случае атлеты должны применять базовые, компаундные упражнения с большими весами и продолжительным отдыхом между сетами.
Для улучшения работы фосфагенной системы не лишним будет и включение в тренировочную программу различных вариаций темповых тяжелоатлетических упражнений.
Примеры силовой и мощностной программ, способных улучшить энергетические системы человека, будут приведены ниже. Используйте их как часть периодизированного мезоцикла или просто на пару месяцев включите в свою тренировочную программу. Количество повторений указано для максимального веса — то есть того, с которым вы сможете выполнить упражнение указанное количество раз. Как вы увидите, программы состоят из базовых, компаундных упражнений с добавлением некоторых тяжелоатлетических движений. Рабочие веса очень большие и требуют продолжительного отдыха между подходами.
Фосфагенную систему можно тренировать плиометрическими упражнениями и спринтами. Забеги, длящиеся менее 10 секунд, повысят способности нижней части тела к хранению и использованию креатина фосфата. А плиометрию можно применять и к нижней, и к верхней части тела, и даже к прессу.
Пищевые добавки и фосфагенная энергетическая система
Отрегулировав тренировочную программу, можно приступить к изменениям в питании. Лучшими добавками для достижения ваших целей будут креатин и рибоза. Креатин эффективен в любой форме — как моногидрат, так и фосфат (3). Креатиносодержащие добавки повысят уровень фосфата креатина в мышцах, что позволит поддерживать высокий уровень АТФ в течение тренировки, а, следовательно, вы сможете заниматься интенсивней и дольше (6). Из-за своей способности влиять на фосфагенную энергетическую систему креатин увеличивает силу, мощность и скорость (4). Рибоза работает в паре с ним и еще больше повышает уровень фосфата креатина.
Как лучше их применять? Ко мне часто обращаются атлеты с жалобой на то, что после приема креатина в течение 2-3 лет они не почувствовали результатов. Вот мои рекомендации.
1. Загрузочная фаза. Принимайте по 20 г креатина в день на протяжении пяти дней, деля их на четыре дозы по 5 г (2,4).
2. Поддерживающая фаза. Снизьте дозу до 2-3 г в день и принимайте ее только в дни тренировок.
3. Если вы пользуетесь порошковым креатином, принимайте его вместе с пищей, а если растворимым — то через 1,5 часа после еды.
4. Снизьте до минимума потребление кофеина, так как он препятствует абсорбции креатина (4).
5. Когда вы входите в фазу суперинтенсивного тренинга (креатин в этом случае вам особенно необходим), возвращайтесь к загрузочной дозе.
6. Помните, что прием креатина необходимо циклировать.
8. Если вы хотите максимизировать энергетику, принимайте 2,2 г рибозы до и после тренировки.
Возникает вопрос, почему бы, начав с загрузочной фазы, так и не остаться на этом уровне приема? Будет ли это причиной всех тех побочных эффектов, о которых ходят слухи? Есть старое правило: больше — не всегда лучше. Очевидно, что мышцы способны сохранить определенную дозу креатина. Затем они насыщаются и теряют способность принимать такое количество (2). Большинство исследований показывают, что после пяти дней загрузки мышцы полностью насыщаются креатином. Если после этого продолжать загрузку, организм будет вынужден куда-то девать излишки — следовательно, все они пойдут через печень и почки. Таким образом, если вы принимаете креатин в очень больших количествах и длительное время, вы перегружаете печень и почки, что может стать причиной их заболевания. Поэтому так важно циклировать прием креатиносодержащих добавок.
Во второй части этой статьи, которая будет опубликована в следующем номере журнала, мы поговорим о способах улучшения анаэробного гликолиза.
Ссылки:
1. Bessman, S.P., and Carpenter, C.L. (1985). The creatine-creatine phosphate energy shuttle. Annual Reviews of Biochemistry. 54:831-862.
2. Greenhaff, P.L. (1995). Creatine and its application as an ergogenic aid. International Journal of Sports Nutrition. 5(supplement):S100-110.
3. Peters, B.M.; Lantz, C.D.; and Mayhew, J.L. (1999). Effect of oral creatine monohydrate and creatine phosphate supplementation on maximal strength indices, body composition and blood pressure. Journal of Strength and Conditioning Research. 13(1):3-9.
4. Plisk, S.S., and Kreider, R.B. (1999). Creatine controversy? Strength and Conditioning Journal. 21(1):14-23.
5. Stryer, L. Biochemistry, 3rd edition. New York: W.H. Freeman and Company. 1988. 315-320, 935-936.
ironman.ru
Фосфагенная энергетическая система — Физкультура на 5
Восстановление АТФ осуществляется во время мышечного сокращения почти мгновенно, как только он расщепляется до АДФ. Оно происходит за счет энергии другого высокоэнергетического фосфатного соединения – креатинфосфата (КрФ). При отщеплении от КрФ фосфатной группы освобождается большое количество энергии – в среднем 10,5 ккал при расщеплении 1 м КрФ (М-1моля). Конечными продуктами расщепления КрФ являют креатин и свободный фосфат. Энергия от расщепления КрФ тут же используется для ресинтеза АТФ. Таким образом, КрФ – это первый энергетический резерв мышцы, действующий как немедленный источник для восстановления АТФ. АТФ и КрФ действуют как единая фосфагенная система энергообеспечения. Деятельность этой системы не зависит ни от сложной последовательности химических реакций, ни от доставки О2 к мышцам и, поэтому фосфагенная система представляет собой наиболее быстрый источник энергии, используемый мышечной клеткой с начала мышечных сокращений. Фосфагенная система обладает наибольшей мощностью по сравнению с другими системами. Она примерно в 3 раза превышает максимальную мощность лактацидной (гликолитической) и в 4-10 раз максимальную мощность кислородной системы. Поэтому фосфагенная система играет решающую роль в энергообеспечении работ предельной мощности (спринт, штанга, прыжки и др.), осуществляемых с максимально возможными по силе и скорости (мощности) сокращениями мышц. Емкость фосфагенной системы невелика, поскольку запасы АТФ и КрФ в мышце ограничены. Емкость фосфагенной системы, оцениваемая запасом АТФ в 20 кг мышц и количеством АТФ, которое может ресинтезироваться за счет КрФ, содержащейся в той же мышечной массе, составляет всю около 0,5 моль АТФ, т.е. около 5 ккал. Поэтому такая работа может продолжаться в течение нескольких секунд. Например, при спринтерском беге энергетический запрос может составлять около 1 ккал/сек. Поэтому теоретически такая работа может продолжаться в течение 5 сек за счет фосфагенной системы. Поэтому, если работа выполняется большей продолжительности, то ее энергетическое обеспечение не может происходить только за счет наличных запасов АТФ и КрФ.
fizkultura-na5.ru
АТФ
АТФ — Аденозин Три-Фосфорная кислота
АТФ формулаАТФ (аденозин трифосфат: аденин, связанный с тремя фосфатными группами) — молекула, которая служит источником энергии для всех процессов в организме, в том числе для движения. Сокращение мышечного волокна происходит при одновременном расщеплении молекулы АТФ, в результате чего выделяется энергия, которая идет на осуществление сокращения. В организме АТФ синтезируется из инозина.
АТФ должна пройти через несколько ступеней, чтобы дать нам энергию. Сначала при помощи специального коэнзима отделяется один из трех фосфатов (каждый из которых дает десять калорий), высвобождается энергия и получается аденозин дифосфат (АДФ). Если энергии требуется больше, то отделяется следующий фосфат, формируя аденозин монофосфат (АМФ). Главным источником для производства АТФ служит глюкоза, которая в клетке инициально расщепляется на пируват и цитозол.
Во время отдыха происходит обратная реакция – при помощи АДФ, фосфагена и гликогена фосфатная группа вновь присоединяется к молекуле, формируя АТФ. Для этих целей из запасов гликогена берется глюкоза. Вновь созданный АТФ готов к следующему использованию. В сущности АТФ работает как молекулярная батарея, сохраняя энергию, когда она не нужна, и высвобождая в случае необходимости.
Структура АТФ
Молекула АТФ состоит из трех компонентов:
1. Рибоза (тот же самый пятиуглеродный сахар, что формирует основу ДНК)
2. Аденин (соединенные атомы углерода и азота)
3. Трифосфат
Молекула рибозы располагается в центре молекулы АТФ, край которой служит базой для аденозина. Цепочка из трех фосфатов располагается с другой стороны молекулы рибозы. АТФ насыщает длинные, тонкие волокна, содержащие протеин, называемый миозином, который формирует основу наших мышечных клеток.
Системы АТФ
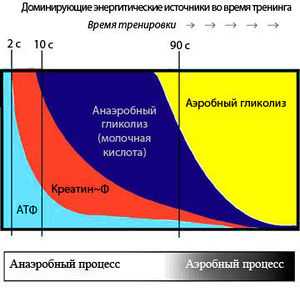 Последовательное включение энергетических систем во время выполнения упражнений
Последовательное включение энергетических систем во время выполнения упражненийЗапасов АТФ достаточно только на первые 2-3 секунды двигательной активности, однако мышцы могут работать только при наличии АТФ. Для этого существуют специальные системы, которые постоянно синтезируют новые молекулы АТФ, они включаются в зависимости от продолжительности нагрузки (см. рисунок). Это три основные биохимические системы:
1. Фосфагенная система (Креатин-фосфат)
2. Система гликогена и молочной кислоты
3. Аэробное дыхание
Фосфагенная система
Когда мышцам предстоит короткая, но интенсивная активность (приблизительно 8-10 секунд), используется фосфагенная система – АДФ соединяется с креатина фосфатом. Фосфагенная система обеспечивает постоянную циркуляцию небольшого количества АТФ в наших мышечных клетках. Мышечные клетки также содержат высокоэнергетический фосфат – фосфат креатина, который используется для восстановления уровня АТФ после кратковременной, высокоинтенсивной работы. Энзим креатин киназа отнимает фосфатную группу у креатина фосфата и быстро передает ее АДФ для формирования АТФ. Итак, мышечная клетка превращает АТФ в АДФ, а фосфаген быстро восстанавливает АДФ до АТФ. Уровень креатина фосфата начинает снижаться уже через 10 секунд высокоинтенсивной активности. Пример использования фосфагенной системы энергоснабжения – это спринт на 100 метров.
Система гликогена и молочной кислоты
Система гликогена и молочной кислоты снабжает организм энергией медленнее, чем фосфагенная система, и предоставляет достаточно АТФ примерно для 90 секунд высокоинтенсивной активности. В ходе процесса из глюкозы мышечных клеток в результате анаэробного метаболизма происходит формирование молочной кислоты.
Учитывая тот факт, что в анаэробном состоянии организм не использует кислород, эта система дает кратковременную энергию без активации кардио-респираторной системы точно так же, как и аэробная система, но с экономией времени. Более того, когда в анаэробном режиме мышцы работают быстро, они очень мощно сокращаются, перекрывая поступление кислорода, так как сосуды оказываются сжатыми. Эту систему еще можно назвать анаэробно-респираторной, и хорошим примером работы организма в этом режиме послужит 400-метровый спринт. Обычно продолжать работать таким образом атлетам не дает мышечная болезненность, возникающая в результате накопления молочной кислоты в тканях.
Аэробное дыхание
Если упражнения длятся более двух минут, в работу включается аэробная система, и мышцы получают АТФ вначале из углеводов, потом из жиров и наконец из аминокислот (протеинов). Протеин используется для получения энергии в основном в условиях голода (диеты в некоторых случаях). При аэробном дыхании производство АТФ проходит наиболее медленно, но энергии получается достаточно, чтобы поддерживать физическую активность на протяжении нескольких часов. Это происходит, потому что глюкоза распадается на диоксид углерода и воду беспрепятственно, не испытывая противодействия со стороны, например, молочной кислоты, как в случае анаэробной работы.
Читайте также
sportguardian.ru
Читать книгу Физиология силы Руслана Городничева : онлайн чтение
Текущая страница: 4 (всего у книги 14 страниц) [доступный отрывок для чтения: 10 страниц]
Глава 3
Биоэнергетика силы
Единственным непосредственным источником энергии для мышечного сокращения служит АТФ. При активации мышцы повышение внутриклеточной концентрации ионов кальция приводит к сокращению и усиленному расщеплению АТФ. Интенсивность метаболизма мышцы возрастает при этом в 100-1000 раз. АТФ гидролитически расщепляется с помощью миозин-АТФ-азы до аденозиндифосфата (АДФ) и неорганического фосфата. Расщепление одного моля АТФ обеспечивает около 8 кДж энергии. Только 40–50 % этой энергии преобразуется в механическую энергию работы. Остальные 50–60 % превращаются в тепло. В естественных условиях лишь 20–30 % всех энерготрат идет на механическую работу, поскольку часть энергии используется для работы ионных насосов и окислительного восстановления АТФ. Чем больше произведенная работа, тем больше расходуются источники энергии и значительнее выделение тепла. Наибольшую работу мышца совершает при средних величинах внешней нагрузки. Эта закономерность названа законом средних нагрузок.
Для поддержания довольно длительной мышечной деятельности необходимо постоянное восстановление АТФ с той же скоростью, с которой он расходуется. Необходимая для ресинтеза АТФ энергия поступает в результате окисления углеводов, жиров, белков, а также за счет расщепления креатинфосфата и гликогена (глюкозы). Распад этих веществ сопровождается освобождением энергии, запасенной в их химических связях. Эта свободная энергия обеспечивает связывание АДФ и фосфата с образованием АТФ.
Ресинтез АТФ осуществляется в мышце двумя основными путями – анаэробным (без участия кислорода) и аэробным (с участием кислорода). Для ресинтеза и использования АТФ в качестве источника энергии в сокращающейся мышце могут действовать три энергетические системы: фосфагенная, гликолитическая и окислительная.
Фосфагенная и гликолитическая системы работают по анаэробному пути, а окислительная – по аэробному. Эти три системы отличаются по энергетической емкости – максимальному количеству образуемой энергии, или максимальному количеству АТФ, которое может ресинтезироваться за счет энергии каждой из этих систем. Неодинакова и их энергетическая мощность – максимальное количество энергии, выделяющейся за единицу времени, или максимальное количество АТФ, образующейся в единицу времени за счет энергии этих систем.
От емкости энергетической системы зависит максимальный объем работы, выполняемый за счет данной системы. Мощность системы ограничивает предельную интенсивность работы, которая может быть выполнена за счет энергии данной системы. Степень участия той или иной системы в ресинтезе АТФ определяется интенсивностью и продолжительностью мышечной деятельности (табл. 3.1).
Таблица 3.1
Взаимосвязь между продолжительностью работы и энергетическими системами
Фосфагенная энергетическая система. К группе фосфагенов относят АТФ и креатинфосфат (КрФ). При расщеплении КрФ освобождается большое количество энергии, причем креатинфос-фат взаимодействует с АДФ, образуя АТФ с выделением креатина. КрФ содержится в мышцах в большем количестве, чем АТФ. КрФ и АТФ действуют как единая система энергопродукции. Фосфагенная система обеспечивает наиболее быстрое восстановление АТФ, поэтому используется с самого начала мышечных сокращений. Она обладает наибольшей мощностью по сравнению с другими системами. В связи с этим фосфагенная система играет решающую роль в энергообеспечении кратковременных максимальных мышечных сокращений. В силу ограниченности запасов АТФ и КрФ в мышце емкость фосфагенной системы сравнительно невелика. Теоретический расчет показывает, что работа с максимальной мощностью, которая обеспечивается исключительно анаэробной фосфагенной системой, может продолжаться не более 5 с. Поэтому при выполнении более длительной работы энергетическое обеспечение осуществляется другими системами.
Систематическая силовая тренировка может приводить к изменению концентрации фосфагенов в скелетных мышцах. Так, под влиянием 5-недельной силовой тренировки происходит заметное увеличение содержания фосфагенов в икроножной мышце (J.D. MacDougall et al., 1977). Такое повышение концентрации фосфагенов связано с избирательной гипертрофией мышечных волокон типа II, которые могут содержать большее количество фосфагенов, чем волокна типа I (J. D. MacDougall, 1986).
Гликолитическая энергетическая система. В основе деятельности этой системы лежит анаэробное расщепление углеводов (гликогена, глюкозы) до молочной кислоты. Совокупность идущих при этом химических реакций называется анаэробным гликогенолизом или гликолизом. При гликолизе освобождается энергия, позволяющая ресинтезировать АТФ. В результате анаэробного гликогенолиза каждая молекула глюкозы дает энергию для образования трех молекул АТФ.
В процессе гликолиза возрастает концентрация молочной кислоты в мышцах и крови, что приводит к сдвигу их активной реакции в кислую сторону. Это затрудняет обменные процессы, снижает активность ключевых гликолитических ферментов. В результате понижается скорость гликолиза и количество энергии (АТФ), образующейся в единицу времени за счет этой системы. Отсюда следует, что емкость гликолитической системы ограничивается не содержанием гликогена и глюкозы, а концентрацией (количеством) конечного продукта деятельности этой системы – молочной кислоты.
Гликолитическая система мышц характеризуется большой мощностью, величина которой в 1,5 раза выше, чем окислительной системы, но примерно в 3 раза ниже мощности фосфагенной системы. Энергетическая емкость гликолитической системы значительно меньше, чем окислительной, но в 2,5 раза больше по сравнению с фосфагенной.
Гликолитическая система функционирует в том случае, когда сокращающиеся мышцы снабжаются кислородом в недостаточной степени. Ей принадлежит решающая роль в энергообеспечении повторных мышечных сокращений большой мощности, продолжительность которых от 20 с до 1–2 мин. Гликолитическая система также обеспечивает ресинтез АТФ в самом начале любой работы, когда потребности мышц в кислороде еще не удовлетворяются. Скорость гликолиза зависит от интенсивности силовых упражнений. Например, скорость гликолиза в течение 6 подходов по 6 повторений в каждом с силой в 70 % ПМ вдвое больше, чем при аналогичном количестве подходов и повторений, но с усилием в 35 % ПМ (C. P. Lambert et al., 1991). Энергетическая роль глико-литичееской системы снижается по мере увеличения продолжительности выполняемой работы. Заметим, что понижение мышечного гликогена и повышение уровня молочной кислоты может быть лимитирующим (ограничивающим) фактором при выполнении силовых тренировочных программ, поддерживаемых в начале работы анаэробными гликолитическими механизмами.
Окислительная энергетическая система. При продолжительной малоинтенсивной мышечной деятельности аэробный ресинтез АТФ происходит главным образом за счет окислительного фосфорилирования. Необходимая для ресинтеза АТФ энергия поступает в результате окисления углеводов или жиров. Преимущественное использование того или иного энергетического субстрата зависит от мощности выполняемой работы. Чем больше мощность работы, тем относительно выше энергетический вклад окисляемых углеводов и соответственно меньше вклад окисляемых жиров в общую энергопродукцию сокращающихся мышц. При работе очень большой мощности окисляются в основном углеводы, а при малоинтенсивной работе – жиры.
При полном аэробном расщеплении одной молекулы глюкозы образуются 38 молекул АТФ. Следовательно, аэробное расщепление глюкозы дает гораздо больше энергии по сравнению с анаэробным гликолизом. В процессе аэробного распада молекулы жирной кислоты образуется в несколько раз больше молекул АТФ, чем при аэробном расщеплении молекулы глюкозы. Таким образом, при окислении одинакового количества углеводов и жиров последние обеспечивают значительно большую энергопродукцию.
Окислительная система, использующая в качестве источников окисления жиры и углеводы, обладает наибольшей энергетической емкостью по сравнению с другими энергетическими системами. Ее емкость в тысячи раз превышает емкость фосфагенной и гликолитической систем. Именно поэтому окислительная энергетическая система обеспечивает возможность выполнения работы в течение длительного времени – от 3–5 мин до нескольких часов.
Биоэнергетические факторы, ограничивающие физическую работоспособность. Достижение максимальной физической работоспособности ограничивается многими причинами, в том числе и биоэнергетическими факторами в результате развивающегося под влиянием работы утомления. В табл. 3.2 приведены некоторые лимитирующие факторы, основанные на снижении энергетических ресурсов и увеличении в скелетных мышцах содержания ионов водорода – [H+].
Как следует из табл. 3.2, значение того или иного фактора в ограничении физической работоспособности зависит от длительности и интенсивности выполняемой работы. Так, при марафонском беге основным лимитирующим фактором работоспособности является снижение гликогена в мышцах, а при силовой тренировке – снижение АТФ, креатинфосфата и увеличение в мышечной ткани концентрации ионов водорода.
Таблица 3.2
Биоэнергетические лимитирующие факторы
Примечание: 5 – наибольшее значение фактора в ограничении работоспособности.
Восстановление после силовых тренировочных нагрузок. Известно, что строго регламентированные периоды отдыха между подходами и отдельными повторениями в конкретных подходах имеют большое значение, поскольку во многом определяют суммарную рабочую нагрузку (В.Н. Платонов, 2004; Г.П. Виноградов, 2009). Целенаправленная регламентация периодов отдыха может обеспечить необходимое увеличение суммарного кислородного долга и повышение общего расхода калорий. Силовые тренировки с разным объемом нагрузки приводят к нарушению постоянства внутренней среды организма и продуцируют кислородный долг от умеренных до больших величин (M.A. Burleson et al., 1998). Более того, тренировка со свободными весами может приводить к большему кислородному долгу, чем типичные аэробные упражнения. Такую закономерность необходимо использовать при проведении занятий по укреплению здоровья, а также в планировании и организации тренировочного процесса в профессиональном спорте.
Следует учитывать, что недостаточные для восстановления между тренировочными занятиями периоды отдыха могут изменять физиологические эффекты проведенной тренировки и уменьшать способность к выполнению последующей двигательной нагрузки. Понижение последующей двигательной активности способно уменьшить ожидаемые тренировочные адаптации к проведенной тренировке. Кроме того, неполное восстановление может быть одним из факторов, которые вызывают перетренированность у спортсменов (M.H. Stone et al., 1991). Несмотря на наличие убедительных доказательств о влиянии предшествующих тренировок на последующие, точные методические указания и конкретные временны е границы для восстановления после силовых тренировочных занятий разной направленности до сих пор неизвестны. Поэтому тренерам и спортсменам следует планировать длительность отдыха, достаточную для адекватного восстановления после силовых тренировок, направленных на решение тех или иных тренировочных задач.
Метаболическая специфичность тренировок. Реализация силовых тренировочных программ, предусматривающих соответствующую интенсивность и продолжительность работы, регламентированные интервалы отдыха, может обеспечить целенаправленную активацию определенных энергетических систем в процессе тренировки в результате специфических адаптаций при выполнении различных силовых упражнений (G.A. Brooks et al., 1996). Несмотря на то что силовая тренировка и другие формы анаэробной мышечной работы могут увеличивать аэробную мощность в незначительной степени (4–9 %) и на короткое время (J. P. McCarthy et al., 1995), главные эффекты таких тренировок связаны с изменениями анаэробных процессов. Силовая, спринтерская и другие формы анаэробной тренировки способны потенциально увеличить запасы фосфагенов и гликогена и повысить миокиназные реакции (G.A. Brooks et al., 1971; Н.И. Волков, В.И. Олейников, 2011). Тренировка такой направленности приводит к преимущественной гипертрофии волокон типа II (P.D. Gollnick, B. Saltin, 1982) и, в основном, повышает анаэробный метаболизм (P. J. Abernethy et al., 1990), обеспечивая увеличение двигательных возможностей спортсменов.
Окислительный метаболизм имеет важное значение в восстановлении организма после тяжелых анаэробных упражнений – силовой тренировки (G.A. Brooks et al., 1996). В то же время нужно проявлять осторожность в использовании средств аэробной тренировки в анаэробных видах спорта. Это связано с наличием экспериментальных доказательств, что аэробная тренировка даже с низкими объемами может ухудшать потенциальные анаэробные возможности при выполнении силовой и скоростной двигательной деятельности (K. Hakkinen et al., 2003). Механизм такого влияния заключается в понижении способности к производству анаэробной энергии при выполнении аэробных мышечных нагрузок (V.A. Vihko et al., 1978). В ряде исследований показано, что комбинированная анаэробно-аэробная тренировка может уменьшить прирост обхвата мышц, максимальную силу и, в особенности, скоростно-силовые способности (G.A. Dudley и R. Diamil, 1985). Хотя биохимические и физиологические механизмы этого процесса до сих пор неясны (M.H Stone et al., 1991), понижение прироста названных выше показателей после комбинированной тренировки может быть в определенной степени связано с изменением свойств мышечных волокон и концентрации тестостерона и кортизола в крови (WJ. Kraemer et al., 1995). Результаты некоторых исследований свидетельствуют, что анаэробные тренировки, включая силовые, способны повышать выносливость при выполнении силовых упражнений низкой интенсивности (JJ. Bastiaans et al., 2001).
Имеются сведения о том, что аэробная тренировка, сопровождающая занятия анаэробной направленности, ускоряет восстановление спортсменов (S.S. Plisk, 1991), поскольку восстановительные процессы обеспечиваются главным образом аэробными механизмами. Количественные характеристики влияния повышения аэробной мощности на процессы восстановления не совсем ясны. В некоторых работах показана зависимость между величиной максимальной аэробной производительности и показателями, характеризующими процесс восстановления после выполнения упражнений: устранение лактата, потребление O2 (J.R. Hoffman, 1997). В других работах такая закономерность не наблюдалась (G.J. Bell et al., 1997). J.R. Hoffman (1997) показал, что способность к восстановлению повышается тренировками аэробной направленности только до определенной степени, превышение которой не дает дополнительной выгоды. Следовательно, имеется оптимальный лимит аэробной производительности и превышение его не влияет на восстановление названных выше параметров. Лимитирующая величина максимальной аэробной производительности, по данным J.R. Hoffman, составляет для мужчин приблизительно 45 мл/кг-1/мин-1.
В заключение отметим, что специфические анаэробные программы могут стимулировать увеличение аэробной мощности и повышать маркеры восстановления (J.L. McMillan et al., 1993). Кросс-поперечные и лонгитудинальные исследования (K.R. Pierce et al., 1987) показали, что силовая тренировка, а в особенности тренировка с высоким объемом нагрузки, может улучшать параметры восстановления, включая более быстрое возвращение к дорабочему уровню: сердечной деятельности (повседневного ЧСС), лактата, аммония и различных гормонов. Таким образом, интенсивная аэробная тренировка не является обязательной для восстановления после анаэробных нагрузок и может быть контрпродуктивной для большинства силовых и скоростно-силовых видов спорта (L.P. Koziris et al., 1996).
Глава 4
Эндокринология силы
В обеспечении двигательной деятельности человека важное значение имеют гормоны – вещества высокой биологической активности, выделяемые железами внутренней секреции (эндокринными железами). Гормоны поступают в кровь, распространяются по кровеносным сосудам со скоростью кровотока и воздействуют на органы и ткани, расположенные вдали от той железы, в которой они продуцируются. Эффект гормона характеризуется специфичностью. Каждый гормон воздействует только на те структуры, в клетках которых располагаются особые рецепторы, связывающие данный гормон. Взаимодействие определенного гормона с его рецептором приводит к строго определенным и последовательным изменениям в цепи обменных процессов, уровне активности катализирующих их ферментов (А.А. Виру, 1981). Гормоны относительно быстро разрушаются, поэтому для поддержания оптимального количества каждого из них в крови необходимо постоянное продуцирование его соответствующей железой внутренней секреции. Заметим, что мышечные усилия, обеспечивающие реализацию того или иного двигательного действия, всегда осуществляются на определенном гормональном фоне. Гормональный статус крови, отражающийся в определенной концентрации каждого из имеющихся в ней гормонов, оказывает влияние на параметры выполняемого движения.
Известно, что различные гормональные системы совместно с нервными влияниями обеспечивают адаптацию организма спортсменов к силовым мышечным нагрузкам (А.А. Виру, 1981; M.H. Stone et al., 2007). На рис. 4.1 представлены основные компоненты адаптации к мышечным нагрузкам. В соответствии с этой схемой приспособление к силовым тренировочным программам осуществляется через механизмы мобилизации энергетических ресурсов, пластического резерва организма и адаптивного синтеза ферментных и структурных белков, защитных способностей организма (А.А. Виру, П.К. Кырге, 1983). Белковый синтез является обязательным компонентом как общего, так и специфического приспособления к мышечной работе. В регуляции белкового синтеза существенное значение имеет деятельность симпатоадреналовой системы. С учетом тематики данной книги особого внимания заслуживает рассмотрение гормонов, оказывающих наибольшее влияние на развитие и проявление силовых возможностей у человека. К таким гормонам прежде всего можно отнести тестостерон и соматотропин.
Рис. 4.1. Схема адаптационных процессов в организме человека (А.А. Виру, 1981).
Тестостерон. Тестостерон стимулирует развитие мужских половых органов и формирование вторичных половых признаков (характер оволосения, тембр голоса, распределение жира на теле), а также обладает анаболическими характеристиками, влияющими почти на каждую ткань и функциональные системы, включая центральную и периферическую нервные системы (А.Р. Arnold, 1984). Тестостерон усиливает синтез белков и, следовательно, способствует развитию гипертрофии скелетных мышц. Тестостерон и его производные имеют отношение к различным поведенческим реакциям человека, включая агрессию (М.Н. Stone, 1993). Механизм влияния этого гормона реализуется посредством связывания или вызова изменений в глюкокортикоидных цитоплазматических рецепторах, торможения катаболических эффектов кортизола и увеличения анаболических эффектов (М. Meyer and Р. Rosen, 1975). Тестостерон, вероятно, обеспечивает более высокий гликолитический профиль мышечных волокон типа II в сравнении с мышечными волокнами типа I и усиливает синтез гликогена посредством стимуляции продуцирования синтетиков гликогена (S. Adalphsson, 1973).
Содержание тестостерона находится в тесной связи с мышечным поперечником, величиной развиваемой силы и мощности (С. Bosco et al., 1996). Уменьшение тестостерона с возрастом может сопровождаться уменьшением двигательных способностей нервно-мышечной системы, в особенности у женщин (К. Hakkinen and А. Pakarinen, 1993).
Легкая аэробная тренировка (< 60 % МПК) оказывает малый эффект на концентрацию тестостерона в сыворотке (J.E. Wilker-son et al., 1980), хотя продолжительная аэробная тренировка может вызвать его уменьшение (К. Dessypris et al., 1976) или увеличение (H. Galbo et al., 1977). Как аэробная (D. Jezova et al., 1985), так и анаэробная тренировка (J.H. Jensen et al., 1991) обычно вызывают увеличение тестостерона, которое зависит от их интенсивности и объема, а также от объема активированной мышечной массы. Основные механизмы увеличения тестостерона, выработанного в процессе тренировки, не вполне ясны, но, возможно, могут быть результатом катехоламиновой стимуляции рецепторов B2 семенников (D. Jezova et al., 1985) или уменьшения выделительных процессов из-за понижения внутреннего кровяного потока (R. Teriung, 1979). При увеличении длительности тренировки на выносливость наблюдается тенденция к понижению концентрации тестостерона до уровня, характерного для состояния мышечного покоя (W. A. Kindermann et al., 1982).
Исследования концентрации тестостерона в сыворотке или плазме крови силовых тренировочных занятий показали противоречивые результаты. Длительная тренировка на выносливость вызывала уменьшение тестостерона в сыворотке крови животных (L. Dohm and T. M. Louis, 1978). У человека аэробная тренировка не приводила к изменениям концентрации тестостерона (N. Fell-mann et al., 1985), а также могла понизить (R. Frey et al., 1983) или увеличить (RJ. Young and A.H. Ismaril, 1978) ее в сравнении с состоянием мышечного покоя. Силовая тренировка также вызывала различные эффекты. Не наблюдалось изменений содержания тестостерона у умеренно тренирующихся молодых мужчин (K.J. Ostrowski et al., 1997) и у мужчин среднего возраста (B.J. Nik-las et al., 1995). Концентрация тестостерона возрастала после силовых тренировок в сравнении с покоем у мальчиков (C. D. Tsolakis et al., 2000), юношей (R.S. Staron et al., 1994) и молодых женщин (J.D. Marx et al., 2001).
Показано, что кратковременные силовые тренировочные программы (1–4 недели) с очень большим объемом (веса) могут уменьшать концентрацию тестостерона у хорошо тренированных штангистов (T.K. Busso et al., 1992). Продолжительные тренировки со штангой мало влияли на содержание тестостерона в состоянии покоя (K.A. Hakkinen et al., 1987). Тем не менее, понижение объема тренировки у штангистов после предшествующих больших нагрузок приводит к увеличению концентрации тестостерона в покое (T.K. Busso et al., 1992). У молодых штангистов (14–20 лет) как кратковременная, так и длительная тренировка со свободными весами может приводить к повышению концентрации тестостерона в покое, а также к увеличению тестостеронового ответа на последующую тренировку (A.C. Fry et al., 1994).
Как отмечают A.C. Fry и W.J. Kraemer (1997), изменения в концентрации гормонов в течение длительного периода тренировок могут быть трудно уловимыми. Содержание тотального тестостерона в состоянии покоя статистически значимо не изменяется. Достоверно повышается лишь концентрация свободного тестостерона, его соотношение с кортизолом и соотношение тотального тестостерона с кортизолом (K. A. Hakkinen et al., 1988). В некоторых работах описано увеличение общего содержания тестостерона, которое реорганизует и изменяет рецепторную активность (M.A. Alen et al., 1988).
Особенности адаптации к разным силовым тренировочным программам, отражающиеся в динамике содержания тестостерона, могут быть результатом различий в уровне тренированности, возрасте, общем состоянии здоровья и физических кондиций обследуемых (W. J. Kraemer et al., 1992). Такие особенности могут быть связаны с видом и интенсивностью тренировки или размером вовлекаемой мышечной массы (D. Blessing et al., 1986). Перечисленные выше факты показывают наличие взаимосвязи между большими или внезапными увеличениями объема тренировки и концентрацией тестостерона и других связанных с этим гормоном параметров в состоянии мышечного покоя.
Возможность прироста мышечной массы в результате тренировки может также определяться соотношением концентрации тестостерона со связанными с ним другими гормонами, например такими, как кортизол (Т: К) (K. Hakkinen et al., 1989).
Соотношение Т: К характеризует доминирование анаболического или катаболического статуса (H. Aldercruetz et al., 1986). Такой показатель может частично оценить и объяснить баланс между андрогено-анаболической активностью (тестостерон) и катаболической активностью (кортизол) в течение длительного периода силовой тренировки (M. Alen and K. Hakkinen., 1987).
Показано, что силовая тренировка с высоким объемом нагрузки вызывает понижение тестостерона и соотношения Т: К с одновременным увеличением лютенизирующего гормона (K. Hakkinen et al., 1985). В течение периода «нормализации» или понижения тренировочных нагрузок концентрация кортизола и лютенизирующего гормона уменьшается.
Продолжительная по времени силовая тренировка может приводить к увеличению и уменьшению показателя Т: К (A.C. Fry and WJ. Kraemer, 1997). Тем не менее даже трудно уловимые изменения в Т: К, сопровождающие тренировочные программы, тесно коррелируют с силовыми возможностями (V. Alen et al., 1988). Объем и интенсивность силовых тренировочных программ проявляется в количественных изменениях этих отношений. На рис. 4.2 представлены данные, показывающие изменение соотношения Т: К у американских пауэрлифтеров национального уровня за 12-недельный период. Из рисунка видно, что изменения Т: К имеют тенденцию быть обратно взаимосвязанными с изменениями в тренировочном объеме нагрузки.
Есть основания утверждать, что это соотношение (Т: К) способно быть чувствительным индикатором тренировочного стресса и общего стресса. Следовательно, изменения Т: К у спортсмена могут быть критериальным параметром для выявления перенапряжения или перетренированности (M. Alen and K. Hakkinen., 1987). Кроме того, соотношение Т: К может использоваться в качестве индикатора «готовности» к проявлению серьезного уровня физической активности. Например, если оно высокое, спортсмен обладает большим потенциалом для хорошего выступления.
Понижение тренировочных нагрузок может иногда увеличивать соотношение T: C выше основного уровня (суперкомпенсационный эффект) и следовательно потенциально повышать «готовность» к достижению высокого спортивного результата.
Рис. 4.2. Изменение соотношения Т: К у американских пауэрлифтеров национального уровня за 12-недельный период тренировки (M.H. Stone et al., 2007).
Соматотропин (гормон роста) вырабатывается в передней доле гипофиза. Он ускоряет процессы роста у детей. При избыточном вырабатывании этого гормона в детском возрасте развивается гигантизм: длина тела в таком случае может достигать иногда даже 230–250 см. У детей количество соматотропина существенно больше, чем у взрослых. С возрастом секреция гормона роста уменьшается. Соматотропин повышает скорость синтеза белка, способствует транспорту аминокислот в клетку, усиливает освобождение жирных кислот из жировой ткани. В определенных условиях гормон роста способствует уменьшению жировых запасов и угнетает использование углеводов тканями.
Содержание гормона роста в крови возрастает после выполнения однократной аэробной и анаэробной мышечной нагрузки. Увеличение концентрации гормона зависит от длительности и интенсивности выполняемой однократной нагрузки (M. H. Stone et al., 2007). Многократные тренировки в течение одного дня не приводят к дальнейшему повышению концентрации соматропина. Повышенный уровень гормона, вызванный одноразовой тренировкой, может сохраняться в течение 30 мин и более после ее окончания. У хорошо тренированных спортсменов наблюдается менее значительное увеличение концентрации соматотропина на стандартную нагрузку (поднимание свободных весов) и более быстрое восстановление в сравнении со спортсменами низкой квалификации.
Значение соматотропина в повышении синтеза белка привело к применению спортсменами его синтетического аналога для повышения физической работоспособности, в том числе и для улучшения силовых возможностей. Однако использование такого препарата в качестве допинга оправданно запрещено, поскольку его длительное применение нарушает гормональный статус организма.
iknigi.net
Приложение 709261417. Энергетика мышечного сокращения — Мегаобучалка
Единственным прямым (непосредственным) источником энергии для мышечного сокращения служит аденозинтрифосфат (АТФ). При гидролизе АТФ образуется аденозиндифосфат (АДФ), отщепляется фосфатная группа и выделяется около 10 ккал свободной энергии[Мф39] .
Ресинтез АТФ
Для того чтобы мышечные волокна могли поддерживать сколько-нибудь длительное сокращение, необходимо постоянное восстановление АТФ с такой же скоростью, с какой он расщепляется (расходуется).
Энергия для восстановления АТФ образуется за счет питательных («горючих») веществ — углеводов, жиров, белков. При распаде (полном или частичном) этих веществ часть энергии, запасенная в их химических связях, освобождается. Эта свободная энергия обеспечивает связывание АДФ и фосфата с образованием АТФ.
Ресинтез АТФ в мышце может происходить двумя основными путями — анаэробным (без участия кислорода) и аэробным (при участии кислорода). Если энергетическое обеспечение работы осуществляется в основном анаэробным путем, такая работа называется анаэробной, а если преимущественно аэробным путем — аэробной работой.
Для образования и использования АТФ в качестве непосредственного источника энергии в сокращающейся мышце могут действовать три химические (энергетические) системы:
1) фосфагенная (АТФ-КФ-система)
2) гликолитическая (лактацидная)
3) окислительная (кислородная)
Фосфагенная и гликолитическая работают по анаэробному пути, окислительная — по аэробному.
Системы отличаются друг от друга по энергетической ёмкости и мощности[Мф40] .
Энергетическая ёмкость системы — это максимальное количество образуемой энергии, или максимальное количество АТФ, которое может_ресинтезироваться за счет_энергии рассматриваемой системы (табл. 210210903).
Энергетическая мощность системы — это максимальное количество энергии, выделяющейся в единицу времени, или максимальное количество АТФ, образующейся в единицу времени за счет энергии рассматриваемой системы (табл. 210210903).
Емкость энергетической системы ограничивает максимальный объем работы, который может быть выполнен за счет данной системы.
Мощность системы лимитирует предельную интенсивность работы, выполняемой за счет энергии данной системы.
Таблица 210210903
Максимальная мощность и ёмкость трех энергетических систем (примерные величины из расчета на 20 кг активной мышечной массы*)
| Системы | Максимальная мощность, моль АТФ/мин | Емкость, общее число молей АТФ | |
| Фосфагенная | 3,6 | 0,5 | |
| Гликолитическая | 1,2 | 1,2 | |
| Окислительная | Окисление гликогена и глюкозы | 0,8 | |
| Окисление жиров | 0,4 |
* У «стандартного» мужчины весом 70 кг мышечная масса составляет около 40 % веса тела, т.е. примерно 30 кг. При наиболее типичных видах мышечной деятельности (локомоциях) участвует не более 2/3 всей мышечной массы. Общая масса сокращающихся мышц — активная мышечная масса — не превышает обычно 20 кг.
Степень участия трех систем в энергообеспечении (ресинтезе АТФ) зависит от силы и продолжительности мышечных сокращений, т.е. от мощности и длительности работы, а также от условий работы мышц, особенно от уровня их обеспечения кислородом.
Фосфагенная энергетическая система
К этой группе относятся высокоэнергетические фосфатные соединение АТФ и креатинфосфат (КрФ).
Восстановление (ресинтез, регенерация) АТФ осуществляется во время мышечных сокращений почти мгновенно, как только он расщепляется до АДФ.
КрФ — первый энергетический резерв мышцы, действующий как немедленный источник восстановления АТФ.
Конечными продуктами расщепления КрФ являются креатин и свободный фосфат. Энергия от расщепления КрФ немедленно и непосредственно используется для ресинтеза АТФ.
При отщеплении фосфатной группы от КрФ освобождается в среднем 10,5 ккал[V.G.41] [Мф42] .
Мощность фосфагенной системы наибольшая по сравнению с другими системами (таблица 210210903). Максимальная скорость энергообразования, выраженная в количестве АТФ, ресинтезируемого в единицу времени за счет активности фосфагенной системы, примерно в 3 раза превышает максимальную мощность гликолитической системы и в 4—10 раз — максимальную мощность кислородной системы. Поэтому фосфагенная система играет решающую роль в энергообеспечении работ предельной мощности (спринтерский бег, кратковременные мышечные усилия «взрывного характера» — при подъеме штанги, метаниях, прыжках и т. п.), осуществляемых с максимально возможными по силе и скорости (мощности) сокращениями мышц.
Емкость фосфагенной системы невелика, так как запасы АТФ и КФ в мышце весьма ограниченны (таблица 210210903). Емкость фосфагенной системы, оцениваемая запасом АТФ в 20 кг мышц и количеством АТФ, которое может ресинтезироваться за счет КрФ, содержащегося в этой же мышечной массе, составляет всего около 0,5 М АТФ, т.е. около 5 ккал.
Поэтому работа с предельной мощностью, которая обеспечивается почти исключительно анаэробной фосфагенной системой, может продолжаться с энергетической точки зрения не более нескольких секунд. Например, при спринтерском беге энергетический запрос работающих мышц может достигать около 1 ккал/с. Поскольку максимальное количество энергии, которое может обеспечить фосфагенная система, составляет около 5 ккал, такая работа теоретически может продолжаться за счет фосфагенной системы не более 5 с.
Если выполняется работа большей продолжительности, её энергетическое обеспечение не может происходить только за счет наличных запасов мышечных фосфагенов (АТФ и КрФ).
Гликолитическая энергетическая система
В основе деятельности этой системы, обеспечивающей ресинтез АТФ и КФ, лежит цепь химических реакций анаэробного расщепления гликогена (и глюкозы). Совокупность этих реакций называется анаэробный гликогенолиз, или гликолиз (или цикл Эмбден — Мейерхофа). В результате таких реакций образуется молочная кислота — лактацид. Отсюда другое название этой энергетической системы — лактацидная. В результате гликолиза освобождается энергия, которая позволяет ресинтезировать АТФ.
Энергетическими субстратами лактацидной мышечной системы служат углеводы, главным образом гликоген, запасенный в мышечных клетках, и в меньшей степени — глюкоза. Молекула гликогена — это цепочка молекул глюкозы. При расщеплении молекулы гликогена от нее последовательно отщепляются молекулы глюкозы (глюкозные единицы). В мышечные клетки поступает также глюкоза из крови. В свою очередь, глюкоза попадает в кровь из печени. Анаэробное расщепление гликогена (гликогенолиз) в мышечных клетках имеет энергетическое преимущество перед гликолизом (анаэробным расщеплением глюкозы), так как каждая глюкозная единица из молекулы мышечного гликогена в процессе гликолиза дает энергию для образования 3 молекул АТФ, а каждая молекула глюкозы — только для 2 молекул АТФ.
В результате анаэробного гликогенолиза из каждой глюкозной единицы (глюк. ед.) образуются 2 молекулы молочной кислоты. Химические реакции анаэробного гликогенолиза (гликолиза) происходят исключительно в саркоплазме, где находятся все ферменты, катализирующие гликолиз (гликолитические ферменты).
Анаэробный гликогенолиз развертывается практически с начала мышечной работы, но достигает максимальной мощности (наибольшей скорости образования молочной кислоты) лишь через 30—40 с. Поэтому наибольшие концентрации молочной кислоты в мышцах (и в крови) обнаруживаются лишь при работах, которые продолжаются более 20—30 с. При более кратковременной работе энергетическая роль лактацидной системы относительно меньше.
Мощность лактацидной системы мышц велика. При работах очень большой мощности, которые могут продолжаться лишь несколько десятков секунд, скорость расходования гликогена в мышцах достигает 10 мМ глюк.ед/кг веса мышцы в минуту или около 0,2 г/мин (рис[V.G.43] .).
Определение содержания и расходования в мышцах гликогена и других энергетических субсгратов у человека стало возможным благодаря использованию метода игольчатой биопсии — взятию малой порции мышечной ткани с помощью специальной иглы (Д.Бергстрём) до работы и в разные периоды ее выполнения, а также после работы.
Таким образом, скорость расщепления гликогена в активной мышечной массе в 20 кг равна 200 мМ глюк. ед/мин. Если расщепление гликогена идет анаэробным путем, то при этом образуется 400 мМ молочной кислоты в минуту. Образование каждой молекулы молочной кислоты из гликогена обеспечивает ресинтез 3 молекул АТФ. Следовательно, при образовании 400 мМ молочной кислоты ресинтезируется 1,2 М АТФ/мин. Такова примерная максимальная мощность лактацидной системы (см. табл. 2).
Мощность лактацидной системы в 1,5 раза выше, чем кислородной, но примерно в 3 раза ниже мощности фосфагенной системы (табл. 210210903).
Лактацидная система играет решающую роль в энергетическом обеспечении работ очень большой мощности, которые могут продолжаться от 20 с до 1—2 мин и связаны с сильными мышечными сокращениями, требующими большой скорости расщепления и соответственно ресинтеза АТФ. К таким видам мышечной работы относятся, например, бег на дистанциях от 200 до 800 м, плавание на дистанциях от 50 до 200 м и т.д.
По мере дальнейшего уменьшения мощности выполняемой работы (увеличения её продолжительности) энергетическая роль лактацидной системы снижается.
Лактацидная система функционирует в тех случаях, когда сокращающиеся мышцы испытывают недостаток в снабжении кислородом. Такие условия возникают:
при работах очень большой мощности,
в самом начале любой работы когда снабжение мышц кислородом отстает от потребности в нем [p],
при статическом сокращении мышц даже небольшой силы (превышающей 20% от максимальной).
При статическом сокращении мышц даже небольшой силы (превышающей 20% от максимальной) из-за внутримышечного давления резко ограничивается кровоснабжение, а, следовательно, и обеспечение мышц кислородом.
Образующаяся в процессе анаэробного гликогенолиза (гликолиза) молочная кислота медленно диффундирует в кровь. Поэтому по мере образования она скапливается внутри сокращающихся мышечных клеток, что ведет к сдвигу их активной реакции в кислую сторону. По мере накопления молочной кислоты происходит угнетение вплоть до полного торможения активности ключевых гликолитических ферментов мышечных клеток, в частности фосфорилазы и фосфофруктокиназы. В результате снижается скорость гликолиза (образования молочной кислоты) и количество энергии (АТФ), образующейся в единицу времени за счет лактацидной системы.
Таким образом, скорость образования (концентрация) в сокращающихся мышцах молочной кислоты регулируется по механизму отрицательной обратной связи — торможением — конечным продуктом реакции: чем больше скорость образования (накопления) молочной кислоты, тем сильнее торможение, замедляющее анаэробный гликогенолиз (гликолиз). Поэтому емкость лактацидной системы в основном лимитируется не содержанием ее энергетических субстратов — углеводов (мышечного гликогена и глюкозы крови), а количеством (концентрацией) конечного продукта деятельности этой системы — молочной кислоты.
В этой связи при выполнении, мышечной работы, энергетическое обеспечение которой осуществляется преимущественно лактацидной системой, никогда не происходит резкого истощения гликогена в работающих мышцах, а тем более в печени.
Ёмкость лактацидной системы. Подсчитаем её.
Среднее содержание гликогена в мышцах — около 15 г/кг, или около 80 мМ глюк. ед (молекулярный вес глюкозы = 180). При работах с максимальной для лактацидной системы нагрузкой в результате анаэробного гликогенолиза в рабочих мышцах расходуется, по-видимому, не более 20 мМ глюк. ед/кг. Таким образом, во всей активной мышечной массе (20 кг) расходуется в целом примерно 400 мМ, или 0,4 М глюк. ед. При анаэробном глюкогенолизе 1 М глюк. ед. дает энергию, достаточную для образования 3 М АТФ.
Следовательно, при анаэробном расщеплении 0,4 М глюк. ед. возможен ресинтез 1,2 М АТФ, что эквивалентно общей энергопродукции за счет анаэробного расщепления мышечного гликогена в 12000 ккал. Эти величины и характеризуют максимальную энергетическую емкость лактацидной системы (см. табл. 210210903).
Другой способ подсчета емкости лактацидной системы — по максимальному количеству образующейся в мышцах молочной кислоты.
Максимальная концентрация молочной кислоты в работающих мышцах достигает у нетренированного человека 250 мг/100 г., или 2,5 г/кг. В этом случае во всей массе активных мышц (20 кг) общее содержание молочной кислоты составляет 50 г, а при учете ее молекулярного веса (90) — около 0,6 М. При анаэробном расщеплении одной глюкозной единицы гликогена освобождается энергия, достаточная для ресинтеза 3 молекул АТФ. При этом образуются 2 молекулы молочной кислоты.
Следовательно, при образовании 0,6 М молочной кислоты освобождается энергия, достаточная для ресинтеза — 0,9 М АТФ (около 9 ккал).
В действительности же при работе общее количество образующейся в мышцах молочной кислоты превышает рассчитанное по ее максимальной концентрации, так как молочная кислота непрерывно диффундирует из мышечных клеток в кровь. Следовательно, и емкость лактацидной системы должна превышать те величины, которые рассчитываются по максимальной концентрации лактата. Энергетическая емкость лактацидной системы примерно в 2,5 раза больше, чем фосфагенной системы (табл. 210210903).
Кислородная (окислительная) энергетическая система
При непрерывном поступлении кислорода в митохондрии мышечных клеток действует кислородная система энергопродукции (ресинтеза АТФ). При работах аэробного характера с повышением интенсивности (мощности) выполняемой нагрузки увеличивается количество O2, потребляемого мышцами (и всем организмом в целом) в единицу времени.
Поскольку между скоростью потребления O2 (л/мин) и мощностью работы (Вт) аэробного характера существует прямо пропорциональная (линейная) зависимость, интенсивность аэробной работы можно характеризовать скоростью потребления кислорода (рис[V.G.44] .). При определенной, индивидуальной для каждого человека нагрузке достигается максимально возможная для него скорость потребления O2 — максимальное потребление кислорода (МПК). Для физиологической оценки относительной мощности выполняемой человеком аэробной мышечной работы её оценивают по относительной скорости потребления O2, т.е. выраженному в процентах отношению скорости потребления O2 (л/мин) при выполнении данной работы к МПК (% МПК).
Энергетические субстраты. Для энергетического обеспечения мышечной работы кислородная система может использовать в качестве субстратов окисления («горючего») все основные питательные вещества — углеводы (гликоген и глюкозу), жиры (жирные кислоты) и белки (аминокислоты).
Вклад белков (аминокислот) в общую аэробную энергопродукцию мышц весьма мал и потому может практически не учитываться. До тех пор, пока углеводы и жиры доступны мышце, именно они используются как субстраты аэробной (окислительной) энергопродукции для поддержания мышечных сокращений.
Соотношение между окисляемыми углеводами и жирами определяется относительной мощностью аэробной работы (% МПК): чем больше относительная мощность выполняемой работы, тем относительно выше энергетический вклад окисляемых углеводов и соответственно меньше вклад окисляемых жиров в общую энергопродукцию работающих мышц (рис[V.G.45] .).
Во время выполнения легкой работы при потреблении O2 до 50 % МПК (с предельной продолжительностью до нескольких часов) большая часть энергии для сокращающихся мышц образуется за счет окисления жиров. Во время более тяжелой работы (при потреблении O2 более 60 % МПК) значительную часть энергопродукции обеспечивают углеводы. При работах, близких к МПК, подавляющая часть аэробной энергопродукции идет за счет окисления углеводов.
Таким образом, при работе очень большой мощности основными энергетическими субстратами в работающих мышцах служат углеводы. Они расщепляются главным образом аэробно (окисляются) при работе продолжительностью до нескольких десятков минут и в значительной мере анаэробно (гликолитически) при менее продолжительной работе.
Аэробное расщепление углеводов (гликогена и глюкозы) вплоть до стадии образования пировиноградной кислоты идет по тому же пути, что и при анаэробном гликолизе. В последнем случае из-за недостатка O2 (или уменьшения отношения НАД / НАДН+) пировиноградная кислота превращается (восстанавливается) в молочную кислоту при участии фермента лактатдегидрогеназы (ЛДГ). В аэробных условиях пировиноградная кислота не восстанавливается до молочной кислоты, а окисляется. При этом образуются конечные продукты окисления — CO2 и H2O. Суммарное уравнение для аэробного расщепления (окисления) глюкозы (C6H12O6) выглядит следующим образом:
Из этого уравнения следует, что при полном аэробном расщеплении одной молекулы глюкозы образуются 38 молекул АТФ, из которых 2 молекулы АТФ образуются на гликолитическом этапе расщепления глюкозы до пировиноградной кислоты, а 36 молекул АТФ — за счет кислородной системы. На этапах анаэробного расщепления из одной глюкозной единицы мышечного гликогена образуются 3 молекулы АТФ. Поэтому при полном окислении одной глюкозной единицы гликогена образуются 39 молекул АТФ (3 + 36). Следовательно, аэробное расщепление глюкозы дает в 19 раз больше продукции АТФ, чем анаэробный гликолиз (38 молекул АТФ против 2 молекул АТФ).
Окисление глюкозной единицы из мышечного гликогена обеспечивает образование в 13 раз большего числа молекул АТФ, чем при анаэробном гликогенолизе (39 молекул АТФ против 3 молекул АТФ).
Таким образом, с точки зрения расходования горючего (углеводов) кислородная система значительно более эффективна, чем анаэробная лактацидная: при одинаковом расходовании углеводов за счет их окислительного расщепления ресинтезируется во много раз больше АТФ, чем при анаэробном гликогенолизе (гликолизе).
Приведенное выше уравнение позволяет также вычислить кислородный эквивалент АТФ при окислении углеводов, т.е. количество O2, необходимое для окислительного ресинтеза 1 моля АТФ.
Как следует из уравнения, для образования 1 моля АТФ при аэробном расщеплении глюкозы требуется 0,158 молей O2 (6 : 38). 1 моль O2, равен 22,4 л O2, следовательно, на образование 1 моля АТФ требуется 3,54л O2 (0,158х22,4). Для образования же 1 моля АТФ при аэробном расщеплении гликогена требуется несколько меньшее количество кислорода: (6 : 39) × 22,4 = 3,45 л O2. Соответственно при окислении гликогена расходование 1 моля O2 ведет к образованию 6,5 моля АТФ (39 : 6), а при окислении глюкозы — 6,3 моля АТФ (38 : 6).
Таким образом, с точки зрения энергетической эффективности O2 мышечный гликоген — более выгодный субстрат для аэробной энергопродукции по сравнению с глюкозой, которую мышцы получают из крови.
Мышечный гликоген является предпочтительным субстратом окисления во время интенсивной мышечной работы. Скорость его расходования находится в прямой связи с относительной мощностью работы (% МПК) и в обратной связи с содержанием в мышце. Чем больше мощность работы (сила сокращений мышц), тем выше скорость расходования мышечного гликогена (см. рис[V.G.46] .). До мощности работы примерно 70 % МПК гликоген подвергается главным образом (если не исключительно) аэробному гликогенолизу. При более высоких нагрузках резко увеличивается скорость анаэробного гликогенолиза. При анаэробном гликогенолизе ресинтезируется в 13 раз меньше АТФ, чем при аэробном расщеплении гликогена. Поэтому для образования одинакового количества АТФ скорость расходования гликогена при анаэробном расщеплении (лактацидной системой) должна в 13 раз превышать скорость расходования гликогена окислительной системой.
Это объясняет резкое (экспоненциальное) повышение скорости расходования гликогена при увеличении мощности работы сверх 70% МПК.
По мере уменьшения содержания мышечного гликогена в процессе работы постоянной мощности скорость его расходования снижается (см. рис. 23, Б), а использование работающими мышцами глюкозы из крови увеличивается. Скорость утилизации глюкозы из крови работающими мышцами также растет с увеличением мощности работы (в дополнение к повышению скорости расходования мышечного гликогена). Таким образом, соотношение между расходованием гликогена и глюкозы зависит как от мощности, так и от продолжительности работы. Например, в начале работы на уровне 70% МПК (длительностью 1,5—2 ч) потребление глюкозы мышцами составляет лишь 10—15% от общего расхода углеводов, а в конце работы — до 50% и более.
При расчете ёмкости кислородной системы, обеспечиваемой углеводами, следует учитывать запас гликогена в мышцах и печени, а также возможность печени в процессе работы образовывать глюкозу не только за счет расщепления гликогена (гликогенолиза), но и за счет образования глюкозы (глюконеогенеза) из лактата и других веществ (аминокислот, пирувата, глицерола), попадающих в печень с кровью.
Если принять среднюю концентрацию гликогена в мышцах равной 15 г/кг, то общее его содержание в 20 кг активной мышечной массы равно 300 г (или около 1,7 М глюк. ед). Степень расходования гликогена разными мышцами неодинакова, так как зависит от степени участия каждой из мышц в выполняемой работе. В целом можно допустить, что во всех 20 кг активных мышц содержание гликогена может уменьшиться максимально на 80%. Таким образом, максимальное количество расходуемого гликогена вряд ли превышает 250 г (около 1,5 М глюк. ед.). При окислении этого количества гликогена кислородная система продуцирует энергию, достаточную для образования примерно 60 М АТФ (1,5×39).
У мужчин весом около 70 кг общий запас гликогена в печени равен примерно 70—80 г. Во время работы его расходуется не более 60 г. Если даже принять, что вся глюкоза, образующаяся из этого количества гликогена, используется мышцами, то она обеспечивает дополнительно ресинтез еще около 15 М АТФ. К последней величине следует прибавить некоторое количество глюкозы, образующейся в печени за счет глюконеогенеза. Эта величина, по-видимому, составляет не более 10—20% от общей продукции глюкозы печенью при кратковременной тяжелой и до 45% при длительной работе. Таким образом, общее количество АТФ, которое могут продуцировать рабочие мышцы за счет потребления глюкозы из крови, вряд ли превышает 20 М.
Приведенные расчеты позволяют заключить, что емкость кислородной системы, использующей в качестве субстрата окисления углеводы, составляет величину порядка 80 М АТФ, или 800 ккал. Только за счет окисления доступных запасов углеводов нетренированный человек может, например, пробежать дистанцию в 15 км.
Другим важнейшим субстратом кислородной системы служат жиры (липиды). Наибольшая часть жиров в теле человека находится в форме триглицеридов. Молекула триглицерида состоит из четырех частей — одной молекулы глицерида (глицерола) и трех молекул жирных кислот. Образование АТФ происходит в основном за счет окисления молекул жирных кислот, которые освобождаются при расщеплении триглицерида. Поэтому чаще всего говорят об образовании АТФ за счет окисления жирных кислот. Этот процесс происходит только в митохондриях и называется бета-окислением.
В среднем 1 моль смеси жирных кислот, характерных для человека, обеспечивает образование 138 молей АТФ. Таким образом, при аэробном расщеплении молекулы жирной кислоты образуется в несколько раз больше молекул АТФ, чем при аэробном распаде молекулы глюкозы (или глюкозной единицы из гликогена).
При аэробном использовании 1 моля глюкозы образуется 38 молей АТФ, т. е. 1 г окисленной глюкозы обеспечивает ресинтез 0,21 М АТФ. Это соответствует 2,1 ккал. При полном сгорании 1 г углеводов в калорической бомбе выделяется, как известно, 4,1 ккал. В мышце 2,1 ккал идет на ресинтез фосфагенов, а остальные 2 ккал (около 50 % энергии) выделяются в виде тепла. То же справедливо и в отношении жиров, полная энергия 1 г которых равна 9,1 ккал, однако для ресинтеза АТФ используются лишь 5 ккал.
При окислении 1 моля пальмитиновой кислоты (молекулярный вес = 256) образуется 128 молей АТФ, т.е. каждый грамм этой жирной кислоты при полном окислении обеспечивает ресинтез 0,5 моля АТФ, что эквивалентно примерно 5 ккал. Иначе говоря, при одинаковом по весу расходе углеводов и жиров последние могут обеспечить ресинтез в два с лишним раза большего количества АТФ, чем при окислении углеводов.
Жиры обладают наибольшей энергетической емкостью из всех остальных мышечных источников энергии. 1 моль АТФ дает около 10 ккал, 1 моль КФ — около 10,5 ккал, 1 моль глюкозы при анаэробном расщеплении — около 50 ккал, при аэробном расщеплении (окислении) — около 700 ккал, а 1 моль жиров при окислении обеспечивает 2400 ккал.
Необходимо отметить, что запасы жиров в теле человека велики — жиры составляют от 10 до 30 % его веса. Только небольшая часть жиров (до 5 % всех жировых запасов) находится в мышцах в виде триглицеридов, концентрация которых в мышцах составляет от 5 до 15 г/кг. Основная масса жира находится в жировых депо тела. Отсюда освободившиеся при расщеплении (липолизе) свободные жирные кислоты попадают в ток крови, а затем захватываются из крови мышцами.
Во время работы мышцы используют как свободные жирные кислоты из крови, так и собственные освобождающиеся из триглицеридов. При работе на уровне 50 – 70 % МПК вклад этих двух источников примерно одинаков. Приблизительные подсчеты показывают, что за счет окисления всех запасенных в теле жиров активная мышечная масса в 20 кг может ресинтезировать несколько тысяч молей АТФ (см. табл. 210210903). Эта величина характеризует огромную энергетическую емкость кислородной системы, использующей жиры в качестве субстрата окисления. Запасов жиров вполне достаточно для энергетического обеспечения непрерывной ходьбы в течение 7—10 дней.
Кислородная система, использующая в качестве субстратов окисления как углеводы, так и жиры, обладает наибольшей энергетической емкостью из всех других энергетических систем. Ее емкость во много тысяч (!) раз превышает емкость лактацидной и фосфагенной систем (см. табл. 210210903). Поэтому кислородная энергетическая система обеспечивает возможность выполнения работы большой продолжительности — от нескольких минут до многих часов и даже дней.
Приведенные выше данные позволяют сопоставить энергетические характеристики углеводов и жиров как субстратов окисления (табл. 3). Жиры являются самым легким «горючим», если сравнивать энергию, содержащуюся в единице веса вещества (удельная энергетическая емкость). Для образования одинакового количества АТФ расходуется по весу почти в 2,5 раза меньше жиров, чем глюкозы. В мышце каждый грамм гликогена связан с 2,7 г воды. Поэтому каждому грамму гликогена соответствует 3,7 г комплекса «гликоген + вода». Это снижает энергетическую удельную емкость мышечного гликогена до 0,07 моль АТФ/г комплекса «гликоген + вода». В действительности гликоген — самое тяжелое «горючее». Жиры имеют также преимущество перед углеводами по общей энергетической емкости. Все это делает их пригодными для выполнения длительной работы относительно небольшой мощности.
Однако углеводы имеют серьезное преимущество перед жирами, когда сравнивается количество АТФ, образующееся на единицу потребляемого O2, (энергетическая эффективность O2). При таком сравнении углеводы на 10—13% эффективнее, чем жиры. Если выполняется очень интенсивная аэробная работа (близкая к МПК, т. е. околомаксимальная аэробная работа), она в большой степени лимитируется скоростью потребления O2. В этом случае углеводы как субстрат окисления имеют неоспоримое преимущество перед жирами, так как для образования одинакового количества энергии (АТФ) при окислении углеводов затрачивается меньшее количество O2, чем при окислении жиров. Особенно эффективно в этом случае окисление мышечного гликогена, имеющего наибольшую энергетическую эффективность O2.
Таблица 210231404
Энергетические характеристики окислительного использования углеводов и жиров
| Показатели | Мышечный гликоген (глюк.ед.) | Глюкоза | Жирные кислоты |
| Удельная энергетическая емкость: | |||
| моль АТФ/моль субстрата | |||
| моль АТФ/г субстрата | 0,22 | 0,22 | 0,50 |
| Общая энергетическая емкость (на 20 кг мышечной массы), моль АТФ | |||
| Энергетическая эффективность O2, моль АТФ/моль O2 | 6,50 | 6,34 | 5,61 |
| Кислородный эквивалент АТФ, л O2 / моль АТФ | 3,45 | 3,54 | 4,05 |
| Максимальная мощность (на 20 кг активной мышечной массы), моль АТФ/моль субстрата · с-1 | 0,8 | 0,5 | 0,4 |
Наконец, общее количество энергии (АТФ), которое может продуцироваться в единицу времени за счет окисления углеводов (особенно мышечного гликогена), вдвое больше, чем при окислении жиров. Такая относительно сниженная мощность энергопродукции при окислении жиров по сравнению с окислением углеводов позволяет понять, почему участие жиров в энергообеспечении уменьшается по мере повышения мощности выполняемой работы.
[a] Мышечное волокно
[b] Структурные компоненты миона: эндомизий (коллагеновые волокна, фиброциты, гемокапилляр, моторная бляшка) + мышечное волокно (базальная мембрана, миосателлитоцит, миосимпласт ).
[c] Синонимы – нейро‑моторная единица, двигательная единица.
[d] Выраженность этой способности у различных атипичных кардиомиоцитов существенно разнится. У рабочих кардиомиоцитов практически нет.
[e] Расположены в капсуле, в составе нервно-мышечного веретена fusus neuromuscularis
[f] Синоним — фазические
[g] Под световым микроскопом мышечное веретено представляет собой вытянутую структуру, расширенную посередине за счет капсулы и напоминающую по форме веретено, что и обусловило ее название.
[h] Тонус [лат. tonus < гр. tonos напряжение] — состояние длительного возбуждения нервной или мышечной ткани, не сопровождающееся развитием утомления.
[i] Подробнее читайте в Приложение 709261417. Энергетика мышечного сокращения.
[j] Синоним – окислительные.
[k] Huxley H.E., Hanson J.
[l] Синоним – электромеханический каплинг
[m] Потенциалуправляемых (электроуправляемых)
[n] Также у нейронов
[o] инозитолтрисфосфатом
[p] Разность между запросом и реальным потреблением кислорода в этот период называется кислородным дефицитом (см. 19.3).
[1] Необходимо знать ХОРОШО! С материалом этого раздела Вы должны быть уже знакомы.
[2] Необходимо знать ХОРОШО! Но с материалом этого раздела Вы должны быть уже хорошо знакомы. Подробности в учебнике.
[3] Необходимо знать ХОРОШО! Но с материалом этого раздела Вы должны быть уже хорошо знакомы. Подробности в учебнике.
[4] Необходимо знать ХОРОШО! Но с материалом этого раздела Вы должны быть уже хорошо знакомы.
[5] Необходимо знать ОЧЕНЬ ОЧЕНЬ ХОРОШО, в том числе и двоечникам! Но с материалом этого раздела Вы должны быть уже хорошо знакомы.
[6] Это Вы должны уже знать. Если забыли, вспомните. Воспроизводить могут только отличники и хорошисты.
[7] Необходимо знать ХОРОШО! Но с материалом этого раздела Вы должны быть уже хорошо знакомы.
[8] Необходимо знать ХОРОШО!
[9] Необходимо знать ХОРОШО! Подробнее мы будем знакомиться с этим типом мышечных волокон при изучении сенсорных систем.
[10] Необходимо знать ХОРОШО!
[11] Для ознакомления! Подробно в учебнике.
[12] Необходимо знать ХОРОШО!
[13] Необходимо знать ОЧЕНЬ ХОРОШО!
[14] Необходимо знать ОЧЕНЬ ОЧЕНЬ ХОРОШО, в том числе и двоечникам!
[15] Необходимо знать ОЧЕНЬ ОЧЕНЬ ХОРОШО, в том числе и двоечникам!
[16] Необходимо знать ОЧЕНЬ ОЧЕНЬ ХОРОШО, в том числе и двоечникам!
[V.G.1](см. рис. 21)
[V.G.2](см. также рис. 21, е)
[Мф3]++501+C.74
[Мф4]++501+C.74
[Мф5]++501+C.74
[V.G.6](см. рис. 22, б)
[V.G.7]210211139
[V.G.8]210211146
[V.G.9]210211148
[Мф10]++760+ С.57
[Мф11]++597+ С.294
[Мф12]Вопрос 2.
[Мф13]++597+ С.294
[Мф14]
[Мф15]++597+ С.292
[Мф16]I Сам придумал
[Мф17]в презентации лучшего качества
[Мф18]++51+ С.693
[Мф19]Камкин С.329
[Мф20]++736+ С.125
[Мф21]++736+ С.125
[Мф22]Камкин С.329
[Мф23]Современная теория мышечного сокращения и расслабления.
[Мф24]++501+С.74; —147-С.23
[Мф25]—147-С.23
[Мф26]++601+С.76
[Мф27]++501+С.74
[Мф28]Распространение электрической волны внутрь волокна приводит, в свою очередь, к деполяризации мембран цистерн продольных трубочек саркоплазматического ретикулума (Коц).
[Мф29]++597+С.293
[Мф30]++501+С.72
[Мф31]++810+ С.101 Орлов С.
[Мф32]++810+ С.101 Орлов С.
[Мф33]++810+ С.101 Орлов С.
[Мф34]++810+ С.101 Орлов С.
[Мф35]++810+ С.101 Орлов С.
[Мф36]++597+ С.294 №7
[V.G.37](см. 6.5.2)
[Мф38]++597+ С.294 №8
[Мф39]++736+С.100
[Мф40]++736+С.102
[V.G.41]при расщеплении 1 М КрФ (М-1 моля) ???
[Мф42]++736+С.101
[V.G.43]23
[V.G.44]24
[V.G.45]25
[V.G.46]23
megaobuchalka.ru
Гликолитическая энергетическая система — Физкультура на 5
В основе этой системы, обеспечивающей ресинтез АТФ и КрФ, лежит цепь химических реакций анаэробного характера расщепления гликогена (и глюкозы). В результате этих реакций образуется молочная кислота (лактацид). Совокупность этих реакций называется анаэробный гликогенолиз или гликолиз. В результате гликолиза освобождается энергия, которая расходуется на ресинтез АТФ. Энергетическими субстратами лактацидной энергетической системы являются углеводы, главным образом гликоген, запасенный в мышечных клетках и в меньшей степени глюкоза.
Молекула гликогена
Молекула гликогена – это цепочка молекул глюкозы. При расщеплении молекулы гликогена от нее последовательно отщепляются молекулы глюкозы (глюкозные единицы). В мышечные клетки поступает также глюкоза из крови. В свою очередь в кровь она поступает из печени. Анаэробное расщепление гликогена в мышечных клетках (гликогенному) имеет энергетическое преимущество перед анаэробным расщеплением глюкозы (гликолиз) так как в первом случае при расщеплении одной глюкозной единицы выделяется энергия для образования 3х молекул АТФ, а во втором – только две. В результате гликогенолиза из каждой глюкозной единицы образуется две молекулы молочной кислоты. Анаэробный гликогенолиз разворачивается практически с начала мышечной работы, но достигает максимальной мощности ( наибольшей скорости образования La лишь через 30-40 сек. Поэтому наибольшие концентрации Za в мышцах (и крови) обнаруживаются при работах которые продолжаются более 20-30 секунд. При более кратковременной работе роль лактацидной системы менее значима.
Лактацидная система мышц характеризуется большой мощностью, т.е. высокой скоростью энергообразования. При работах очень большой мощности, которые могут продолжаться лишь несколько десятков секунд, скорость расходования гликогена достигает 10 мМ глюк. ед/кг веса мышцы в минуту или около 0,2 г/мин. Образование каждой молекулы Lа из гликогена обеспечивает Ресинтез 3-х молекул АТФ. Мощность лактацидной системы в 1,5 раза выше, чем кислородной, но примерно в 3 раза ниже мощности фосфагенной. Лактацидная система играет решающую роль в энергообеспечении работ очень большой мощности, которые продолжаются от 20 сек до 1-2 мин и связаны с сильными мышечными сокращениями, требующими большой скорости расщепления и, соответственно, ресинтеза АТФ (бег 200-800 м, плавание 50-200 м. и т.д.). По мере уменьшения мощности работы (увеличения продолжительности) роль лактацидной системы снижается. Лактацидная система функционирует тогда. Когда сокращающиеся мышцы испытывают недостаток в снабжении кислородом. Такие условия возникают не только при работах большой мощности, но и в самом начале любой работы, когда снабжение мышц О2 отстает от потребности в нем (кислородный дефицит), а также при статическом сокращении мышц даже небольшой силы когда из-за внутреннего давления в мышцах резко ограничивается кровоснабжение, а следовательно, и обеспечение мышц О2 Образующаяся в процессе анаэробного гликогенолиза, Lа медленно дифференцирует в кровь. Она накапливается в мышечных клетках, что ведет к сдвигу их активной реакции в кислую сторону. Накопление Lа вызывает угнетение вплоть до полного торможения активности ключевых гликолитических ферментов мышечных клеток, в частности фосфорилазы фосфоруктокиназы. В результате снижается скорость гликолиза (образование Lа) и количество энергии (АТФ), образующейся в единицу времени за счет лактацидной системы. Таким образом, скорость образования Lа в сокращающихся мышцах регулируется по принципу обратной отрицательной связиторможением — конечным продуктом реакции:чем больше скорость накопления Lа, тем сильнее торможение, замедляющее анаэробный гликогенолиз (гликолиз). Поэтому емкость лактацидной системы в основном лимитируется не содержанием ее энергетических субстратов (углеводов мышечного гликогена и глюкозы крови), а количеством конечного продукта этой системы – Lа. Емкость лактацидной системы можно рассчитать по количеству Lа в мышцах. Максимальная концентрация Lа в мышцах нетренированных людей 250 мг/100 г или 2,5 /кг. В этом случае во всей массе активных мышц (20 кг) общее содержание Lа составит 50 г, а при учете ее молекулярного веса (90) – около 0,6 моль. При анаэробном расщеплении глюкозной единицы гликогена освобождается энергия, достаточная для ресинтеза 3 молекул АТФ. При этом образуется 2 молекулы Lа. Следовательно, при образовании 0,6 моль Lа освобождается энергия, достаточная для ресинтеза 0,9 моль АТФ (около 9 ккал). Энергетическая емкость лактацидной системы примерно в 5 раза больше фосфагенной системы.
fizkultura-na5.ru
АТФ в бодибилдинге — SportWiki энциклопедия
АТФ (аденозин трифосфат: аденин, связанный с тремя фосфатными группами) — молекула, которая служит источником энергии для всех процессов в организме, в том числе для движения. Сокращение мышечного волокна происходит при одновременном расщеплении молекулы АТФ, в результате чего выделяется энергия, которая идёт на осуществление сокращения. В организме АТФ синтезируется из инозина.
АТФ должна пройти через несколько ступеней, чтобы дать нам энергию. Сначала при помощи специального коэнзима отделяется один из трёх фосфатов (каждый из которых даёт десять калорий), высвобождается энергия и получается аденозин дифосфат (АДФ). Если энергии требуется больше, то отделяется следующий фосфат, формируя аденозин монофосфат (АМФ). Главным источником для производства АТФ служит глюкоза, которая в клетке инициально расщепляется на пируват и цитозол.
Во время отдыха происходит обратная реакция – при помощи АДФ, фосфагена и гликогена фосфатная группа вновь присоединяется к молекуле, формируя АТФ. Для этих целей из запасов гликогена берётся глюкоза. Вновь созданный АТФ готов к следующему использованию. В сущности АТФ работает как молекулярная батарея, сохраняя энергию, когда она не нужна, и высвобождая в случае необходимости.
Структура АТФ[править | править код]
Молекула АТФ состоит из трёх компонентов:
1. Рибоза (тот же самый пятиуглеродный сахар, что формирует основу ДНК)
2. Аденин (соединённые атомы углерода и азота)
3. Трифосфат
Молекула рибозы располагается в центре молекулы АТФ, край которой служит базой для аденозина. Цепочка из трёх фосфатов располагается с другой стороны молекулы рибозы. АТФ насыщает длинные, тонкие волокна, содержащие протеин, называемый миозином, который формирует основу наших мышечных клеток.
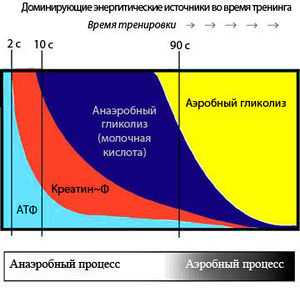 Последовательное включение энергетических систем во время выполнения упражнений
Последовательное включение энергетических систем во время выполнения упражненийЗапасов АТФ достаточно только на первые 2-3 секунды двигательной активности, однако мышцы могут работать только при наличии АТФ. Для этого существуют специальные системы, которые постоянно синтезируют новые молекулы АТФ, они включаются в зависимости от продолжительности нагрузки (см. рисунок). Это три основные биохимические системы:
1. Фосфагенная система (Креатин-фосфат)
2. Система гликогена и молочной кислоты
3. Аэробное дыхание
Фосфагенная система[править | править код]
Когда мышцам предстоит короткая, но интенсивная активность (приблизительно 8-10 секунд), используется фосфагенная система – АДФ соединяется с креатина фосфатом. Фосфагенная система обеспечивает постоянную циркуляцию небольшого количества АТФ в наших мышечных клетках. Мышечные клетки также содержат высокоэнергетический фосфат – фосфат креатина, который используется для восстановления уровня АТФ после кратковременной, высокоинтенсивной работы. Энзим креатин киназа отнимает фосфатную группу у креатина фосфата и быстро передаёт её АДФ для формирования АТФ. Итак, мышечная клетка превращает АТФ в АДФ, а фосфаген быстро восстанавливает АДФ до АТФ. Уровень креатина фосфата начинает снижаться уже через 10 секунд высокоинтенсивной активности. Пример использования фосфагенной системы энергоснабжения – это спринт на 100 метров.
Система гликогена и молочной кислоты[править | править код]
Система гликогена и молочной кислоты снабжает организм энергией медленнее, чем фосфагенная система, и предоставляет достаточно АТФ примерно для 90 секунд высокоинтенсивной активности. В ходе процесса из глюкозы мышечных клеток в результате анаэробного метаболизма происходит формирование молочной кислоты.
Учитывая тот факт, что в анаэробном состоянии организм не использует кислород, эта система даёт кратковременную энергию без активации кардио-респираторной системы точно так же, как и аэробная система, но с экономией времени. Более того, когда в анаэробном режиме мышцы работают быстро, они очень мощно сокращаются, перекрывая поступление кислорода, так как сосуды оказываются сжатыми. Эту систему ещё можно назвать анаэробно-респираторной, и хорошим примером работы организма в этом режиме послужит 400-метровый спринт. Обычно продолжать работать таким образом атлетам не даёт мышечная болезненность, возникающая в результате накопления молочной кислоты в тканях.
Аэробное дыхание[править | править код]
Если упражнения длятся более двух минут, в работу включается аэробная система, и мышцы получают АТФ вначале из углеводов, потом из жиров и наконец из аминокислот (протеинов). Протеин используется для получения энергии в основном в условиях голода (диеты в некоторых случаях). При аэробном дыхании производство АТФ проходит наиболее медленно, но энергии получается достаточно, чтобы поддерживать физическую активность на протяжении нескольких часов. Это происходит, потому что глюкоза распадается на диоксид углерода и воду беспрепятственно, не испытывая противодействия со стороны, например, молочной кислоты, как в случае анаэробной работы.
sportwiki.to
