
Общий обзор строения и функций нервной системы
Центральный и периферический отделы. В нервной системе различают центральный и периферический отделы (цв. табл. V). К центральному отделу относятся спинной мозг, расположенный в позвоночном канале, и головной мозг, находящийся внутри мозгового черепа. Снаружи головной и спинной мозг покрыты оболочками.
На разрезе мозга (как головного, так и спинного) можно заметить, что в одних местах нервная ткань имеет более темный оттенок— серое вещество, а в других более светлый — белое вещество. Серое вещество мозга состоит из огромного количества нервных клеток; его белое вещество представляет собой скопление нервных волокон, по которым проходит возбуждение.
Часть нервных волокон выходит за пределы мозга, образуя нервы, которые направляются к различным органам тела. Совокупность нервов составляет периферический отдел нервной системы.
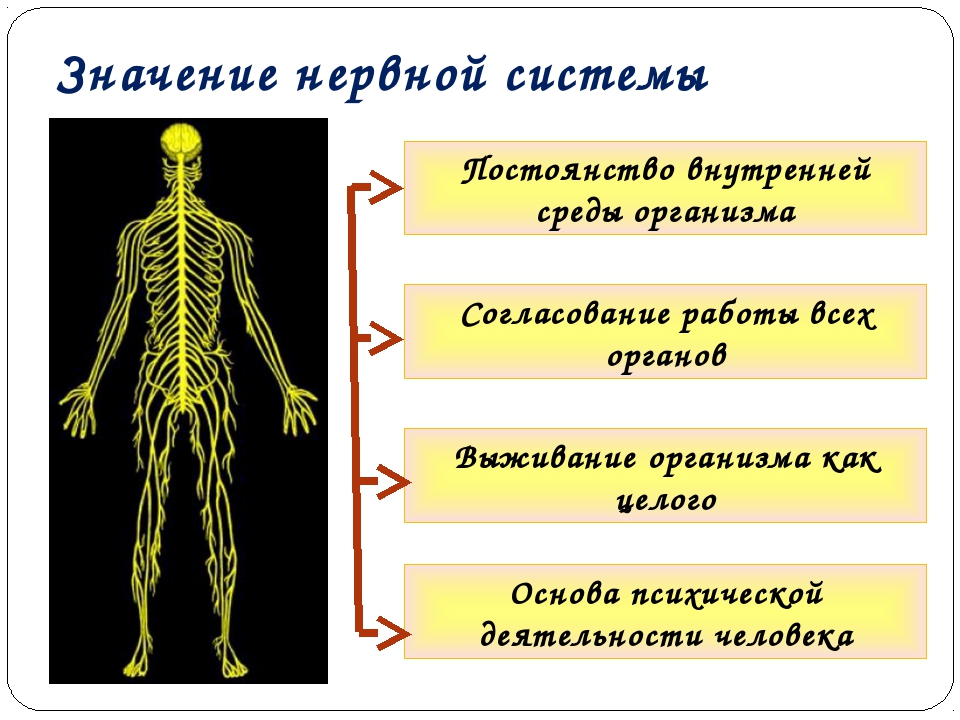
Значение нервной системы. Существование организма невозможно без координации, т. е. согласования деятельности отдельных органов. Точной и быстрой координации требует выполнение любого двигательного акта. Согласованно должны работать и другие органы. Так, в зависимости от выполняемой работы мышцы по-разному должны снабжаться кровью, что обеспечивается сужением или расширением сосудов; одновременно изменяется частота и сила сердечных сокращений и дыхательных движений. Регуляция и координация работы органов, строгое подчинение их деятельности потребностям организма как единого целого осуществляются нервной системой. Она обеспечивает приспособление организма к постоянно меняющимся условиям жизни, ориентировку в окружающем мире. Нервная система может быть названа органом не только регуляции и координации деятельности организма, но и позна-
ния окружающего мира С деятельностью нервной системы связано мышление человека и все его поведение.
Рефлекс — основа нервной деятельности. С каждым органом тела нервная система связана двойными путями: по одним она получает информацию о том, что происходит как в самом организме, так и в окружающей среде, а по другим посылает ответные импульсы, воздействующие на работу органа. Нервные волокна, передающие возбуждение в центральную нервную систему, называются афферентными (т. е. приносящими) или центростремительными, а передающие его органам называются эфферентными (т. е. уносящими) или центробежными
Реакции организма, возникающие в ответ на раздражение и протекающие при участии нервной системы, называются рефлексами. Они лежат в основе всей деятельности нервной системы Всякое изменение, возникающее в любом органе тела, вызывает возбуждение соответствующих окончаний центростремительных нервов, что и влечет за собой рефлекторные реакции органов.
Путь, по которому пробегают импульсы, вызывающие ответную реакцию, называется рефлекторной дугой (рис 25).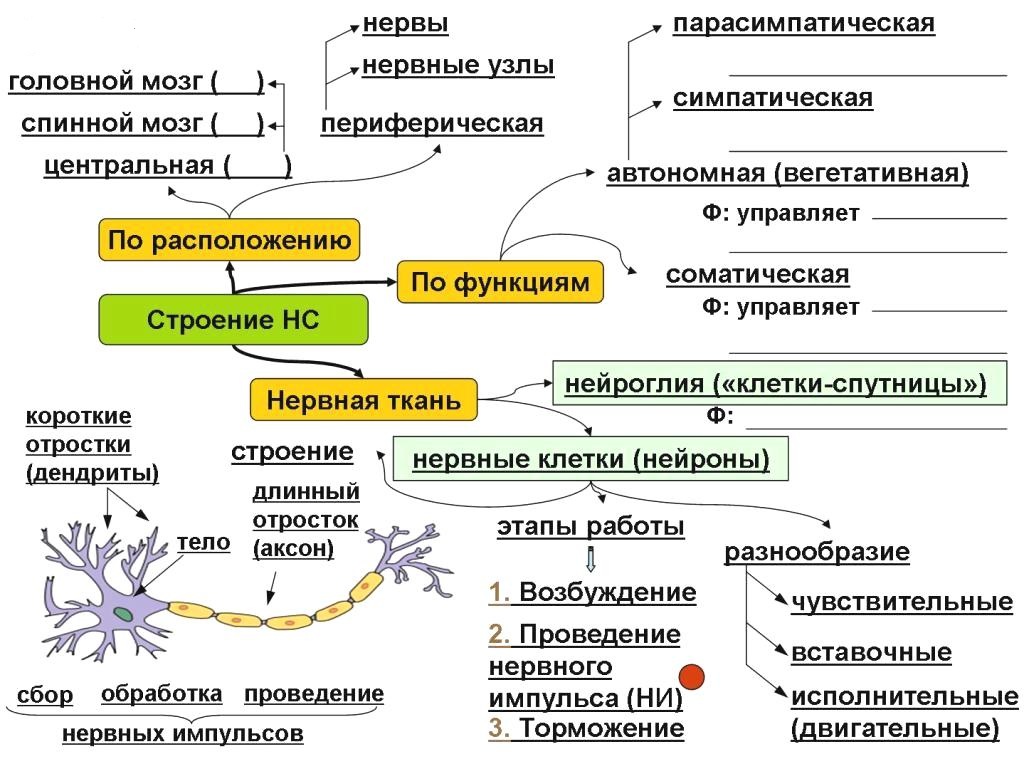 а’также центры внутренних органе!Нервныеволокна, составляющие основную массу белого вещества, образуют проводящие пути. Они осуществляют связь между нейронами различных отделов мозга
а’также центры внутренних органе!Нервныеволокна, составляющие основную массу белого вещества, образуют проводящие пути. Они осуществляют связь между нейронами различных отделов мозга
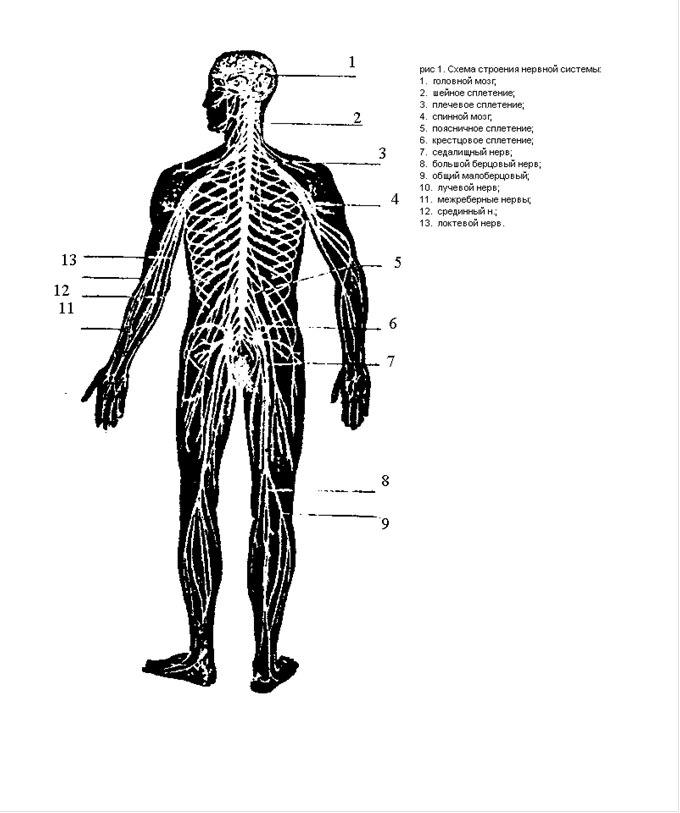
Между каждой парой соседних позвонков от спинного мозга отходят слева и справа спинномозговые нервы — всего 31 пара Каждый нерв начинается двумя корешками — передним и задним. Через задние корешки возбуждение передается с периферии в спинной мозг, а через передние—от спинного мозга к мышцам и другим органам. Поэтому педедние корешки называют двигательными, а задние чувствующими» Оба корешка по выходе-дз -мозга ел ива -ются в один нерв Таким образом, нервы, идущие от спинного
Рис 26 Спинной мозг.
вверху — поперечный разрез спинного моз га, внизу—спинной мозг в почвоночном канале / — передние корешки нервов 2 — задние корешки», 3 — пограничный симпатический ствол.
Рис 25 Общая схема рефлекторной дуги
/ — рецептор 2 — центростремительный нерв if—центральная нервная сисгема, 4 — центробежный нерв 5 — орган, даю щий ответную реакцию
Рис 27 Серое и белое вещество на поперечном разрезе головного мозга / — кора больших полушарий 2 — подкор ковые двигательные ядра, 3 — бугры промежуточного мозга 4 — подбугровая область, справа — схема разреза
мозга, смешанные, в них находятся нервные волокна афферентные, (центростремительные) и афферентные (центробежные).
Головной мозг. Строение головного мозга изображено на цветной таблице VI. Участок головного мозга, который представляет собой непосредственное продолжение спинного, называется продолговатым мозгом. Он хорошо виден на нижней поверхности головного мозга, а также если разрезать весь головной мозг в продольном направлении на две равные части (цв. табл. VII). Впереди продолговатого мозга в виде поперечного вала расположен мост.
С продолговатым мозгом и мостом связана большая часть нервов головного мозга. Сюда идут центростремительные импульсы от органа слуха, кожи головы, слизистой оболочки полости рта. Отсюда выходят центробежные импульсы ко всем мышцам головы и слюнным железам. Через блуждающий нерв продолговатый мрзг связан с органами кровообращения, пищева.йеыи.я, и дыхания. Сле-довательно,, его деятельность имеет непосредственное отношение к важнейшим жизненным отправлениям: здесь находятся центры дыхательный, сердечно-сосудистый, мимических мышц лица, жевания, глотания, рвоты, кашля, слюноотделения и многие другие.
В четверохолмии находятся центры ориентировочных рефлексов на свет и звуки. Известно, что всякий внезапный звук или вспышка света вызывает рефлекторную реакцию: человек настораживается, поворачивает голову в сторону раздражителя. Подобные реакции имеют огромное биологическое значение. Они ориентируют организм в явлениях, происходящих в окружающей среде, т. е. помогают в них разбираться. Отсюда и их название.
Остальная часть мозга называется передним мозгом. Он состоит из межуточного мозга, граничащего со средним мозгом, и
В межуточном мозге различают бугры, подбугровую область и некоторые другие участки (рис. 27). Через ядра бугров и смежных с ними образований проходят к большим полушариям афферентные пути от всех частей тела. кв. м. В коре больших полушарий находится около 14 миллиардов клеток, что, составляеГоольше половины всех нерв-ных клеток головного и спинного мозга.
кв. м. В коре больших полушарий находится около 14 миллиардов клеток, что, составляеГоольше половины всех нерв-ных клеток головного и спинного мозга.
Функция отдельных участков коры неодинакова (цб. табл, VII). Информация с органов зрения поступает в затылочную область, с органов слуха—в височную, с кожи—в заднюю центральную область (позади центральной борозды), с мышц и сухожилий—в переднюю центральную область. Внутри каждой области можно наблюдать очень дробную локализацию. Так, в передней центральной извилине каждая мышца имеет свое определенное представительство в коре, иными словами, участок, раздражение которого вызывает ее сокращение. Мышцы, принимающие участие в более сложных и разнообразных движениях, имеют большую площадь коркового-представительства. Это особенно заметно при сравнении корковых центров мышц ноги туловища, отдельных частей руки, языка, голосовых связок, С особыми участками мозга связана членораздельная человеческая речь: двигательный центр речи связан с произнесением слов, слуховой центр — со слуховым восприятием речи, зрительный — с чтением, центр письма — с написанием слов.
Большая часть нервных путей, идущих как к коре, так и от коры, перекрещивается, а потому правое полушарие связано главным образом с левой половиной тела, а левое — с правой.
от грудного и верхней части поясничного отделов спинного мозга, гГпарасимпашческие—ог некоторых участков головного мозга (в основном от продолговатого) и от нижнего отдела спинного мозга. Симпатические и парасимпатические нервы образуют единую вегетативную систему (цв. табл. VIII).
Путь от центральной нервной системы к мышцам состоит из одного нейрона: его тело расположено в сером веществе спинного или стволовой части головного мозга, а длинный отросток (аксон), не прерываясь, тянется до мышцы. замедляет их. Парасимпатические нервы иннервируют главным образом внутренние органы, а симпатические направляются ко всем органам. Симпатические волокна подходят даже к скоплениям нервных клеток в спинном и головном мозге. Столь широкое распространение- симпатических нервов объясняется тем, что их функция связана с регуляпирй процессов рйдена • веществ, проте-кающих в каждой живой клетке.
замедляет их. Парасимпатические нервы иннервируют главным образом внутренние органы, а симпатические направляются ко всем органам. Симпатические волокна подходят даже к скоплениям нервных клеток в спинном и головном мозге. Столь широкое распространение- симпатических нервов объясняется тем, что их функция связана с регуляпирй процессов рйдена • веществ, проте-кающих в каждой живой клетке.
Читайте также:
Центральная нервная система
Уникальность нервной системы – центральная нервная система сформировалась за многие тысячи лет развития человеческой расы, в том, что взаимосвязь осуществляется сразу в двух направлениях. Каждый орган и подчиняется головному мозгу, и оказывает влияние на его деятельность. Основные структурные единицы ЦНС – головной, а также спинной мозг. Именно благодаря им, люди отличаются от иных представителей животного мира.
Общая характеристика
Эволюционные процессы, которым подвергались многоклеточные организмы на протяжении столетий, привели к необходимости регулирования, а также координирования всех жизненных функций.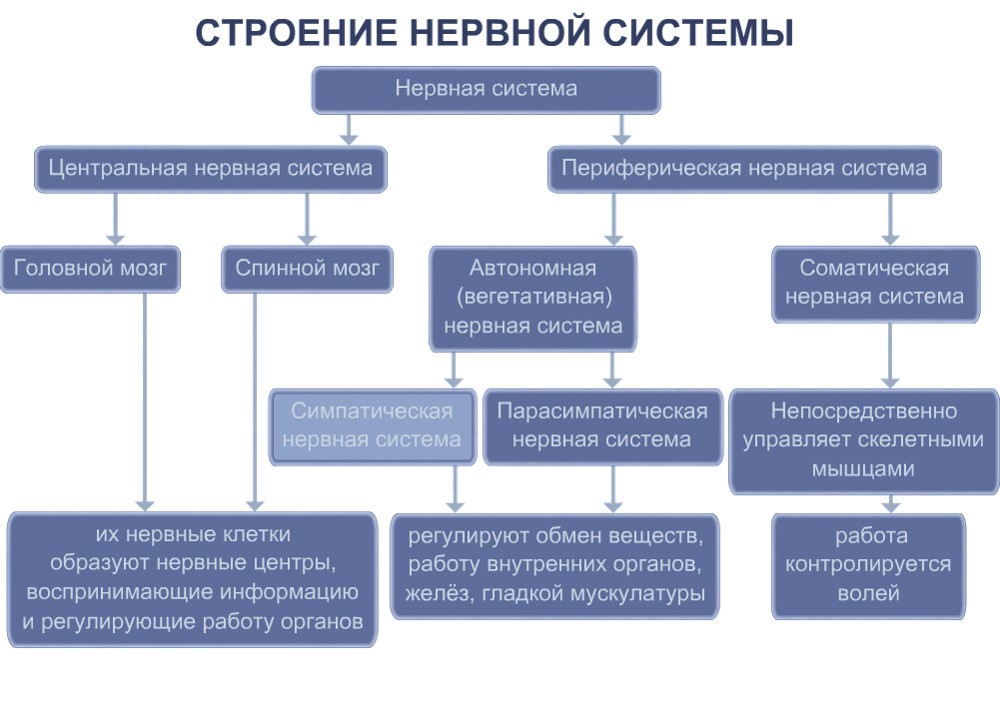
По сути, центральная нервная система – это высокоорганизованная совокупность специальных структурных единиц, которые объединяют и координируют деятельность каждой ткани, системы и органа, как изнутри, так и при взаимодействии со средой извне. Она представлена двумя важнейшими органами – внутричерепным, а также внутрипозвоночным мозгом. Тогда как черепно-мозговые нервные волокна к ЦНС не имеют отношения. Это уже периферическая система иннервации.
В основном, относящиеся к головному и спинному мозгу функциональные единицы несут ответственность за восприятие информации от внешних/внутренних раздражителей, ее переработку и формирование адекватного ответа. Благодаря этой являющейся частью целого организма системе, и обеспечивается полноценное взаимодействие людей с миром вокруг – через память, мышление, эмоции, творческие процессы.
Особенности строения
У людей с момента оплодотворения яйцеклетки начинается развитие и формирование ЦНС – из непосредственно нервной трубки образуются головной, а также спинной мозг. Их защищают костные каркасы – черепная коробка и позвонки. Ниже расположены три оболочки – твердая с паутинной и сосудистой. В их пределах находятся жидкие среды – ликвор с кровью.
Традиционно строение ЦНС подразумевает, что клетки – нейроны, объединяются в особые скопления – нервные центры. Тела нейронов образуют серое вещество, тогда как их короткие и длинные отростки – белую субстанцию, проводящие сигнальные импульсы пути.
Помимо этого, в ЦНС присутствует нейроглия, состоящая их глиальных клеток. Их количество в несколько раз превышает число нейронов. Поэтому они составляют большую часть массы центрального отдела нервной системы.
В головном отделе принято выделять несколько сегментов – мозжечок с большими полушариями, а также продолговатый, средний, промежуточный и задний участки. Каждый из них несет свою ответственность за правильное функционирование органа отдельно, и всего организма и систем в целом. В спинном мозге градация осуществляется согласно сегментам позвоночного ствола – от шейного, до грудного и пояснично-крестцового.
Каждый из них несет свою ответственность за правильное функционирование органа отдельно, и всего организма и систем в целом. В спинном мозге градация осуществляется согласно сегментам позвоночного ствола – от шейного, до грудного и пояснично-крестцового.
Анатомия головного мозга
В составе центральной нервной системы главенствующее место, безусловно, занимает головной мозг. Внутри черепной коробки он представлен двумя крупными полушариями, испещренными глубокими и мелкими бороздами, под которыми расположены иные структурные единицы:
- Продолговатый участок – локализуется на блюменбаховом скате. Книзу он плавно трансформируется в спинной мозг. На его передней поверхности определяется продольная щель, по бокам от которой специалисты выделяют 2 своеобразных возвышения в виде валиков. Их именуют пирамидами с оливами. Тогда как подобную же борозду на задней поверхности структуры с двумя задними канатиками принято называть столбами.
- Над продолговатой зоной расположен задний мозг – в форме Варолиева моста, а также мозжечка.
 Внешне схожи с большими полушариями, но функционально имеются свои особенности. В глубине ткани находятся скопления ядер, от которых берут свое начало черепно-мозговые нервы.
Внешне схожи с большими полушариями, но функционально имеются свои особенности. В глубине ткани находятся скопления ядер, от которых берут свое начало черепно-мозговые нервы. - Взаимосвязь между продолговатой единицей и вышерасположенными отделами осуществляет средний мозг – представлен ножками, нервными пучками, а также четверохолмием. Переоценить их значение невозможно – именно в этой зоне пролегает множество важнейших нервных путей и расположены ядра нескольких пар нервов.
- Промежуточный участок – известен как зрительные бугры с подбугровой областью, локализуется дальше от центра головного мозга. Они содержат первичные клетки зрительной системы, а также чувствительные проводниковые волокна. Гипоталамус, он же подбугровый участок, принимает участие в обменных процессах.
Каждая из перечисленных единиц системы – от полушарий и мозжечка, до ствола головного мозга имеет свое значение для жизнедеятельности людей. Если сбой происходит в одной зоне – оболочки ЦНС, к примеру, опухоли мозга, то влияние будет оказываться на все участки органа.
Анатомические особенности спинного мозга
Прочный каркас позвоночного столба надежно защищает еще одну структурную единицу центральной нервной системы – спинной мозг. Его протяженность впечатляет – от затылочного отверстия черепной коробки до поясничных позвонков.
Визуально с двух его сторон можно отметить присутствие продольных борозд, тогда как непосредственно по центру – спинномозговой канал. С внешней стороны размещена белая субстанция – множество отростков нервных клеток, которые объединены в волокна. Серое же вещество представлено преимущественно боковыми, а также задними с передними роговыми зонами – напоминает бабочку в полете.
Двигательные нервные клетки локализуются в передних рогах, а задние образования имеют вставочные нейроны, которые осуществляют взаимосвязь с чувствительными клетками. Они расположены в узловых сегментах нервной системы.
В месте соединения передних волокон с задними зонами формируются корешки спинного мозга. Они, по сути – проводники между центральной и периферической системой. Именно за счет подобного анатомического строения осуществляется взаимосвязь между разными участками человеческого организма, сохраняется равновесие внутренней среды – спинномозговыми рефлексами.
Именно за счет подобного анатомического строения осуществляется взаимосвязь между разными участками человеческого организма, сохраняется равновесие внутренней среды – спинномозговыми рефлексами.
Функции ЦНС
Исходя из особенностей строения и расположения структур центральной нервной системы, следуют ее основные функции:
- интегративная – установление взаимосвязи между клетками в тканях, органах в системах для образования единого высокоорганизованного человеческого организма;
- координирующая – обеспечение согласованности действий различных органов для достижения единой стоящей перед человеком задачи, к примеру, адаптация к резко изменившейся ситуации;
- регулирующая – структуры нервной системы контролируют все протекающие внутри процессы, без их участия не происходит ни одна деятельность;
- трофическая – выполняет регуляцию трофики, а также интенсивности обменных процессов в тканях, чтобы реакция на изменения была адекватной и быстрой;
- приспособительная – анализ и последующий синтез поступающей извне информации для приспособления к внешней среде.

Помимо всех перечисленных функций, значение ЦНС заключается в формировании ненаправленного поведения – в соответствии с доминирующей у человека потребности. По мере удовлетворения нижележащих – питание, дыхание, люди начинают искать свое внутреннее «Я», духовные ценности.
Несмотря на обилие научных работ о центральной нервной системе, в этой области таится много неизвестного. Человечество ожидает еще множество открытий.
Основные функции ЦНС организма — Студопедия
Нервная система обеспечивает жизнедеятельность организма как целого по отношению к внешней и внутренней среде. Основными функциями нервной системы являются:
Быстрая и точная передача информации о состоянии внешней и внутренней среды – сенсорная функция;
Анализ и интеграция всей информации;
Организация адаптивного реагирования на внешние сигналы – моторная функция;
Регуляция деятельности внутренних органов и внутренней среды – висцеральная функция;
Регуляция и координация деятельности всех органов и систем в соответствии с изменяющимися условиями внешней и внутренней среды.
Нервная система объединяет организм человека в единое целое, регулирует и координирует функции всех органов и систем, поддерживает постоянство внутренней среды организма (гомеостаз), устанавливает взаимоотношения организма с внешней средой.
Для нервной системы характерны точная направленность нервных импульсов, большая скорость проведения информации, быстрая приспособляемость к изменяющимся условиям внешней среды. У человека нервная система создает основу психической деятельности, анализа и синтеза поступающей в организм информации (мышления, речи, сложных форм социального поведения).
Эти сложнейшие и жизненно важные задачи решаются с помощью нейронов, выполняющих функцию восприятия, передачи, обработки и хранения информации. Сигналы (нервные импульсы) от органов и тканей человека и из внешней среды, воздействующей на поверхность тела и органы чувств, поступают по нервам в спинной и головной мозг. В мозге человека происходят сложные процессы обработки информации. В результате из мозга также по нервам к органам и тканям идут ответные сигналы, вызывающие реакцию организма, которая проявляется в виде мышечной или секреторной деятельности. В ответ на поступившие из мозга импульсы происходит сокращение скелетных мышц или мускулатуры в стенках внутренних органов, кровеносных сосудов, а также секреция различных желез — слюнных, желудочных, кишечных, потовых и других (выделение слюны, желудочного сока, желчи, гормонов железами внутренней секреции).
Сигналы (нервные импульсы) от органов и тканей человека и из внешней среды, воздействующей на поверхность тела и органы чувств, поступают по нервам в спинной и головной мозг. В мозге человека происходят сложные процессы обработки информации. В результате из мозга также по нервам к органам и тканям идут ответные сигналы, вызывающие реакцию организма, которая проявляется в виде мышечной или секреторной деятельности. В ответ на поступившие из мозга импульсы происходит сокращение скелетных мышц или мускулатуры в стенках внутренних органов, кровеносных сосудов, а также секреция различных желез — слюнных, желудочных, кишечных, потовых и других (выделение слюны, желудочного сока, желчи, гормонов железами внутренней секреции).
Из мозга к рабочим органам (мышцам, железам) нервные импульсы также следуют по цепям нейронов. Ответную реакцию организма на воздействия внешней среды или изменения его внутреннего состояния, выполняемую с участием нервной системы, называют рефлексом (от лат. reflexus — отражение, ответная реакция). Путь, состоящий из цепей нейронов, по которому нервный импульс проходит от чувствительных нервных клеток до рабочего органа, называют рефлекторной дугой. У каждой рефлекторной дуги можно выделить первый нейрон — чувствительный, или приносящий, который воспринимает воздействия, образует нервный импульс и приносит его в ЦНС. Следующие нейроны (один или несколько) являются вставочными, проводниковыми нейронами, расположенными в головном мозге. Вставочные нейроны проводят нервные импульсы от приносящего, чувствительного нейрона к последнему, выносящему, эфферентному нейрону. Последний нейрон выносит нервный импульс из мозга к рабочему органу (мышце, железе), включает этот орган в работу, вызывает эффект, поэтому называется также эффекторным нейроном.
Путь, состоящий из цепей нейронов, по которому нервный импульс проходит от чувствительных нервных клеток до рабочего органа, называют рефлекторной дугой. У каждой рефлекторной дуги можно выделить первый нейрон — чувствительный, или приносящий, который воспринимает воздействия, образует нервный импульс и приносит его в ЦНС. Следующие нейроны (один или несколько) являются вставочными, проводниковыми нейронами, расположенными в головном мозге. Вставочные нейроны проводят нервные импульсы от приносящего, чувствительного нейрона к последнему, выносящему, эфферентному нейрону. Последний нейрон выносит нервный импульс из мозга к рабочему органу (мышце, железе), включает этот орган в работу, вызывает эффект, поэтому называется также эффекторным нейроном.
Основными функциями ЦНС являются:
— объединение всех частей организма в единое целое и их регуляция;
— управление состоянием и поведением организма в соответствии с условиями внешней среды и его потребностями.
Главная и специфическая функция ЦНС — осуществление простых и сложных высокодифференцированных отражательных реакций, получивших название рефлексов.
У высших животных и человека низшие и средние отделы ЦНС — спинной мозг, продолговатый мозг, средний мозг, промежуточный мозг и мозжечок — регулируют деятельность отдельных органов и систем высокоразвитого организма, осуществляют связь и взаимодействие между ними, обеспечивают единство организма и целостность его деятельности.
Высший отдел ЦНС — кора больших полушарий головного мозга и ближайшие подкорковые образования — в основном регулирует связь и взаимоотношения организма как единого целого с окружающей средой.
Практически все отделы центральной и периферической нервной системы участвуют в переработке информации, поступающей через внешние и внутренние, расположенные на периферии тела и в самих органах рецепторы.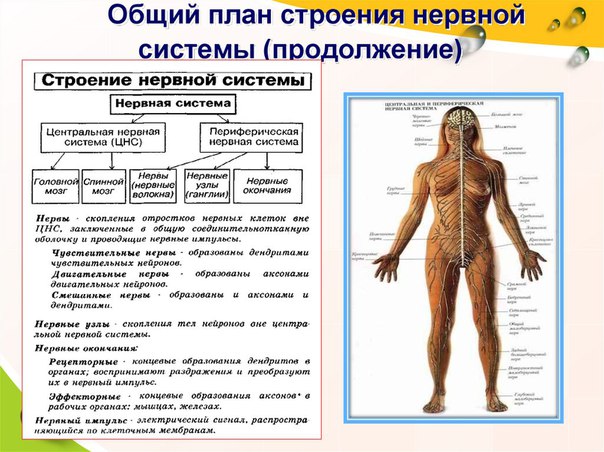 С высшими психическими функциями, с мышлением и сознанием человека связана работа коры головного мозги и подкорковых структур, входящих в передний мозг.
С высшими психическими функциями, с мышлением и сознанием человека связана работа коры головного мозги и подкорковых структур, входящих в передний мозг.
Основным принципом функционирования ЦНС является процесс регуляции, управления физиологическими функциями, которые направлены на поддержание постоянства свойств и состава внутренней среды организма. ЦНС обеспечивает оптимальные взаимоотношения организма с окружающей средой, устойчивость, целостность, оптимальный уровень жизнедеятельности организма.
Различают два основных вида регуляции: гуморальный и нервный.
Гуморальный процесс управления предусматривает изменение физиологической активности организма под влиянием химических веществ, которые доставляются жидкими средами организма. Источником передачи информации являются химические вещества – утилизоны, продукты метаболизма (углекислый газ, глюкоза, жирные кислоты), информоны, гормоны желез внутренней секреции, местные или тканевые гормоны.
Нервный процесс регуляции предусматривает управление изменениями физиологических функций по нервным волокнам при помощи потенциала возбуждения под влиянием передачи информации.
В организме нервный и гуморальный механизмы работают как единая система нейрогуморального управления. Это комбинированная форма, где одновременно используются два механизма управления, они взаимосвязаны и взаимообусловлены.
Нервная система представляет собой совокупность нервных клеток, или нейронов.
По локализации различают:
1) центральный отдел – головной и спинной мозг;
2) периферический – отростки нервных клеток головного и спинного мозга.
По функциональным особенностям различают:
1) соматический отдел, регулирующий двигательную активность;
2) вегетативный, регулирующий деятельность внутренних органов, желез внутренней секреции, сосудов, трофическую иннервацию мышц и самой ЦНС.
Функции нервной системы:
1) интегративно-коордиационная функция. Обеспечивает функции различных органов и физиологических систем, согласует их деятельность между собой;
2) обеспечение тесных связей организма человека с окружающей средой на биологическом и социальном уровнях;
3) регуляция уровня обменных процессов в различных органах и тканях, а также в самой себе;
4) обеспечение психической деятельности высшими отделами ЦНС.
Центральная нервная система: строение и функции
Организм человека работает как единое целое. Слаженность и взаимодействие всех органов обеспечивает центральная нервная система. Она имеется во всех живых существах и состоит из нервных клеток и их отростков.
ЦНС у позвоночных представлена головным и спинным мозгом, у беспозвоночных – системой объединенных нервных узлов.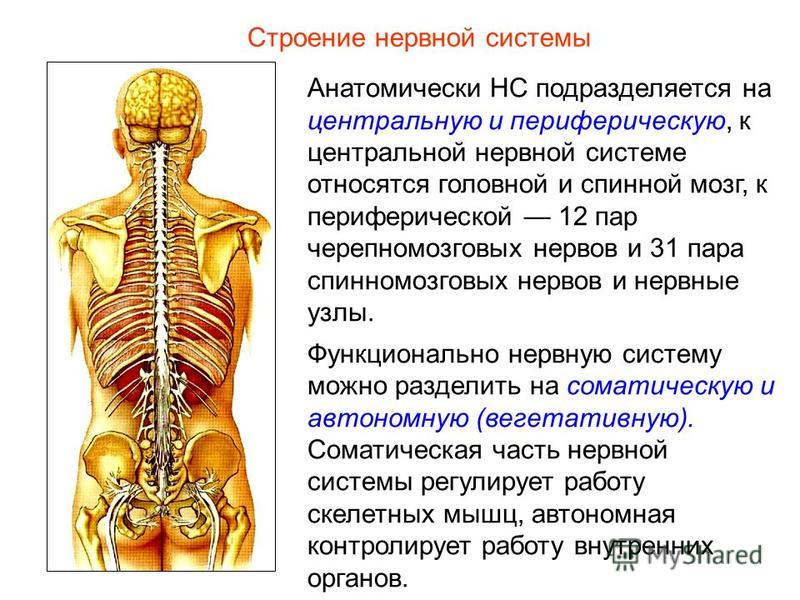 Центральную нервную систему защищают костные образования скелета: черепная коробка и позвоночник.
Центральную нервную систему защищают костные образования скелета: черепная коробка и позвоночник.
Строение центральной нервной системы
Анатомия центральной нервной системы изучает строение головного и спинного мозга, которые связаны с каждым органом посредством периферической НС.
ЦНС отвечает за такие чувства, как:
- слух;
- зрение;
- нюх;
- осязание;
- эмоции;
- память;
- мышление.
Структура мозга центральной нервной системы главным образом содержит белую и серую субстанции.
Серая – это нервные клетки с небольшими отростками. Располагаясь в спинном мозге, она занимает центральную часть, опоясывая спинномозговой канал. Что касается мозга головы, то в этом органе серое вещество составляет его кору и имеет отдельные образования в белом веществе. Белая субстанция располагается под серой. Ее структура содержит нервные волокна, формирующие нервные пучки. Некоторое количество таких «связок» составляют нерв.
Головной и спинной мозг окружают три оболочки:
- Твердая. Это наружная оболочка. Она располагается во внутренней полости черепной коробки и позвоночного канала.
- Паутинная. Этот покров находится под твердой частью. В своей структуре он имеет нервы и сосуды.
- Сосудистая. Данная оболочка напрямую соединена с мозгом. Она заходит в его борозды. Образована из множества кровеносных артерий. Паутинная от сосудистой оболочки отделяется полостью, которая заполнена мозговым веществом.
Спинной мозг как часть ЦНС
Эта составляющая ЦНС располагается в позвоночном канале. Он тянется от затылка до поясничного отдела. С двух сторон у мозга есть продольные борозды, а в центре – спинномозговой канал. С внешней стороны мозга спины размещается белая субстанция.
Серый элемент преимущественно составляют боковые, задние и передние роговые участки. Передние рога содержат двигательные нервные клетки, задние – имеют вставочные, производящие контакт чувствительных (лежащих в узловых отделах) и двигательных клеток. К передним роговым участкам двигательных частиц присоединены отростки, составляющие волокна. Те нейроны, которые создают задние корешки, присоединяются к задним роговым зонам.
К передним роговым участкам двигательных частиц присоединены отростки, составляющие волокна. Те нейроны, которые создают задние корешки, присоединяются к задним роговым зонам.
Эти корешки являются посредниками между нервной периферической системой и мозгом спины. Возбуждение, приходящее в мозг, поступает на вставочный нейрон, а затем через аксон поступает к нужному органу. Достигая отверстия между позвонками, чувствительные клетки соединяются с двигательными собратьями. После этого они разделяются на задние и передние веточки, состоящие также из двигательных и чувствительных волокон. От каждого позвонка в две стороны отходят 62 смешанных нерва.
Мозг головы человека
Этот орган расположился в мозговом отделе черепной коробки. Условно он имеет пять участков, внутри него имеются четыре полости, которые заполнены спинномозговой жидкостью. Большую часть органа составляют полушария (80%). Вторую по величине долю занимает ствол.
Он имеет такие структурные участки:
- средний;
- мозговой;
- продолговатый;
- промежуточный.

Участки головного мозга
- Продолговатый мозг. Эта область продолжает спинной мозг и имеет подобное ему строение. Его структура образована из белого вещества с областями серой субстанции, от которой отступают нервы черепа. Верхний участок заканчивается варолиевым мостом, а к бокам присоединяются от мозжечка нижние ножки. Практически весь этот мозг прикрывается полушариями. В сером элементе этой части мозга располагаются центры, отвечающие за функционирование легких, работу сердца, глотание, кашель, слезы, выделение слюны и образование желудочного сока. Какое-либо повреждение этого участка может прекратить дыхание и деятельность сердца, то есть привести к смерти.
- Задний мозг. К этой части относят мозжечок и варолиевый мост. Варолиев мост – это участок, начинающийся от продолговатого и заканчивающийся вверху «ножками». Боковые части его формируют средние ножки мозжечка. В варолиевый мост входят: лицевой, тройничный, отводящий и слуховой нервы. Мозжечок находится за мостом и продолговатым мозгом.
 Эта часть органа состоит из серого компонента, которым является кора, и белой субстанции с серыми участками. Мозжечок составляют: два полушария, средний участок и три пары ножек. Именно посредством этих ножек, которые состоят из нервных волокон, он и связан с другими областями головного мозга. Благодаря мозжечку человек может координировать свои движения, сохранять равновесие, держать мышцы в тонусе, выполнять четкие и плавные движения. Через проводящие пути центральной нервной системы мозжечок передает импульсы к мышечным тканям. Но его работа контролируется корой больших мозговых полушарий.
Эта часть органа состоит из серого компонента, которым является кора, и белой субстанции с серыми участками. Мозжечок составляют: два полушария, средний участок и три пары ножек. Именно посредством этих ножек, которые состоят из нервных волокон, он и связан с другими областями головного мозга. Благодаря мозжечку человек может координировать свои движения, сохранять равновесие, держать мышцы в тонусе, выполнять четкие и плавные движения. Через проводящие пути центральной нервной системы мозжечок передает импульсы к мышечным тканям. Но его работа контролируется корой больших мозговых полушарий. - Средний мозг. Анатомически размещается перед варолиевым мостом. Состоит из четырех холмиков и ножек мозга. По центру располагается канал, соединяющий третий и четвертый желудочки. Этот проток обрамляет серый элемент. В ножках мозга располагаются проводящие пути, объединяющие продолговатый и варолиев мост с полушариями. Благодаря среднему мозгу возможно поддержание тонуса и реализация рефлексов.
 Он позволяет выполнять такие действия, как стояние и хождение. Помимо этого в буграх четверохолмия размещены чувствительные ядра, имеющие связь со зрением и слухом. Они осуществляют световые и звуковые рефлексы.
Он позволяет выполнять такие действия, как стояние и хождение. Помимо этого в буграх четверохолмия размещены чувствительные ядра, имеющие связь со зрением и слухом. Они осуществляют световые и звуковые рефлексы. - Промежуточный. Он располагается перед мозговыми «ножками». Отделы этой части центральной нервной системы – это пара зрительных бугров, коленчатые тела, надбугорная и подбугорная области. Строение промежуточного мозга включает белое вещество и скопления серой субстанции. Здесь располагаются основные центры чувствительности – зрительные бугры. Именно сюда попадают импульсы со всего тела и далее направляются к коре мозга. Под буграми располагается гипоталамус, где вегетативная система показана подкорковым высшим центром. Благодаря ему происходит обмен веществ и теплоотдача. Этот центр поддерживает стабильность внутренней среды. В коленчатых телах расположены слуховые и зрительные нервы.
- Передний мозг. Его структура – это большие полушария с соединительной средней частью.
 Эти полушария разделены «проходом», внизу его расположилось мозолистое тело. Оно соединяет обе части нервными клеточными отростками. Верх полушарий – кора мозга, состоящая из нейронов и отростков. Под ней размещается белое вещество, выполняющее функцию проводящих путей. Оно объединяет центры полушария в одно целое. Эта субстанция состоит из нервных клеток, образующих подкорковые ядра серого элемента. Кора мозговых полушарий имеет довольно сложное строение. Она состоит из более чем 14 миллиардов нервных частиц, располагающихся шестью шарами. Они имеют разные формы, величины и связи.
Эти полушария разделены «проходом», внизу его расположилось мозолистое тело. Оно соединяет обе части нервными клеточными отростками. Верх полушарий – кора мозга, состоящая из нейронов и отростков. Под ней размещается белое вещество, выполняющее функцию проводящих путей. Оно объединяет центры полушария в одно целое. Эта субстанция состоит из нервных клеток, образующих подкорковые ядра серого элемента. Кора мозговых полушарий имеет довольно сложное строение. Она состоит из более чем 14 миллиардов нервных частиц, располагающихся шестью шарами. Они имеют разные формы, величины и связи.
Кора мозга головы обладает извилинами и бороздами.
Те, в свою очередь, разделяют поверхность на четыре участка:
- затылочный;
- лобный;
- теменной;
- висок.
Центральная и височная борозды относятся к самым глубоким. Первая проходит через полушария, вторая отделяет височный участок мозга от других. На участке лобной доли, перед центральной бороздой, располагается центральная передняя извилина.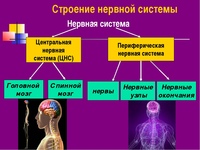 Задняя центральная извилина находится за основной бороздой.
Задняя центральная извилина находится за основной бороздой.
Основание мозга составляют нижняя зона полушарий и ствол. Каждому участку коры головного мозга отвечает своя часть тела. В этом сегменте располагаются центры практически всех чувствительных систем. Анализ поступающей информации проходит в коре мозговых полушарий. Главными участками коры являются: обонятельный, двигательный, чувствительный, слуховой, зрительный.
Строение ЦНС у высших и низших живых организмов имеет отличия. Система низших животных имеет структуру сетчатого типа, высшие организмы (в том числе и человек) имеют нейрогенный тип конструкции НС. В первом случае импульсы могут передаваться диффузно, во втором варианте каждая клетка функционирует как отдельная единица, хотя и связана с другими нейронами. Афферентная нервная система передает импульсы от всех органов к ЦНС.
Точки соединения этих частиц называют синапсами. Участок между клеткой и ее отростком заполнен глией. Это совокупность особых частиц, которые в отличие от нейронов способны делиться. Самый распространенный вид таких частиц – астроциты. Они чистят внеклеточный промежуток от лишних ионов и медиаторов, устраняют химические проблемы, препятствующие согласованным реакциям на поверхности нервных клеток. Помимо этого астроциты обеспечивают глюкозой активные клетки и меняют направление переноса кислорода.
Самый распространенный вид таких частиц – астроциты. Они чистят внеклеточный промежуток от лишних ионов и медиаторов, устраняют химические проблемы, препятствующие согласованным реакциям на поверхности нервных клеток. Помимо этого астроциты обеспечивают глюкозой активные клетки и меняют направление переноса кислорода.
В отделах центральной нервной системы происходит много нервных процессов. Простые и сложные высокодифференцированные отражательные реакции осуществляются благодаря этой системе. Функции центральной нервной системы можно охарактеризовать двумя назначениями: связь и взаимодействие живого организма и внешней среды и регуляция работы органов. Это является одним из необходимых условий для нормального функционирования организма.
5.5. Функции центральной нервной системы
Центральная
нервная система (ЦНС) в организме
выполняет интегрирующую роль. Она
объединяет в единое целое все ткани,
органы, координируя их специфическую
активность в составе целостных
гомеостатических и поведенческих
функциональных систем (П. К. Анохин).
Основными частными функциями ЦНС
являются следующие.
Она
объединяет в единое целое все ткани,
органы, координируя их специфическую
активность в составе целостных
гомеостатических и поведенческих
функциональных систем (П. К. Анохин).
Основными частными функциями ЦНС
являются следующие.
\
Управление деятельностью опорно-двигательного аппарата. ЦНС регулирует тонус мышц и посредством его перераспределения поддерживает естественную позу, а при нарушении восстанавливает ее, инициирует все виды двигательной активности (физическая работа, физкультура, спорт, любое перемещение организма).
Регуляция работы внутренних органов осуществляется вегетативной нервной системой и эндокринными железами; обеспечивает интенсивность их функционирования согласно потребностям организма в различных условиях его жизнедеятельности.
86
Обеспечение сознания и всех видов психической деятельности.
 Психическая
деятельность — это идеальная, субъективно
осознаваемая деятельность организма,
осуществляемая с помощью нейрофизиологических
процессов. И. П. Павлов ввел представление
о высшей и низшей нервной деятельности. Высшая нервная
деятельность — это
совокупность нейрофизиологических
процессов, обеспечивающих сознание,
подсознательную переработку
информации и целенаправленное поведение
организма в окружающей среде. Психическая
деятельность осуществляется с помощью
высшей нервной деятельности и протекает
осознанно, т.е. во время бодрствования,
независимо от того, сопровождается она
физической работой или нет. Высшая
нервная деятельность протекает
во время бодрствования и сна (см. разделы
15.8, 15.9, 15.10). Низшая
нервная деятельность — это совокупность
нейрофизиологических
процессов, обеспечивающих осуществление
безусловных рефлексов.
Психическая
деятельность — это идеальная, субъективно
осознаваемая деятельность организма,
осуществляемая с помощью нейрофизиологических
процессов. И. П. Павлов ввел представление
о высшей и низшей нервной деятельности. Высшая нервная
деятельность — это
совокупность нейрофизиологических
процессов, обеспечивающих сознание,
подсознательную переработку
информации и целенаправленное поведение
организма в окружающей среде. Психическая
деятельность осуществляется с помощью
высшей нервной деятельности и протекает
осознанно, т.е. во время бодрствования,
независимо от того, сопровождается она
физической работой или нет. Высшая
нервная деятельность протекает
во время бодрствования и сна (см. разделы
15.8, 15.9, 15.10). Низшая
нервная деятельность — это совокупность
нейрофизиологических
процессов, обеспечивающих осуществление
безусловных рефлексов.Формирование взаимодействия организма с окружающей средой.
 Это
реализуется, например, с помощью
избегания или избавления
от неприятных раздражителей (защитные
реакции организма),
регуляции интенсивности обмена веществ
при изменении температуры окружающей
среды. Изменения внутренней среды
организма, воспринимаемые субъективно
в виде ощущений, также побуждают
организм к той или иной целенаправленной
двигательной активности. Так,
например, в случае недостатка воды и
при повышении
осмотического давления жидкостей
организма возникает жажда, которая
инициирует поведение, направленное на
поиск и прием воды. Любая деятельность
самой ЦНС реализуется в конечном
итоге с помощью функционирования
отдельных клеток.
Это
реализуется, например, с помощью
избегания или избавления
от неприятных раздражителей (защитные
реакции организма),
регуляции интенсивности обмена веществ
при изменении температуры окружающей
среды. Изменения внутренней среды
организма, воспринимаемые субъективно
в виде ощущений, также побуждают
организм к той или иной целенаправленной
двигательной активности. Так,
например, в случае недостатка воды и
при повышении
осмотического давления жидкостей
организма возникает жажда, которая
инициирует поведение, направленное на
поиск и прием воды. Любая деятельность
самой ЦНС реализуется в конечном
итоге с помощью функционирования
отдельных клеток.
5.6. Функции клеток цнс и ликвора, классификация нейронов цнс, их медиаторы и рецепторы
Мозг
человека содержит около 50 миллиардов
нервных клеток, взаимодействие
между которыми осуществляется посредством
множества синапсов, число которых в
тысячи раз больше количества самих
клеток (1015-1016),
так как их аксоны делятся многократно
дихотомически, поэтому один
нейрон может образовывать
до тысячи синапсов с другими нейронами. Нейроны
оказывают свое
влияние на органы и ткани также посредством
синапсов.
Нейроны
оказывают свое
влияние на органы и ткани также посредством
синапсов.
А. Нервная клетка (нейрон) является структурной и функциональной единицей ЦНС, она состоит из сомы (тела клетки с яд-
87
ром) и отростков, представляющих собой большое число дендри-тов и один аксон (рис. 5.5). Потенциал покоя (ПП) нейрона составляет 60-80 мВ, потенциал действия (ПД) -80-110 мВ. Сома и дендриты покрыты нервными окончаниями — синаптическими бутонами и отростками глиальных клеток. На одном нейроне число синаптических бутонов может достигать 10 тысяч (см. рис. 5.5). Аксон начинается от тела клетки аксонным холмиком. Диаметр тела клетки составляет 10-100 мкм, аксона — 1-6 мкм, на периферии длина аксона может достигать метра и более. Нейроны мозга образуют колонки, ядра и слои, выполняющие определенные функции.
88
Клеточные
скопления образуют серое вещество
мозга. Между клетками
проходят немиелинизированные и
миелинизированные нервные волокна
(дендриты и аксоны нейронов).
Между клетками
проходят немиелинизированные и
миелинизированные нервные волокна
(дендриты и аксоны нейронов).
Функциями нервной клетки являются получение, переработка и хранение информации, передача сигнала другим нервным клеткам, регуляция деятельности эффекторных клеток различных органов и тканей организма. Целесообразно выделить следующие функциональные структуры нейрона.
Структуры, обеспечивающие синтез макромолекул, — это сома (тело нейрона), выполняющая трофическую функцию по отношению к отросткам (аксону и дендритам) и клеткам-эффекторам. Отросток, лишенный связи с телом нейрона, дегенерирует. Макромолекулы транспортируются по аксону и дендритам.
Структуры, воспринимающие импульсы от других нервных клеток, — это тело и дендриты нейрона с расположенными на них шипиками, занимающими до 40% поверхности сомы нейрона и ден-дритов. Причем, если шипики не получают импульсацию, они исчезают.
 Импульсы могут поступать и к окончанию
аксона — аксо-аксонные
синапсы, например, в случае пресинаптического
торможения.
Импульсы могут поступать и к окончанию
аксона — аксо-аксонные
синапсы, например, в случае пресинаптического
торможения.Структуры, где обычно возникает потенциал действия (генераторный пункт ПД), — аксонный холмик.
Структуры, проводящие возбуждение к другому нейрону или к эффектору, — аксон.
Структуры, передающие импульсы на другие клетки, — синапсы.
Б. Классификация нейронов ЦНС. Нейроны делят на следующие основные группы.
В зависимости от отдела ЦНС выделяют нейроны соматической и вегетативной нервной системы.
По источнику или направлению информации нейроны подразделяют на: а) афферентные, воспринимающие с помощью рецепторов информацию о внешней и внутренней среде организма и передающие ее в вышележащие отделы ЦНС; б) эфферентные, передающие информацию к рабочим органам — эффекторам; нервные клетки, иннервирующие эффекторы, иногда называют эф-фекторными; эффекторные нейроны спинного мозга (мотонейроны) делят на а- иу-мотонейроны; в) вставочные (интернейроны), обеспечивающие взаимодействие между нейронами ЦНС.

По медиатору, выделяющемуся в окончаниях аксонов, различают нейроны адренергические, холинергические, серотонинер-гические и т. д.
По влиянию — возбуждающие и тормозящие.
89
В. Глиальные
клетки (нейроглия
— «нервный клей») более многочисленны,
чем нейроны, составляют около 50% объема
ЦНС. Они способны к делению в течение
всей жизни. Размеры глиальных
клеток в 3—4 раза меньше нервных, с
возрастом их число увеличивается
(число нейронов уменьшается). Тела
нейронов, как и их аксоны, окружены
глиальными клетками. Глиальные клетки выполняют
несколько функций: опорную,
защитную, изолирующую, обменную
(снабжение нейронов питательными
веществами). Микроглиальные клетки
способны к фагоцитозу, ритмическому
изменению своего объема (период
«сокращения» -1,5 мин, «расслабления» —
4 мин). Циклы изменения объема повторяются
через каждые 2-20 час. Полагают, что
пульсация способствует продвижению
аксоплазмы в нейронах и влияет на ток
межклеточной жидкости. Мембранный
потенциал клеток нейроглии составляет
70-90 мВ, однако ПД они не генерируют,
возникают только лишь локальные токи,
электротонически распространяющиеся
от одной клетки к другой. Процессы
возбуждения в нейронах и электрические
явления в глиальных клетках, по-видимому,
взаимодействуют.»
Полагают, что
пульсация способствует продвижению
аксоплазмы в нейронах и влияет на ток
межклеточной жидкости. Мембранный
потенциал клеток нейроглии составляет
70-90 мВ, однако ПД они не генерируют,
возникают только лишь локальные токи,
электротонически распространяющиеся
от одной клетки к другой. Процессы
возбуждения в нейронах и электрические
явления в глиальных клетках, по-видимому,
взаимодействуют.»
Г. Ликвор — бесцветная
прозрачная жидкость, заполняющая
мозговые
желудочки, Спинномозговой канал и
субарахноидальное пространство. Ее
происхождение связано с интерстициальной
жидкостью
мозга, значительная часть ликвора
образуется сосудистыми сплетениями
желудочков мозга. Непосредственной питательной средой
клеток мозга является интерстициальная
жидкость, в которую клетки выделяют
также и продукты своего обмена. Лик-вор
представляет собой совокупность
фильтрата плазмы крови и интерстициальной
жидкости: она содержит около 90% воды и
около
10% сухого остатка (2% — органические, 8% —
неорганические вещества).
Д. Медиаторы и рецепторы синапсов ЦНС. Медиаторами синапсов ЦНС являются многие химические вещества, разнородные в структурном отношении (в головном мозге к настоящему времени обнаружено около 30 биологически активных веществ). Вещество, из которого синтезируется медиатор (предшественник медиатора), попадает в нейрон или его окончание из крови или ликвора, в результате биохимических реакций под действием ферментов в нервных окончаниях превращается в соответствующий медиатор и накапливается в синаптических везикулах. По химическому строению медиаторы можно разделить на несколько групп, главными из которых являются амины, аминокислоты, полипептиды. Достаточно широко распространенным медиатором является ацетилхолин.
Согласно
принципу Дейла, один
нейрон синтезирует и использует
один и тот же медиатор или одни и те же
медиаторы во всех разветвлениях своего
аксона («один
нейрон — один медиатор»).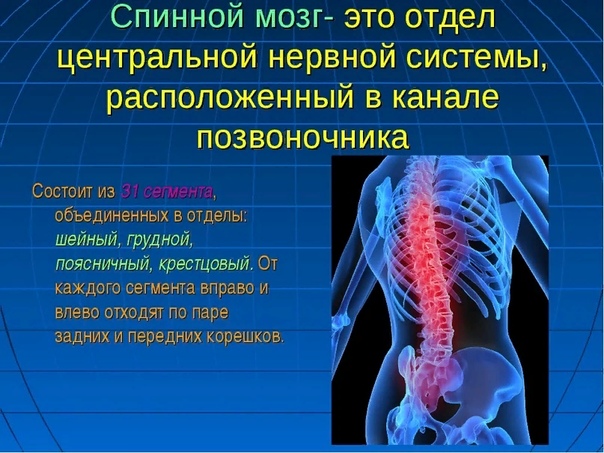 Кроме основного медиатора, как выяснилось,
в окончаниях аксона
могут выделяться и другие — сопутствующие
медиаторы (ко-медиаторы), играющие
модулирующую роль и более медленно
действующие.
Однако в спинном мозге установлено два
быстродействующих
медиатора в одном тормозном нейроне —
ГАМК и глицин и даже один тормозной
(ГАМК) и один возбуждающий (АТФ). Поэтому
принцип Дейла в новой редакции сначала
звучал: «Один нейрон — один быстрый
медиатор», а затем: «Один нейрон — один
быстрый синаптический эффект»
(предполагаются и другие варианты).
Кроме основного медиатора, как выяснилось,
в окончаниях аксона
могут выделяться и другие — сопутствующие
медиаторы (ко-медиаторы), играющие
модулирующую роль и более медленно
действующие.
Однако в спинном мозге установлено два
быстродействующих
медиатора в одном тормозном нейроне —
ГАМК и глицин и даже один тормозной
(ГАМК) и один возбуждающий (АТФ). Поэтому
принцип Дейла в новой редакции сначала
звучал: «Один нейрон — один быстрый
медиатор», а затем: «Один нейрон — один
быстрый синаптический эффект»
(предполагаются и другие варианты).
Эффект
действия медиатора
зависит в основном от свойств
постсинаптической
мембраны и вторых посредников. Это
явление особенно
ярко демонстрируется при сравнении
эффектов отдельных медиаторов
в ЦНС и в периферических синапсах
организма. Ацетилхолин,
например, в коре мозга при микроаппликациях
на разные нейроны
может вызывать возбуждение и торможение,
в синапсах сердца
— торможение, в синапсах гладкой
мускулатуры желудочно-кишечного
тракта — возбуждение.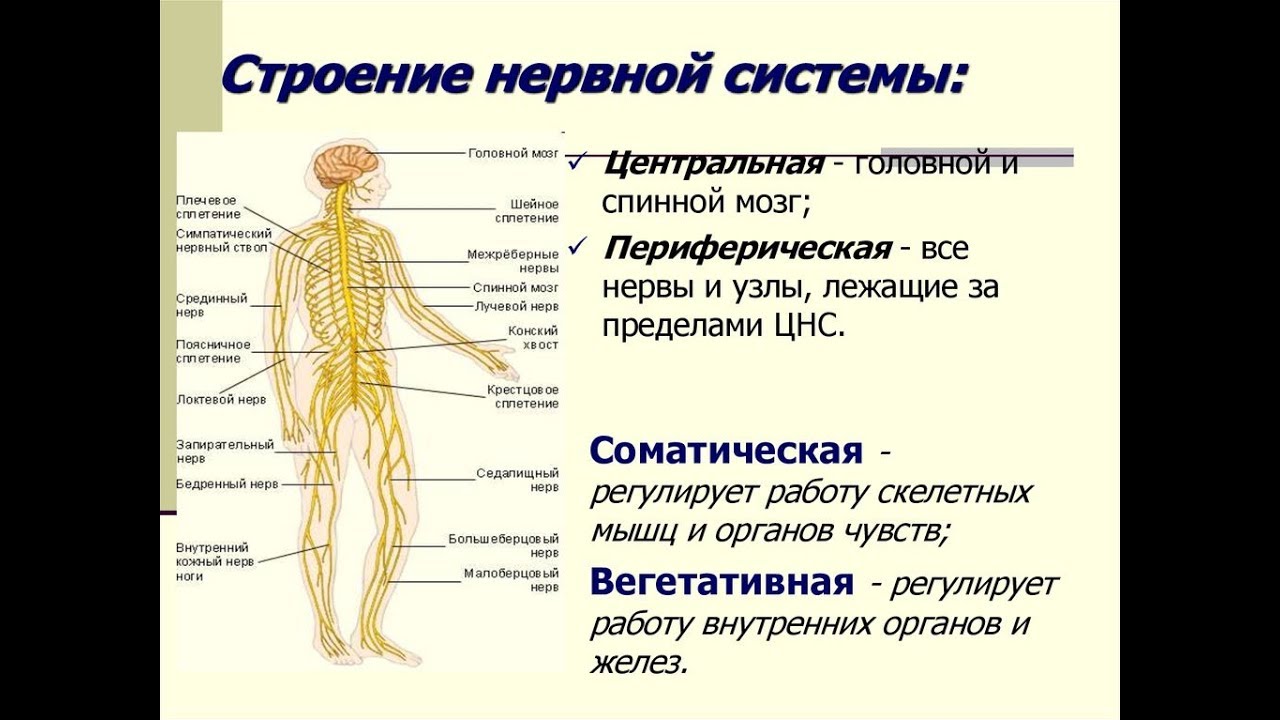 Катехоламины
стимулируют сердечную
деятельность, но тормозят сокращения
желудка и кишечника.
Катехоламины
стимулируют сердечную
деятельность, но тормозят сокращения
желудка и кишечника.
5.7. МЕХАНИЗМ ВОЗБУЖДЕНИЯ НЕЙРОНОВ ЦНС
В любых химических синапсах (ЦНС, вегетативных ганглиях, в нервно-мышечном) механизмы передачи сигнала в общих чертах подобны (см. раздел 2.1). Однако в возбуждении нейронов ЦНС имеются характерные особенности, основными из которых являются следующие.
Для возбуждения нейрона (возникновения ПД) необходимы поток афферентных импульсов и их взаимодействие. Это объясняется тем, что один пришедший к нейрону импульс вызывает небольшой возбуждающий постсинаптический потенциал (ВПСП, рис. 5.6) — всего 0,05 мВ (миниатюрный ВПСП). Один пузырек содержит до нескольких десятков тысяч молекул медиатора, например ацетилхолина. Если учесть, что пороговый потенциал нейрона 5-10 мВ, ясно, что для возбуждения нейрона требуется множество импульсов.
Место возникновения генераторных ВПСП, вызывающих ПД нейрона.
 Подавляющее
большинство нейрональных синапсов
находится на дендритах нейрона. Однако
наиболее эффективно вызывают
возбуждение нейрона синаптические
контакты,
Подавляющее
большинство нейрональных синапсов
находится на дендритах нейрона. Однако
наиболее эффективно вызывают
возбуждение нейрона синаптические
контакты,
90
91
расположенные на теле нейрона. Это связано с тем, что постси-наптические мембраны этих синапсов располагаются в непосредственной близости от места первичного возникновения ПД, располагающегося в аксонном холмике. Близость соматических синапсов к аксонному холмику обеспечивает участие их ВПСП в механизмах генерации ПД. В этой связи некоторые авторы предлагают называть их генераторными синапсами.
Генераторный пункт нейрона, т.е. место возникновения ПД, — аксонный холмик. Синапсьг на нем отсутствуют, отличительной особенностью мембраны аксонного холмика является вы-» сокая возбудимость, в 3-4 раза превосходящая возбудимость сома-дендритной мембраны нейрона, что объясняется более высокой концентрацией Ыа-каналов на аксонном холмике. ВПСП элек-тротонически достигают аксонный холмик, обеспечивая здесь уменьшение мембранного потенциала до критического уровня. В этот момент возникает ПД. Возникший в аксонном холмике ПД, с одной стороны, ортодромно переходит на аксон, с другой — антидромно на тело нейрона.
Роль дендритов в возникновении возбуждения до сих пор дискутируется. Полагают, что множество ВПСП, возникающих на дендритах, электротонически управляют возбудимостью нейрона. В этой связи дендритные синапсы получили название модуляторных синапсов.
5.8. ХАРАКТЕРИСТИКА РАСПРОСТРАНЕНИЯ ВОЗБУЖДЕНИЯ В ЦНС
Особенности распространения возбуждения в ЦНС объясняются ее нейронным строением — наличием химических синапсов, многократным ветвлением аксонов нейронов, наличием замкнутых нейронных путей. Этими особенностями являются следующие.
92
Одностороннее распространение возбуждения в нейронных цепях, в рефлекторных дугах. Одностороннее распространение возбуждения от аксона одного нейрона к телу или дендритам другого нейрона, но не обратно, объясняется свойствами химических синапсов, которые проводят возбуждение только в одном направлении.
Замедленное распространение возбуждения в ЦНС по сравнению с нервным волокном объясняется наличием на путях распространения возбуждения множества химических синапсов. Суммарная задержка передачи возбуждения в нейроне до возникновения ПД достигает величины порядка 2 мс.
Иррадиация (дивергенция) возбуждения в ЦНС объясняется ветвлением аксонов нейронов, их способностью устанавливать многочисленные связи с другими нейронами, наличием вставочных нейронов, аксоны которых также ветвятся (рис. 5.7 — А).
Конвергенция возбуждения (принцип общего конечного пути) — схождение возбуждения различного происхождения по нескольким путям к одному и тому же нейрону или нейронному пулу (принцип шеррингтоновской воронки). Объясняется наличием многих аксонных коллатералей, вставочных нейронов, а также тем, что афферентных путей в несколько раз больше, чем эфферентных нейронов. На одном нейроне ЦНС могут располагаться до 10 000 синапсов, на мотонейронах — до 20 000 (рис. 5.7 — Б).
Циркуляция возбуждения по замкнутым нейронным цепям, которая может продолжаться минутами и даже часами (рис. 5.8).
93
6. Распространение возбуждения в центральной нервной системе легко блокируется фармакологическими препаратами, что находит широкое применение в клинической практике. В физиологических условиях ограничения распространения возбуждения по ЦНС связаны с включением нейрофизиологических механизмов торможения нейронов.
Рассмотренные особенности распространения возбуждения дают возможность подойти к пониманию отличительных свойств нервных центров.
Центральная нервная система — Википедия
Центра́льная не́рвная систе́ма (ЦНС) — основная часть нервной системы и животных и людей, состоящая из нейронов, их отростков и вспомогательной глии; у беспозвоночных представлена системой тесно связанных между собой нервных узлов (ганглиев), у позвоночных животных (включая человека) — спинным и головным мозгом.
Главная и специфическая функция ЦНС — осуществление простых и сложных рефлексов. У человека и других высших животных низшие и средние отделы ЦНС — спинной мозг, продолговатый мозг, средний мозг, промежуточный мозг и мозжечок — регулируют деятельность отдельных органов и систем высокоразвитого организма, осуществляют связь и взаимодействие между ними, обеспечивают единство организма и целостность его деятельности. Высший отдел ЦНС — кора больших полушарий головного мозга и ближайшие подкорковые образования — в основном регулирует связь и взаимоотношения организма как единого целого с окружающей средой.
Строение и функции
Центральная нервная система связана со всеми органами и тканями организма через периферическую нервную систему, включающую у позвоночных черепные нервы, от головного мозга, спинномозговые — от спинного мозга, нервные узлы, находящиеся между позвонков; периферические отделы. Вегетативная нервная система — нервные узлы с нервными волокнами, подходящих и отходящих к ним. В состав ЦНС входят клетки нейроглии, которые выполняют в ней опорную и защитную функцию, участвуют в метаболизме нервных клеток. Головной и спинной мозг окружены тремя мозговыми оболочками: твёрдой, паутинной и сосудистой. Головной мозг заключён в защитную капсулу — череп, а спинной — в позвоночник.
Чувствительные (афферентные) нервы несут возбуждение в ЦНС от периферических рецепторов; по отводящим эфферентным (двигательным и вегетативным) нервным волокнам возбуждение из ЦНС направляется к клеткам исполнительных рабочих аппаратов (мышцы, железы, сосуды и др.). Афферентные и эфферентные клетки своими отростками могут контактировать между собой и составлять двухнейронную рефлекторную дугу, осуществляющую элементарные рефлексы (например сухожильные рефлексы спинного мозга). Но, как правило, в рефлекторной дуге между афферентными и эфферентными нейронами расположены вставочные нейроны. Связь между различными отделами ЦНС осуществляется также с помощью множества отростков афферентных, эфферентных и вставочных нейронов этих отделов, образующих внутрицентральные короткие и длинные проводящие пути.
Твёрдая — наружная, соединительнотканная, выстилает внутреннюю полость черепа и позвоночного канала. Паутинная расположена под твёрдой — это тонкая оболочка с небольшим количеством нервов и сосудов. Сосудистая оболочка сращена с мозгом, заходит в борозды и содержит много кровеносных сосудов.
Спинной мозг находится в позвоночном канале и имеет вид белого тяжа. По передней и задней поверхности спинного мозга расположены продольные борозды. В центре проходит спинно-мозговой канал, вокруг него сосредоточено серое вещество — скопление огромного количества нервных клеток, образующих контур бабочки.
Белое вещество спинного мозга образует проводящие пути, которые тянутся вдоль спинного мозга, соединяя как отдельные его сегменты друг с другом, так и спинной мозг с головным. Одни проводящие пути называются восходящими или чувствительными, передающими возбуждение в головной мозг, другие — нисходящими или двигательными, которые проводят импульсы от головного мозга к определённым сегментам спинного мозга. Они выполняют две функции — рефлекторную и проводниковую. Деятельность спинного мозга находится под контролем головного мозга, который регулирует спинномозговые рефлексы.
Головной мозг человека расположен в мозговом отделе черепа. Средняя его масса 1300—1400 г. Рост мозга продолжается до 20 лет. Состоит он из 5-ти отделов: переднего, промежуточного, среднего, заднего и продолговатого мозга. Внутри головного мозга находятся 4 сообщающиеся между собой полости — мозговые желудочки. Они заполнены спинномозговой жидкостью. Филогенетически более древняя часть — ствол головного мозга. Ствол включает продолговатый мозг, варолиев мост, средний и промежуточный мозг. 12 пар черепных нервов лежат в стволе мозга. Стволовая часть мозга прикрыта полушариями головного мозга.
Продолговатый мозг — продолжение спинного мозга и повторяет его строение; на передней и задней поверхности залегают борозды. Он состоит из белого вещества, где рассеяны скопления серого вещества — ядра, от которых берут начало черепные нервы — с 9 по 12-ю пару.
Задний мозг включает варолиев мост и мозжечок. Варолиев мост снизу ограничен продолговатым мозгом, сверху переходит в ножки мозга, боковые его отделы образуют средние ножки мозжечка. Мозжечок расположен сзади моста и продолговатого мозга. Поверхность его состоит из серого вещества (кора). Под корой — ядра.
Средний мозг расположен впереди варолиева моста, он представлен четверохолмием и ножками мозга. Промежуточный мозг занимает самое высокое положение и лежит спереди ножек мозга. Состоит из зрительных бугров, надбугорной, подбугорной области и коленчатых тел. По периферии промежуточного мозга находится белое вещество. Передний мозг состоит из сильно развитых полушарий и соединяющей их срединной части. Борозды делят поверхность полушарий на доли; в каждом полушарии различают 4 доли: лобную, теменную, височную и затылочную.
Деятельность анализаторов отражает в нашем сознании внешний материальный мир. Деятельность коры головного мозга человека и высших животных определена И. П. Павловым как высшая нервная деятельность, представляющая собой условно-рефлекторную функцию коры головного мозга.
Ссылки
Центральная нервная система (ЦНС)
Центральная нервная система (ЦНС) состоит из головного мозга, спинного мозга и сетчатки. Это командный центр тела, приводящий в действие все, что мы делаем, и интерпретирующий все вокруг нас.
Эта система продолжает развиваться на протяжении всей жизни человека, постоянно создавая и удаляя нейроны, чтобы поддерживать нормальное функционирование нервной системы в норме.
Центральная нервная система работает, принимая информацию, собранную периферической нервной системой , и перераспределяя информацию на основе этого.Он контролирует все, от чихания до частоты сердечных сокращений.
ЦНС содержит около 86 миллиардов нервов. в общей сложности это более чем в 12 раз больше, чем на планете Земля. прямо сейчас! Все они связаны триллионами нервных волокон — аксонов и дендриты — которые способствуют передаче информации по всему вся нервная система.
Мозг
У людей самый большой мозг среди всех позвоночных животных относительно размера тела (Lewis, 2018).Однако, несмотря на то, что вы, возможно, слышали, размер на самом деле не имеет значения.
Многие уже довольно давно считали, что размер мозга прямо коррелирует с интеллектом у видов животных, однако научных доказательств, подтверждающих эту идею, не так много. Точнее сказать, что соотношение размера мозга и размера тела является более надежным показателем интеллекта.
Помимо самого большого мозга по сравнению с размером тела, у людей также больше нейронов на единицу объема, чем у нескольких других видов .Основная причина этого — площадь поверхности человеческого мозга. Поверхность головного мозга образована всеми складками ткани. Чем больше складок, тем больше площадь поверхности.
Человеческий мозг весит около 3,5 фунтов и составляет примерно 2% от средней массы тела взрослого человека и контролирует почти все, что мы делаем. «Подожди, почти?» — недоверчиво спросите вы . Да, почти… вроде.
Мозг не контролирует такие движения напрямую, как рефлексы — эти действия проходят через спинной мозг , самое большее из-за скорости, с которой они должны происходить.Но вот из чего состоит мозг и что он контролирует.
Структуры и функции человеческого мозга
Вся ЦНС состоит из «серого вещества», и «белого вещества». Серое вещество состоит из нейронов, типа клеток, которые составляют основу всех типов нервных клеток.
Самая большая часть человеческого мозга — это головной мозг, который происходит от переднего мозга или переднего мозга. Головной мозг разделен на два полушария, которые соединены мозолистым телом .Эта область мозга управляет сенсорными, моторными и когнитивными функциями мозга.
Левое полушарие головного мозга содержит важные области, которые играют важную роль в речи и языке. Это также связано с математикой и поиском информации.
Правое полушарие мозга играет ключевую роль в обработке зрения и слуха, пространственных навыках и художественных способностях. Но , все эти функции задействованы в обоих полушариях мозга, поэтому популярное понятие «левое или правое полушарие» плохо подтверждается доказательствами.Извините, что убил атмосферу.
Наружный слой головного мозга, известный поскольку кора головного мозга считается «центром мысли» и состоит следующих структур:
- Лобная доля: контролирует эмоции, решение проблем, обучение, память и многое другое.
- Теменная доля: управляет пространственной ориентацией и навигацией путем интеграции информации, собранной многочисленными сенсорными системами.
- Височная доля: обрабатывает слуховые стимулы и помогает в речи; включает гиппокамп и миндалевидное тело, которые играют основные роли в памяти и эмоциях, соответственно.
- Затылочная доля: регулирует обработку изображений
Corpus callosum .Да, это определенно звучит как заклинание, которое вы выучили в Хогвартсе, но, поверьте мне, это не так (я пробовала). Это совокупность нервных волокон, которые соединяют левое и правое полушария мозга.
Как ни странно, левое полушарие управляет всеми мышцами правой стороны тела и наоборот. Левосторонность или праворукость — это результат того, что одно полушарие доминирует над другим.
Ствол мозга происходит от среднего мозга (mesencephalon) и заднего мозга (rombencephalon) и является связующим звеном между спинным мозгом и мозгом.Он состоит из продолговатого мозга, моста и среднего мозга. Основные обязанности ствола мозга —
- Передача информации между мозгом. и тело
- Поставляют черепные нервы к лицо и голова (нервы идут от этой области мозга к этим областям)
- Выполняют важные функции в контроль сердца, дыхания и сознания.
Между головным мозгом и стволом головного мозга находятся таламус и гипоталамус. Гипоталамус передает сенсорные и двигательные сигналы к коре и участвует в регулировании сознания, сна и бдительности. Гипоталамус соединяет нервную систему с эндокринной системой через гипофиз
Мозжечок, который развивается из задний мозг важен для управления моторикой, поскольку он частично регулирует координацию и баланс. Эта структура также может выполнять некоторые когнитивные функции.
Защита и основа центральной нервной системы
Органы нервной системы очень хорошо защищены и надежно поддерживаются тканями и структурами органов по всему телу.
Как и должно быть! Вы же не хотите, чтобы ваши нервные импульсы вертелись вокруг каждый раз, когда вы делаете движение, не так ли? Конечно, нет!
Чтобы получить максимальную защиту, частям CNS предоставляется до трех различных форм защиты . И головной, и спинной мозг сначала защищены слоями мембраны, называемыми мозговыми оболочками. Мозг дополнительно инкапсулирован черепом, а спинной мозг — позвонками.
В-третьих, это спинномозговая жидкость: это невероятно важно, поскольку она обеспечивает буфер, который снижает силу любого потенциального удара между мозгом и черепом или между спинным мозгом и позвонками.
Мозг дополнительно инкапсулирован черепом, а спинной мозг — позвонками. В-третьих, существует спинномозговая жидкость : это невероятно важно, так как обеспечивает буфер, который снижает силу любого потенциального удара между мозгом и черепом или между спинным мозгом и позвонками.
Уже упоминалось, что CNS полностью состоит из двух основных категорий нервных клеток: серого вещества и белое вещество. Серое вещество состоит из тел нейронов и их ассоциированные дендриты, глиальные клетки, капилляры (они обеспечивают кровоснабжение через ткань головного мозга, что и делает его розовым!).
Серое вещество находится в основном во внешних слоях головного мозга и спинном мозге. Вы можете идентифицировать это в спинном мозге, поскольку это то, что образует внутреннюю форму «бабочки», которую вы можете увидеть в поперечном сечении спинного мозга.
Белое вещество, с другой стороны, составляет области ЦНС, в которых находится большинство аксонов. Белое вещество внизу серое вещество в головном мозге, а в спинном мозге это внешний слой вокруг серого ядра.
Спинной мозг
Спинной мозг весит всего около 35 г и имеет всего около 1 см в диаметре.Он защищен 26 позвонками и может быть разделен на пять областей, всего 31 пара нервов простирается от боковых сторон позвоночного столба к дистальным частям тела. Пять отделов спинного мозга следующие:
- Шейный (шея): 8 пар нервов
- Грудной (грудная клетка): 12 пар нервов
- Поясничный (брюшной): 5 пар нервов нервы
- Крестцовый (тазовый): 1 пара нервов
- Копчиковый (копчиковый): 1 пара нервов
Серое вещество в спинном мозге составляет маслянистый силуэт.Каждое «крыло» обозначается как «рожок ». ”
Эти рога представляют собой скопление сенсорных нейронов, и все они окружены белым веществом, функционирующим так же, как миелиновая оболочка аксона. (Таким образом, вы можете думать о спинном мозге как о Матери всех нейронов, где каждый нерв периферической нервной системы действует как дендриты, спинной мозг — как аксон, а мозг — как ядро.)
Сетчатка
сетчатка — это нервная часть глаза .Это происходит из-за «загибания» промежуточного мозга, называемого зрительным пузырьком, который подвергается инвагинации, образуя глазной бокал.
В частности, внутренняя стенка этого глазного бокала дает начало сетчатке, а внешняя стенка дает начало пигментному эпителию. Пигментный эпителий — это то, что делает ваш глаз голубым, коричневым и т. Д., Поскольку он содержит меланин.
Меланин нужен не только для красоты. Он работает, чтобы уменьшить обратное рассеяние света, попадающего в глаз, делая его легче (и возможно) увидеть.
Пигментный эпителий также играет важную роль в поддержании фоторецепторов, обновляя фотопигменты и фагоцитируя диски фоторецепторов. Это делается с невероятно высокой скоростью, необходимой для поддержания зрения.
Фоторецепторы посылают электрические импульсы на сетчатка, нейроны которой затем отправляют эту информацию в мозг через зрительный нерв.
В сетчатке есть пять типов нейронов:
- Фоторецепторы
- Биполярные клетки
- Ганглиозные клетки
- Горизонтальные клетки
- Амакриновые клетки
светочувствительных нервных клеток в сетчатке — палочки и колбочки.Оба имеют внешний сегмент, который состоит из мембранных дисков, содержащих фотопигмент, что позволяет воспринимать слуховые стимулы, специфичные для цвета.
Все эти структуры, возглавляемые головным, спинным мозгом и сетчаткой и связанные триллионами синапсов, регулируют основные процессы, необходимые нашему телу для поддержания жизни и дыхания.
Ссылки
Purves D, Augustine GJ, Fitzpatrick D, et al., Редакторы. Неврология. 2-е издание. Сандерленд (Массачусетс): Sinauer Associates; 2001 г.Сетчатка. Доступно по адресу: https://www.ncbi.nlm.nih.gov/books/NBK10885/
Frontiers | Роль ТРЕК-1 в здоровье и болезнях, фокус на центральной нервной системе
Введение
В головном мозге ионные каналы регулируют множество клеточных процессов, таких как высвобождение нейротрансмиттеров, возбудимость нейронов и пластичность. Они имеют решающее значение для генерации и распространения потенциалов действия. Нарушение функции этих каналов вызывает несколько патологий, называемых каннелопатиями.Для лечения этих каннелопатий многие ионные каналы были мишенями для небольших молекул и биологических препаратов. В целом ионные каналы составляют 19% всех белковых мишеней человека (Santos et al., 2017). На сегодняшний день 177 препаратов на основе ионных каналов одобрены для лечения различных патологий и расстройств (Santos et al., 2017). Патологии центральной нервной системы (ЦНС) были связаны с нарушением регуляции ионных каналов, таких как потенциалзависимые каналы Na + , K + и Ca 2+ .Кальциевые каналы являются наиболее изученными среди ионных каналов, поскольку ионы Ca 2+ помимо контроля возбудимости клетки действуют как вторичные посредники, преобразующие электрические сигналы в химическую активность (Pietrobon, 2002). С другой стороны, чтобы контролировать возбудимость нейронов, калиевые каналы реполяризуют нейроны, вытесняя ионы K + во внеклеточное пространство. Неврологические калиевые каналы в основном вызваны нарушением регуляции потенциал-зависимых калиевых каналов (Benatar, 2000), а также внутренне выпрямляющих калиевых каналов (Kir), что вызывает гипервозбудимость, наблюдаемую при эпилепсии (Spillane et al., 2016). Мутации в гене KCNT1, кодирующем активируемые натрием калиевые каналы, вызывают редкую инфантильную энцефалопатию, называемую мигрирующими парциальными припадками младенчества (Barcia et al., 2012). В других случаях мутации KCNQ2, кодирующего канал Kv7.2, провоцируют тяжелые эпилептические энцефалопатии (Weckhuysen et al., 2012). Калиевый канал, выпрямляющий внутрь, Kir 6.2 кодируется геном KCNJ11, который образует главную субъединицу АТФ-чувствительного калиевого канала. Мутации в этом гене вызывают синдром DEND, очень редкую тяжелую форму неонатального сахарного диабета, характеризующуюся задержкой развития, эпилепсией и неонатальным диабетом (Gloyn et al., 2004). Kv7.2 и Kv7.3 являются основными компонентами медленного потенциал-управляемого М-канала, который регулирует возбудимость нейронов (Brown and Passmore, 2009). Мутации с потерей функции генов KCNQ2 и KCNQ3, кодирующих, соответственно, Kv7.2 и Kv7.3, вызывают доброкачественные семейные неонатальные судороги (Spillane et al., 2016). Доминантная миссенс-мутация в гене KCNMA1, кодирующем порообразующую α-субъединицу большого проводящего кальций-чувствительного калиевого канала (ВК), была связана с формой генерализованной эпилепсии и пароксизмальной дискинезии (Du et al., 2005).
Сообщалось, что некоторые мутации в каналах K 2P вызывают несколько патологий. Синдром дисморфизма умственной отсталости Бирка Барела вызван миссенс-мутацией в материнской копии гена KCNK9, который кодирует TASK-3 (K 2P 9.1). Потеря функции канала наблюдалась в гомодимерах, а также когда он образует гетеродимер с TASK-1 (K 2P .3.1) (Barel et al., 2008). Недавно сообщалось, что de novo усиление функциональной миссенс-мутации гена KCNK4, кодирующего канал TRAAK, вызывает распознаваемый синдром неврологического развития, характеризующийся лицевым дисморфизмом, гипертрихозом, эпилепсией, умственной отсталостью / задержкой развития и чрезмерным ростом десен (Bauer et al., 2018). Мутация сдвига рамки считывания (F139Wfsx24) в KCNK18, кодирующая активированный кальцием K 2P канал TRESK канала, была связана с мигренью с аурой (Lafreniere et al., 2010). Каннелопатии, вызванные дисфункцией калиевых каналов, представляют большой интерес для исследователей, поскольку могут представлять интересную мишень для потенциальных методов лечения.
Последнее открытое семейство двухпоровых калиевых каналов (K 2P ) регулируется множеством химических и физических стимулов (Lesage and Lazdunski, 2000).В этом обзоре мы сосредоточимся на описании роли одного из наиболее изученных каналов K 2P , канала TREK-1, в здоровье и болезнях. Помимо обсуждения последних фармакологических модуляторов его активности.
Трек-1 канал
TREK-1, названный KCNK2 или K 2P 2.1, принадлежит к большому семейству каналов K 2P , содержащему 15 членов, сгруппированных в шесть подсемейств. Каналы K 2P — это самый последний обнаруженный класс каналов K + .K 2P каналы или двухпоровые доменные калиевые каналы представляют собой тандемы четырех трансмембранных сегментов (M1-M4), содержащих двухпористый домен (P1 и P2) (Figures 1, 2). Они имеют протяженную внеклеточную петлю M1-P1 и цитозольные N- и C-концы. Каналы K 2P имеют уникальную последовательность сигнатуры поры Gly-Tyr (Phe) -Gly в 1-й поре (P1) и Gly-Leu (Phe) -Gly во 2-й поре (P2) (Honore, 2007; Рисунок 1) . TREK-1 был впервые клонирован из мозга мыши (Fink et al., 1996; рис. 3).TREK-1 был назван в честь канала TWIK-1, первого клонированного канала K 2P (Lesage et al., 1996). TREK-1 имеет 28% гомологию последовательности с каналом TWIK-1. TREK-1 высоко экспрессируется в головном мозге и легких, но также присутствует в почках, сердце и скелетных мышцах. Когда мы смотрим на локализацию мозга, TREK-1 высоко экспрессируется в нескольких областях мозга, таких как обонятельная луковица, гиппокамп, мозжечок и кора головного мозга (Fink et al., 1996).
Рисунок 1. Структура и классификация каналов К 2П . Семейство каналов K 2P состоит из 15 членов, сгруппированных в шесть подсемейств. Каналы K 2P представляют собой двухпоровые калиевые каналы и самый последний обнаруженный класс каналов K + . Они собираются как димеры четырех трансмембранных сегментов (M1 – M4) и двухпорового домена (P1 и P2). У них есть протяженная внеклеточная петля M1-P1 и цитозольные N- и C-концы. Каналы K 2P имеют уникальную последовательность сигнатуры пор Gly-Tyr (Phe) -Gly в 1-й поре (P1) и Gly-Leu (Phe) -Gly во 2-й поре (P2).
Рисунок 2. Регламент Полимодальный ТРЭК-1. ТРЕК-1 мультирегулируется множеством физических и химических раздражителей. TREK-1 обладает различными белками-партнерами, такими как AKAP150, β-COP, Mtap2 и сортилин. Сортилин взаимодействует с ТРЕК-1 и направляет его на плазматическую мембрану. Спадин представляет собой синтетический пептид, полученный из сортилина, который, как было показано, блокирует TREK-1 с высоким сродством. По-видимому, антидепрессивная активность спадина опосредуется активацией PI3K и Akt. TREK-1 участвует в многочисленных патологиях ЦНС, таких как депрессия, ишемия, эпилепсия и боль.
Рисунок 3. Ключевые этапы открытия каналов ТРЭК-1. На схеме представлены основные даты от клонирования каналов TREK-1 до открытия его роли в физиологии и патологии. АК, арахидоновая кислота; GPCR, рецептор, связанный с G-белком; STAR ∗ D, Альтернативы последовательного лечения для облегчения депрессии. Зеленые и красные треугольники представляют процессы активации и ингибирования TREK-1 соответственно.
TREK-1 показывает выпрямление наружу в симметричном состоянии K + из-за внешнего блока Mg 2+ , имеющего отрицательный потенциал, и механизма зависимости от напряжения (Maingret et al., 2002). Несмотря на отсутствие области измерения напряжения в каналах K 2P , TREK-1 и некоторые другие каналы K 2P демонстрируют сильную зависимость от напряжения. Источником этой чувствительности к напряжению является механизм стробирования ионного потока и перемещение от трех до четырех ионов K + в сильное электрическое поле неактивного селективного фильтра (Schewe et al., 2016). Однако эта зависимость от напряжения отключается физиологическими стимулами, такими как арахидоновая кислота (АК) и фосфатидилинозитолбисфосфат (PIP 2 ), которые превращают TREK-1 в классические каналы утечки (Schewe et al., 2016). ТРЕК-1 с ТРЕК-2 и ТРААК являются механо- и термочувствительными каналами K 2P (Honore, 2007). Они открываются за счет растяжения и набухания клеток. Нокаутные мыши TREK-1, TREK-2 и TRAAK гиперчувствительны к механической силе, у них проявляется механическая аллодиния и гипералгезия во время воспаления (Brohawn, 2015). Сообщалось, что механическая сила передается на TREK-1 и TRAAK непосредственно через липидный бислой (Brohawn et al., 2014). Эти каналы быстро открываются в ответ на напряжение и имеют низкий порог и широкий диапазон активации напряжения (Brohawn, 2015).
Физиологическая регуляция каналов ТРЕК-1
КаналТРЭК-1 регулируется различными физическими и химическими стимулами (рис. 2). TREK-1 открывается за счет механического растяжения в конфигурациях прикрепленных к клеткам и наизнанку как в клетках, трансфицированных COS, так и в ооцитах (Patel et al., 1998). Открытие ТРЭК-1 происходит при всех положительных и отрицательных потенциалах. Когда цитоскелет разрушается колхицином или цитохалазином D, активация TREK-1 за счет растяжения не изменяется (Patel et al., 1998).Показано, что TREK-1 чувствителен к изменению температуры в различных типах клеток, экспрессирующих TREK-1. Нагрев постепенно и обратимо активирует ТРЕК-1, достигая максимума активации при 37 ° C. Однако при 12 ° C базальный ток ТРЕК-1 подавляется (Maingret et al., 2000). Активация тепла TREK-1 нуждается в целостности клеток и цитозольных компонентах, поскольку TREK-1 нечувствителен к теплу, когда иссекается мембрана пятна (Maingret et al., 2000). TREK-1 активируется полиненасыщенными жирными кислотами (ПНЖК), такими как АК, в различных конфигурациях патч-зажима: цельноклеточная, клеточно-прикрепленная, внутренняя и внешняя (Patel et al., 1998). AA активирует TREK-1 дозозависимым образом и требует C-концевой части канала, который имеет решающее значение для активации и ингибирования после фосфорилирования протеинкиназой A (PKA). PIP 2 стимулирует нативный стриарный ток TREK-1 в конфигурации пластыря наизнанку (Chemin et al., 2005). В ооцитах Xenopus, экспрессирующих TREK-1, гидролиз PIP 2 ингибирует канал TREK-1, модулируя его зависимость от напряжения (Lopes et al., 2005). Также известно, что внутренний ацидоз вызывает активацию каналов TREK-1 с вовлечением С-концевой части (Maingret et al., 1999). Члены подсемейства TREK, включая TREK-1, TREK-2 и TRAAK, чувствительны к фосфатидной кислоте (PA), которая возникает в результате гидролиза фосфатидилхолина фосфолипазой D (PLD). Было показано, что TREK-1 и TREK-2, но не TRAAK, позитивно регулируются с помощью PLD2, который избирательно связывается с их C-концевыми доменами (Comoglio et al., 2014).
Белки-партнеры TREK-1
Протеомный подход, основанный на иммунопреципитации и масс-спектрометрическом анализе комплексов нативных каналов, позволил идентифицировать каркасный белок под названием AKAP150 (Sandoz et al., 2006; Фигура 2). Заякоренный белок А-киназы (AKAP) закрепляет регуляторную субъединицу PKA вблизи субстратов. AKAP150 образует сигнальные комплексы внутри нейрона, эти комплексы состоят из PKA, PKC, PP2B (протеинфосфатаза 2B), PSD-95, SAP97 и ионных каналов (Esseltine and Scott, 2013). Когда TREK-1 закрепляется с помощью AKAP150 в пост-M4 области, он становится активным каналом утечки, нечувствительным к стимулам, внутреннему закислению AA или механическому растяжению (Sandoz et al., 2006). Связанный с микротрубочками белок Mtap2 — еще один белок, который, как сообщается, связан с TREK-1 в головном мозге (Sandoz et al., 2008). Mtap2 связывается с микротрубочками и стабилизирует их. TREK-1 и Mtap2 совместно локализуются в нескольких областях мозга, таких как гиппокамп, мозжечок, обонятельная луковица, полосатое тело и кора. Подобно AKAP150, связывание Mtap2 с TREK-1 увеличивает его активность (Sandoz et al., 2008). Это увеличение амплитуды тока TREK-1 происходит не из-за прямого взаимодействия с Mtap2, а из-за увеличения уровня экспрессии канала на плазматической мембране. Внутри нейрона TREK-1, AKAP150 и Mtap2 одновременно обнаруживаются в постсинаптических окончаниях.Этот сигнальный комплекс регулирует активность канала TREK-1 и его доставку к плазматической мембране.
При использовании дрожжевой двугибридной техники библиотеки кДНК человека на N-концевой области TREK-1 сообщалось о прямом взаимодействии β-COP-TREK-1 (Kim et al., 2010). β-COP является субъединицей комплекса белков оболочки I (COPI), роль которого заключается в формировании покрытых везикул и управлении ретроградным движением от Гольджи обратно к эндоплазматическому ретикулуму (ER) или между различными компартментами внутри Гольджи (Гомес-Наварро и Миллер). , 2016).Было обнаружено, что истощение β-COP снижает мембранную экспрессию канала трансмембранного регулятора проводимости при муковисцидозе (CFTR). Мутации в этом хлоридном канале вызывают кистозный фиброз легких (Rennolds et al., 2008). β-COP увеличивает поверхностную экспрессию TREK-1 и амплитуду тока. β-COP участвует в прямом транспорте TREK-1 посредством прямого взаимодействия с N-концевой частью канала (Kim et al., 2010).
Самым недавно обнаруженным белком-партнером TREK-1 является сортилин (Petersen et al., 1997; Рисунок 2), также известный как рецептор нейротензина-3 (NTSR 3 ) (Mazella et al., 1998). Сортилин синтезируется в виде предшественника, называемого просортилином, который расщепляется в компартменте транс-Гольджи пропротеинконвертазой фурином с образованием зрелого сортилина и высвобождением 44 аминокислот, названных пропептидом (PE) (Munck Petersen et al., 1999 ). NTSR 3 состоит из большого просветного домена, одного трансмембранного сегмента и короткого С-концевого фрагмента. Количество NTSR 3 , экспрессируемых на плазматической мембране, не превышает 10%, в то время как большая часть экспрессируется внутриклеточно и участвует в основном во внутриклеточном движении.И NTSR 3 , и TREK-1 заметно экспрессируются в префронтальной (PFC) и поясной коре, миндалевидном теле, гиппокампе, прилежащем ядре, дорсальном ядре шва (DRN) и гипоталамусе. Экспрессия TREK-1 усиливалась на плазматической мембране в клетках COS-7, котрансфицированных NTSR 3 (Mazella et al., 2010).
ТРЭК-1 Гетеродимеризация
Первый член K 2P , клонированный TWIK-1 (Lesage et al., 1996), экспрессируется с TREK-1 в астроцитах (Zhou et al., 2009). Экспрессия функционального канала TWIK-1 делает необходимым его гетеродимеризацию с TREK-1, чтобы обеспечивать пассивную проводимость в астроцитах (Hwang et al., 2014). Как упоминалось ранее в этом обзоре, подсемейство TREK включает в себя TREK-1, TREK-2 и TRAAK. Несмотря на 78% гомологии между TREK-1 и TREK-2, эти два липид- и механоактивированных канала K + регулируются по-разному. Недавние исследования показали, что функциональные гетеродимеры TREK-1 образуются с каналами TREK-2 и TRAAK (Blin et al., 2016; Левиц и др., 2016). Гетеродимер ТРЕК-1-ТРЕК-2 имел минимальную активность при физиологическом pH. Однако комплекс ТРЕК-1-ТРЕК-2 активируется как в щелочной, так и в кислой среде. Гетеродимер TREK-1-TRAAK проявляет активацию как за счет внутриклеточного закисления, так и за счет подщелачивания (Levitz et al., 2016). Функциональные гетеродимеры TREK-1-TRAAK образуются и обладают уникальными биофизическими свойствами и различными типами регуляций (Blin et al., 2016). Гетеромеризация подсемейства TREK приводит к диверсификации каналов K 2P и обеспечивает различные виды регулирования.Недавно было показано, что усеченная форма (TRESK-MT) канала TRESK, K 2P , часто ассоциированная с мигренью, гетеромеризуется и ингибирует TREK-1 и TREK-2 (Royal et al., 2019). Гетеродимер TRESK-TREK действует как доминантно-отрицательный, повышающий возбудимость сенсорных нейронов тройничного нерва и мигренеподобный фенотип у мышей (Royal et al., 2019).
Фармакология каналов ТРЕК-1
Блокираторы ТРЭК-1
С момента открытия участия TREK-1 в нескольких патологиях ЦНС, очень большой интерес был посвящен поиску молекул, модуляторов его активности.Как показано в таблице 1, поиск блокаторов активности TREK-1 начался с открытия роли TREK-1 в процессах депрессии. Модель депрессии на мышах, лишенная kcnk2 (ген, кодирующий TREK-1), обнаруживает фенотип, устойчивый к развитию депрессии (Heurteaux et al., 2006). Спадин был первой молекулой, которая была разработана с целью блокировать канал TREK-1 и имитировать антидепрессантоподобный фенотип мышей kcnk2 — / — (Mazella et al., 2010; Djillani et al., 2018). Спадин — это пептид, синтезируемый из эндогенного PE, высвобождающегося после расщепления просортилина фурином. Позднее были разработаны несколько аналогов спадина для улучшения стабильности и антидепрессивной активности спадина in vivo (Veyssiere et al., 2015; Djillani et al., 2017). Из библиотеки из 22 пептидов более короткая пептидная последовательность PE 22–28 проявила более высокую эффективность ингибирования TREK-1, связанную с улучшенной стабильностью in vivo и антидепрессивными свойствами (Djillani et al., 2017). Подобные свойства были обнаружены у его биотинилированного продукта или аналога G / A-PE 22–28 (где Gly заменен на Ala). Эти аналоги спадина демонстрируют чрезвычайно высокое сродство к TREK-1, поскольку ингибирование происходит при наномолярных концентрациях. Более интересно то, что спадин и его аналоги являются специфическими блокаторами TREK-1, поскольку не наблюдалось никакого эффекта как на TREK-2, так и на TRAAK, двух ближайших к TREK-1 членов K 2P , которые принадлежат к одному подсемейству и имеют почти 80% гомология (Moha Ou Maati et al., 2012; Джиллани и др., 2017).
Таблица 1. Блокираторы ТРЭК-1.
Селективные ингибиторы обратного захвата серотонина (СИОЗС) — это семейство небольших молекул с высоким сродством к переносчику серотонина, SERT (Hirano et al., 2005). Блокада SERT вызывает накопление серотонина в синаптических щелях и проявляет антидепрессивный эффект в клинике. Кроме того, в клинических концентрациях СИОЗС и флуоксетин противодействуют каналу TREK-1 (IC 50 = 19 мкМ для флуоксетина и IC 50 = 9 мкМ для норфлуоксетина, его активного метаболита) (Kennard et al., 2005). Другие СИОЗС, такие как пароксетин, циталопрам или эсциталопрам, также блокируют TREK-1 с различной активностью. Однако SSRI неспецифичны для TREK-1, поскольку они нацелены на каналы TREK-2, Nav1.5 и L-типа Ca 2+ (Wong et al., 2005; Poulin et al., 2014; Mcclenaghan et al. , 2016).
Антипсихотические препараты, используемые для лечения психозов, таких как шизофрения и биполярное расстройство, оказались мощными блокаторами каналов TREK-1. Они противодействуют TREK-1 и TREK-2, но не влияют на каналы TRAAK (Таблица 1; Thummler et al., 2007). Блокада ТРЕК-1 наблюдалась как при применении типичного, так и атипичного антипсихотика. Например, хлорпромазин и локсапин дозозависимо блокируют каналы TREK-1 с IC 50 , равным 2,7 и 19,7 мкМ (Thummler et al., 2007). Эти данные демонстрируют потенциальную связь между TREK-1, шизофренией и биполярным расстройством.
Аналоги дигидропиридина, такие как амлодипин или нигулдипин, являются блокаторами каналов Ca 2+ L-типа. Было показано, что они также блокируют TREK-1 с высоким сродством (IC 50 = 0.43 и 0,75 мкМ для амлодипина и нигулдипина соответственно). Кроме того, мыши, лишенные канала L-типа Cav1.3, обнаруживают антидепрессант-подобный фенотип (Busquet et al., 2010). Соединение SID1900 блокирует TREK-1 с IC 50 ∼30 мкМ и, как было показано, обладает антидепрессантоподобными свойствами в модели хронического непредсказуемого легкого стресса (CUMS) на крысах (Ye et al., 2015). Эффект SID1900 был сопоставим со спадином. По-прежнему нет никаких свидетельств о других возможных целях SID1900, и необходимы дополнительные данные, чтобы сделать вывод о его специфичности.Наконец, сообщалось, что другие молекулы блокируют каналы TREK-1 и обеспечивают нейрозащиту от ишемии и инсульта, такие как 3-н-бутилфталид (NBP) и его аналог lig4-4 (Ji et al., 2011; Wang et al., 2018) . Однако, если информация о специфичности NBP отсутствует, сообщалось, что lig4-4 влияет на канал hERG, потенциал-зависимые каналы K + (K v ), нейронные каналы Na + и Ca 2+ , что может ограничить его развитие в качестве потенциальной нейрозащитной молекулы.
Активаторы ТРЭК-1
КаналTREK-1 активируется летучими средствами для общей анестезии, такими как хлороформ, диэтиловый эфир, галотан и изофлуран, и было показано, что С-концевая часть имеет решающее значение для его активации (Таблица 2; Patel et al., 1999). Если хлороформ кажется селективным для каналов TREK-1 с EC 50 (0,2–1,6 мМ), галотан и изофлуран активируют каналы TREK-1 и TASK. Кроме того, диэтиловый эфир открывает TREK-1, но снижает активность канала TASK (Patel et al., 1999). В отличие от общих анестетиков, местные анестетики, такие как бупивакаин, действуют как ингибиторы TREK-1 (Kindler et al., 1999). AA и другие PUFA открывают каналы TREK-1 дозозависимым образом (Patel et al., 1998). Считается, что открытие канала под действием AA, α-линоленовой кислоты (ALA) и докозагексаеновой кислоты (DHA) способствует нейрозащитному процессу (Lauritzen et al., 2000). Если антидепрессанты СИОЗС и нейролептики действуют как блокаторы TREK-1, сообщалось, что стабилизаторы настроения, такие как хлорид лития, и противоэпилептические препараты, такие как габапентин, вальпроат и карбамазепин, активируют канал TREK-1 (Kim et al., 2017). Соединение BL-1249 на основе тетразола показало интересную активацию TREK-1 с ЕС 50 около 1,5 мкМ in vitro в культивируемых миоцитах мочевого пузыря человека (Тертышникова и др., 2005). В клеточной линии карциномы поджелудочной железы BL-1249 активирует TREK-1 дозозависимо с IC 50 = 2 ± 2 мкМ (Sauter et al., 2016). Недавние исследования позволили лучше понять механизм действия BL-1249. Действительно, было показано, что BL-1249 активирует затвор фильтра селективности C-типа и избирательно активирует каналы TREK-1 и TREK-2 в 10 раз сильнее, чем каналы TRAAK (Pope et al., 2018). Кроме того, для действия BL-1249 необходим С-концевой хвост канала для активации, а трансмембранные домены M2 и M3 критичны для его селективности (Pope et al., 2018). После скрининга более 106 000 малых молекул были идентифицированы активатор ТРЕК-1, названный ML67, и его оптимизированный аналог ML67-33 (Багрянцев и др., 2013). Затем в той же лаборатории были недавно разработаны две молекулы с более высокой эффективностью, ML335 и ML402, которые усиливают активность TREK-1 с ЕС 50 из 5.2 ± 0,8 мкМ и 5,9 ± 1,6 мкМ соответственно (таблица 2; Lolicato et al., 2017). ML335 и ML402 связываются и активируют скрытый связывающий карман в фильтре селективности ворот C-типа канала TREK-1 (Lolicato et al., 2017). Сообщалось, что нестероидные противовоспалительные препараты (НПВП), такие как флуфенамовая кислота, нифлумовая кислота и мефенамовая кислота, активируют каналы TREK-1 независимо от ингибирования циклооксигеназы ЦОГ (Veale et al., 2014). Сообщалось, что производные кофейной кислоты активируют канал TREK-1 и оказывают сильное обезболивающее действие in vivo (Rodrigues et al., 2014; Vivier et al., 2017). Наконец, малая молекула GI-530159 была описана как открыватель TREK-1, который снижает возбудимость нейронов малых DRG (Loucif et al., 2018). Однако GI-530159 не является селективным для каналов TREK-1, поскольку он также активирует TREK-2 без значительного влияния на каналы TRAAK (Loucif et al., 2018).
Таблица 2. Активаторы ТРЭК-1.
Модуляторы ТРЭК-1
Рилузол (таблица 2) — нейропротекторная молекула, продаваемая как противосудорожное средство.Механизм его действия включает блокаду рецепторов глутамата. Также его назначают для продления выживаемости пациента, страдающего боковым амиотрофическим склерозом. Кроме того, было показано, что рилузол действует как временный активатор TREK-1 в течение 30 секунд и сильный ингибитор через 90 секунд (Duprat et al., 2000). Считается, что двойная активность в отношении TREK-1 опосредована посредством активации PKA циклическим аденозинмонофосфатом (цАМФ). С другой стороны, рилузол вызывает длительную активацию TRAAK без ингибирования.
ТРЕК-1 и заболевания ЦНС
каналов TREK-1 обнаруживают широкое распространение в головном мозге крыс и мышей (Hervieu et al., 2001; Talley et al., 2001). Рационально представить себе несколько ролей, которые ТРЕК-1 может играть в ЦНС. В этой части обзора мы подробно объясним, почему канал ТРЕК-1 стал очень многообещающей мишенью при ряде патологий, поражающих ЦНС (Рисунок 3).
ТРЭК-1 в депрессии
Роль TREK-1 в депрессии была продемонстрирована на мышах, лишенных kcnk2 , гена, кодирующего TREK-1 (Heurteaux et al., 2006). Нокаут-мыши TREK-1 демонстрировали фенотип, устойчивый к депрессии, в пяти различных моделях депрессии на мышах: тест принудительного плавания Порсолта (FST), тест подвешивания за хвост (TST), тест условного подавления подвижности (CSMT), усвоенный тест на беспомощность (LH) и тест с подавлением новизны (NSF) (таблица 3; Heurteaux et al., 2006). Поведение мышей kcnk2 — / — было похоже на поведение мышей, получавших острое или хроническое лечение классическими антидепрессантами, такими как флуоксетин (Heurteaux et al., 2006). Kcnk2 -дефицитные мыши показывают увеличение нейротрансмиссии серотонина (5-HT) в нейронах DRN. Удаление TREK-1 увеличивает нейрогенез, индуцированный длительным лечением антидепрессантами (Heurteaux et al., 2006). Блокирование ТРЕК-1 стало новой стратегией разработки антидепрессантов нового поколения. В клинически используемых концентрациях СИОЗС подавляют каналы TREK-1 (Kennard et al., 2005).
Таблица 3. ТРЕК-1 при патологиях ЦНС.
Спадин ранее был описан как специфический блокатор TREK-1 с высоким сродством (IC 50 = 40–70 нМ) (Mazella et al., 2010; Djillani et al., 2017, 2018). Спадин состоит из 17-аминокислотного пептида, синтезированного из пропептида (РЕ). PE представляет собой 44 аминокислоты, образованные в результате созревания сортилина в результате расщепления просортилина фурином (Munck Petersen et al., 1999). Спадин с высокой аффинностью связывает как TREK-1, так и рецептор сортилина / нейротензина 3 (NTSR3). Спадин, блокируя TREK-1, генерирует мышей с фенотипом устойчивости к депрессии в нескольких тестах на депрессию уже через 4 дня лечения. Подобно мышам kcnk2 — / — , спадин усиливает нейротрансмиссию 5-HT в серотонинергических нейронах DRN (Mazella et al., 2010). В экспериментах in vitro спадин стимулирует пути MAPK и PI3K во времени и в зависимости от концентрации. Спадин в концентрации 100 нМ увеличивает фосфорилирование ERK1 / 2 и Akt, но не влияет на фосфорилирование mTOR, что позволяет предположить оригинальный механизм действия спадина, отличный от другого быстродействующего антидепрессанта кетамина, который, как было показано, является mTOR-зависимым (Devader et al. др., 2015). Спадин оказывает защитное действие на нейроны против апоптоза каспазы-3, индуцированного стауроспорином, посредством специфической активации пути PI3K.Спадин временно увеличивает экспрессию мРНК PSD-95 и синапсина, двух маркеров синаптогенеза в головном мозге, через 8 часов. Это увеличение связано с кратковременным увеличением BDNF только через 5 часов. Спадин также увеличивает долю зрелых шипов в культуре корковых нейронов. In vivo ежедневное введение спадина в течение 4 дней увеличивало экспрессию мРНК BDNF, PSD-95 и синапсина всего через 7 дней в гиппокампе. Однако в префронтальной коре (ПФК) только BDNF усиливался через 3 недели (Devader et al., 2015).
Антидепрессивная активность спадина FST исчезает через 7 часов после острого лечения. Для улучшения биодоступности и стабильности in vivo спадина были сконструированы ретро-инверсные аналоги спадина и проведен скрининг на клеточной линии, стабильно экспрессирующей человеческий TREK-1 (Veyssiere et al., 2015). Были идентифицированы два аналога, 3 и 8. Они продлевают антидепрессивную активность с 7 до 16 часов при FST. Однако из-за некоторых проблем, связанных с токсичностью in vitro аналогов 3 и 8 при более высоких концентрациях, были применены другие стратегии для улучшения соотношения польза / риск.Путем сокращения последовательности спадина пептид PE 22-28, содержащий всего 7 аминокислот, был идентифицирован как самая короткая эффективная молекула, проявляющая антидепрессивную активность (Djillani et al., 2017, 2018). Роль TREK-1 у мышей была подтверждена другими исследованиями на людях, таких как STAR ( * ) D, которые подчеркивают тесную связь между устойчивостью к СИОЗС и наличием четырех однонуклеотидных полиморфизмов (SNP) в гене kcnk2 (Perlis и др., 2008). Другой SNP, rs6686529, расположенный в гене kcnk2 , оказался связанным с большим депрессивным расстройством и ответом на лечение антидепрессантами (Liou et al., 2009). Все эти данные подчеркивают ключевую роль TREK-1 в депрессии и необходимость создания блокаторов TREK-1 как многообещающего оригинального антидепрессанта (рис. 3).
ТРЭК-1 в нейропротекции
ТРЕК-1 в эпилептогенезе
TREK-1 экспрессируется в ГАМК-содержащих интернейронах в коре и гиппокампе (Hervieu et al., 2001). In vivo . Сообщалось, что активаторы TREK-1 ПНЖК, такие как α-линоленовая кислота, защищают крыс, получавших агонист рецептора глутамата, каиновую кислоту (КА) от судорог и поражений гиппокампа (Lauritzen et al., 2000). Более того, нейрозащита ПНЖК наблюдалась в другой модели приступов с использованием глутаматергических нейронов. Этот эффект был связан с ингибированием глутаматергической нейротрансмиссии (Lauritzen et al., 2000). Было показано, что мыши с дефицитом TREK-1 более уязвимы для развития эпилептических припадков, вызванных KA и пентилентетразолом (PTZ, антагонист рецептора GABA A ) (Heurteaux et al., 2004). Экспрессия c-fos , маркера возбудимости нейронов, была увеличена в пирамидных нейронах СА3 после лечения КА (Heurteaux et al., 2004). Линоленовая кислота и лизофосфатидилхолин (LPC) уменьшают KA-индуцированные судороги у мышей дикого типа. Однако не наблюдалось никакого эффекта у мышей, лишенных каналов TREK-1 (Heurteaux et al., 2004). Защитные эффекты линоленовой кислоты и LPC требуют наличия каналов TREK-1. Эпилептический статус (SE) — это постоянная повторяющаяся судорожная активность более 5 мин. При разработке мутанта TREK-1 (TREK-M) были получены постоянно открытые каналы TREK-1, которые показали устойчивость к подавлению PKA и PKC.ТРЕК-М был способен гиперполяризовать плазматическую мембрану и уменьшать спонтанное возбуждение нейронов гиппокампа в культуре (Dey et al., 2014). In vivo введение рекомбинантного аденоассоциированного вируса (AAV) — опосредованной доставки TREK-M как в энторинальную кору, так и в области CA3 гиппокампа, уменьшило на 50% продолжительность SE в мышиной модели SE, индуцированной литием и пилокарпином. AAV-TREK-M предотвращал гибель нейронов как в энторинальной коре, так и в CA3 после инъекции (таблица 3; Dey et al., 2014). Поскольку открытие TREK-1 приводит к нейропротекции против эпилептических эпизодов, блокирование этого активированного АА канала K 2P , напротив, может быть вредным. Удивительно, но антагонист TREK-1 спадин не усиливает припадки, индуцированные KA или PTZ, при введении мышам. Что еще более интересно, мыши, получавшие спадин, проявляют большую устойчивость к развитию генерализованных судорог и к смерти (Moha Ou Maati et al., 2012; Djillani et al., 2018).
ТРЭК-1 при ишемии
Используя модель глобальной ишемии, результаты транзиторной двусторонней окклюзии общих сонных артерий у мышей дикого типа и мышей TREK-1 с нокаутом показали, что 74% мышей TREK — / — умерли через 3 дня после ишемии, тогда как только 34% мышей погибли в группе дикого типа (Heurteaux et al., 2004). Здесь снова линоленовая кислота или предварительная обработка LPC не смогли защитить от ишемии у мышей с дефицитом TREK-1, в отличие от мышей дикого типа, у которых выживаемость мышей была значительно увеличена (Heurteaux et al., 2004). Ишемия спинного мозга у мышей индуцируется окклюзией дуги аорты и левой подключичной артерии. Сообщалось, что 75% мышей kcnk2 — / — умерли через 3 часа после 10-минутной ишемии, по сравнению с только 14% мышей дикого типа через 24 часа после ишемии (Heurteaux et al., 2004). Кроме того, у выживших мышей TREK-1 — / — развился тяжелый паралич задних конечностей. Однако у мышей дикого типа неврологического дефицита не наблюдалось (Heurteaux et al., 2004). Было показано, что помимо нейронов важную роль в ишемии мозга играют астроциты. TREK-1 способствует поддержанию крайне отрицательного мембранного потенциала астроцитов, который имеет решающее значение для контроля возбудимости клеток (Zhou et al., 2009). В культивируемых корковых астроцитах и в срезах гиппокампа широко экспрессируется TREK-1.Кортикальные и гиппокампальные каналы TREK-1 активируются во время астроглиоза после фокальной ишемии (Wang et al., 2012). В условиях гипоксии экспрессия белка TREK-1 в астроцитах также повышается, что увеличивает клиренс глутамата, подавляет астроцитарный маркер S100β и блокирует гибель нейронов (Таблица 3; Wu et al., 2013). Глутамат — главный возбуждающий нейротрансмиттер в головном мозге. Астроциты выделяют глутамат после активации GPCR. DH Woo et al. демонстрируют, что канал TREK-1 участвует в быстром высвобождении глутамата, что требует взаимодействия между N-концевым доменом TREK-1 и G βγ ПЦР G αi (Woo et al., 2012).
ТРЭК-1 под наркозом
При общей анестезии при клинических дозах, используемых для изофлурана, диэтилового эфира, галотана и хлороформа, активируются каналы TREK-1 (Patel et al., 1999). Активация канала TREK-1 подразумевает С-концевой фрагмент и приводит к гиперполяризации клеток, снижению потенциала действия и нейрозащите. Веселящий газ закись азота и ксенон в основном оказывают свое анестезирующее действие за счет антагонизма рецепторов NMDA, поскольку они обеспечивают обезболивание, эйфорию и нейрозащиту.Они не усиливают рецепторы GABA A (Gruss et al., 2004). Было показано, что закись азота и ксенон открывают каналы TREK-1 с Glu306 в качестве критически важной аминокислоты для анестезирующей активации TREK-1 (Gruss et al., 2004). Хлоралгидрат используется в педиатрии при некоторых формах эпилепсии, таких как прогрессирующий миоклонус и рефрактерная детская эпилепсия. При введении хлоралгидрат быстро метаболизируется в его активный метаболит 2,2,2-трихлорэтанол (TCE), который, как считается, отвечает за активность хлоралгидрата (Harinath and Sikdar, 2004).TCE вызывает угнетение ЦНС, усиливая рецепторы GABA A и ингибируя рецепторы NMDA, AMPA и Kainate. Сообщалось, что TCE также активирует каналы TREK-1 и TRAAK и способствует центральному анестезирующему эффекту (Harinath and Sikdar, 2004). In vivo , мыши, лишенные каналов TREK-1, проявляли более низкую чувствительность к наиболее сильным активаторам TREK-1, хлороформу и галотану, а также к севофлурану, десфлурану и изофлурану (Heurteaux et al., 2004). Порог провоцирования анестезии и дозы, необходимые для анестезирующего действия, были выше.Фенобарбитал, общий анестетик, который не модулирует каналы TREK-1, не может воспроизвести эти эффекты у мышей kcnk2 — / — . Таким образом, наблюдаемые эффекты были обусловлены именно отсутствием канала ТРЕК. Недавно было показано, что прекондиционирование in vitro, и in vivo, севофлураном обеспечивает нейрозащиту за счет активации каналов TREK-1 (Tong et al., 2014). Крысы, получавшие анестетик севофлуран после MCAO, проявляют нейрозащиту, которая, по-видимому, опосредуется каналами TREK-1 (Таблица 3; Pan et al., 2017).
Роль ТРЭК-1 в постинсультной депрессии
Учитывая нейрозащитную роль каналов TREK-1 при инсульте и их участие в процессе депрессии. Имеет смысл рассмотреть роль этого канала K 2P в физиопатологии постинсультной депрессии (PSD). В последнее время несколько лабораторий начали исследовать потенциальную роль каналов TREK-1 в этой патологии. PSD — нервно-психическое расстройство с высокой распространенностью, которое возникает после начала инсульта.Было показано, что уровни экспрессии канала TREK-1 были повышены в гиппокампе и PFC в модели PSD крыс (Table 3; Lin et al., 2015). Примечательно, что активация канала TREK-1 подавлялась после длительного лечения блокатором TREK-1 и SSRI, эсциталопрамом. В этой модели крысы PSD были воспроизведены основные действующие лица, вызывающие PSD у людей. Например, операция по окклюзии средней мозговой артерии (MCAO) была создана для имитации инсульта и хронического легкого стресса (CMS) и изоляции для имитации стресса и психосоциальных состояний соответственно (Lin et al., 2015).
TREK-1 в восприятии боли
Известно, чтоканалов ТРЭК-1 модулируются теплом. Эти каналы K 2P постепенно и обратимо открываются в зависимости от температуры. Примечательно, что открытие канала TREK-1 реверсируется цАМФ и простагландином E2, двумя сенсибилизаторами периферических и центральных терморецепторов. Блокада TREK-1 происходит из-за фосфорилирования Ser333 на его С-конце. Термочувствительный канал TREK-1 высоко экспрессируется и локализуется в нейронах малых ганглия задних корешков (DRG) и в нейронах центрального гипоталамуса, областях, которые сильно вовлечены в боль (Maingret et al., 2000). Шестьдесят процентов сенсорных нейронов экспрессируют TREK-1, и большинство из них связано с веществом P. Более 40% нейронов DRG, которые экспрессируют каналы TREK-1, также экспрессируют каналы теплового ноцицептора TRPV1. In vivo , по сравнению с мышами дикого типа, мыши, лишенные гена kcnk2 , очень чувствительны к низкопороговой тепловой боли и механическим стимулам (Alloui et al., 2006). Мыши с дефицитом TREK-1 показали усиленный очаговый воспалительный ответ после травмы спинного мозга (SCI).Кроме того, дефицит TREK-1 усиливает астроглиоз, апоптоз нейронов, демиелинизацию и замедляет восстановление моторики (Fang et al., 2017). Тем не менее, в других опубликованных исследованиях в DRG экспрессия микроРНК miR-183 была снижена, а экспрессия TREK-1 увеличена при нейропатической боли, вызванной хроническим сокращением седалищного нерва (CCI) (Table 3; Han et al., 2016; Shi et al., 2018). Интратекальная инъекция miR-183 уменьшала боль у крыс с моделью CCI и подавляла экспрессию TREK-1 (Shi et al., 2018). Недавно было сообщено, что модулятор TREK-1 рилузол предотвращает как сенсорный, так и моторный дефицит нейропатической боли, вызванной химиотерапевтическим препаратом оксалиплатином (Poupon et al., 2018). Тем не менее, выводы, сделанные в этой статье, не являются полностью убедительными, поскольку авторы рассматривают рилузол только как активатор и не принимают во внимание его ингибирующие эффекты. Наконец, каналы TREK-2 и TRAAK, другие члены подсемейства TREK, охарактеризованы в нейронах DRG небольшого размера крыс (Viatchenko-Karpinski et al., 2018).
Периферийные роли каналов ТРЭК-1
ТРЕК-1 в Сердце
В сердце активируемые растяжением каналы K + (SAK) реполяризуют клеточную мембрану и уравновешивают активность активируемых растяжением катионных каналов (SAC), которые, напротив, увеличивают приток катионов и деполяризацию клеток. Канал TREK-1 представляет собой серьезного кандидата на формирование SAK в сердце с высокой проводимостью Ca 2+ -активированного канала K + (BK Ca ) и каналов K ATP (Decher et al., 2017а). В сердце крысы все гены канала K 2P были обнаружены по меньшей мере в одной камере сердца с преобладанием экспрессии TWIK-2, TASK-1 и TREK-1. Канал TREK-1 был высоко экспрессирован в правом желудочке (Liu and Saint, 2004). В миоцитах предсердий новорожденных крыс с помощью одноканального пластыря, вывернутого наизнанку, ток, подобный TREK-1, активировался АА и внутренним ацидозом (Kim and Clapham, 1989). Было показано, что этот ток, подобный TREK-1, обратимо активируется летучими анестетиками, такими как хлороформ, галотан и изофлуран.Он также подавлялся аналогами цАМФ и агонистами β-адренергических рецепторов (Terrenoire et al., 2001). Ток SAK имеет идентичные биофизические свойства с рекомбинантным током TREK-1, такие как проводимость одного канала, отсутствие зависимости от напряжения и чувствительность к летучим анестетикам. В кардиомиоцитах желудочков были обнаружены два варианта TREK-1: большая проводимость и низкая проводимость с (132 ± 5 пСм) и (41 ± 5 пСм), соответственно, при положительных потенциалах (Xian Tao et al., 2006).Вариант TREK-1 с низкой проводимостью был вскрыт механическим растяжением, внутренним подкислением и АК. Биофизические свойства двух каналов TREK-1 были аналогичны тем, которые демонстрируют рекомбинантные каналы TREK-1, экспрессируемые в клетках HEK293. Используя кардиоспецифичных мышей с дефицитом TREK-1, было предположено, что TREK-1 играет существенную роль в возбудимости клеток синоатриального узла (Unudurthi et al., 2016). Недавно у пациентов с диагнозом тахикардия оттока правого желудочка (RVOT) была идентифицирована гетерозиготная точечная мутация в фильтре селективности канала TREK-1 (Decher et al., 2017б). Что еще более интересно, ТРЕК-М показал повышенную чувствительность к растяжению и проницаемость для ионов натрия (Decher et al., 2017b; Рисунок 3).
Роль ТРЕК-1 в других тканях
В дополнение к центральной роли каналов TREK-1 в головном мозге было совершенно рационально думать, что модуляция каналов TREK-1 может играть периферическую роль, поскольку этот канал K 2P экспрессируется в ряде тканей, таких как поджелудочная железа, клетки простаты и гладких мышц (Medhurst et al., 2001).Поджелудочная железа и особенно инсулин-секретирующие β-клетки представляют собой одну из основных мишеней периферических тканей, где TREK-1 и другие каналы K 2P играют важную роль в гомеостазе глюкозы (Kang et al., 2004; Mazella et al. , 2010; Dadi et al., 2014; Vierra et al., 2015). Учитывая важность канала TREK-1 как ключевого модулятора потенциала плазматической мембраны в дополнение к его регуляции различными физическими и химическими стимулами, его роль в гомеостазе глюкозы была исследована с использованием спадина (Hivelin et al., 2016). Спадин ингибирует каналы TREK-1 в линии β-клеток поджелудочной железы β-TC3 (Mazella et al., 2010). Известно, что регуляция секреции инсулина β-клетками поджелудочной железы тонко настраивается с помощью различных гормонов, таких как глюкагоноподобный пептид-1 (GLP-1), лептин, эстроген, мелатонин и гормон роста (Fu et al. , 2013). Хорошо известно, что АТФ-чувствительные калиевые каналы (K ATP ) играют критическую роль в поддержании гомеостаза глюкозы (Mctaggart et al., 2010). Каналы K ATP служат метаболическим сенсором для β-клеток поджелудочной железы.Когда уровни глюкозы повышаются, каналы K ATP закрываются, вызывая деполяризацию мембраны, приводящую к открытию каналов Ca 2+ , а также к секреции многочисленных гормонов и усилению эффектов глюкозы и других стимуляторов секреции, таких как GLP-1. (Mctaggart et al., 2010). Блокада TREK-1 спадином вызывает секрецию инсулина только при стимуляции глюкозой. Однако, в отличие от инкретиновых гормонов, спадин вызывает повышение Ca 2+ и высвобождение инсулина посредством оригинального PKA-независимого механизма (Hivelin et al., 2016). Каналы TREK-1 также ингибируются фосфорилированием PKA Ser333 в C-концевой части канала (Murbartián et al., 2005), что является следствием активации различных рецепторов, связанных с G-белком, в плазматическая мембрана. Связываясь со своими рецепторами GLP-1, GLP-1 и его аналоги, такие как эксендин-4, способны запускать синтез цАМФ и, таким образом, индуцировать активацию PKA, ведущую к секреции инсулина (Göke et al., 1993). Затем высвобождение инсулина β-клетками усиливается за счет закрытия TREK-1 после фосфорилирования PKA.Спадин или его аналоги могут заменить действие гормона инкретина и предложить терапевтическую альтернативу в случае неэффективности противодиабетического лечения с использованием миметиков GLP-1. Среди других каналов K 2P механочувствительный канал TREK-1 K 2P преимущественно экспрессируется в канале детрузора человека (Andersson and Arner, 2004). Недавнее исследование, оценивающее взаимосвязь между шестью SNP и возникновением симптомов гиперактивных нижних мочевыводящих путей (СНМП), выявило сильную связь между полиморфизмами TREK-1 (генотип rs758937019-CT и аллель rs758937019-T) и сверхактивными СНМП у человека (Недумаран и другие., 2019).
Заключение и перспективы
С момента их клонирования 20 лет назад физиологическое значение каналов TREK-1 продолжало расти (Рисунок 3). Сегодня было показано, что каналы TREK-1 важны, и их присутствие необходимо при ряде физиопатологических процессов. Их участие в этих различных процессах демонстрирует необходимость разработки фармакологических модуляторов, активаторов или ингибиторов этих каналов для коррекции любых дисфункций, связанных с TREK-1.
Несмотря на ряд исследований и скрининга многих молекул, было идентифицировано лишь несколько предполагаемых новых лекарств. Интересные результаты показывают активаторы серий ML и BL. Однако они демонстрируют более низкое сродство (в микромолярном диапазоне) и не специфичны для каналов TREK-1, поскольку открывают другие каналы K 2P . Открыватели ТРЕК-1 необходимы при таких патологиях, как ишемия, эпилепсия и боли. Задача состоит в том, чтобы улучшить их сродство, специфичность и способность преодолевать гематоэнцефалический барьер.Новые серии активаторов TREK ML и BL еще не тестировались при таких патологиях, как эпилепсия и ишемия. Было бы интересно исследовать любые нейрозащитные эффекты этих молекул на моделях эпилепсии или ишемии на грызунах. В то же время их специфичность должна быть улучшена, чтобы избежать каких-либо нецелевых эффектов, возникающих в результате модуляции других каналов K 2P .
В качестве ингибиторов наиболее перспективными молекулами являются спадин и его аналоги. Эти пептиды очень специфичны в семействе K 2P и обладают очень высоким сродством в пикомолярном диапазоне для более коротких аналогов (Djillani et al., 2017). Депрессия — это сложное расстройство настроения, в котором задействованы разные актеры. TREK-1 определенно является одним из наиболее многообещающих ионных каналов, играющих ключевую роль в нейробиологии депрессии. Блокаторы TREK-1, такие как спадин и короткие аналоги, обладают огромной терапевтической ценностью по сравнению с классическими антидепрессантами, назначаемыми в настоящее время. Они быстродействующие, обладают высокой аффинностью и специфичностью к ТРЕК-1. Что еще более интересно, они преодолевают гематоэнцефалический барьер, что облегчает их дальнейшую оптимизацию и развитие в качестве антидепрессантов, полезных в клинике.
В будущем, помимо поиска новых модуляторов TREK-1 для лечения нейропсихиатрических расстройств, актуальным будет изучение роли каналов TREK-1 в периферических тканях и понимание их участия в физиопатологии. Возникающая область, к которой ТРЕК-1 проявляет растущий интерес, — это сердце. Экспрессия TREK-1, как было установлено, изменяется при гипертрофии сердца (Wang et al., 2013). Кроме того, было обнаружено, что ТРЕК-М изменяет проницаемость каналов для натрия и вызывает желудочковую тахикардию (Decher et al., 2017б).
В заключение следует отметить, что широкое распространение TREK-1 в организме человека, разнообразные стимулы и регуляции, на которые он может реагировать, а также плейотропные роли, которые он может играть в физиологии и физиопатологии, делают TREK-1 многообещающей целью для более глубокого изучения, но в то же время очень сложно разработать модуляторы, специфичные для ткани.
Авторские взносы
г. н.э. написал рукопись. MB, CH и JM исправили это.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось в отсутствие каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Ссылки
Alloui, A., Zimmermann, K., Mamet, J., Duprat, F., Noel, J., Chemin, J., et al. (2006). TREK-1, K + канал, участвующий в полимодальном восприятии боли. EMBO J. 25, 2368–2376. DOI: 10.1038 / sj.emboj.7601116
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Багрянцев С. Н., Анг, К. Х., Галлардо-Годой, А., Кларк, К. А., Аркин, М. Р., Ренсло, А. Р. и др. (2013). Функциональный экран с высокой пропускной способностью идентифицирует небольшие молекулы-регуляторы температурно- и механо-чувствительных каналов K2P. ACS Chem. Биол. 8, 1841–1851. DOI: 10.1021 / cb400289x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Barcia, G., Fleming, M. R., Deligniere, A., Gazula, V. R., Brown, M. R., Langouet, M., et al. (2012). De novo мутации канала KCNT1 с усилением функции вызывают злокачественные мигрирующие парциальные припадки в младенчестве. Нат. Genet. 44, 1255–1259. DOI: 10,1038 / нг.2441
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Барел, О., Шалев, С.А., Офир, Р., Коэн, А., Злотогора, Дж., Шорер, З. и др. (2008). Унаследованный от матери синдром дисморфизма умственной отсталости Бирка Барела, вызванный мутацией в геномно импринтированном калиевом канале KCNK9. Am. J. Hum. Genet. 83, 193–199. DOI: 10.1016 / j.ajhg.2008.07.010
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бауэр, К. К., Каллигари, П., Радио, Ф. К., Капуто, В., Дентичи, М. Л., Фалах, Н. и др. (2018). Мутации в KCNK4, влияющие на стробирование, вызывают узнаваемый синдром нервного развития. Am. J. Hum. Genet. 103, 621–630. DOI: 10.1016 / j.ajhg.2018.09.001
PubMed Аннотация | CrossRef Полный текст |
| Нервная система Классы: 5 — 12 Нервная система для детей — На уроке рассматривается нервная система и все, за что она отвечает ибо в теле. 12:00 | |
| Нейроны и как они работают Класс: 5 — 12 Узнайте о нейронах — основных строительных блоках нервной системы. 4:50 | |
| Анатомия нейрона Класс: 5 — 12 Введение в нейрон и его анатомию. 6:15 | |
| Что, если бы мы могли заглянуть внутрь человеческого мозга? Класс: 5 — 12 Мозг делает нас теми, кто мы есть, но мы так мало понимаем, как он работает. Врачи узнают больше о мозге, используя новую технологию для наблюдения за пациентами с эпилепсией во время операции. 4:00 | |
| Человеческий мозг: как мы принимаем решения? Класс: 5 — 12 Взгляните на мозг, когда мы принимаем решения, будь то в проходе с хлопьями в местном магазине или во время пилотирования самолета. 2:35 | |
| Почему мы чувствуем боль? Классы: 5–12 В презентации рассказывается, как и почему мы чувствуем боль. 5:00 | |
| Анатомия глаза и как мы видим? Класс: 5 — 12 Урок о человеческом глазу с объяснением его анатомии и функций. 2:00 | |
| Анатомия уха и как мы слышим? Классы: 5–12 На этом уроке вы узнаете о человеческом ухе. Наше ухо состоит из трех частей: наружного, среднего и внутреннего уха. Все они работают вместе, чтобы передавать звук через ухо и в наш мозг. 2:15 | |
| Анатомия носа и как мы нюхаем? Класс: 5 — 12 Узнайте, как работает обоняние и как мы распознаем ароматы. 4:20 | |
| Наш язык и его вкус? Оценка: 5 — 12 Узнайте о чувстве вкуса — как что-то вкусное или неприятное на вкус! 2:30 | |
| Наша кожа и осязание Оценка: 5–12 Урок о нашем осязании. Кожа человека, наш самый большой орган, всегда усердно работает, чтобы держать мозг в курсе окружающей среды. 1:20 | |
| Скажите мне, почему: Головокружение Оценка: 5 — 12 Узнайте, почему у вас кружится голова, когда вы несколько раз вращаетесь. 3:10 | |
| Почему мы забываем вещи? Класс: 5 — 12 Вы, должно быть, заметили, как можно забыть, куда положить ключи от машины, но при этом помнить, как ездить на велосипеде. Из этого видео вы узнаете, почему мы забываем одни вещи и вспоминаем другие. 1:35 | |
| Как наш мозг заставляет нас забыть? Класс: 5 — 12 Знаете ли вы, что ваш мозг на самом деле стирает ваши воспоминания? Видео проливает свет на то, почему мы со временем забываем некоторые воспоминания. 3:40 | |
| Почему вашему мозгу нужен сон? Класс: 5 — 12 Сон может показаться пустой тратой времени.Но ученые только что обнаружили, насколько это важно для нашего здоровья. Узнайте, что именно происходит во время сна. 3:05 | |
| Как работает ваш мозг? Класс: 5 — 12 Человеческий мозг, возможно, самый сложный объект в известной вселенной, не является безупречным. Посмотрите эти уловки и иллюзии, чтобы понять, как работает мозг. 4:40 | |
| Как работает наша память? Класс: 5 — 12 Видео объясняет, как хранятся наши воспоминания и что мы можем сделать, чтобы улучшить нашу память. 2:40 | |
| Как мы создаем воспоминания? Класс: 5–12 Узнайте, как наш мозг по-разному создает и вспоминает воспоминания. 9:55 | |
| Чудо в человеческом мозге Оценка: 5 — 12 Узнайте, как наш мозг работает с сотнями триллионов удивительных связей между миллиардами нейронных клеток. 1:25 | |
| Анатомия и функции мозга Класс: 5–12 На уроке объясняются структуры и функции основных частей человеческого мозга — ствола мозга, мозжечка, таламуса. , и головной мозг.Здесь также обсуждаются эволюционные аспекты мозга. 13:55 | |
| Human Brain Project Оценка: 5–12 Узнайте о проекте Human Brain Project — исследовательском проекте, целью которого является моделирование человеческого мозга с помощью суперкомпьютеров для лучшего понимания как это работает. 2:30 | |
| Нервные системы Оценка: 8–12 Урок нервной системы.Он дает обзор мозга. На уроке обсуждаются основные части нейрона, принципы его работы и объясняется, как нейротрансмиттеры передают сообщения через синапс. 17:40 | |
| Сенсорная система Оценка: 8–12 Урок наших чувств. Он объясняет, как мы воспринимаем окружающую среду с помощью нашей сенсорной системы. Он подробно описывает то, как люди обоняют, видят и слышат. 10:30 | |
| Нервная система A&P Урок, часть 1 Класс: 8 — 12 Урок, посвященный компонентам нервной системы центральной и периферической нервной системы.Центральная нервная система состоит из головного и спинного мозга, периферическая нервная система состоит из всех других нервных тканей в организме, таких как нервы, идущие к спинному мозгу и от него, и нервы в конечностях. 10:00 | |
| Нервная система A&P Урок, часть 2 Класс: 8-12 10:00 | |
| 33 | 6 Класс: 5 — 12 Урок по эндокринной системе.Он объясняет основные элементы эндокринной системы и то, как железы вырабатывают гормоны, которые являются мишенями для клеток. Он описывает типы гормонов и действие десяти эндокринных желез — шишковидной железы, передней доли гипофиза, задней доли гипофиза, щитовидной железы, паращитовидной железы, поджелудочной железы, коры надпочечников, мозгового вещества надпочечников, яичек и яичников. 10:30 |
| Эндокринная система: как она работает Оценка: 5–12 Узнайте, как работает наша эндокринная система.Эндокринные железы выделяют гормоны, которые регулируют различные функции человека, такие как метаболизм, рост, функции тканей, сон и т. Д. 2:30 | |
Центральная нервная система | Статья о центральной нервной системе по The Free Dictionary
основная часть нервной системы животных и человека, состоящая из нервных клеток (нейронов) и их выступающих частей (отростков). Он включает в себя систему тесно взаимосвязанных групп нервных клеток (ганглиев) у беспозвоночных и спинного и головного мозга у позвоночных, включая человека.Основная и специфическая функция центральной нервной системы (ЦНС) — вызывать простые и сложные высокодифференцированные рефлексы. У высших животных и человека нижние и средние отделы ЦНС — спинной мозг, продолговатый мозг, средний мозг, промежуточный мозг и мозжечок — регулируют деятельность органов и систем высокоразвитого организма, осуществляют контакт и взаимодействие между ними. , и поддерживают единство организма и целостность его деятельности. Высшие отделы ЦНС — кора головного мозга и смежные подкорковые структуры — в значительной степени регулируют взаимоотношения организма с внешней средой.
Структура и функции . ЦНС связана со всеми органами и тканями периферической нервной системой. У позвоночных это включает черепные нервы, выходящие из головного мозга; спинномозговые нервы, которые отходят от спинного мозга; ганглии межпозвонковых нервов; и периферическая часть вегетативной нервной системы, то есть система нервных ганглиев с эфферентными нервными волокнами, которые входят (преганглионарные) или выходят (постганглионарные) ганглии. Сенсорные или афферентные аддукторные волокна проводят возбуждение в ЦНС от периферических рецепторов, а эфферентные (моторные и вегетативные) отводящие волокна проводят возбуждение от ЦНС к клеткам таких эффекторов, как мышцы, железы и кровь.
Рисунок 1 . Схема рефлекторной дуги спинномозгового рефлекса: (а) трехнейронная дуга, (б) двухнейронная дуга, (R) рецептор, (E) эффектор, (1) афферентный нейрон, (2) интеркалярный нейрон, ( 3) эфферентный нейрон
сосудов. Во всех отделах ЦНС есть афферентные нейроны, которые получают стимулы с периферии, а также эфферентные нейроны, передающие нервные импульсы различным эффекторным органам на периферии. Афферентные и эфферентные клетки способны взаимодействовать посредством своих отростков и образовывать двухнейронную рефлекторную дугу, которая влияет на такие элементарные рефлексы, как сухожильные рефлексы спинного мозга.В целом, однако, есть вставочные нервные клетки или интернейроны, расположенные в рефлекторной дуге между афферентными и эфферентными нейронами (см. Рисунок 1). Контакт между различными отделами ЦНС также устанавливается многочисленными процессами их афферентных, эфферентных и интеркалярных нейронов, которые образуют короткие и длинные проводящие пути. ЦНС также включает нейроглиальные клетки, которые выполняют вспомогательную функцию и участвуют в метаболизме нервных клеток.
Рефлекторный принцип деятельности ЦНС .Рефлекторный принцип деятельности ЦНС был экспериментально установлен еще до XIX века, но изучался только применительно к деятельности тех отделов, которые расположены ниже уровня полушарий головного мозга. Ученые, занимающиеся ЦНС, выяснили общие механизмы и адаптивный характер рефлекторной деятельности ЦНС, а также функциональные характеристики ее отделов. В 19 веке И. М. Сеченов и ряд других прогрессивных ученых выдвинули смелую идею о том, что деятельность высших отделов ЦНС, в том числе и умственная, носит рефлекторный характер.Эта прогрессивная, материалистическая идея легла в основу классических экспериментальных и теоретических исследований И. П. Павлова, которые привели к его теории высшей нервной деятельности. Павлов установил, что рефлексы, вызываемые у высокоразвитых организмов корой головного мозга, в отличие от рефлексов нижних отделов ЦНС, не являются врожденными, а развиваются в течение жизни организма при его взаимодействии с окружающей средой; Рефлексы коры головного мозга позволяют организму с максимальной эффективностью адаптироваться к окружающей среде.Павлов назвал этот новый класс рефлексов условными рефлексами, в отличие от врожденных или безусловных рефлексов.
Отвергая первоначальный, рудиментарный взгляд на рефлекс как на стереотипную, механическую и пассивную реакцию ЦНС, классические исследования нейрофизиологов Э. Пфлюгера, Сеченова, Павлова и К. Шеррингтона установили, что безусловные и, в частности, условные рефлексы очень динамичны и изменчивы: реакции на одни и те же стимулы меняются в зависимости от внешних и внутренних условий окружающей среды и от функционального состояния самой ЦНС.
Основные паттерны активности ЦНС тесно связаны с характеристиками рефлекторной дуги, которая является структурной основой каждого рефлекторного действия. Рефлекторная дуга проводит возбуждение только в одном направлении, от рецепторного конца к эффектору, благодаря структурной и функциональной поляризации, присущей всем нервным клеткам. Микроструктуры, называемые синапсами, которые расположены на концевых ветвях аксона каждого нейрона, позволяют нейрону вступать в контакт с телами или дендритами других нейронов и передавать им свою активность в одностороннем порядке (закон Белла-Мажанди).Разнообразные внешние и внутренние рецепторы в ходе эволюции стали специализироваться в своей способности точно реагировать на качественно различные типы энергии — световую, акустическую, тепловую, механическую и химическую. Рецепторы трансформируют эту энергию в процессе нервного возбуждения, которая последовательно передается от одних элементов рефлекторной дуги к другим в виде ритмических импульсов. Возбуждение претерпевает значительные изменения в ритме, интенсивности, скорости и характере на своем многоступенчатом пути к последнему элементу.Рефлекторное возбуждение может вызывать в эффекторах различные эффекты из-за структурных и функциональных характеристик самих эффекторов (мышц, желез или кровеносных сосудов).
Функции центральных отделов рефлекторной дуги, в отличие от функций простых нервных проводников, характеризуются относительно медленным началом и наличием возбуждения и фазовых колебаний уровня возбудимости, производимых волнами. возбуждения. Эта уникальная функциональная инертность — относительно длительное сохранение состояния высокой возбудимости — вызывает феномен суммирования (слабые, неэффективные стимулы становятся эффективными при повторении), а также связанный с ним феномен ослабления.
Центральные элементы рефлекторной дуги могут изменять или трансформировать ритм раздражения. Возникающее в результате возбуждение обычно возникает со своим собственным ритмом волн, ритмом, который иногда быстрее, а иногда медленнее, чем ритм первоначальной стимуляции. Между силой раздражения и интенсивностью рефлекторной реакции существует только относительное соответствие, которое обычно наблюдается после умеренного раздражения. Если стимуляция сильная и продолжительная, нервные центры тормозятся, и в этом случае слабая стимуляция начинает вызывать больший рефлекторный эффект, чем сильная стимуляция, явление, известное как парадоксальный рефлекс.Относительно высокая предрасположенность к утомлению и выраженная чувствительность к биохимическим изменениям внутренней среды организма, особенно к недостатку кислорода в крови и к некоторым токсинам, также характерны для центральных элементов рефлекторной дуги. Все эти особенности обусловлены в основном свойствами синапсов, дендритов и тел самих нейронов.
Рефлекторная дуга обычно изображается в упрощенном виде, как цепь отдельных рецепторных, афферентных, интеркалярных, эфферентных и эффекторных клеток.В действительности же рефлекторная дуга состоит из множества таких цепочек, звеньями которых являются не отдельные клетки различного типа, а совокупность однородных взаимосвязанных клеток. Совокупность рецепторных клеток образует рефлексогенную зону рефлекса, группа эффекторных клеток составляет рабочий механизм рефлекса, а совокупность нейронов в ЦНС образует соответствующий нервный центр.
Павлов определил нервный центр как совокупность нервных элементов, расположенных в разных частях ЦНС, тесно связанных между собой, составляющих единую систему и регулирующих определенную функцию организма.Развивая и конкретизируя эту концепцию, Э. А. Асратян предложил рассматривать центральную часть безусловной рефлекторной дуги как многоступенчатую структуру, состоящую из нескольких ветвей; каждая ветвь проходит через один из отделов ЦНС, участвующих в данном рефлексе, и характеризуется специфичностью (см. рис. 2). Эти ветви имеют неодинаковое значение в реализации рефлексов: ветви, расположенные на одних уровнях, используются для определенных рефлексов, а ветви на других уровнях используются для других рефлексов.Например, основная ветвь дуги, связанная с сердечно-сосудистыми и респираторными рефлексами, проходит через продолговатый мозг, а основная ветвь дуги, связанная с пищеварительными, защитными и сексуальными рефлексами, проходит через промежуточный мозг. Ветвь многоступенчатой дуги, проходящая через кору головного мозга, соответствует тому, что Павлов назвал корковым представлением безусловного рефлекса, который является основой для образования соответствующих условных рефлексов.
Рисунок 2. Схема рефлекторной дуги с многоступенчатым центральным делением: (A) афферентный нейрон, (E) эфферентный нейрон, (I – V) уровни ветвей центрального отдела дуги
Координация рефлексов . ЦНС воздействует на множество разнообразных рефлексов, различающихся рецепторными, центральными и эффекторными механизмами, а также типом, характером, направлением, происхождением и степенью сложности. Тем не менее, эти рефлексы не возникают или возникают нескоординированно или хаотично, а имеют определенный состав, порядок и последовательность в соответствии с текущими потребностями организма в единой, интегрированной и адаптивной деятельности.Координирующая деятельность ЦНС определяется ее структурными и функциональными характеристиками. Степень генетического родства между различными структурными элементами ЦНС и характер их взаимоотношений являются важными предпосылками их взаимодействия и координации рефлексов. Особое значение в этой связи имеют структурные характеристики, известные как расходящиеся и сходящиеся пути. Посредством расходящихся путей многочисленные конечные ветви аксонов приводят каждый афферентный нейрон в контакт со многими эфферентными нейронами либо напрямую, либо через промежуточный нейрон (см. Рис. 3, а).Это позволяет одному афферентному нейрону активировать множество близких и удаленных эфферентных нейронов и связанных с ними рефлексов в определенной последовательности. Например, умеренное раздражение подушечки задней лапы децеребрированной кошки вначале вызывает рефлекс сгибания таранно-пяточного сустава. Постепенное усиление возбуждающего тока последовательно вызывает рефлексы сгибания коленного и тазобедренного суставов одной конечности, рефлекс разгибания противоположной задней ноги, рефлекс разгибания одноименной передней ноги и, наконец, рефлекс сгибания. противоположная передняя нога.Эти реакции являются результатом постепенного рассеивания (облучения) возбуждения, возникающего в афферентном элементе первичного рефлекса, на родственные структуры близлежащих и удаленных рефлексов спинного мозга посредством коротких и длинных внутриспинальных проводящих путей.
Рисунок 3. Диаграмма, изображающая расхождение (а) и конвергенцию (б) нервных путей в спинном мозге: (1) афферентный нейрон, (2) интернейрон, (3) эфферентный нейрон
С помощью конвергентного pathways ряд афферентных нейронов вступает в контакт с эфферентным нейроном терминальными ветвями аксонов либо напрямую, либо через промежуточные нейроны (см. рисунок 3, b).В последнем случае ветви нескольких афферентных нейронов сходятся на одном интернейроне, который служит уникальным общим путем. Ветви нескольких таких интернейронов, в свою очередь, сходятся к одному эфферентному нейрону или их совокупности, как к общему терминальному пути, который блокирован для других потоков возбуждения. Конвергенция путей в ЦНС делает возможным использование одного эфферентного нейрона множеством однородных и гетерогенных афферентных нейронов, которых в несколько раз больше, чем эфферентных нейронов.Например, отдельная умеренная стимуляция двух разных областей рецептивного поля рефлекса царапания не вызывает соответствующих рефлексов. Однако одновременное раздражение обеих областей током одинаковой силы вызывает отчетливый рефлекс из-за суммирования подпороговых возбуждений. Взаимное усиление рефлексов достигается также одновременной стимуляцией рецептивных полей двух разных рефлексов, имеющих общий терминальный путь и вызывающих одинаковый эффект.
Феномены дивергенции и конвергенции первоначально были обнаружены и изучены в связи с активностью спинного мозга, но позже было обнаружено, что они существуют и в других отделах ЦНС, особенно в коре головного мозга. Поскольку афферентные нервные элементы значительно превосходят эфферентные элементы в высших отделах ЦНС, принцип конвергенции множественных путей к сенсомоторной области, главному кортикальному общему пути, весьма выражен. Это дает возможность афферентным нервам влиять на различные безусловные двигательные рефлексы, а различные условные рефлексы могут развиваться из этих безусловных рефлексов.
Недавно разработанные электрофизиологические методы позволили таким современным нейрофизиологам, как итальянец Дж. Моруцци и П. Г. Костюк, открыть и изучить микроструктурные и микрофункциональные основы отдельных нейронов. Эти ученые также идентифицировали и изучили механизмы дивергенции и конвергенции, особенно в нейронах спинного мозга, ретикулярной формации, таламусе, коре головного мозга и других отделах ЦНС. Было обнаружено, что тело и дендриты каждого нейрона в ретикулярной формации и коре головного мозга имеют синаптические контакты с множеством других нейронов, которые активируются стимулами разных модальностей и оказывают как возбуждающее, так и тормозное влияние на общий путь.
Такие факторы, как исходное функциональное состояние нервных структур, участвующих в рефлексе, сила, биологическая роль и продолжительность внешнего раздражителя, а также влияние внутренних нервных и гуморальных факторов, имеют большое значение для координации рефлексов и в деятельности ЦНС. Однако главной функциональной основой координации рефлексов является способность ЦНС активировать посредством возбуждения определенные синаптические контакты и пути и, в частности, одновременно блокировать ингибирование других синаптических контактов и путей.Эти процессы происходят в большом количестве комбинаций и пространственных и временных отношений в соответствии с текущими потребностями организма и существующими условиями внешней среды.
Ключевая роль торможения в координирующей деятельности ЦНС четко проявляется в антагонистических взаимодействиях рефлексов, особенно когда одновременно стимулируются разные рецепторы или рецептивные поля, имеющие общий эффектор, но вызывающие разные виды рефлексов. Такие взаимно тормозящие отношения существуют между локомоторными, защитными и отложенными рефлексами, в которых роль играют идентичные мотонейроны-разгибатели и мышцы конечностей, иннервируемых ими.Каждый из этих рефлексов может возникать отдельно и беспрепятственно, если стимулировать только его рецептивное поле. Но если во время одного из рефлексов стимулируется рецептивное поле другого рефлекса или если стимулируются рецептивные поля обоих рефлексов, возникает конфликт из-за контроля общего терминального пути. Преобладает рефлекс, рецептивное поле которого в данный момент стимулируется сильнее, и тот, который более существенен для организма в данных условиях.Остальные рефлексы блокируются тормозным процессом, создавая тем самым благоприятные условия для беспрепятственного осуществления преодолевающего рефлекса.
Другой пример такой координации рефлексов — реципрокная иннервация антагонистических мышц конечностей, дыхательной и других систем. Шеррингтон и российские физиологи П. А. Спиро и Н. Е. Введенский показали, что рефлекторное возбуждение мотонейронов мышц-сгибателей конечностей сопровождается угнетением мотонейронов анатомических антагонистов этих мышц, мышц-разгибателей, и наоборот.Антагонистические взаимоотношения также могут проявляться рефлексами, принадлежащими разным функциональным системам; например, при глотании на короткое время останавливаются дыхательные движения. Большое значение торможения в координирующей функции ЦНС может быть ясно показано путем введения экспериментальному животному стрихнина, который временно лишает ЦНС способности развивать торможение. У животного почти полностью исчезает координация рефлексов: любое локальное раздражение вызывает одновременную двигательную реакцию всех мышц тела.
Сеченов и голландский физиолог Р. Магнус обнаружили важность исходного функционального состояния ЦНС с точки зрения ее координирующей деятельности. Стимуляция данного рецептивного поля задней лапы животного после перерезки его спинного мозга вызывает противоположные эффекты, в зависимости от исходного состояния стимулируемой конечности. Если конечность разгибается, стимуляция вызывает рефлекс сгибателя, а если конечность сгибается, стимуляция вызывает рефлекс разгибателя.Исходное положение конечности отражается в соответствующем нервном центре через возбуждение соответствующих нервных путей кожи и мышц; в результате функциональное состояние центра меняется.
Рецепторы, встроенные в эффекторы, и особенно в двигательный аппарат, информируют соответствующие структуры ЦНС об исходном состоянии эффектора в покое, а также о природе, интенсивности, продолжительности и динамике активности эффектора. Непрерывный поток информации от рецепторов функционирующих эффекторов играет важную роль в настройке и саморегуляции деятельности в соответствии с текущими потребностями организма и данной ситуацией, с которой он сталкивается.Физиологам давно известно о важной роли, которую играет в ЦНС этот важный принцип рефлекторной саморегуляции функций. Названный термином «обратная связь», он в конечном итоге стал фундаментальным принципом кибернетики.
Уникальное круговое взаимодействие между центральными и периферийными элементами рефлекторной дуги иногда проявляется во взаимной активации, а иногда в круговом вращении возбуждения внутри элементов дуги. Это круговое взаимодействие также проявляется в формировании особой цепочки рефлексов: рефлекторное сокращение мышцы стимулирует ее рецепторы, которые, в свою очередь, вызывают новое рефлекторное сокращение той же мышцы (C.Белл, А. Ф. Самойлов). Круговое взаимодействие также происходит между нейронами центрального элемента рефлекторной дуги и проявляется в разных формах. Примером может служить явление, известное как торможение Реншоу, названное в честь американского нейрофизиолога Б. Реншоу. В этом явлении аксон спинномозгового мотонейрона посылает в спинной мозг повторяющийся коллатераль, который контактирует с тем же мотонейроном посредством тормозящего интернейрона и который подавляет этот мотонейрон, если он становится чрезмерно возбужденным.Этот феномен отрицательной обратной связи был описан в отношении пирамидных нервных клеток коры головного мозга и клеток других структур ЦНС.
Примером положительной обратной связи является феномен неуклонно усиливающегося нервного возбуждения, описанный испанским физиологом Р. Лоренте де Нет. Это явление вызвано продолжительной циркуляцией возбуждения в многоразветвленной замкнутой цепи нейронов в рефлекторном центре: рекуррентный коллатераль одного из этих нейронов входит в контакт с одним или несколькими интернейронами.Эти интернейроны, когда они входят в контакт с исходным нейроном, замыкают цепь и формируют структурную основу для повторяющейся циркуляции возбуждения и уникального типа самоусиления.
Скоординированное и антагонистическое взаимодействия рефлексов неразрывно связаны и составляют разные аспекты единого координирующего процесса. Когда существует общий терминальный путь, согласованные рефлексы накапливаются и усиливают друг друга. С другой стороны, антагонистические рефлексы вступают в конфликт на этом пути, а преодолевающий рефлекс подавляет своих соперников на определенный период времени.В определенных ситуациях, например, под влиянием таких внешних или внутренних факторов, как местное механическое давление, гормон, токсин или поляризующий ток, возбудимость центрального механизма рефлекса существенно и равномерно увеличивается, и этот механизм временно увеличивается. становится уникальным общим терминальным путем.
Когда стимулируются рецептивные поля разнородных рефлексов, а не рефлексы, специфичные для этих полей, результатом является начало или усиление рефлекса, центр которого ранее испытывал усиленное возбуждение.Этот тип координации рефлексов был идентифицирован и изучен в нескольких модификациях, которые в современной нейрофизиологии называют облегчением путей, рефлексом суммирования, ослаблением и доминантой. Считается, что нервный центр, находясь в состоянии повышенной возбудимости или раздражения, подавляет определенные элементы центральных структур различных рефлексов. Он также отвлекает от обычного маршрута возбуждение, идущее по начальным элементам рефлекторной дуги, как бы притягивая их к себе.Наконец, он достигает кульминации с собственным возбуждением и инициирует или усиливает рефлекс, который важен для организма в данный момент.
Согласованность рефлексов зависит также от функционального состояния их центрального механизма. Например, снижение возбудимости этого механизма из-за усталости, нейрогуморальных факторов или токсинов приводит к тому, что даже ранее доминировавшие рефлексы легко уступают общий терминальный путь другим рефлексам. Таким образом, разнообразные формы координации рефлексов, такие как координационная деятельность ЦНС в целом, основаны на активных нервных процессах, которые имеют противоположные эффекты.К ним относятся возбуждение и торможение, различные комбинации и вариации распределения возбуждения и торможения в очень сложных макроструктурных и микроструктурных элементах ЦНС, сложная динамика, взаимное проникновение, а также конфликт и взаимодействие между этими процессами.
Подчинение, специализация и локализация функций . Для деятельности ЦНС характерно функциональное подчинение, то есть иерархическое ранжирование подразделений системы, которое развивалось в течение длительного периода времени.Это иерархическое ранжирование, а также структурная неоднородность и функциональное неравенство ЦНС проявляются на относительно ранних стадиях исторического развития организмов. Центральные нервные образования и рецепторы головы развиваются раньше, чем в других частях тела. Главный отдел ЦНС развивается за счет увеличения ее массы, непрерывной структурной дифференциации и функциональной специализации существующих отделов системы. Главный отдел ЦНС также развивается за счет постоянного появления в ней новых центральных образований, рефлекторная активность которых находится на все более высоком уровне и имеет все большее значение; эти образования направляют и регулируют деятельность всех нижних отделов ЦНС.Этот процесс непрерывного развития, специализации, локализации и подчинения функций ЦНС достигает своего наиболее сложного уровня у высших млекопитающих, особенно у человекообразных обезьян и человека.
ЦНС находится в состоянии тонуса даже при отсутствии видимых внешних признаков ее активности. Тоническое возбуждение ЦНС — это проявление общей функциональной готовности к началу и продолжению деятельности. Подчинение структур, особенно в продолговатом мозге, среднем и промежуточном мозге, может быть отчетливо обнаружено при тоническом возбуждении ЦНС.У высших животных, например, хирургическое рассечение или криогенная блокада путей между продолговатым мозгом и спинным мозгом (то есть прекращение подчиняющего влияния продолговатого мозга на спинной мозг) приводит к спинному шоку — глубокому и глубокому поражению. длительное угнетение спинномозговых рефлексов. Следствием такой перерезки на уровне верхних бугорков является феномен децеребральной жесткости, то есть сильное тоническое сокращение всех мышц-разгибателей и потеря способности животного принимать или сохранять нормальную позу.
Подчиненность отделов ЦНС еще более очевидна в их деятельности. Каждый высший элемент ЦНС воздействует на рефлексы, которые становятся все более сложными с точки зрения структуры и состава, и интегрирует их более полно, одновременно задействуя в своей деятельности рефлексы, которые регулируются низшими элементами ЦНС. Характеристики рефлекторной деятельности основных отделов ЦНС можно описать следующим образом. Рефлексы сегментов спинного мозга затрагивают только отдельные части тела, например, конечности.Более сложные рефлексы продолговатого мозга распространяются на пищеварительную, дыхательную, сердечно-сосудистую и двигательную системы. Меэнцефалические рефлексы задействуют всю скелетную мускулатуру тела и координируют такие сложные двигательные функции, как стояние и ходьба. Рефлексы диэнцефальных структур регулируют и координируют деятельность внутренних органов всех систем организма во всех возможных комбинациях и в гармонии с их жизненно важными безусловными рефлексами, например, хватанием пищи, защитными и сексуальными рефлексами.Полушария головного мозга способны совершенствовать все эти рефлексы, объединять их в комплексы рефлексов и создавать качественно новые типы рефлексов — условные рефлексы. Таким образом, чем выше уровень развития животного и чем выше уровень организации его ЦНС, тем больше преобладание высших отделов над низшими и тем значительнее их участие в регуляции функций организма.
Возрастающее значение высших отделов ЦНС в ходе эволюции жизненных процессов организма называется церебрализацией, энцефализацией или кортикализацией функций.Однако нижние отделы ЦНС также влияют на высшие, и вся информация от внешних и внутренних органов постепенно передается вверх. Следовательно, феномен подчинения ЦНС следует рассматривать просто как выражение преобладающего направления в сложном и разнообразном взаимодействии между нейронными структурами разных уровней. Например, ретикулярная формация сильно стимулирует и подавляет функциональное состояние почти всех отделов ЦНС, включая кору головного мозга.В свою очередь, кора головного мозга влияет на функциональное состояние и активность ретикулярной формации и других глубоких структур мозга, включая промежуточные ганглии восходящих путей, регулируя тем самым поток информации, которую они передают. Круговое взаимодействие между структурами ЦНС, а также саморегуляция их функций подтверждают правильность точки зрения Павлова о том, что кора головного мозга играет важную роль в объединенной и интегративной деятельности всей ЦНС.
Структурные и функциональные характеристики ЦНС определяют разнообразие и эффективность действий, которые она выполняет для удовлетворения нормальных потребностей организма, а также его требований к новым формам координации. Обильные резервные возможности и компенсаторные приспособления ЦНС имеют биологическое значение как во время нормального существования организма, так и после повреждения периферических сенсорных и эффекторных органов, афферентных и эфферентных нервных структур и органов самой ЦНС.
СПИСОК ЛИТЕРАТУРЫ
Prohaska J. Физиология, или наука о естественном человеческом . СПб., 1822.Орбели Л.А. Лекции по физиологии нервной системы , 3-е изд. Москва-Ленинград, 1938.
Декарт, Р. Избр. произв. Москва, 1950. (Пер. С французского и латинского)
Ухтомский А.А. Собр. соч. , т. 1. Ленинград, 1950. Павлов,
, И. П. Полн. собр. соч. , 2-е изд., тт. 1–6. Москва-Ленинград, 1951–52.
Введенский, Н.Е. Полн. собр. соч. , т. 4. Ленинград, 1953.
Сеченов, И. М. Избр. произв ., тт. 1–2. Москва, 1952–56.
Ройтбак, А.И. Биоэлектрические излучения в коре больших полушарий , парти. Тбилиси, 1955.
Магнус, р. Услановка тел. . Москва-Ленинград, 1962.
Магоун Х. Бодрствующий мозг , 2-е изд. Москва, 1965. (Пер. С англ.)
Беритов И.С. Общая физиология мышечной и нервной системы , 3-е изд., Т. 2. Москва, 1966.
Самойлов А.Ф. Избр. Труды . Москва, 1967.
Анохин П.К. Биология и нейрофизиология условного рефлекса . М., 1968.
Русинов В.С. Доминанта: Электрофизиологические исследования . М., 1969.
Асратян Э.А. Очки по физиологии условных рефлексов . Москва, 1970.
Физиология высшей нервной деятельности , ч. 1–2.Москва, 1970–71.
Костюк П.Г. Физиология центральной нервной системы . Киев, 1977.
Ливанов М. Н. Пространственная организация апроцессов головного мозга . Москва, 1972.
Рабинович М. Я. Замыкательная функция мозга . Москва, 1975.
Большая Советская Энциклопедия, 3-е издание (1970-1979). © 2010 The Gale Group, Inc. Все права защищены.
Функциональные области мозга и их удивительные возможности
Классическое представление о человеческом мозге — это командный центр вашей нервной системы.Однако исследования показывают, что это намного сложнее, чем просто коммутатор между ушами.
Ваш мозг настолько силен и разнообразен, что сложно назвать все его функции. Нейробиологи попытались разбить различные части мозга, чтобы объяснить его структуру.
Они известны как функциональных областей мозга .
В этой статье мы исследуем эти различные области мозга и их функции.
Как работает мозг?
Структура человеческого мозга — это иерархия, состоящая из примерно 85 миллиардов нейронов.Эти нейроны посылают сигналы через 18–32 триллиона крошечных пространств, называемых синапсами. Каждый синапс может передавать сигнал 0,1–2 раза в секунду. Это соответствует от 18 до 640 триллионов сигналов, которые ваш мозг отправляет и принимает каждую секунду.
Вдобавок ко всему, он хранит часть передаваемой информации в виде памяти. Как система хранения данных долговременный жесткий диск мозга может быть бесконечным. Например, электрические импульсы могут вызывать воспоминания, которые, как предполагалось, были давно забыты.Считается, что мозг может запоминать все от рождения до смерти.
Какая основная часть мозга?
Кора головного мозга — это то, что отличает человеческий мозг от других организмов. Он организован во многих физических масштабах, начиная с уровня отдельных нейронов и заканчивая функциональными системами.
Текущие исследования функциональной визуализации классифицируют структуру мозга на три уровня. Это области, сети и системы.
В основном известно, что мозг состоит из функциональных областей, которые объединены в сети.Однако абсолютный вклад каждой области в поведение еще предстоит должным образом определить.
Новые крупные исследования объединяют данные изображений мозга и поведенческие данные. Это позволяет исследователям изучать более широкие данные о моделях сканирования мозга, сравнивая нейронные маркеры и функциональные результаты.
Левое полушарие мозга и правое полушарие
Во-первых, мозг человека делится на левое и правое полушария. Они связаны пучком нервных волокон, называемым мозолистым телом.Две стороны сильно, хотя и не полностью симметричны.
Левое полушарие управляет мышцами правой стороны тела. Затем правое полушарие контролирует левую сторону. Одно полушарие может быть немного доминирующим. Вроде левша или правша.
В чем разница между мышлением левого и правого полушария?
Идея «левого полушария» и «правого полушария» — это качества, не подтвержденные исследованиями. Однако между этими областями есть некоторые важные различия.Левое полушарие содержит области, участвующие в речи и языке (область Брока и область Вернике). Он также выполняет математические вычисления и поиск фактов. Это считается логической или академической стороной мозга.
Правое полушарие мозга играет роль в визуальных, слуховых и пространственных навыках. Обычно считается, что они приравниваются к художественным способностям, которые считаются более инстинктивными или творческими по своей природе. Однако в этих функциях задействованы оба полушария.
Сканирование изображений показывает, что люди используют разные пропорции своего мозга.Когда вы используете свой мозг, вы задействуете и левую, и правую функциональные области.
Функциональные области коры головного мозга
Различные области коры головного мозга контролируют разные функции.
Самый большой участок мозга человека — это головной мозг. Он также известен как кора головного мозга и контролирует многие мысли и процессы, которые мы приравниваем к человеческому разуму.
