Анаболизм и катаболизм. Тема: Здоровье | by Eggheado | Eggheado: Health
Человек, занимающийся спортом, должен хотя бы отдаленно иметь представление о процессах, происходящих в его организме. Это позволит ему составить правильный режим питания и тренировок, что, в свою очередь, приведет к достижению отличного результата. Сегодня мы поговорим о важнейшем процессе в организме человека — обмене веществ и его составляющих, анаболизме и катаболизме.
Итак, обмен веществ или метаболизм — это совокупность химических реакций, протекающих в организме, обеспечивающих его рост, развитие и процессы жизнедеятельности, взаимодействие с окружающей средой и т.д.
Человек получает готовые органические вещества с пищей, но чтобы они смогли участвовать в обмене их необходимо расщепить на элементарные частицы, т.к. организму необходимо использовать во всех процессах свои, присущие только ему жиры, белки и углеводы. Эти процессы происходят в пищеварительной системе.
Белки расщепляются ферментами до аминокислот. В клетках из них строятся белки тела. Белки входят в состав клеток, участвуют в процессах свертываемости крови, транспортировки газов, входят в состав костей. Они способны к окислению с выделением энергии, которая в дальнейшем будет использоваться организмом.
Жиры распадаются в организме на глицерин и жирные кислоты. Образуется жир, характерный для организма. Далее он отправляется в депо клетки, там он используется как запасное вещество и строительный материал. Жиры входят в состав мембран клеток, выполняют защитную функцию, сохраняют тепло. Более того, жиры — источник энергии, они способны выделять при окислении больше энергии, чем белки и углеводы.
Углеводы расщепляются в организме до глюкозы и других простых углеводов. Издержки сахаров превращаются в гликоген и другие соединения, а остальные распределяются между всеми клетками. Глюкоза — отличный источник энергии.
Одной из составляющих обмена веществ является анаболизм, или по-другому, пластический обмен.
Анаболизм — это совокупность химических реакций, направленных на образование клеток и тканей. В результате образуется новый материал для построения клеток и их роста, а так же запасается энергия.
Примерами анаболизма могут служить следующие процессы: создание новых клеток или мышечных волокон, синтез белков и т.д.
Простыми словами анаболизм — это создание новых веществ или тканей в организме.
Анаболизм неразрывно связан с обратным ему, катаболизмом, т.е. разрушением на более простые вещества.
Этот термин приобрел негативную окраску среди спортсменов и это совсем не правильно. Ведь расщепление жиров и углеводов с дальнейшим выделением энергии — это тоже катаболизм. А эта энергия расходуется при работе мышц на тренировках и т.д.
Также в ходе катаболизма происходит распад устарелых тканей и клеточных элементов. В дальнейшем продукты этого распада удаляются из организма. Именно катаболизм и анаболизм имеют большое значение для атлета, серьезно относящегося к своей спортивной карьере. Эти процессы протекают в организме одновременно, но в разные периоды времени один процесс преобладает над другим. Например, после еды преобладают анаболические процессы, после сна — катаболические. Более того первая стадия анаболизма является последней стадией катаболизма.
Но катаболизм действительно может оказывать негативное влияние на результаты спортсмена, т.к. в ходе него разрушается мышечная ткань. Разнообразные диеты, стрессы, недосыпание усиливают катаболические процессы в организме спортсмена.
Уменьшить это разрушительное влияние поможет правильно питание, питание до и после тренировки, употребление ВСАА, протеина, а так же пищи, богатой белком.
По материалам: paladincenter.ru
КАТАБОЛИЗМ — это… Что такое КАТАБОЛИЗМ?
КАТАБОЛИЗМ — (от греч. katabole низвержение, разрушение), термин для обозначения процессов распада веществ в организме вообще, т.е. процессов диссимиляции (см.), и в этом смысле противополагаемый анаболизму (см.). Т. о. анаболизм и К. в совокупности обнимают… … Большая медицинская энциклопедия
КАТАБОЛИЗМ — (от греч. katabole сбрасывание разрушение) (диссимиляция), совокупность протекающих в живом организме ферментативных реакций расщепления сложных органических веществ (в т. ч. пищевых). В процессе катаболизма происходит освобождение энергии,… … Большой Энциклопедический словарь
КАТАБОЛИЗМ — КАТАБОЛИЗМ, см. МЕТАБОЛИЗМ … Научно-технический энциклопедический словарь
катаболизм — ДИССИМИЛЯЦИЯ – совокупность реакций метаболизма, приводящих к расщеплению сложных органических веществ. Сопровождается синтезом АТФ (по этой причине под К. понимают обычно энергетический обмен клетки), низкомолекулярных соединений,… … Словарь микробиологии
катаболизм — сущ., кол во синонимов: 1 • диссимиляция (3) Словарь синонимов ASIS. В.Н. Тришин. 2013 … Словарь синонимов
катаболизм — катаболизм. См. диссимиляция. (Источник: «Англо русский толковый словарь генетических терминов». Арефьев В. А., Лисовенко Л.А., Москва: Изд во ВНИРО, 1995 г.) … Молекулярная биология и генетика. Толковый словарь.
КАТАБОЛИЗМ — см. Диссимиляция. Экологический энциклопедический словарь. Кишинев: Главная редакция Молдавской советской энциклопедии. И.И. Дедю. 1989 … Экологический словарь
катаболизм — а, м. catabolisme m. <гр. katabole сбрасывание вниз. биол. Вид метаболизма: совокупность реакций обмена веществ в организме, соответствующее диссимиляции и направленных на распад органических веществ составных частей клеток и тканей: противоп … Исторический словарь галлицизмов русского языка
катаболизм — – совокупность реакций, направленных на распад сложных молекул, как поступивших с пищей, так и уже входящих в состав клеток организма, до более простых веществ (конечных продуктов обмена) … Краткий словарь биохимических терминов
катаболизм — Процесс разложения органических веществ в живом организме с выделением энергии [http://www. dunwoodypress.com/148/PDF/Biotech Eng Rus.pdf] Тематики биотехнологии EN catabolism … Справочник технического переводчика
Что такое анаболизм и катаболизм? | Спорт
Анаболизм и катаболизм одинаково нужные в организме процессы,и стоит узнать о них подробнее, чтобы не верить многочисленным мифам.
Записавшись в спортзал, от тренера вы часто будете слышать такие термины, как анаболизм, катаболизм и обмен веществ.
Слово “катаболизм” может вселять страх, ведь это распад мышц, как объяснил инструктор, а анаболизму, наоборот, воздаются оды и каждый тренирующийся должен к нему непременно стремиться, закрывая углеводное окно, или намешивая протеиновые коктейли прямо между подходами.
Но не все так просто. И анаболизм, и катаболизм одинаково нужные в организме процессы, поэтому стоит узнать о них подробнее, чтобы не доверять многочисленным мифам на эту тему.
Какая взаимосвязь между анаболизмом, анаболиками и анаболическим эффектом?
Анаболизм – это биохимический процесс в организме человека, благодаря которому создаются новые соединения на молекулярном уровне. Простыми словами – это генерация клеток и синтез белков и гормонов, благодаря которым происходит рост мышечных волокон, чего добиваются все спортсмены.
Анаболизм происходит под воздействием питательных веществ, минералов и витаминов, поступающих в организм в достаточном количестве.
С анаболизмом связано несколько понятий в микробиологии и медицине, одно из них – анаболический эффект.
Это взрывной рост клеток в организме, вследствие реакции на интенсивный тренинг, смену режима питания, спортивные добавки или анаболики.
Анаболизм может быть не только у мышечной ткани, но и у жировой, в широком смысле слова это понятие означает рост и обновление любых клеток в организме человека.
Но если говорить об анаболизме, как о процессе увеличения мышечных волокон, то он зависит от многих факторов:
Режим питания, сна и отдыха.
2. Регулярность тренировок и смена тренировочных программ.
3. Отсутствие стресса и полноценное восстановление.
4. Конституция тела и индивидуальный метаболизм.
Метаболизм или обмен веществ напрямую связан с анаболическим и катаболическим процессами, которые являются его составляющими. Скорость метаболизма отличается у людей разного телосложения, образа жизни и возраста.
У детей метаболизм очень быстрый, поэтому они так любят сладкую пищу, изобилующую быстрыми углеводами, которые нужны для получения мгновенной энергии, которую растущий организм растрачивает полностью.
У людей разного типа телосложения метаболические процессы различаются.
Выделяют три типа телосложения:
• Эктоморф
• Мезоморф
• Эндоморф
Эктоморфы – худые от природы, у них быстрый метаболизм, и им требуется гораздо больше усилий для анаболизма мышц, так как катаболические процессы преобладают в их организме.
Мезоморфы имеют от природы атлетическое телосложение, их мышцы легко отзываются на нагрузку, анаболизм и катаболизм находятся в балансе.
Эндоморфы склонны к полноте, анаболизм превалирует над катаболизмом, они легко растят как мышечную, так и жировую ткань.
В зависимости от типа телосложения следует подбирать режим тренировок и рацион питания.
К примеру, эндоморфам нужно есть больше белковых продуктов и сокращать жиры и углеводы, а эктоморфам не следует бояться жиров и углеводов, ведь если их будет недостаточно в рационе, организм будет брать энергию из протеинов, и рост мышц будет очень медленным.
Отдых между тренировками важен, так как во время полноценного отдых происходит полное восстановление организма, это время активного роста мышц, поэтому не стоит пренебрегать днями отдыха от спортзала.
Особенно, если вы не занимаетесь спортом профессионально. Да, тренирующиеся спортсмены проводят до двух тренировок в один день и почти во все дни недели, умудряясь не только не терять массу, но и набирать ее.
Это им удается благодаря бесчисленным спортивным добавкам, которые помогают быстрее восстанавливаться и тренироваться эффективнее, протеину и мегакалорийному рациону питания с большим количеством белка.
Обычному любителю достаточно 3-4 тренировок в неделю на постоянной основе, чтобы видеть прогресс в развитии силы и выносливости, изменения в теле и прирост мышечной массы.
Но, даже занимаясь регулярно, можно прийти к тому, что вы перестанете замечать собственную эволюцию в тренинге.
Многие в этот период начинают принимать различные препараты и покупать спортивное питание.
Но прежде всего, нужно обратить внимание на свою программу тренировок, которую желательно изменять или обновлять каждые три месяца. Не лишним будет изменить род физических нагрузок, например, заняться любым новым видом фитнесса.
Рацион спортсмена должен быть богат белковой пищей. Чем больше мышечной массы вы имеете, тем больше белка должно быть в рационе. Белок нужен для недопущения процесса распада мышц, для их поддержания и роста.
Сколько белка нужно именно вам, можно рассчитать по специальным формулам, которые легко найти в интернете, но не забывайте корректировать усредненные цифры, ориентируясь на индивидуальную конституцию тела.
Время сна – это время восстановления и обновления всех функций организма на клеточном уровне.
Для анаболизма мышц сон особенно важен, ведь во время сна затягиваются микротравмы мышечных волокон, полученные в результате тренинга, и, регенерируя, мышцы гипертрофируются.
Стоит ли бояться катаболизма?
Процесс, противоположный анаболическому – катаболизм. Это расщепление веществ на молекулярном уровне, распад сложных соединений на простые.
Катаболическим называется процесс расщепления белков, жиров и углеводов, получаемых из пищи, чтобы организм мог нормально функционировать.
Благодаря одному процессу происходит другой, процессы анаболизма и катаболизма взаимосвязаны и вместе они представляют собой метаболизм (обмен веществ) в организме.
Без одного процесса невозможен второй, поэтому глупо бояться катаболизма и верить мифам о нем.
Но если применять термины эмпирически, то понятно, что спортсмены боятся не катаболизма в целом, а потери мышечной массы, которую не так просто набрать, особенно эктоморфам.
Как не допустить катаболизма мышц:
Тренироваться регулярно и периодически менять программу тренировок.
2. Спать по 8-9 часов в сутки, регулярно отдыхать, отвлекаясь от забот и проблем.
3. Избегать стресса и потрясений, расслабляться.
4. Хорошо питаться, есть много белка или добирать его протеином.
Хороший, быстрый обмен веществ – это признак здорового человека. Если у вас есть какие-либо проблемы с организмом, недомогания или заболевания, лучше пройти медобследование перед посещением зала.
От уровня метаболизма зависит скорость его основных процессов, а значит, время и силы на постройку мышц.
Теперь вы знаете значение анаболизма и катаболизма в процессе строительства собственного тела, а значит, сумеете грамотно применить полученные знания на практике, чтобы тренироваться максимально эффективно и получать регулярный и полноценный анаболизм.
2. Энергетический обмен (катаболизм, диссимиляция)
Универсальным источником энергии во всех клетках служит АТФ (аденозинтрифосфат, или аденозинтрифосфорная кислота).
Все энергетические затраты любой клетки обеспечиваются за счёт универсального энергетического вещества — АТФ.
АТФ синтезируется в результате реакции фосфорилирования, то есть присоединения одного остатка фосфорной кислоты к молекуле АДФ (аденозиндифосфата):
АДФ + h4PO4+ 40 кДж = АТФ + h3O.
Энергия запасается в форме энергии химических связей АТФ. Химические связи АТФ, при разрыве которых выделяется много энергии, называются макроэргическими.
При распаде АТФ до АДФ клетка за счёт разрыва макроэргической связи получит приблизительно \(40\) кДж энергии.
Энергия для синтеза АТФ из АДФ выделяется в процессе диссимиляции.
Энергетический обмен (диссимиляция, катаболизм) — это совокупность химических реакций постепенного распада органических соединений, сопровождающихся высвобождением энергии, часть которой расходуется на синтез АТФ.
В зависимости от среды обитания организма, диссимиляция может проходить в два или в три этапа.
Процессы расщепления органических соединений у аэробных организмов происходят в три этапа: подготовительный, бескислородный и кислородный.
В результате этого органические вещества распадаются до простейших неорганических соединений.
У анаэробных организмов, обитающих в бескислородной среде и не нуждающихся в кислороде (а также у аэробных организмов при недостатке кислорода), диссимиляция происходит в два этапа: подготовительный и бескислородный.
В двухэтапном энергетическом обмене энергии запасается гораздо меньше, чем в трёхэтапном.
Первый этап — подготовительный
Подготовительный этап заключается в распаде крупных органических молекул до более простых: полисахаридов — до моносахаридов, липидов — до глицерина и жирных кислот, белков — до аминокислот.
Этот процесс называется пищеварением. У многоклеточных организмов он осуществляется в желудочно-кишечном тракте с помощью пищеварительных ферментов. У одноклеточных организмов — происходит под действием ферментов лизосом.
В ходе биохимических реакций, происходящих на этом этапе, энергии выделяется мало, она рассеивается в виде тепла, и АТФ не образуется.
Второй этап — бескислородный (гликолиз)
Второй (бескислородный) этап заключается в ферментативном расщеплении органических веществ, которые были получены в ходе подготовительного этапа. Кислород в реакциях этого этапа не участвует.
Биологический смысл второго этапа заключается в начале постепенного расщепления и окисления глюкозы с накоплением энергии в виде \(2\) молекул АТФ.
Процесс бескислородного расщепления глюкозы называется гликолиз.
Гликолиз происходит в цитоплазме клеток.
Он состоит из нескольких последовательных реакций превращения молекулы глюкозы C6h22O6 в две молекулы пировиноградной кислоты — ПВК C3h5O3 и две молекулы АТФ (в виде которой запасается примерно \(40\) % энергии, выделившейся при гликолизе). Остальная энергия (около \(60\) %) рассеивается в виде тепла.
Получившаяся пировиноградная кислота при недостатке кислорода в клетках животных, а также клетках многих грибов и микроорганизмов, превращается в молочную кислоту C3H6O3.
В мышцах человека при больших нагрузках и нехватке кислорода образуется молочная кислота и появляется боль. У нетренированных людей это происходит быстрее, чем у людей тренированных.
При недостатке кислорода в клетках растений, а также в клетках некоторых грибов (например, дрожжей), вместо гликолиза происходит спиртовое брожение: пировиноградная кислота распадается на этиловый спирт C2H5OH и углекислый газ CO2:
В результате гликолиза глюкоза распадается не до конечных продуктов (CO2 и h3O), а до богатых энергией соединений (молочная кислота, этиловый спирт) которые, окисляясь дальше, могут дать её в больших количествах. Поэтому у аэробных организмов после гликолиза (или спиртового брожения) следует третий, завершающий этап энергетического обмена — полное кислородное расщепление, или клеточное дыхание.
Этот этап происходит на кристах митохондрий.
Третий этап, так же как и гликолиз, является многостадийным и состоит из двух последовательных процессов — цикла Кребса и окислительного фосфорилирования.
Третий (кислородный) этап заключается в том, что при кислородном дыхании ПВК окисляется до окончательных продуктов — углекислого газа и воды, а энергия, выделяющаяся при окислении, запасается в виде \(36\) молекул АТФ (\(2\) молекулы в цикле Кребса и \(34\) молекулы в ходе окислительного фосфорилирования).
Этот этап можно представить себе в следующем виде:
Вспомним, что ещё две молекулы АТФ запасаются в ходе бескислородного расщепления каждой молекулы глюкозы (на втором, бескислородном, этапе). Значит, суммарный результат полного окисления молекулы глюкозы составляет \(38\) молекул АТФ.
Суммарное уравнение энергетического обмена:
C6h22O6+6O2=6CO2+6h3O+38АТФ.
В реакциях энергетического обмена используется не только глюкоза, но и липиды, белки. Но главным источником энергии в большинстве клеток являются углеводы.
Катаболизм веществ его этапы и процессы (Общая схема)
Катаболизм (диссимиляция ) — это энергетический обмен, часть метаболизма, распада сложных веществ на более простые (или окисления вещества), который протекает с освобождением энергии в виде тепла и в виде молекулы АТФ, универсального источника энергии всех биохимических процессов.
Схема катаболизма пищевых веществ, его этапы, пути и процессы
На схеме наглядно представлен катаболизм пищевых веществ, который состоит из 3-х основных этапов (стадий), первый и второй этапы относятся к специфическим путям катаболизма, а третий этап относится к общему пути катаболизма.
ATP (АТФ) — это аденозинтрифосфорная кислота (нуклеотид), универсальный источник и переносчик энергии, который участвует во всех биохимических процессах.
NAD (НАД) — никотинамидадениндинуклеотидфосфата, является коферментом, переносит электроны из одной реакции в другую в метаболизме.
NADH (НАД*Н) — востановленная форма NAD.
Пируват — это соли пировиноградной кислоты, конечный продукт в процессе гликолиза.
Этапы катаболизма
Протекание процессов катаболизма
Энергетическая ценность
1-й этап катаболизма (подготовительный)
расщепление больших макромолекул на простые субьединицы
На первом этапе происходит расщепление пищевых веществ (белки, полисохариды, жиры) до мономеров. У многоклеточных организмов это осуществляется в пищеварительном тракте (у простейших в лизосомах при самообновленнии клеток) под воздействием соответствующих ферментов, после чего полученные мономеры всасываются в кровь (моносахариды аминокислоты) и в лимфу (жирные кислоты).
Небольшое количество энергии рассеивается ввиде тепла
2-й этап катаболизма (бескислородный)
расщепление простых субьединиц на ацетил-СоА, сопровождающийся образованием ограниченного количества АТP и NADH
На втором этапе все пищевые продукты которые поступают в клетку из крови образуют ацетилкоэнзим А (ацетил-СоА). Это соединение, а также другие ферменты, включающие в себя КоА, являются ключевыми звеньями множества разнообразных биохимических реакций.
— При гликолизе моносахариды образуют пировиноградную кислоту.
— При катаболизме аминокислот образуются ацетил-CoA, пируват, другие кетокислоты
— При β-окислении жирных кислот образуется ацетил-CoA
Второй этап происходит в цитозоли и митохондрии.
При расщеплении глюкозы 60% выделившейся энергии дает тепло, 40% идет на синтез 2х молекул ATP, эта часть энергии запасается.
3-й этап катаболизма (кислородный)
при полном окислении ацетил-СоА до H2O и CO2 образуется большое количество NADH, что обеспечивает синтез большого количества ATP при переносе электронов
Третий этап и его реакции проходят в митохондриях. Ацетил-CoA участвует в реакциях цикла лимонной кислоты (цикл трикарбоновых кислот Кребса), там углероды окисляются до углекислого газ CO2. Происходит полное окисление ацетильной группы ацетил-СоА до Н2O и СO2, при этом большое количество электронов и протонов запасается на молекулах NADH (процесс «окислительное фосфолирование»). В дальнейшем энергия электронов используется для образования протонного градиента, что обеспечивает последующий синтез АТР.
2C3H6O3 + 6O
2 + 36H3PO4 + 36ADP —> 6CO2 + 38H2O + 36ATP
При окислении 2х молекул кислоты образуется 36 молекул ATP
_______________
Источник информации: Биология для поступающих в вузы / Г. Л. Билич, В.А. Крыжановский. — 2008
Анаболизм и катаболизм
Анаболизм
и катаболизм – это основные метаболические
процессы.
Катаболизм
– это ферментативное расщепление
сложных органических соединений,
осуществляющееся внутри клетки за счет
реакций окисления. Катаболизм
сопровождается выделением энергии и
запасанием ее в макроэргических фосфатных
связях АТФ.
Анаболизм
– это синтез сложных органических
соединений – белков, нуклеиновых кислот,
полисахаридов – из простых предшественников,
поступающих в клетку из окружающей
среды или образующихся в процессе
катаболизма. Процессы синтеза связаны
с потреблением свободной энергии,
которая поставляется АТФ (рис. 31).
Рис.
31 Схема путей метаболизма в бактериальной
клетке
В
зависимости от биохимии процесса
диссимиляции (катаболизма) различают
дыхание и брожение.
Дыхание– это сложный процесс биологического
окисления различных соединений),
сопряженный с образованием большого
количества энергии, аккумулируемой в
виде макроэргических связей в структуре
АТФ (аденозинтрифосфат), УТФ (уридинтрифосфат)
и т.д., и образованием углекислого газа
и воды. Различают аэробное и анаэробное
дыхание.
Брожение– неполный распад органических соединений
с образованием незначительного количества
энергии и продуктов, богатых энергией.
Анаболизм
включает процессы синтеза, при которых
используется энергия, вырабатываемая
в процессе катаболизма. В живой клетке
одновременно и непрерывно протекают
процессы катаболизма и анаболизма.
Многие реакции и промежуточные продукты
являются для них общими.
Живые
организмы классифицируют в соответствии
с тем, какой источник энергии или углерода
они используют. Углерод – основной
элемент живой материи. В конструктивном
метаболизме ему принадлежит ведущая
роль.
В
зависимости от источника клеточного
углерода все организмы, включая
прокариотные, делят на автотрофы и
гетеротрофы.
Автотрофыиспользуют CO2 в качестве единственного
источника углерода, восстанавливая его
водородом, который отщепляется от воды
или другого вещества. Органические
вещества они синтезируют из простых
неорганических соединений в процессе
фото- или хемосинтеза.
Гетеротрофы получают углерод из органических
соединений.
Живые
организмы могут использовать световую
или химическую энергию. Организмы,
живущие за счет энергии света, называют фототрофными.Органические вещества
они синтезируют, поглощая электромагнитное
излучение Солнца (свет). К ним относятся
растения, сине-зеленые водоросли, зеленые
и пурпурные серобактерии.
Организмы,
получающие энергию из субстратов,
источников питания (энергия окисления
неорганических веществ), называют хемотрофами.Кхемогетеротрофамотносятся большинство бактерий, а так
же грибы и животные.
Существует
немногочисленная группа хемоавтотрофов.
К таким хемосинтезирующим микроорганизмам
относятся нитрифицирующие бактерии,
которые, окисляя аммиак до азотистой
кислоты, высвобождают необходимую для
синтеза энергию. К хемосинтетикам
относятся также водородные бактерии,
получающие энергию в процессе окисления
молекулярного водорода.
Углеводы как источник энергии
У
большинства организмов расщепление
органических веществ происходит в
присутствии кислорода – аэробный обмен.
В результате такого обмена остаются
бедные энергией конечные продукты (СО2и Н2О), но высвобождается много
энергии. Процесс аэробного обмена
называется дыханием, анаэробного –
брожением.
Углеводы
– основной энергетический материал,
который клетки используют в первую
очередь для получения химической
энергии. Кроме того, при дыхании могут
использоваться также белки и жиры, а
при брожении – спирты и органические
кислоты.
Расщепление
углеводов организмы осуществляют
разными путями, в которых важнейшим
промежуточным продуктом является
пировиноградная кислота (пируват).
Пируват занимает центральное место в
метаболизме при дыхании и брожении.
Выделяют три основных механизма
образования ПВК.
1.Фруктозодифосфатный (гликолиз) или
путь Эмбдена-Мейергофа-Парнаса–
универсальный путь.
Процесс
начинается с фосфорилирования (рис.
32). При участии фермента гексокиназы и
АТФ глюкоза фосфорилируется по шестому
углеродному атому с образованием
глюкозо-6-фосфата. Это активная форма
глюкозы. Она служит исходным продуктом
при расщеплении углеводов любым из трех
путей.
При
гликолизе глюкозо-6-фосфат изомеризуется
во фруктозо-6-фосфат, а затем под действием
6-фосфофруктокиназы фосфорилируется
по первому углеродному атому. Образовавшийся
фруктозо-1,6-дифосфат под действием
фермента альдолазы легко распадается
на две триозы: фосфоглицериновый альдегид
и дигидроксиацетонфосфат. Дальнейшее
превращение С3-углеводов
осуществляется за счет переноса водорода
и фосфорных остатков через ряд органических
кислот с участием специфических
дегидрогеназ. Все реакции этого пути,
за исключением трех, протекающих с
участием гексокиназы, 6-фосфофруктокиназы
и пируваткиназы, полностью обратимы.
На стадии образования пировиноградной
кислоты заканчивается анаэробная фаза
превращения углеводов.
Баланс:
Максимальное
количество энергии, получаемое клеткой
при окислении одной молекулы углеводов
гликолитическим путем, равно 2·105Дж.
Рис.32.
Фруктозодифосфатный путь расщепления
глюкозы
2.Пентозофосфатный (Варбурга-Дикенса-Хорекера)путь характерен также для большинства
организмов (в большей степени для
растений, а для микроорганизмов играет
вспомогательную роль). В отличие от
гликолиза ПФ путь не образует пируват.
Глюкозо-6-фосфат
превращается в 6-фосфоглюколактон,
который декарбоксилируется (рис. 33). При
этом образуется рибулозо-5-фосфат, на
котором завершается процесс окисления.
Последующие реакции рассматриваются
как процессы превращения пентозофосфатов
в гексозофосфаты и обратно, т.е. образуется
цикл. Считают, что пентозофосфатный
путь на одном из этапов переходит в
гликолиз.
При
прохождении через ПФ путь каждых шести
молекул глюкозы происходит полное
окисление одной молекулы глюкозо-6-фосфата
до CO2и восстановление 6 молекул
НАДФ+до НАДФ·Н2. Как механизм
получения энергии этот путь в два раза
менее эффективен, чем гликолитический:
на каждую молекулу глюкозы образуется
1 молекула АТФ.
Рис.
33. Пентозофосфатный путь расщепления
глюкозо-6-фосфата
Основное
назначение этого пути – поставлять
пентозы, необходимые для синтеза
нуклеиновых кислот, и обеспечивать
образование большей части НАДФ·Н2,
необходимого для синтеза жирных кислот,
стероидов.
3.Путь Энтнера-Дудорова
(кетодезоксифосфоглюконатный или
КДФГ-путь)встречается только у
бактерий. Глюкоза фосфорилируется
молекулой АТФ при участии фермента
гексокиназы (рис. 34).
Рис.34.
Путь Энтнера-Дудорова расщепления
глюкозы
Продукт
фосфорилирования – глюкозо-6-фосфат –
дегидрируется до 6-фосфоглюконата. Под
действием фермента фосфоглюконатдегидрогеназы
от него отщепляется вода и образуется
2-кето-3-дезокси-6-фосфоглюконат (КДФГ).
Последний расщепляется специфичной
альдолазой на пируват и глицеральдегид-3-фосфат.
Глицеральдегид далее подвергается
действию ферментов гликолитического
пути и трансформируется во вторую
молекулу пирувата. Кроме того, этот путь
поставляет клетке 1 молекулу АТФ и 2
молекулы НАД·Н2.
Таким
образом, основным промежуточным продуктом
окислительного расщепления углеводов
является пировиноградная кислота,
которая при участии ферментов превращается
в различные вещества. Образовавшаяся
одним из путей ПВК в клетке подвергается
дальнейшему окислению. Освобождающиеся
углерод и водород удаляются из клетки.
Углерод выделяется в форме CO2,
водород передается на различные
акцепторы. Причем может передаваться
либо ион водорода, либо электрон, поэтому
перенос водорода равноценен переносу
электрона. В зависимости от конечного
акцептора водорода (электрона) различают
аэробное дыхание, анаэробное дыхание
и брожение.
Дыхание
Дыхание
– окислительно-восстановительный
процесс, идущий с образованием АТФ; роль
доноров водорода (электронов) в нем
играют органические или неорганические
соединения, акцепторами водорода
(электронов) в большинстве случаев
служат неорганические соединения.
Если
конечный акцептор электронов –
молекулярный кислород, дыхательный
процесс называют аэробным
дыханием.
У некоторых микроорганизмов конечным
акцептором электронов служат такие
соединения, как нитраты, сульфаты и
карбонаты. Этот процесс называется анаэробным
дыханием.
Аэробное
дыхание – процесс полного окисления субстратов
до CO2 и
Н2О
с образованием большого количества
энергии в форме АТФ.
Полное
окисление пировиноградной кислоты
происходит в аэробных условиях в цикле
трикарбоновых кислот (ЦТК или цикл
Кребса) и дыхательной цепи.
Аэробное
дыхание состоит из двух фаз:
1).
Образующийся в процессе гликолиза
пируват окисляется до ацетил-КоА, а
затем до CO2,
а освобождающиеся атомы водорода
перемещаются к акцепторам. Так
осуществляется ЦТК.
2).
Атомы водорода, отщепленные дегидрогеназами,
акцептируются коферментами анаэробных
и аэробных дегидрогеназ. Затем они
переносятся по дыхательной цепи, на
отдельных участках которой образуется
значительное количество свободной
энергии в виде высокоэнергетических
фосфатов.
Цикл
трикарбоновых кислот (цикл Кребса, ЦТК)
Пируват,
образующийся в процессе гликолиза, при
участии мультиферментного комплекса
пируватдегидрогеназы декарбоксилируется
до ацетальдегида. Ацетальдегид, соединяясь
с коферментом одного из окислительных
ферментов – коферментом А (КоА-SH),
образует «активированную уксусную
кислоту» — ацетил-КоА – высокоэнергетическое
соединение.
Ацетил-КоА
под действием цитрат-синтетазы вступает
в реакцию со щавелевоуксусной кислотой
(оксалоацетат), образуя лимонную кислоту
(цитрат С6),
которая является основным звеном ЦТК
(рис. 35). Цитрат после изомеризации
превращается в изоцитрат. Затем следует
окислительное (отщепление Н)
декарбоксилирование (отщепление СО2)
изоцитрата, продуктом которого является
2-оксоглутарат (С5).
Под влиянием ферментного комплекса
ɑ-кетоглутаратдегидрогеназы с активной
группой НАД он превращается в сукцинат,
теряя СО2 и два атома водорода. Сукцинат затем
окисляется в фумарат (С4),
а последний гидратируется (присоединение
Н2О)
в малат. В завершающей цикл Кребса
реакции происходит окисление малата,
что приводит к регенерации оксалоацетата
(С4). Оксалоацетат взаимодействует с
ацетил-КоА, и цикл повторяется снова.
Каждая из 10 реакций ЦТК, за исключением
одной, легко обратима. В цикл вступают
два атома углерода в виде ацетил-КоА и
такое же количество атомов углерода
покидают этот цикл в виде СО2.
В
результате четырех окислительно-восстановительных
реакций цикла Кребса осуществляется
перенос трех пар электронов на НАД и
одной пары электронов на ФАД. Восстановленные
таким путем переносчики электронов НАД
и ФАД подвергаются затем окислению уже
в цепи переноса электронов. В цикле
образуется одна молекула АТФ, 2 молекулы
СО2 и 8 атомов водорода.
Биологическое
значение цикла Кребса заключается в
том, что он является мощным поставщиком
энергии и «строительных блоков» для
биосинтетических процессов. Цикл Кребса
действует только в аэробных условиях,
в анаэробных он разомкнут на уровне
α-кетоглутаратдегидрогеназы.
Дыхательная
цепь
Последней
стадией катаболизма является окислительное
фосфорилирование. В ходе этого процесса
высвобождается большая часть метаболической
энергии.
Восстановленные
в цикле Кребса переносчики электронов
НАД и ФАД подвергаются окислению в
дыхательной цепи или цепи транспорта
электронов. Молекулы-переносчики – это
дегидрогеназы, хиноны и цитохромы.
Обе
ферментные системы у прокариот находятся
в плазматической мембране, а у эукариот
– во внутренней мембране митохондрий.
Электроны от атомов водорода (НАД, ФАД)
по сложной цепи переносчиков переходят
к молекулярному кислороду, восстанавливая
его, при этом образуется вода.
Баланс. Расчеты энергетического баланса
показали, что при расщеплении глюкозы
гликолитическим путем и через цикл
Кребса с последующим окислением в
дыхательной цепи до СО2 и Н2О
на каждую молекулу глюкозы образуется
38 молекул АТФ. Причем максимальное
количество АТФ образуется в дыхательной
цепи – 34 молекулы, 2 молекулы — в ЭМП-пути
и 2 молекулы – в ЦТК (рис. 36).
Рис.
36. Схема ассимиляции глюкозы при
аэробном дыхании
Неполное
окисление органических соединений
Дыхание
обычно связано с полным окислением
органического субстрата, т.е. конечными
продуктами распада являются СО2 и Н2О.
Однако
некоторые бактерии и ряд грибов не до
конца окисляют углеводы. Конечными
продуктами неполного окисления являются
органические кислоты: уксусная, лимонная,
фумаровая, глюконовая и др., которые
аккумулируются в среде. Этот окислительный
процесс используется микроорганизмами
для получения энергии. Однако общий
выход энергии при этом значительно
меньший, чем при полном окислении. Часть
энергии окисляемого исходного субстрата
сохраняется в образующихся органических
кислотах.
Микроорганизмы,
развивающиеся за счет энергии неполного
окисления, используются в микробиологической
промышленности для получения органических
кислот и аминокислот.
Урок 24. энергетика живой клетки — Естествознание — 10 класс
Метаболизм (обмен веществ) – сложная цепь превращений веществ в организме начиная с момента их поступления из внешней среды и кончая удалением продуктов распада. Представляет собой совокупность процессов катаболизма и анаболизма.
Катаболизм (диссимиляция), совокупность ферментативных реакций в живом организме, направленных на расщепление сложных органических соединений до более простых.
Анаболизм (ассимиляция), биохимические реакции, в результате которых из более простых веществ синтезируются более сложные, что приводит к запасанию энергии, к образованию новых материалов для построения клеток и к росту.
Энергетический обмен – это совокупность химических реакций постепенного распада органических соединений, сопровождающихся высвобождением энергии, часть которой расходуется на синтез АТФ. Синтезированная АТФ становится универсальным источником энергии для жизнедеятельности организмов. Значение энергетического обмена – снабжение клетки энергией, которая необходима для жизнедеятельности.
Пластический обмен (ассимиляция, анаболизм, биосинтез) – это когда из простых веществ с затратой энергии образуются (синтезируются) более сложные. Непосредственным поставщиком энергии в клетках выступает АТФ.
Фотосинтез – процесс образования органических веществ из неорганических (углекислого газа и воды) с использованием солнечной энергии. Проходит в два этапа: световая фаза (происходит улавливание и фиксация энергии света в АТФ) и темновая (связывание углекислого газа в молекулы глюкзы с затратой энергии АТФ).
Хемосинтез – процесс образования органических веществ из неорганических (углекислого газа и воды) с использованием энергии окисления неорганических веществ. Например такой тип питания используют азотфиксирующие бактерии.
Клеточное или тканевое дыхание – совокупность биохимических реакций, протекающих в клетках живых организмов, в ходе которых происходит окисление углеводов, липидов и аминокислот до углекислого газа и воды.
Гликолиз – многоступенчатый процесс бескислородного расщепления глюкозы (С6Н1206) до пировиноградной кислоты (С3Н403). Реакции гликолиза катализируются специальными ферментами и протекают в цитоплазме клеток.
Аэробное окисление – окисление пировиноградной кислоты в присутствии атмосферного кислорода, протекающее во много раз быстрее, чем неполный процесс анаэробного дыхания и с выделением большего количества энергии. Конечными продуктами выступают СО2 и Н2О.
Брожение – это анаэробный (происходящий без участия кислорода) метаболический распад молекул питательных веществ, например глюкозы под действием ферментов.
Катаболизм — определение и примеры
Определение катаболизма
Катаболизм — это часть метаболизма , отвечающая за разрушение сложных молекул на более мелкие. Другая часть метаболизма, анаболизм , превращает простые молекулы в более сложные. Во время катаболизма энергия высвобождается из разрушающихся связей больших молекул. Обычно эта энергия затем сохраняется в связях аденозинтрифосфата (АТФ).Катаболизм увеличивает концентрацию АТФ в клетке, поскольку он расщепляет питательные вещества и пищу. АТФ в таких высоких концентрациях с большей вероятностью откажется от своей энергии при высвобождении фосфата. Затем анаболизм использует эту энергию для объединения простых предшественников в сложные молекулы, которые добавляются к клетке и накапливают энергию для деления клетки.
Многие пути катаболизма имеют аналогичные версии в анаболизме. Например, большие молекулы жира в пище организма должны расщепляться на мелкие жирные кислоты, из которых он состоит.Затем, чтобы организм мог запасать энергию на зиму, необходимо создавать и хранить большие молекулы жира. Катаболические реакции расщепляют жиры, а анаболические пути их восстанавливают. Эти метаболические пути часто используют одни и те же ферменты. Чтобы уменьшить вероятность того, что эти пути будут препятствовать развитию друг друга, эти пути часто подавляют друг друга и у эукариот разделены на разные органеллы.
Примеры катаболизма
Углеводный и липидный катаболизм
Почти все организмы используют сахар , глюкозу в качестве источника энергии и углеродных цепей.Глюкоза хранится организмами в более крупных молекулах, называемых полисахаридами . Эти полисахариды могут быть крахмалом, гликогеном или другими простыми сахарами, такими как сахароза. Когда клеткам животного нужна энергия, они посылают сигналы тем частям тела, которые хранят глюкозу, или потребляют пищу. Глюкоза высвобождается из углеводов специальными ферментами в первой стадии катаболизма. Затем глюкоза распределяется по организму для использования другими клетками в качестве энергии. Катаболический путь , гликолиз , затем еще больше расщепляет глюкозу, высвобождая энергию, которая хранится в АТФ.Из глюкозы образуются молекулы пирувата. Дальнейшие катаболические пути создают ацетат , который является ключевой промежуточной молекулой метаболизма. Ацетат может представлять собой самые разные молекулы, от фосфолипидов до молекул пигмента, гормонов и витаминов.
Жиры, представляющие собой большие липидные молекулы, также расщепляются в процессе метаболизма с образованием энергии и других молекул. Подобно углеводам, липиды хранятся в виде больших молекул, но могут расщепляться на отдельные жирные кислоты.Эти жирные кислоты затем превращаются посредством бета-окисления в ацетат. Опять же, ацетат может использоваться анаболизмом для производства более крупных молекул или как часть цикла лимонной кислоты , который управляет дыханием и производством АТФ. Животные используют жиры для хранения большого количества энергии для использования в будущем. В отличие от крахмала и углеводов, липиды гидрофобны и исключают воду. Таким образом можно сохранить много энергии без того, чтобы тяжелый вес воды замедлял работу организма.
Большинство катаболических путей конвергентно в том смысле, что они заканчиваются одной и той же молекулой. Это позволяет организмам потреблять и накапливать энергию в различных формах, в то же время имея возможность производить все необходимые молекулы в анаболических путях. Другие катаболические пути, такие как катаболизм белков, обсуждаемый ниже, создают различные промежуточные молекулы — предшественники, известные как аминокислот , для создания новых белков.
Катаболизм белков
Все известные в мире белки состоят из одних и тех же 20 аминокислот.Это означает, что белки растений, животных и бактерий представляют собой всего лишь различные комбинации 20 аминокислот. Когда организм потребляет меньший организм, весь белок в этом организме должен перевариваться в результате катаболизма. Ферменты, известные как протеиназы , разрывают связи между аминокислотами в каждом белке, пока кислоты не будут полностью разделены. После разделения аминокислоты могут быть распределены по клеткам организма. Согласно ДНК организма, аминокислоты будут рекомбинированы в новые белки.
Если источник глюкозы отсутствует или слишком много аминокислот, молекулы вступят в дальнейшие катаболические пути и распадутся на углеродные скелеты. Эти небольшие молекулы могут быть объединены в глюконеогенезе для создания новой глюкозы, которую клетки могут использовать в качестве энергии или накапливать в больших молекулах. Во время голодания клеточные белки могут подвергаться катаболизму, позволяя организму выживать в собственных тканях, пока не будет найдено больше пищи. Таким образом, организмы могут жить с небольшим количеством воды в течение очень долгого времени.Это делает их более устойчивыми к изменяющимся условиям окружающей среды.
Анаболизм — Часть метаболизма, которая строит большие молекулы из более мелких.
Метаболизм — анаболизм и катаболизм вместе, или все ферментативные реакции в клетке.
Метаболический путь — Последовательные химические реакции, организуемые внутри клеток.
Катаболический путь — Одиночная серия реакций, которые разрушают определенную молекулу.
Викторина
1. Дрожжи — это одноклеточные организмы, используемые для производства алкоголя. В среде с низким содержанием кислорода или его отсутствием дрожжи создают спирт как побочный продукт высвобождения энергии из глюкозы. Является ли производство алкоголя частью анаболического пути, катаболического пути или ни одного из них? A. Анаболический путь B. Катаболический путь C. Ни один из
Ответ на вопрос № 1
B является правильным.Хотя алкоголь является побочным продуктом, он возникает во время катаболизма глюкозы. Как и все клетки, дрожжи должны использовать глюкозу для получения энергии. Без кислорода дрожжи развили катаболический путь, известный как ферментация , при котором энергия все еще может собираться, но без кислорода. Вместо этого спирты создаются и выбрасываются в окружающую среду. Пивоварни, виноградники и винокурни используют этот изящный прием глюкозы для создания спирта из сахаров. Из разных источников сахара получаются напитки с разными вкусами.В вине используется виноградный сахар, в пиве используется ячменный крахмал, а в других спиртных напитках используется множество различных сахаров, например, картофель в водке и рис в саке.
2. Плотоядные животные могут производить всю необходимую им глюкозу из животного белка. Всю необходимую глюкозу травоядные животные получают из растений. Почему нельзя принуждать плотоядных есть растения или заставлять травоядных есть мясо, чтобы получить энергию? A. Они не умеют. B. Они не производят необходимых ферментов. C. Могут! Всеядное животное — это просто хищник, который научился есть растения.
Ответ на вопрос № 2
B правильный. Облигатные плотоядные животные могут есть только мясо, потому что у них отсутствуют необходимые катаболические пути, разрушающие растения. Эволюция, выбирая неиспользуемые и неэффективные пути, выбирает организмы, заполняющие определенные ниши. Если эта ниша предлагает очень мало растительного материала, катаболизм изменяется, и определенные пути теряются. Таким образом, даже если вы научите плотоядное животное есть и собирать растения, его организм не сможет перерабатывать питательные вещества.Точно так же травоядное животное может получать питательные вещества только из растительного сырья. Всеядные животные эволюционировали в нише, для использования которой требуется энергия из обоих источников. У этих животных катаболизм способен переваривать оба вида пищи.
3. Бактерии, не имеющие специализированных отделов внутри своих клеток, должны регулировать анаболизм и катаболизм, чтобы работать вместе. Ученый добавляет к бактериям химическое вещество, которое отключает анаболизм, постоянно обеспечивая только катаболизм.Что будет с клеткой? A. Он умрет. Б. Будет расти. C. Он будет производить много энергии.
Ответ на вопрос № 3
A правильный. В то время как катаболизм будет производить много энергии, в конечном итоге у него закончатся молекулы для разложения, и энергия прекратится. Клетка не смогла бы расти без анаболизма, создающего новые молекулы. Таким образом, даже если клетка может давать энергию, без процесса, который восстанавливает и добавляет к клетке, она в конечном итоге развалится.И анаболизм, и катаболизм необходимы для обеспечения нормального метаболизма в организме.
Коротко- и среднецепочечные жирные кислоты в энергетическом метаболизме: клеточная перспектива
Abstract
Коротко- и среднецепочечные жирные кислоты (SCFAs и MCFAs), независимо от их клеточных сигнальных функций, являются важными субстратами энергетического метаболизма и анаболические процессы у млекопитающих. SCFAs в основном вырабатываются бактериями толстой кишки и преимущественно метаболизируются энтероцитами и печенью, тогда как MCFA возникают в основном из пищевых триглицеридов, в том числе молока и молочных продуктов.Общей чертой SCFA и MCFA является их карнитин-независимое поглощение и интрамитохондриальная активация тиоэфиров ацил-КоА. В отличие от длинноцепочечных жирных кислот, клеточный метаболизм SCFAs и MCFAs в меньшей степени зависит от белков, связывающих жирные кислоты. SCFAs и MCFAs модулируют тканевой метаболизм углеводов и липидов, что проявляется главным образом ингибирующим действием на гликолиз и стимуляцией липогенеза или глюконеогенеза. SCFAs и MCFAs не обладают протонофорной и литической активностью в митохондриях или проявляют лишь слабую их активность и не оказывают значительного влияния на транспорт электронов в дыхательной цепи.SCFAs и MCFAs модулируют выработку митохондриальной энергии двумя механизмами: они обеспечивают восстановительные эквиваленты дыхательной цепи и частично снижают эффективность окислительного синтеза АТФ.
Коротко- и среднецепочечные жирные кислоты (SCFAs и MCFAs), наряду с более многочисленными длинноцепочечными жирными кислотами (LCFAs), являются естественными соединения, присутствующие в тканях как животных, так и растений, которые участвуют в метаболизме клеток.КЦЖК и СЦЖК также являются важными составляющими пищевых продуктов, где они в основном находятся в форме триглицеридов в некоторых растительных маслах и молоке (1). Тем не менее, бактериальная ферментация амилазо-устойчивого крахмала и некрахмальных полисахаридов в кишечнике, вероятно, является наиболее важным источником SCFAs у людей и большинства видов млекопитающих (2–4).
Помимо своей роли энергоснабжающего топлива, SCFAs и MCFAs обладают различными регулирующими и сигнальными функциями. Известно, что бутират и другие SCFA вызывают апоптоз в определенных условиях и, таким образом, контролируют пролиферацию клеток (5, 6).В настоящее время все больше внимания уделяется SCFAs в связи с их предполагаемой ролью в патогенезе аллергии, а также аутоиммунных, метаболических и неврологических заболеваний [обзор в (7)]. В последние два десятилетия также была охарактеризована роль MCFA как агонистов рецепторов, активируемых пролифератором пероксисом (8). Более того, накапливающиеся данные показывают, что SCFAs, генерируемые микробиотой кишечника, оказывают влияние на потребление пищи, тем самым регулируя энергетический гомеостаз и массу тела [обзор в (9-11)].SCFAs и MCFAs также играют важную роль во внутриклеточной передаче сигналов и вносят вклад в регуляцию клеточного метаболизма [rev. (12–16)]. Наконец, MCFAs и SCFAs могут контролировать гибель и выживаемость клеток (17-20). Эти важные регуляторные функции MCFA и SCFA и их влияние на здоровье человека и патологии являются предметом ряда превосходных всесторонних обзоров (1, 7, 21, 22). Здесь мы хотим сконцентрироваться на некоторых особенностях метаболических особенностей SCFAs и MCFAs, которые отличаются от таковых LCFAs, и подвести итоги текущего понимания их роли в клеточном энергетическом метаболизме.Некоторые аспекты участия SCFA и MCFA в энергозависимых митохондриальных процессах уже были кратко рассмотрены нами ранее (23). , 25). Жирные кислоты с общим числом атомов углерода от 1 до 6 обычно классифицируются как SCFA, тогда как кислоты с числом атомов углерода от 7 до 12 определяются как MCFA.Жирные кислоты с более короткими цепями, содержащими до 9 атомов углерода, при комнатной температуре являются жидкими () (26). Запах первых членов резкий, тогда как запах более высоких членов прогорклый или отсутствует. Липофильность SCFAs и MCFAs, измеряемая как распределение свободной кислоты между водой и гептаном, постепенно увеличивается с увеличением длины цепи углеродных атомов, так что MCFAs становятся в этом аспекте сопоставимыми с LCFAs (27–30). В связи с меньшей липофильностью по сравнению с LCFAs SCFAs не образуют мицеллярных структур и не участвуют в формировании биологических мембран (31).SCFAs и MCFAs являются слабыми кислотами, pK a составляет около 4,8, за исключением муравьиной кислоты, pK a которой примерно на одну единицу ниже (). Таким образом, их соли щелочных металлов значительно гидролизуются в водных растворах. Водорастворимые члены этого семейства имеют высокую тенденцию к образованию бимолекулярных ассоциатов в водном растворе. Интересно, что включение SCFAs и MCFAs в двухслойные мембраны, как известно, увеличивает их значения pK a аналогично, как и в случае LCFAs (32, 33).
ТАБЛИЦА 1.
Номенклатура и основные физические свойства SCFA и MCFA
Число атомов углерода
Систематическое название
Общее название
Формула
Температура плавления (° C)
Температура кипения (° C) C)
pK a
1
Метановая кислота
Муравьиная кислота
HCOOH
8,6
100,8
3.75
2
Этановая кислота
Уксусная кислота
CH 3 COOH
16,5
118,1
4,75
314
Пропионовая кислота 2 COOH
−22,0
140,9
4,88
4
Бутановая кислота
Масляная кислота
CH 3 (CH 2 60 2 COO9
162,5
4,81
5
Пентановая кислота
Валериановая кислота
CH 3 (CH 2 ) 3 COOH
0 9017 90100 9017 9010 9017 901 6
Гексановая кислота
Капроновая кислота
CH 3 (CH 2 ) 4 COOH
−3,9
205
4,84
Гептановая кислота
7 CH 3 (CH 2 ) 5 COOH
−7.5
223
4,84
8
Октановая кислота
Каприловая кислота
CH 3 (CH 2 ) 6 COOH
16,3 901 901 9015
9016 23 901 901
9016 9 901 901
Нонановая кислота
Пеларгоновая кислота
CH 3 (CH 2 ) 7 COOH
12,3
254
—
Capric1
Decanoic кислота 3 (CH 2 ) 8 COOH
31.3
269
—
11
Ундекановая кислота
Ундециловая кислота
CH 3 (CH 2 ) 9 COOH
3
9017 901
9017 29,3 901
12
Додекановая кислота
Лауриновая кислота
CH 3 (CH 2 ) 10 COOH
44
225 a
У человека основным источником SCFAs является ферментация пищевых волокон и непереваренных сахаридов в кишечнике анаэробными бактериями толстой кишки (2) [обзор в (3, 4, 7)].Ацетат в основном образуется за счет восстановительного метилирования CO 2 (34). Есть два основных пути производства пропионата бактериями толстой кишки. В пути метилмалон-КоА (также называемом дикарбоксильным путем) пропионат образуется из лактата, который доставляется бактериями, ферментирующими лактат. Короче говоря, лактат поглощается пропионовыми бактериями и затем дегидрируется до пирувата, который карбоксилируется метилмалонил-КоА-карбоксилтрансферазой до оксалоацетата. Впоследствии последний превращается в пропионат по четырехуглеродному пути, состоящему из промежуточных продуктов, малата, фумарата, сукцината и метилмалонил-КоА (34).Следует отметить, что этот путь генерирует, помимо пропионата, одну молекулу ацетата на две молекулы пропионата. Другие бактерии, такие как Clostridium propionicum и Megasphaera elsdenii , легко продуцируют пропионат из лактата (35, 36). В этом случае CoA-эфир лактата (лактоил-CoA) превращается через акрилоил-CoA в пропионил-CoA, который впоследствии гидролизуется до пропионовой кислоты (акрилатный путь) (34). Бутират образуется в результате конденсации двух молекул ацетил-КоА с образованием ацетоацетил-КоА с последующим восстановительным превращением ацетоацетил-КоА в бутирил-КоА (37).По оценкам, ацетат, пропионат и бутират образуются в толстой кишке человека в соотношении примерно 3: 1: 1 (38, 39). В исследовании in vivo скорость высвобождения SCFA из кишечника в систему кровообращения составила около 35 мкмоль / кг массы тела в час (40). Самые высокие концентрации (70–140 мМ) были обнаружены в проксимальном отделе толстой кишки (2). SCFAs могут составлять примерно 10% от общего потребления калорий человеком (4).
Для новорожденных млекопитающих, включая детей, материнское молоко является важным источником MCFA и SCFA, которые присутствуют в основном в форме триглицеридов и фосфолипидов.Например, содержание СЦЖК (C 7: 0 –C 12: 0 ) составляет 6–17% и 9–28% всех жирных кислот у крупного рогатого скота (1) и человека (41, 42). молоко соответственно. Коровье молоко и молочные продукты остаются основным пищевым источником SCFAs, в основном масляной кислоты, у взрослых людей. Другими естественными источниками MCFA и SCFA являются кокосовое масло и косточковое пальмовое масло [(1) и ссылки в нем]. По сравнению с триглицеридами, содержащими LCFA, те, которые содержат MCFA, быстрее гидролизуются в кишечном тракте и не включаются в хиломикроны.SCFAs и MCFAs транспортируются портальным кровотоком в печень, где они легко метаболизируются (21).
SCFAs и MCFAs могут также образовываться в тканях млекопитающих и человека, в основном в печени. Таким образом, пероксисомное β-окисление LCFAs производит ацил-CoAs с укороченной цепью (43), которые могут гидролизоваться внутри пероксисом с помощью различных ацил-CoA тиоэстераз и высвобождаться в цитозоль. Кроме того, пероксисомы также снабжены карнитин-ацетил- и карнитин-октаноилтрансферазами, и, таким образом, укороченные ацил-КоА превращаются в эфиры карнитина для доставки в митохондрии [обзор в (44)].
При патологических условиях (например, при врожденном дефиците ацил-КоА-дегидрогеназы со средней длиной цепи) октаноат и деканоат накапливаются в значительных количествах в тканях (45), что приводит к нарушению функционирования дыхательных комплексов митохондрий (46). Эти нарушения часто сопровождаются повышенной экскрецией с мочой дикарбоновых MCFA (в основном адипиновой, субериновой и себациновой кислот), которые, по-видимому, возникают в результате микросомального ω-окисления соответствующих ацил-КоА со средней длиной цепи (47, 48).Основываясь на их быстром всасывании, триглицериды СЦЖК были введены в качестве быстро доступного источника энергии в лечебное питание в середине прошлого века. Эмульсии, обогащенные триглицеридами, содержащими MCFA, применяли у пациентов, страдающих различными формами нарушения переваривания нормальных триглицеридов, содержащих LCFA (1). Кроме того, из-за быстрого транспорта MCFAs из кишечника в печень были разработаны дыхательные тесты для неинвазивной клинической диагностики с использованием 13 C-меченного октаноата (49, 50).Таким образом, можно измерить специфические печеночные пути, а также скорость опорожнения желудка [см. Обзор (51)].
ПРИНЦИПЫ МЕТАБОЛИЗМА В ТКАНЯХ ЖИВОТНЫХ
Транспорт из кишечника в печень
Бутират поглощается энтероцитами, предположительно с помощью переносчика монокарбоксилата 1 (MCT-1) и связанного с натрием переносчика монокарбоксилата 1 (SMCT- 1) [обзор в (52, 53)]. Эти элементы используют бутират в основном в качестве топлива. По мнению других авторов, SCFAs также абсорбируются из просвета кишечника путем обмена с Cl — (54) и / или HCO 3 — (55).Однако в последнее время стала благоприятствовать неионная диффузия протонированных SCFAs и MCFAs через эпителий толстой кишки (56). Последний механизм также подтверждается исследованиями на модельных системах (30). Согласно этому механизму всасывание SCFAs в кишечнике зависит от pH, в то время как небольшое закисление pH в просвете, возможно, за счет метаболической активности бактерий, увеличивает преобладание протонированной формы SCFAs. В противном случае транспорт SCFAs из энтероцитов в кровь может быть вызван анионным обменом.Таким образом, кажется вероятным, что транспорт через базолатеральную мембрану основан на анионной форме SCFAs против HCO 3 — (57). Бутират, а также другие SCFA и MCFA, которые не используются энтероцитами, транспортируются портальной веной в печень (40, 58) и метаболизируются гепатоцитами.
В отличие от LCFA, которые этерифицируются до триглицеридов в энтероцитах, включаются в хиломикроны, а затем попадают в лимфатическую систему, SCFA и MCFA из кишечного тракта попадают в воротную вену в виде свободных кислот.Здесь MCFAs частично связываются с альбумином плазмы. Соотношение между связанными с альбумином и свободными MCFA увеличивается с увеличением длины цепи, так что первая константа равновесия (т. Е. Для самого сильного сайта связывания) между связанной с альбумином и свободной формами увеличивается с 1,5 × 10 4 для гексаноата через 3,4 × 10 4 для октаноата и 10 5 для деканоата, до 2,4 × 10 6 для лауреата (59, 60). Следует помнить, что ДЦЖК присутствуют в циркулирующей крови практически полностью связанными с альбумином плазмы, с первой константой равновесия порядка от 10 7 до 10 9 (60).Последующее поглощение SCFA и MCFA, по крайней мере, низших членов этой группы, печенью и мышечными клетками, а также другими тканями, не зависит от белков, связывающих жирные кислоты (1). Точно так же для их поглощения клетками не требуются ни транспортные белки жирных кислот, ни транслоказа жирных кислот, встроенная в плазматическую мембрану, ни цитозольные связывающие жирные кислоты белки. Эти наблюдения дают вероятное объяснение того, почему окисление октаноата изолированными гепатоцитами происходит примерно в пять раз быстрее, чем окисление олеата (61).Более того, внутриклеточный метаболизм SCFAs и MCFAs, по-видимому, не требует или требует гораздо меньшего количества белков, связывающих жирные кислоты (62, 63). Напротив, LCFAs нуждаются в белках, связывающих жирные кислоты, для их клеточного поглощения, внутриклеточного транспорта, регуляторных функций и метаболизма [обзор в (63)]. Связывание свободных LCFA или сложных эфиров LCFA-CoA со связывающими жирными кислотами белками также сводит к минимуму их токсические эффекты, такие как литические свойства или ингибирование ферментов (63). Интересно, что у крыс, дефицитных по одному из белков транспорта жирных кислот (белок CD36), кормление диетой, богатой SCFA и MCFA, устраняет повышенное поглощение глюкозы, гиперинсулинемию и гипертрофию сердца (64).Точно так же у мышей с дефицитом CD36 октаноат смягчал плохую ишемическую толерантность сердца (65). Эти наблюдения могут иметь важное значение для медицины человека.
Обеспечение энергетического метаболизма тканей
Использование различными тканями ацетата, образованного кишечными бактериями, сильно различается у жвачных и нежвачных животных [(4) и ссылки в нем]. Ацетат также вырабатывается эндогенно у взрослых людей в результате окисления этанола, которое действует в основном в печени (66).Таким образом, было показано, что окисление этанола может приводить к 20-кратному увеличению уровня ацетата в периферической крови (67). Кроме того, чистое образование ацетата было обнаружено при окислении жирных кислот в перфузированной печени крыс (68). Образовавшийся ацетат в основном является результатом действия ацетил-КоА-гидролазы (ацетил-КоА-деацилазы), которая в печени крыс обнаруживается преимущественно в митохондриальном матриксе (69, 70). Поскольку гидролаза ацетил-КоА ингибируется свободным CoASH ( K i = 17 мкМ), уровень свободного CoASH должен быть сильно снижен, прежде чем гидролаза сможет производить свободный ацетат из ацетил-КоА (70).С другой стороны, свободный ацетат, образующийся в печени при окислении этанола или как побочный продукт кетогенеза, почти не окисляется в этом органе. Это связано с высоким значением K m митохондриальной ацетил-КоА синтетазы печени (71) или ее отсутствием в этих органеллах [(72), см. Также следующий абзац]. Однако ацетат может переноситься циркуляцией в другие органы, например, сердце и скелетные мышцы, где K m митохондриальной ацетил-КоА-синтетазы намного ниже и где он может использоваться в качестве топлива (70).
Активация SCFAs происходит в печени и некоторых других тканях с помощью ацил-CoA синтетаз (72). Эти ферменты расположены в цитозоле, а также в митохондриальном матриксе, где они слабо связаны с внутренней митохондриальной мембраной. У млекопитающих ацетат активируется до ацетил-КоА двумя различными ацетил-КоА-синтетазами, одна из которых (AceCS1) является цитозольной (78 кДа, K m = 0,11 мМ), а другая (AceCS2) митохондриальной ( 71 кДа, К м = 0.06) (73). По мнению этих авторов (73), AceCS2 присутствует в широком диапазоне тканей, с самым высоким уровнем в сердце (бык и грызуны), но практически отсутствует в печени. Напротив, более ранние исследования (74–76) продемонстрировали наличие активности ацетил-КоА синтетазы в гепатоцитах как в митохондриях (20–50%), так и в цитозоле (50–80%).
Ацетат является важным топливом во время голодания, о чем свидетельствует наблюдение, что в скелетных мышцах мышей, лишенных AceCS2, содержание АТФ снизилось до 50% по сравнению с мышами дикого типа (77).Интересно, что активность цитозольных и митохондриальных ацетил-КоА синтетаз регулируется обратимым ацетилированием. Более того, этот процесс находится под контролем NAD + -зависимых деацетилаз, сиртуина 1 и сиртуина 3 [обзор в (78)]. Сиртуин 1 является ядерным и цитоплазматическим ферментом, тогда как сиртуин 3 преимущественно находится в митохондриях. Таким образом, у млекопитающих ацетат играет важную роль не только в энергетическом гомеостазе, но и как субстрат для сиртуинов; он также участвует в регуляции сайленсинга генов и процессов старения (78).
В отличие от LCFA, которые активируются до ацил-CoA в цитозоле и должны переноситься внутрь митохондрий через карнитиновый челнок, SCFA и MCFA, по крайней мере, с числом атомов углерода до C 8 , проникают внутрь митохондрий. митохондриальные мембраны в неэтерифицированной форме и активируются до их КоА-производных в митохондриальном матриксе. Локализация среднецепочечной ацил-КоА синтетазы в митохондриальном матриксе была впервые описана в конце 60-х годов прошлого века (79) [более свежие сообщения см. (80, 81)].Последнее свойство может иметь важные метаболические последствия при определенных условиях, о чем будет сказано ниже. SCFAs и MCFAs, активированные внутри митохондрий, используются в качестве субстратов в митохондриальном β-окислении и цикле лимонной кислоты. Интересно, что с помощью метода перфузии было продемонстрировано, что в печени и сердце крыс октаноат также может подвергаться пероксисомному β-окислению, тем самым доставляя ацетил-КоА в цитозоль (82, 83).
В качестве источника энергии для тканевого метаболизма триглицериды MCFA имеют ряд преимуществ по сравнению с LCFA.Во-первых, они быстрее перевариваются, а высвободившиеся СЦЖК быстрее всасываются в просвете кишечника (21, 84). Во-вторых, тканевой метаболизм SCFAs и частично MCFAs не зависит от белков для связывания, транспорта и трансмембранной транслокации (см. Раздел «Транспорт из кишечника в печень» выше). Следовательно, они могут служить топливом, обеспечивающим больше энергии, чем ДЦЖК, особенно при патологических состояниях, примером которых является сильное воспаление (85). Наконец, MCFA, имеющие немного более низкое энергосодержание, чем LCFA (8.4 вместо 9,2 ккал / г), уменьшают жировую массу тела и повышают чувствительность тканей к инсулину [см. Обзор (1, 21, 22, 62)].
Как было сказано ранее, SCFA и MCFA переносятся кровью из пищеварительного тракта в печень, где они метаболизируются; следовательно, они не хранятся в жировой ткани. Тем не менее, при длительном кормлении крыс с портокавальными анастомозами (кровообращение, превышающее печень), группе van Itallie (86) удалось значительно обогатить липиды тканевых депо триглицеридами, содержащими высшие MCFA (C 8 , C 9 , и C 10 ).Недавно были обобщены общие особенности метаболизма в организме и физиологические функции СЦЖК, в частности октаноата, самого распространенного СЦЖК (87, 88). Подобно другим MCFA и в отличие от LCFA, октаноат быстро разлагается и хранится в виде триглицеридов в жировой или других тканях только в очень низкой степени. Октаноат, как топливо для энергетического метаболизма у млекопитающих, был изучен в тканях, требующих много энергии, таких как скелетные мышцы, сердце, печень и мозг (89–94).Что касается мозга, важно помнить, что SCFA и MCFA способны проникать через гематоэнцефалический барьер (95).
Действие SCFAs и MCFAs на метаболизм энергии в печени изучали в основном либо методом перфузии изолированной печени крысы (89, 91–93, 96–98), либо с помощью экспериментов по инкубации с изолированными гепатоцитами (99–101). Таким образом, эти исследования показали, что добавление октаноата и, в меньшей степени, бутирата увеличивает потребление кислорода по сравнению с инкубациями с буфером Кребса-Хенселейта, дополненным пируватом или лактатом в качестве субстратов, обеспечивающих энергию.Как стимуляция потребления кислорода, так и связанное с этим повышение клеточного уровня NAD (P) H (102) указывают на то, что эти жирные кислоты эффективно поставляют восстанавливающие эквиваленты (NADH, FADH 2 ) в дыхательную цепь митохондрий. Кроме того, октаноат повышал активность митохондрий, это наблюдение основано на измерении in situ потенциала митохондриальной мембраны (ΔΨ m ) (101). Также проявлялась активизация октаноатом гепатоцитов, окисляющих пируват плюс лактат (100).Однако, в отличие от LCFA (например, олеата), октаноат значительно повышает уровень AMP в ткани (89, 100). Также сообщалось, что кормление крыс диетой, богатой MCFA, увеличивает окислительную способность митохондрий скелетных мышц (62, 103), что частично объясняется повышенной активностью цитрат-синтазы (62).
Поскольку триглицериды, содержащие MCFA, быстро перевариваются в кишечнике и поглощаются энтероцитами и не включаются в хиломикроны, они являются идеальными питательными веществами, доставляющими энергию, в клинических ситуациях, когда переваривание и / или всасывание триглицеридов, содержащих LCFA, затруднено. нарушение или быстрое поглощение энергии организмом.По этой причине триглицериды, содержащие MCFA, используются для питания пациентов с наследственными нарушениями β-окисления LCFA (104). Хотя все больше данных указывает на то, что больное сердце страдает от дефицита энергии, подпитка энергетического метаболизма миокарда с помощью MCFA предлагается в качестве метаболической терапии для лечения пациентов, страдающих определенными кардиомиопатиями (87). Для этой обработки MCFA с нечетным числом атомов углерода оказались лучше, чем с четным числом атомов углерода (105).
Модуляция углеводного и липидного обмена
В отличие от ДЦЖК, на окисление СЦЖК не влияет содержание углеводов в рационе. Таким образом, сообщалось, что адаптация взрослых крыс к диете с низким содержанием жиров и высоким содержанием углеводов или с высоким содержанием жиров и низким содержанием углеводов не изменяет скорость окисления октаноата, измеренную в гепатоцитах. Напротив, окисление олеата снизилось на 50% у крыс, адаптированных к диете с низким содержанием жиров и высоким содержанием углеводов (61). Кроме того, MCFA, полученные в результате переваривания триглицеридов, содержащих MCFA, в основном разлагаются за счет β-окисления митохондрий в печени.Избыток образовавшегося ацетил-КоА используется для синтеза кетоновых тел (в основном ацетоацетата и β-гидроксибутирата), которые доставляются в качестве топлива в непеченочные ткани (21, 22). Триглицериды, содержащие MCFA, предпочтительно гидролизуются по сравнению с триглицеридами, содержащими LCFA, а высвободившиеся MCFA также предпочтительно окисляются в органах, в основном в сердце, мышцах, почках и печени (93, 106). Исследования in vitro на изолированных гепатоцитах и перфузируемой печени крысы показали, что SCFA и MCFA модулируют метаболизм углеводов и липидов в печени.Таким образом, бутират и октаноат ингибируют гликолиз (107, 108) и тем самым проявляют «глюкозосберегающую активность». Напротив, стимулируются анаболические пути образования глюкозы (100, 109–111) и липогенеза (107, 112). В качестве иллюстрации, образование глюкозы гепатоцитами, получавшими пируват плюс лактат в качестве глюконеогенных предшественников, примерно в 2 раза стимулируется октаноатом (100). Было обнаружено, что в отличие от ингибирования гликолиза в гепатоцитах деканоат, но не октаноат, стимулирует гликолиз в астроцитах, что приводит к усиленному высвобождению лактата во внеклеточное пространство (113).Поскольку лактат считается ключевым источником энергии для нейронов, лактатный челнок астроцитов / нейронов поставляет этот субстрат соседним нейронам.
Как правило, обсуждалось, что ферменты митохондриального матрикса, пируваткарбоксилаза и пируватдегидрогеназный комплекс регулируются соотношениями ацетил-КоА / КоА, АТФ / АДФ и НАДН / НАД + и, кроме того, концентрация пирувата. С другой стороны, однако, вызванное жирными кислотами (октаноат, пальмитат) увеличение потока пирувата через оба фермента объясняется исключительно повышенным захватом пирувата в компартмент митохондриального матрикса (109, 111).Утверждалось, что образование ацетоацетата из жирных кислот стимулирует захват пирувата через внутреннюю мембрану митохондрий. Следовательно, есть основания предполагать, что SCFA и MCFA играют вспомогательную роль в использовании физиологических низких концентраций пирувата или лактата для выработки глюкозы (104). Тем не менее ускорения поглощения пирувата недостаточно для объяснения огромной стимуляции жирными кислотами образования глюкозы с аспартатом и глицерином в качестве глюконеогенных предшественников (110).Такую стимуляцию обычно связывают с образованием ацетил-КоА (аллостерический эффектор пируваткарбоксилазы) и его восстанавливающих эквивалентов (114), последний способствует образованию глицеральдегид-3-фосфата. Также стоит помнить, что карбоксилирование пирувата в изолированных митохондриях печени крысы сильно стимулируется L-октаноилкарнитином, тогда как неэтерифицированный бутират и октаноат оказывают сильное ингибирование (100). Кроме того, октаноат оказывает краткосрочное двойное регулирующее действие на синтез жирных кислот в печени, а именно стимуляцию в диапазоне низких концентраций (до 1 мМ) и ингибирование в более высоких концентрациях (107, 112).Стимуляцию липогенеза связывают с активацией ацетил-КоА-карбоксилазы, предположительно путем ковалентной модификации фермента. Более того, исследования гепатоцитов, проницаемых для дигитонина, показали, что стимуляция активности ацетил-КоА-карбоксилазы зависит от длины цепи жирной кислоты (112). Величина стимуляции увеличивалась от капроновой (C 6: 0 ) до лауриновой (C 12: 0 ) кислот, но снижалась с жирными кислотами с более длинной цепью. Малонил-КоА, продукт цитозольной реакции ацетил-КоА-карбоксилазы, действует как субстрат для синтеза жирных кислот, но также как ингибитор карнитин-пальмитоилтрансферазы I.
Продолжается дискуссия о том, что SCFAs и MCFAs активируют печеночную AMP-зависимую киназу (AMPK) [обзор в (115) и ссылки там]. Как правило, активация AMPK ингибирует процессы утилизации АТФ в клетке и стимулирует те, которые производят АТФ. Будучи цитозольным ферментом, AMPK активируется повышением уровня цитозольного AMP. Следовательно, механизм, лежащий в основе активации AMPK SCFA / MCFA, не ясен, потому что активация SCFAs и MCFAs их сложными эфирами ацил-CoA повышает внутримитохондриальный уровень AMP.Интересно, что недавнее исследование мышиных миотрубок L6 предполагает, что AMPK также может активироваться без изменения соотношения цитозольного AMP / ATP. Согласно этому предположению, механизм активации AMPK MCFAs опосредуется внеклеточной Ca 2+ -зависимой Ca 2+ / кальмодулин-зависимой киназой киназы β (116). Другими сообщениями о влиянии SCFA и MCFA на анаболические пути являются ингибирование синтеза триглицеридов в адипоцитах (117) и сдерживающий эффект на накопление гликогена в печени (118).Последняя активность объясняется конкуренцией между жирными кислотами и окислением глюкозы.
Тот факт, что MCFA с углеводородным скелетом с нечетной и четной цепью по-разному влияют на энергетический метаболизм клетки, представляет особый интерес и практическое значение (119, 120). В отличие от MCFA с четной цепью, β-окисление MCFA с нечетной цепью генерирует ацетил-КоА и, кроме того, пропионил-КоА, который анаплеротичен для цикла лимонной кислоты. Пропионил-КоА может войти в цикл лимонной кислоты после превращения в сукцинат.Анаплеротическая функция MCFA с нечетной цепью, по-видимому, имеет решающее значение для поддержания уровня метаболитов цикла лимонной кислоты в различных тканях. Этот биохимический фон объясняет предлагаемое использование тригептаноина, полученного из MCFA (тригептаноата глицерина), в качестве анаплеротического препарата (121, 122) для лечения кардиомиопатий при нарушениях окисления длинноцепочечных жиров (105) и дефиците пируваткарбоксилазы (123). Эта анаплеротическая функция MCFA с нечетной цепью также важна во время эпизодов эпилепсии, когда нейроны становятся чрезмерно возбужденными и тем самым выделяют повышенное количество глутамата (124, 125).Предполагается, что высвобождение глутамата, вероятно, снижает уровень метаболитов цикла лимонной кислоты и тем самым снижает окисление ацетил-КоА митохондриями. Действительно, недавно было показано, что тригептаноин частично восстанавливает уровень метаболитов цикла лимонной кислоты в модели эпилепсии на животных (126). Тригептаноин также способен ослаблять вредные побочные эффекты, связанные с ишемическим инсультом (127). В качестве иллюстрации, когда мыши подвергались временной ишемии, тригептаноин снижал внеклеточный уровень глутамата, высвобождаемого в полосатом теле мыши, поддерживал содержание клеточного АТФ на желаемом уровне и предотвращал снижение дыхательной активности изолированных митохондрий головного мозга.Последние данные убедительно свидетельствуют о том, что регенерация митохондриального АТФ является мишенью действия тригептаноина (127). Также стоит упомянуть, что, в отличие от бутирата и октаноата, SCFA с нечетной цепью, пропионат, не оказывает ингибирующего действия на гликолиз и не стимулирует кетогенез (108). Подобно пропионил-КоА, образующемуся в результате клеточной деградации жирных кислот с нечетной цепью, внешний пропионат обеспечивает глюконеогенный путь своим углеводородным скелетом, активность, которая в основном наблюдается у жвачных животных [обзор в (128)].
ПОТЕНЦИАЛЬНЫЕ ПОБОЧНЫЕ ЭФФЕКТЫ
Энергетическое взаимодействие
LCFAs давно известны как мягкие разобщители окислительного фосфорилирования из-за их протонофорного действия на внутреннюю митохондриальную мембрану [см. Обзор (23)]. Механизм этого эффекта был всесторонне объяснен Скулачевым (129, 130), который показал, что анионы LCFA могут переноситься через внутреннюю митохондриальную мембрану с помощью транслоказы аденин-нуклеотидов, тогда как недиссоциированные молекулы жирных кислот могут перемещаться через мембрану путем переворота. флоп-механизм.В результате происходит чистый перенос протона. Последующие исследования показали, что анионы LCFA могут также переноситься через внутреннюю мембрану митохондрий с помощью ряда других переносчиков анионов внутренней мембраны митохондрий [обзор в (131, 132)]. Этот протонофорный эффект снижает электрохимический протонный градиент через внутреннюю мембрану, тем самым снижая эффективность окислительного фосфорилирования. О такой активности LCFAs неоднократно сообщалось in vitro с изолированными митохондриями, и есть доказательства того, что протонофорное разобщение также может действовать in vivo после гипоксии / реперфузии или диеты с высоким содержанием жиров (133, 134).В случае изолированных митохондрий разобщение LCFAs может быть легко определено количественно как увеличение дыхания в состоянии покоя за счет микромолярных концентраций этих кислот. В отличие от LCFA, способность стимулировать дыхание в состоянии покоя с помощью SCFA или MCFA либо чувствительна к олигомицину (от C 4 до C 8 ), либо меньше, даже при применении в миллимолярных концентрациях (100, 132, 135, 136 ). Кроме того, добавление октаноата или деканоата (в концентрации 100 или 300 мкМ) к культивируемым нейронам или астроцитам не стимулировало их дыхание (113).
Основываясь на гипотезе цикла жирных кислот, это различие между LCFAs с одной стороны и MCFAs и SCFAs с другой стороны может быть обсуждено в двух аспектах, а именно с точки зрения: i ) различной скорости проникновения жирных кислот через митохондрии. внутренняя мембрана; и ii ) различное сродство анионов жирных кислот к митохондриальным анионным носителям в зависимости от длины цепи жирной кислоты. В этом контексте можно ожидать, что SCFA и MCFA проявляют более низкую растворимость в липидном ядре внутренней митохондриальной мембраны из-за их более низкой липофильности.Фактически, было показано (30), что проникновение жирных кислот через бислои фосфатидилэтаноламина зависит от их коэффициента распределения между гексадеканом и водой, и последний уменьшается с уменьшением длины углеводородной цепи (137). Кроме того, есть основания предполагать, что связывание анионных форм SCFAs и MCFAs с митохондриальными переносчиками анионов ниже, чем связывание анионов LCFA. Такая точка зрения подтверждается наблюдением, что ингибирование аденинового нуклеотида-носителя тиоэфирами ацил-КоА снижается с увеличением длины их углеводородной цепи (138).Тем не менее, есть доказательства, в основном из исследований изолированных клеток печени и перфузируемой печени, что SCFAs и MCFAs могут инициировать потерю АТФ (89, 96, 97, 99, 100). Было высказано предположение (100), что этот эффект может быть связан с бесполезным чередованием этерифицированных форм SCFA или MCFA и их тиоэфиров ацил-CoA. Это приведет к чистому использованию АТФ. Таким образом, предполагается высокая скорость гидролиза ацетил-КоА в гепатоцитах крысы, его стимуляция увеличением концентрации ацетата и циклическое переключение субстрата между ацетатом и ацетил-КоА (139–141).В случае ацетата этот цикл, вероятно, происходит в цитоплазме гепатоцитов и может составлять до 1% от общего тепловыделения (141).
Известно, что бутират и октаноат стимулируют поглощение кислорода перфузируемой печенью и изолированными гепатоцитами, повышают активность митохондрий и поддерживают АТФ-зависимую выработку глюкозы (100, 101, 109). Однако неоднократно наблюдалось, что эти две жирные кислоты увеличивают поглощение кислорода и резко снижают соотношение АТФ / О при добавлении в инкубационную или перфузионную среду с добавлением пирувата плюс лактат или только лактата (89, 96, 97, 100), таким образом, указывая на нарушение выработки энергии.Механизм этого эффекта нельзя объяснить протонофорным разобщением окислительного фосфорилирования этими SCFAs и MCFA, поскольку этот механизм типичен для LCFA (см. Выше). Основным аргументом против такого механизма было наблюдение, что олигомицин, известный ингибитор митохондриальной АТФазы / АТФ-синтазы, отменяет большую часть повышенного потребления кислорода печенью (89, 99), а также стимулированное октаноатом поглощение кислорода изолированными крысами. митохондрии печени (100). Другие авторы связывают нарушение энергетического обмена бутиратом и октаноатом со стимуляцией Na + / K + -АТФазы (99) или с увеличением отношения FADH 2 / NADH из-за β-окисления (96 ).Однако эти объяснения кажутся маловероятными, поскольку подобное повышенное поглощение кислорода в присутствии LCFA не было чувствительным к олигомицину. Еще одним ключом к разгадке, по-видимому, был стационарный повышенный уровень АМФ, который не наблюдался с ДЦЖК (89, 100, 142–144). Это указывает на повышенный оборот АТФ в митохондриях, а не на его нарушение производства. Кроме того, этот предполагаемый оборот АТФ конкурировал с внутримитохондриальными АТФ-зависимыми реакциями, то есть карбоксилированием пирувата (100) и синтезом цитруллина (144).Мы показали (100), что это явление связано с усиленной активацией октаноата во внутреннем митохондриальном компартменте, сопровождающейся использованием двух высокоэнергетических связей на каждую образовавшуюся молекулу октаноил-КоА. Поскольку как октаноил-АМФ, так и октаноил-КоА могут быть частично гидролизованы в митохондриальном матриксе, возник бесполезный цикл, ответственный за повышенное внутримитохондриальное потребление АТФ, что привело к снижению потенциала митохондриальной мембраны и, таким образом, к увеличению поглощения кислорода ().Этот механизм может предотвратить резкое истощение внутримитохондриального свободного КоА при большом количестве SCFA и MCFA в воротной вене. Кроме того, связанный с активацией октаноата повышенный уровень АМФ снижает внутримитохондриальный пул обмениваемых адениновых нуклеотидов, АТФ и АДФ, событие, которое замедляет скорость работы адениннуклеотидной транслоказы и, таким образом, увеличивает контролирующую силу этого переносчика в отношении общего потока. окислительной генерации АТФ (145).
Разъединение LCFAs и псевдо-разобщение SCFAs и MCFAs активированных митохондрий.A: Реальное протонофорное разобщение LCFA. Недиссоциированные LCFAs подвергаются спонтанным перекидным движениям через внутреннюю митохондриальную мембрану. В щелочной среде на внутренней (матричной) стороне мембраны они подвергаются диссоциации на протон (H + ) и анион жирной кислоты (RCOO —), который впоследствии переносится переносчиком адениновых нуклеотидов (ANT). и другие переносчики митохондриальных анионов обратно на внешнюю сторону мембраны. Здесь анион LCFA репротонируется и может претерпеть еще один триггерный перенос.B: Псевдо-разобщение по SCFA и MCFA. SCFAs и MCFAs активируются до своих тиоэфиров CoA в компартменте митохондриального матрикса. В этом процессе используется АТФ и высвобождается АМФ и пирофосфат (PP i ). АМФ может впоследствии реагировать с АТФ, давая две молекулы АДФ, которые рефосфорилируются за счет митохондриального трансмембранного потенциала (ΔΨ m ), производя, таким образом, эффект разобщения. Кроме того, как ацил-АМФ, так и ацил-КоА подвергаются медленному гидролизу, что увеличивает производство АМФ и бесполезное использование энергии.
Эти специфические свойства SCFA и MCFA могут объяснить хорошо известный факт, что диеты, богатые этими жирными кислотами, увеличивают расход энергии и уменьшают ожирение (146–148). Сообщалось (103, 149), что MCFA, в отличие от LCFA, способствуют поддержанию высокой чувствительности мышечных клеток к инсулину. Однако эта точка зрения не была подтверждена другими авторами (150). Также стоит отметить, что эфиры CoA SCFAs и MCFAs накапливаются в тканях при различных патологических ситуациях, таких как синдром Рейе (151).Кроме того, сообщалось (152), что октаноил-КоА при низких миллимолярных концентрациях оказывает сильное ингибирование активности комплекса III дыхательной цепи.
Генерация активных форм кислорода
Потребление кислорода митохондриями сопровождается образованием активных форм кислорода (АФК), из которых β-окисление является наиболее важным источником (153–156). Теоретически одноэлектронный перенос молекулярному кислороду с образованием супероксида может происходить от комплекса I дыхательной цепи, а также от ацил-КоА дегидрогеназы, флавопротеина переноса электрона (ETF), ETF-убихинон оксидоредуктазы и комплекса III (подробнее в легенде рисунка).Действительно, для митохондрий скелетных мышц было продемонстрировано, что несколько сайтов вовлечены в образование супероксида, связанное с β-окислением (157). H 2 O 2 Высвобождение митохондриями из (крыс) скелетных мышц, сердца и печени измеряли с карнитиновыми производными пальмитата, октаноата и бутирата в качестве субстратов (156, 158). Эти исследования показали, что генерация АФК, связанных с β-окислением, аналогична ДЦЖК и СЦЖК (156, 158). Напротив, сообщалось, что миотрубки C2C12, обработанные каприновой (C 10 ) или лауриновой (C 12 ) кислотой, генерируют меньше ROS, чем те, которые обработаны LCFA, миристиновой или пальмитиновой кислотой; тогда как потребление кислорода выше у MCFA, чем у LCFA (149).Эти авторы предполагают, что это снижение продукции ROS может быть связано с повышенной экспрессией разобщающего белка-3 с помощью MCFAs. Однако другие авторы (160) не наблюдали повышенной экспрессии разобщающего белка-3 в сердцах крыс, получавших диету, богатую MCFA.
Перенос электрона от жирных кислот к комплексу IV во время β-окисления и возможные места образования супероксида. Показана упрощенная схема, суммирующая сайты образования супероксида, поддерживаемые митохондриальной деградацией тиоэфиров жирных кислот.Электроны передаются от первого фермента пути β-окисления, ацил-CoA-дегидрогеназы (Acyl-CoA-DH), и передаются через ETF к переносящей электроны убихинон-оксидоредуктазе (ETF-QOR). ETF-QOR восстанавливает убихинон (Q) до убихинола (QH 2 ). Наконец, убихинол окисляется до убихинона, а затем электроны переходят в комплекс III. 3-гидроксиацил-КоА-дегидрогеназа (НО-КоА-DH), третий фермент пути β-окисления, который окисляет НО-ацил-КоА до кетоацил-КоА, отдает электроны непосредственно комплексу I.Места образования супероксида отмечены красным.
Наряду с их прямой ролью в генерации АФК в качестве доноров электронов для дыхательной цепи в β-окислении, жирные кислоты также имеют косвенное влияние на производство супероксида из-за изменения как скорости потока электронов вдоль дыхательной цепи, так и степени энергии. связь. Как мы подробно обсуждали в другом месте (160), LCFAs потенцируют генерацию ROS из-за их слабого ингибирования электронного потока на уровнях комплексов I и III, скорее всего, за счет взаимодействия внутри сложной субъединичной структуры, а также между комплексами III и IV из-за к высвобождению цитохрома с из внутренней мембраны.Эти эффекты возникают при генерации АФК при так называемом прямом переносе электронов. С другой стороны, из-за протонофорного действия LCFAs на внутренней митохондриальной мембране («мягкий эффект разобщения») они сильно уменьшают генерацию ROS в обратном режиме транспорта электронов (160). В отличие от этого, SCFAs и MCFAs, по крайней мере, низшие члены последних, на низких физиологических уровнях не влияют на функционирование цепи переноса электронов и не оказывают протонофорного эффекта на внутреннюю мембрану митохондрий.Однако, с другой стороны, чрезмерное накопление MCFAs, которое происходит при врожденном дефиците ацил-CoA-дегидрогеназы со средней длиной цепи (45) и связано с нарушением комплексов митохондриальной дыхательной цепи (46), может приводить к увеличению продукции ROS. Это приводит к усилению перекисного окисления липидов, образованию карбонилов белка (как продуктов перекисного окисления) и снижению неферментативной антиоксидантной защиты (161).
8.2 Катаболизм углеводов — микробиология
Цели обучения
Объясните, почему гликолиз не зависит от кислорода
Определите и опишите чистый выход трехуглеродных молекул, АТФ и НАДН в результате гликолиза
Объясните, как молекулы трехуглеродного пирувата превращаются в двухуглеродные ацетильные группы, которые могут попасть в цикл Кребса.
Определите и опишите чистый выход CO 2 , GTP / ATP, FADH 2 и NADH из цикла Кребса
Объясните, как промежуточные молекулы углерода цикла Кребса можно использовать в ячейке.
Существуют обширные ферментные пути расщепления углеводов с целью захвата энергии в АТФ-связях. Кроме того, многие катаболические пути производят промежуточные молекулы, которые также используются в качестве строительных блоков для анаболизма. Понимание этих процессов важно по нескольким причинам.Во-первых, поскольку основные метаболические процессы являются общими для широкого круга хемогетеротрофных организмов, мы можем многое узнать о метаболизме человека, изучая метаболизм более легко поддающихся манипуляциям бактерий, таких как E. coli . Во-вторых, поскольку патогены животных и человека также являются хемогетеротрофами, изучение деталей метаболизма этих бактерий, включая возможные различия между бактериальными и человеческими путями, полезно для диагностики патогенов, а также для открытия противомикробных методов лечения, направленных на конкретные патогены.Наконец, конкретное изучение путей, участвующих в хемогетеротрофном метаболизме, также служит основой для сравнения других, более необычных метаболических стратегий, используемых микробами. Хотя химический источник электронов, инициирующих перенос электронов, различается у хемогетеророфов и хемоавтотрофов, многие аналогичные процессы используются в обоих типах организмов.
Типичный пример, используемый для ознакомления студентов с понятиями метаболизма, — катаболизм углеводов. Для хемогетеротрофов наши примеры метаболизма начинаются с катаболизма полисахаридов, таких как гликоген, крахмал или целлюлоза.Ферменты, такие как амилаза, расщепляющая гликоген или крахмал, и целлюлазы, расщепляющие целлюлозу, могут вызывать гидролиз гликозидных связей между мономерами глюкозы в этих полимерах, высвобождая глюкозу для дальнейшего катаболизма.
Гликолиз
Для бактерий, эукариот и большинства архей гликолиз является наиболее распространенным путем катаболизма глюкозы; он производит энергию, снижает переносчики электронов и молекулы-предшественники для клеточного метаболизма. Каждый живой организм выполняет ту или иную форму гликолиза, предполагая, что этот механизм является древним универсальным метаболическим процессом.Сам процесс не использует кислород; однако гликолиз может сочетаться с дополнительными метаболическими процессами, которые являются либо аэробными, либо анаэробными. Гликолиз происходит в цитоплазме прокариотических и эукариотических клеток. Он начинается с одной молекулы глюкозы с шестью атомами углерода и заканчивается двумя молекулами трехуглеродного сахара, называемого пируватом. Пируват может расщепляться дальше после гликолиза, чтобы использовать больше энергии за счет аэробного или анаэробного дыхания, но многие организмы, включая многие микробы, могут быть не в состоянии дышать; для этих организмов гликолиз может быть единственным источником образования АТФ.
Тип гликолиза, обнаруживаемый у животных и наиболее часто встречающийся у микробов, — это путь Эмбдена-Мейерхоф-Парнаса (EMP), названный в честь Густава Эмбдена (1874–1933), Отто Мейерхофа (1884–1951) и Якуба Парнаса (1884). –1949). Гликолиз с использованием пути ЭМП состоит из двух отдельных фаз (рис. 8.10). Первая часть пути, называемая фазой инвестирования энергии, использует энергию двух молекул АТФ для модификации молекулы глюкозы, так что молекула шестиуглеродного сахара может быть равномерно разделена на две фосфорилированные трехуглеродные молекулы, называемые глицеральдегид-3-фосфатом (G3P). ).Вторая часть пути, называемая фазой выплаты энергии, извлекает энергию, окисляя G3P до пирувата, производя четыре молекулы АТФ и восстанавливая две молекулы NAD + до двух молекул NADH, используя электроны, происходящие из глюкозы. (Обсуждение и иллюстрация полного пути ЭМП с химическими структурами и названиями ферментов приведены в Приложении C.)
Молекулы АТФ, продуцируемые во время фазы выплаты энергии гликолиза, образуются путем фосфорилирования на уровне субстрата (Рисунок 8.11), один из двух механизмов производства АТФ. При фосфорилировании на уровне субстрата фосфатная группа удаляется из органической молекулы и напрямую переносится на доступную молекулу АДФ, производя АТФ. Во время гликолиза высокоэнергетические фосфатные группы из промежуточных молекул добавляются к АДФ для образования АТФ.
В целом, в этом процессе гликолиза чистая прибыль от распада одной молекулы глюкозы составляет:
две молекулы АТФ
две молекулы НАДН, а
две молекулы пирувата.
Рис. 8.10 Фаза инвестирования энергии в пути гликолиза Эмбдена-Мейерхофа-Парнаса использует две молекулы АТФ для фосфорилирования глюкозы с образованием двух молекул глицеральдегид-3-фосфата (G3P). Фаза выплаты энергии использует энергию молекул G3P, производя четыре молекулы АТФ, две молекулы НАДН и два пирувата.
Рис. 8.11. АТФ, образующийся во время гликолиза, является результатом фосфорилирования на уровне субстрата. Здесь показана одна из двух ферментативных реакций в фазе выплаты энергии гликолиза Эмбдена Мейерхофа-Парнаса, которая таким образом производит АТФ.
Другие гликолитические пути
Когда мы говорим о гликолизе, если не указано иное, мы имеем в виду путь ЭМП, используемый животными и многими бактериями. Однако некоторые прокариоты используют альтернативные гликолитические пути. Одной из важных альтернатив является путь Энтнера-Дудорова (ED), названный в честь его первооткрывателей Натана Энтнера и Майкла Дудорова (1911–1975). Хотя некоторые бактерии, включая условно-патогенный грамотрицательный патоген Pseudomonas aeruginosa , содержат только путь ED для гликолиза, другие бактерии, такие как E.coli , обладают способностью использовать либо путь ED, либо путь EMP.
Третий тип гликолитического пути, который происходит во всех клетках, который сильно отличается от двух предыдущих, — это пентозофосфатный путь (PPP), также называемый фосфоглюконатным путем или гексозо-монофосфатным шунтом. Данные свидетельствуют о том, что PPP может быть самым древним универсальным гликолитическим путем. Промежуточные продукты PPP используются для биосинтеза нуклеотидов и аминокислот.Следовательно, этот гликолитический путь может быть предпочтительным, когда клетка нуждается в синтезе нуклеиновой кислоты и / или белка соответственно. Обсуждение и иллюстрация полного пути ED и PPP с химическими структурами и названиями ферментов приведены в Приложении C.
Проверьте свое понимание
Когда организм может использовать путь ED или PPP для гликолиза?
Реакция перехода, кофермент А и цикл Кребса
При гликолизе образуется пируват, который можно дополнительно окислить для захвата большего количества энергии.Чтобы пируват вступил в следующий окислительный путь, он должен сначала декарбоксилироваться ферментным комплексом пируватдегидрогеназа до двухуглеродной ацетильной группы в реакции перехода, также называемой мостиковой реакцией (см. Приложение C и рисунок 8.12). В реакции перехода электроны также переносятся на NAD + с образованием NADH. Чтобы перейти к следующей фазе этого метаболического процесса, сравнительно крошечный двухуглеродный ацетил должен быть присоединен к очень большому соединению-носителю, называемому коэнзимом A (CoA).Реакция перехода происходит в митохондриальном матриксе эукариот; у прокариот это происходит в цитоплазме, потому что у прокариот отсутствуют заключенные в мембраны органеллы.
Рис. 8.12 (a) Коэнзим A показан здесь без присоединенной ацетильной группы. (b) Коэнзим A показан здесь с присоединенной ацетильной группой.
Цикл Кребса переносит оставшиеся электроны от ацетильной группы, образовавшейся во время реакции перехода, на молекулы-носители электронов, тем самым восстанавливая их. Цикл Кребса также происходит в цитоплазме прокариот вместе с гликолизом и реакцией перехода, но он имеет место в митохондриальном матриксе эукариотических клеток, где также происходит реакция перехода.Цикл Кребса назван в честь его первооткрывателя, британского ученого Ганса Адольфа Кребса (1900–1981) и также называется циклом лимонной кислоты или циклом трикарбоновых кислот (TCA), потому что лимонная кислота имеет три карбоксильные группы в своей структуре. В отличие от гликолиза, цикл Кребса является замкнутым циклом: последняя часть пути регенерирует соединение, используемое на первом этапе (рис. 8.13). Восемь стадий цикла представляют собой серию химических реакций, которые захватывают двухуглеродную ацетильную группу (носитель CoA не входит в цикл Кребса) из переходной реакции, которая добавляется к четырехуглеродному промежуточному продукту в цикле Кребса, производство шестиуглеродного промежуточного соединения — лимонной кислоты (что дает альтернативное название для этого цикла).Когда один оборот цикла возвращается к начальной точке четырехуглеродного интермедиата, цикл производит две молекулы CO 2 , одну молекулу АТФ (или эквивалент, такой как гуанозинтрифосфат [GTP]), продуцируемый фосфорилированием на уровне субстрата. , а также три молекулы НАДН и одна молекула ФАДН 2 . (Обсуждение и подробная иллюстрация полного цикла Кребса приведены в Приложении C.)
Хотя многие организмы используют цикл Кребса, как описано как часть метаболизма глюкозы, некоторые из промежуточных соединений в цикле Кребса могут использоваться для синтеза широкого спектра важных клеточных молекул, включая аминокислоты, хлорофиллы, жирные кислоты и нуклеотиды; следовательно, цикл одновременно анаболический и катаболический (Рисунок 8.14).
Рис. 8.13. Здесь кратко описан цикл Кребса, также известный как цикл лимонной кислоты. Обратите внимание, что входящий двухуглеродный ацетил приводит к основным выходам за один оборот двух молекул CO 2 , трех NADH, одной FADH 2 и одной молекулы АТФ (или GTP), образованных фосфорилированием на уровне субстрата. Два витка цикла Кребса необходимы для обработки всего углерода одной молекулы глюкозы.
Рис. 8.14. Многие организмы используют промежуточные продукты цикла Кребса, такие как аминокислоты, жирные кислоты и нуклеотиды, в качестве строительных блоков для биосинтеза.
Короткоцепочечные жирные кислоты как потенциальные регуляторы метаболизма и функции скелетных мышц
1.
О’Коннор, Э. М. Роль кишечной микробиоты в статусе питания. Curr. Opin. Clin. Nutr. Метаб. Уход 16 , 509–516 (2013).
PubMed
Статья
CAS
Google Scholar
2.
Tremaroli, V. & Bäckhed, F. Функциональные взаимодействия между кишечной микробиотой и метаболизмом хозяина. Природа 489 , 242–249 (2012).
CAS
PubMed
Статья
Google Scholar
3.
den Besten, G. et al. Роль короткоцепочечных жирных кислот во взаимодействии между диетой, кишечной микробиотой и энергетическим обменом хозяина. J. Lipid Res. 54 , 2325–2340 (2013).
Артикул
CAS
Google Scholar
4.
Моррисон Д.Дж. И Престон Т. Образование короткоцепочечных жирных кислот кишечной микробиотой и их влияние на метаболизм человека. Кишечные микробы 7 , 189–200 (2016).
PubMed
PubMed Central
Статья
Google Scholar
5.
Canfora, E. E., Jocken, J. W. & Blaak, E. E. Короткоцепочечные жирные кислоты, контролирующие массу тела и чувствительность к инсулину. Nat. Rev. Endocrinol. 11 , 577–591 (2015).
CAS
PubMed
Статья
Google Scholar
6.
Далиле, Б., Ван Ауденхов, Л., Вервлиет, Б. и Вербеке, К. Роль короткоцепочечных жирных кислот в коммуникации между микробиотой, кишечником и мозгом. Nat. Преподобный Гастроэнтерол. Гепатол. 16 , 461–478 (2019).
PubMed
Статья
Google Scholar
7.
Мандалия, Д. К. и Сешадри, С.Короткоцепочечные жирные кислоты, дисфункция поджелудочной железы и диабет 2 типа. Панкреатология 19 , 280–284 (2019).
CAS
PubMed
Статья
Google Scholar
8.
Чжоу Д. и Фань Дж. Г. Микробные метаболиты при неалкогольной жировой болезни печени. World J. Gastroenterol. 25 , 2019–2028 (2019).
CAS
PubMed
PubMed Central
Статья
Google Scholar
9.
Zurlo, F., Larson, K., Bogardus, C. & Ravussin, E. Метаболизм скелетных мышц является основным фактором, определяющим расход энергии в состоянии покоя. J. Clin. Вкладывать деньги. 86 , 1423–1427 (1990).
CAS
PubMed
PubMed Central
Статья
Google Scholar
10.
Frontera, W. R. & Ochala, J. Скелетные мышцы: краткий обзор структуры и функций. Calcif. Tissue Int. 96 , 183–195 (2015).
CAS
PubMed
Статья
Google Scholar
11.
Ridaura, V. K. et al. Микробиота кишечника близнецов, не согласных с ожирением, модулирует метаболизм у мышей. Наука 341 , 1241214 (2013).
PubMed
Статья
CAS
Google Scholar
12.
Yan, H. et al. Микробиота кишечника может передавать характеристики волокон и липидный метаболический профиль скелетных мышц от свиней к стерильным мышам. Sci. Отчет 6 , 31786 (2016).
CAS
PubMed
PubMed Central
Статья
Google Scholar
13.
Lahiri, S. et al. Микробиота кишечника влияет на массу и функцию скелетных мышц у мышей. Sci. Пер. Med. 11 , eaan5662 (2019).
PubMed
PubMed Central
Статья
CAS
Google Scholar
14.
Нет, К. и др. Кишечные бактерии имеют решающее значение для оптимальной функции мышц: потенциальная связь с гомеостазом глюкозы. Am. J. Physiol. Эндокринол. Метаб. 317 , E158 – E171 (2019).
CAS
PubMed
Статья
Google Scholar
15.
Bindels, L. B. et al. Восстановление определенных уровней лактобацилл снижает маркеры воспаления и мышечной атрофии на мышиной модели с острым лейкозом. PLoS ONE 7 , e37971 (2012).
CAS
PubMed
PubMed Central
Статья
Google Scholar
16.
Chen, Y. M. et al. Lactobacillus plantarum Добавка TWK10 улучшает физическую работоспособность и увеличивает мышечную массу у мышей. Питательные вещества 8 , 205 (2016).
PubMed
PubMed Central
Статья
CAS
Google Scholar
17.
Эверард, А.и другие. Ответы микробиоты кишечника и метаболизма глюкозы и липидов на пребиотики у мышей с генетическим ожирением и резистентностью к лептину, вызванной диетой. Диабет 60 , 2775–2786 (2011).
CAS
PubMed
PubMed Central
Статья
Google Scholar
18.
Buigues, C. et al. Влияние пребиотика на синдром слабости: рандомизированное двойное слепое клиническое исследование. Внутр. J. Mol. Sci. 17 , E932 (2016).
PubMed
Статья
CAS
Google Scholar
19.
Ticinesi, A. et al. Старение кишечной микробиоты на перекрестке между питанием, физической слабостью и саркопенией: существует ли ось кишечник – мышцы? Питательные вещества 9 , 1–20 (2017).
Артикул
CAS
Google Scholar
20.
Grosicki, G.J., Fielding, R.A. & Lustgarten, M. S.Микробиота кишечника способствует возрастным изменениям в размере, составе и функции скелетных мышц: биологическая основа оси кишечник-мышцы. Calcif. Tissue Int. 102 , 433–442 (2018).
CAS
PubMed
Статья
Google Scholar
21.
Evans, W. J. Потеря скелетных мышц: кахексия, саркопения и бездействие. Am. J. Clin. Nutr. 91 , 1123S – 1127S (2010 г.).
CAS
PubMed
Статья
Google Scholar
22.
Penna, F. et al. Скелетная мышца как активный игрок против раковой кахексии. Фронт. Physiol. 10 , 41 (2019).
PubMed
PubMed Central
Статья
Google Scholar
23.
Brown, A. J. et al. Рецепторы GPR41 и GPR43, связанные с белками G-сирот, активируются пропионатом и другими карбоновыми кислотами с короткой цепью. J. Biol. Chem. 278 , 11312–11319 (2003).
CAS
PubMed
Статья
Google Scholar
24.
Nilsson, N.E., Kotarsky, K., Owman, C. & Olde, B. Идентификация рецептора свободных жирных кислот, FFA2R, экспрессируемого на лейкоцитах и активируемого короткоцепочечными жирными кислотами. Biochem. Биофиз. Res. Commun. 303 , 1047–1052 (2003).
CAS
PubMed
Статья
Google Scholar
25.
Gao, Z. et al. Бутират улучшает чувствительность к инсулину и увеличивает расход энергии у мышей. Диабет 58 , 1509–1517 (2009).
CAS
PubMed
PubMed Central
Статья
Google Scholar
26.
Maruta, H. et al. Активация AMP-активированной протеинкиназы и стимуляция энергетического обмена уксусной кислотой в клетках мышечной трубки L6. PLoS ONE 11 , e0158055 (2016).
PubMed
PubMed Central
Статья
CAS
Google Scholar
27.
Hong, J.и другие. Бутират снижает ожирение, вызванное диетой с высоким содержанием жиров, за счет активации адипонектин-опосредованного пути и стимуляции митохондриальной функции в скелетных мышцах мышей. Oncotarget 7 , 56071–56082 (2016).
PubMed
PubMed Central
Статья
Google Scholar
28.
Liu, L., Fu, C. & Li, F. Ацетат влияет на процесс липидного обмена в печени, скелетных мышцах и жировой ткани кролика. Животные (Базель) 9 , e799 (2019).
Артикул
Google Scholar
29.
Walsh, M. E. et al. Бутират, ингибитор гистондеацетилазы, улучшает метаболизм и снижает атрофию мышц при старении. Ячейка старения 14 , 957–970 (2015).
CAS
PubMed
PubMed Central
Статья
Google Scholar
30.
den Besten, G.и другие. Короткоцепочечные жирные кислоты защищают от ожирения, вызванного диетой с высоким содержанием жиров, посредством PPARγ-зависимого перехода от липогенеза к окислению жиров. Диабет 64 , 2398–2408 (2015).
Артикул
CAS
Google Scholar
31.
Pan, J.H. et al. Уксусная кислота повышает выносливость мышей, тренируемых физическими упражнениями, за счет увеличения окислительных свойств скелетных мышц. Biosci. Biotechnol. Biochem. 79 , 1535–1541 (2015).
CAS
PubMed
Статья
Google Scholar
32.
Han, J.H. et al. Влияние пропионата и валерата на чувствительность к инсулину для поглощения глюкозы адипоцитами 3T3-L1 и миотрубками C2C12 через рецептор 41, связанный с G-белком. PLoS ONE 9 , e95268 (2014).
PubMed
PubMed Central
Статья
Google Scholar
33.
Фусими Т.и другие. Кормление уксусной кислотой увеличивает восполнение запасов гликогена в печени и скелетных мышцах крыс. J. Nutr. 131 , 1973–1977 (2001).
CAS
PubMed
Статья
Google Scholar
34.
Nakao, C. et al. Влияние ацетата на восполнение запасов гликогена в печени и скелетных мышцах после изнурительного плавания у крыс. Сканд. J. Med. Sci. Спорт 11 , 33–37 (2001).
CAS
PubMed
Статья
Google Scholar
35.
Фусими Т. и Сато Ю. Влияние кормления уксусной кислотой на циркадные изменения гликогена и метаболитов глюкозы и липидов в печени и скелетных мышцах крыс. Br. J. Nutr. 94 , 714–719 (2005).
CAS
PubMed
Статья
Google Scholar
36.
Сакакибара, С., Ямаути, Т., Осима, Ю., Цукамото, Ю. и Кадоваки, Т. Уксусная кислота активирует печеночный AMPK и снижает гипергликемию у мышей с диабетом KK-A (y). Biochem. Биофиз. Res. Commun. 344 , 597–604 (2006).
CAS
PubMed
Статья
Google Scholar
37.
Chriett, S., Zerzaihi, O., Vidal, H. & Pirola, L. Бутират натрия, ингибитор гистондеацетилазы, улучшает передачу сигналов инсулина при пальмитат-индуцированной резистентности к инсулину в мышечных клетках крыс L6 посредством эпигенетически опосредованного повышения. -регулирование Irs1. Мол. Клетка. Эндокринол. 439 , 224–232 (2017).
CAS
PubMed
Статья
Google Scholar
38.
Wakelam, M. J. & Pette, D. Контроль глюкозо-1,6-бисфосфата с помощью состояния развития и гормональной стимуляции культивируемой мышечной ткани. Biochem. J. 204 , 765–769 (1982).
Yamashita, H. et al. Влияние ацетата на метаболизм липидов в мышцах и жировых тканях крыс Otsuka Long-Evans Tokushima Fatty (OLETF) с диабетом 2 типа. Biosci. Biotechnol. Biochem. 73 , 570–576 (2009).
CAS
PubMed
Статья
Google Scholar
41.
Gonzalez, A. et al. Бутират натрия улучшает инсулинорезистентность и почечную недостаточность у крыс с ХЗП, регулируя кишечную проницаемость и экспрессию муцина. Нефрол. Набирать номер. Пересадка. 34 , 783–794 (2019).
CAS
PubMed
Статья
Google Scholar
42.
Койстинен, Х. А. и Зиерат, Дж. Р. Регулирование транспорта глюкозы в скелетных мышцах человека. Ann. Med. 34 , 410–418 (2002).
CAS
PubMed
Статья
Google Scholar
43.
Типтон К. Д. и Вулф Р. Р. Упражнения, метаболизм белков и рост мышц. Внутр. J. Sport Nutr. Упражнение. Метаб. 11 , 109–132 (2001).
CAS
PubMed
Статья
Google Scholar
44.
Кондо, Т., Киши, М., Фусими, Т. и Кага, Т. Уксусная кислота усиливает экспрессию генов ферментов окисления жирных кислот в печени, подавляя накопление жира в организме. J. Agric. Food Chem. 57 , 5982–5986 (2009).
CAS
PubMed
Статья
Google Scholar
45.
Henagan, T. M. et al. Бутират натрия эпигенетически модулирует вызванную диетой с высоким содержанием жиров митохондриальную адаптацию скелетных мышц, ожирение и инсулинорезистентность посредством позиционирования нуклеосом. Br. J. Pharmacol. 172 , 2782–2798 (2015).
CAS
PubMed
PubMed Central
Статья
Google Scholar
46.
Йоргенсен, Х., Ларсен, Т., Чжао, X.-Q. & Eggum, B.O. Энергетическая ценность короткоцепочечных жирных кислот, введенных в слепую кишку свиней. Br. J. Nutr. 77 , 745–756 (1997).
PubMed
Статья
Google Scholar
47.
Имото, С. и Намиока, С. Питательная ценность ацетата у растущих свиней. J. Anim. Sci. 56 , 858–866 (1983).
CAS
PubMed
Статья
Google Scholar
48.
Clarke, S. F. et al. Физические упражнения и связанные с ними экстремальные диеты влияют на микробное разнообразие кишечника. Кишечник 63 , 1913–1920 (2014).
CAS
PubMed
Статья
Google Scholar
49.
Barton, W. et al. Микробиом профессиональных спортсменов отличается от микробиома людей, ведущих малоподвижный образ жизни, по составу и особенно по функциональному метаболическому уровню. Кишечник 67 , 625–633 (2018).
CAS
PubMed
Google Scholar
50.
Allen, J. M. et al. Упражнения изменяют состав и функции кишечной микробиоты у худых и тучных людей. Med. Sci. Спортивные упражнения. 50 , 747–757 (2018).
PubMed
Статья
Google Scholar
51.
Munukka, E. et al. Шестинедельные упражнения на выносливость изменяют метагеном кишечника, что не отражается на системном метаболизме у полных женщин. Фронт. Microbiol. 9 , 2323 (2018).
PubMed
PubMed Central
Статья
Google Scholar
52.
Okamoto, T. et al. Микробиом усиливает упражнения на выносливость за счет выработки ацетата в кишечнике. Am. J. Physiol. Эндокринол. Метаб. 316 , E956 – E966 (2019).
CAS
PubMed
Статья
Google Scholar
53.
Scheiman, J. et al. Метаомический анализ профессиональных спортсменов выявляет микроб, повышающий производительность, который функционирует посредством метаболизма лактата. Nat. Med. 25 , 1104–1109 (2019).
CAS
PubMed
PubMed Central
Статья
Google Scholar
54.
Клаузен М. Р. и Мортенсен П. Б. Кинетические исследования метаболизма короткоцепочечных жирных кислот и глюкозы в колоноцитах при язвенном колите. Кишечник 37 , 684–689 (1995).
CAS
PubMed
PubMed Central
Статья
Google Scholar
55.
van Loon, L. J. C., Greenhaff, P. L., Constantin-Teodosiu, D., Saris, W. H. M. & Wagenmakers, A. J. M. Влияние увеличения интенсивности упражнений на использование мышечного топлива у людей. J. Physiol. (Лондон) 536 , 295–304 (2001).
Артикул
Google Scholar
56.
Бергстрём, Дж., Хермансен, Л., Халтман, Э. и Салтин, Б. Диета, гликоген в мышцах и физическая работоспособность. Acta Physiol. Сканд. 71 , 140–150 (1967).
PubMed
Статья
PubMed Central
Google Scholar
57.
van der Beek, C. M., Dejong, C. H. C., Troost, F. J., Masclee, A. A. M. и Lenaerts, K. Роль короткоцепочечных жирных кислот в воспалении толстой кишки, канцерогенезе, защите и заживлении слизистой оболочки. Nutr. Ред. 75 , 286–305 (2017).
PubMed
Статья
Google Scholar
58.
den Besten, G. et al. Полученные из кишечника короткоцепочечные жирные кислоты быстро ассимилируются в углеводы и липиды хозяина. Am. J. Physiol. Гастроинтест. Liver Physiol. 305 , G900 – G910 (2013).
Артикул
CAS
Google Scholar
59.
Boets, E. et al. Системная доступность и метаболизм короткоцепочечных жирных кислот толстого кишечника у здоровых субъектов: исследование стабильных изотопов. J. Physiol. (Лондон) 595 , 541–555 (2017).
CAS
Статья
Google Scholar
60.
Михайлова, М. М. и Шоу, Р. Дж. Путь передачи сигналов AMPK координирует рост клеток, аутофагию и метаболизм. Nat. Cell Biol. 13 , 1016–1023 (2011).
CAS
PubMed
PubMed Central
Статья
Google Scholar
61.
Akimoto, T. et al. Физические упражнения стимулируют транскрипцию Pgc-1α в скелетных мышцах за счет активации пути p38 MAPK. J. Biol. Chem. 280 , 19587–19593 (2005).
CAS
PubMed
Статья
Google Scholar
62.
Jäger, S., Handschin, C., St-Pierre, J.И Шпигельман, Б. М. Действие AMP-активированной протеинкиназы (AMPK) в скелетных мышцах посредством прямого фосфорилирования PGC-1α. Proc. Natl Acad. Sci. США 104 , 12017–12022 (2007).
PubMed
Статья
CAS
Google Scholar
63.
Фернандес-Маркос, П. Дж. И Ауверкс, Дж. Регулирование PGC-1α, узлового регулятора митохондриального биогенеза. Am. J. Clin. Nutr. 93 , 884С – 90 (2011).
CAS
PubMed
PubMed Central
Статья
Google Scholar
64.
Конли, К. Э. Митохондрии в движении: оптимизация окислительного фосфорилирования для повышения работоспособности. J. Exp. Биол. 219 , 243–249 (2016).
PubMed
PubMed Central
Статья
Google Scholar
65.
Эренборг, Э. и Крук, А. Регулирование физиологии и метаболизма скелетных мышц дельта рецептора, активируемого пролифератором пероксисом. Pharmacol. Ред. 61 , 373–393 (2009).
CAS
PubMed
Статья
Google Scholar
66.
Krämer, D. K. et al. Прямая активация транспорта глюкозы в первичных мышечных трубках человека после активации дельта рецептора, активируемого пролифератором пероксисом. Диабет 54 , 1157–1163 (2005).
PubMed
Статья
Google Scholar
67.
Fan, W. et al. PPARδ повышает выносливость при беге за счет сохранения глюкозы. Cell Metab. 25 , 1186–1193.e4 (2017).
CAS
PubMed
PubMed Central
Статья
Google Scholar
68.
Krämer, D. K. et al. Роль киназы AMP и PPARδ в регуляции метаболизма липидов и глюкозы в скелетных мышцах человека. J. Biol. Chem. 282 , 19313–19320 (2007).
PubMed
Статья
Google Scholar
69.
Finck, B.N. et al. Возможная связь между передачей сигналов рецептора-альфа, активируемого пролифератором пероксисом в мышцах, и диабетом, связанным с ожирением. Cell Metab. 1 , 133–144 (2005).
CAS
PubMed
Статья
Google Scholar
70.
Амин, Р. Х., Мэтьюз, С. Т., Кэмп, Х. С., Динг, Л. и Лефф, Т. Селективная активация PPARγ в скелетных мышцах индуцирует эндогенное производство адипонектина и защищает мышей от индуцированной диетой резистентности к инсулину. Am. J. Physiol. Эндокринол. Метаб. 298 , E28 – E37 (2010).
CAS
PubMed
Статья
Google Scholar
71.
Уолш, М. Э. и Ван Реммен, Х. Новые роли гистоновых деацетилаз в возрастной атрофии мышц. Nutr. Здоровое старение 4 , 17–30 (2016).
CAS
PubMed
PubMed Central
Статья
Google Scholar
72.
Beharry, A. W. et al. HDAC1 активирует FoxO и является достаточным и необходимым для атрофии скелетных мышц. J. Cell Sci. 127 , 1441–1453 (2014).
CAS
PubMed
PubMed Central
Статья
Google Scholar
73.
Bongers, K. S. et al. Денервация скелетных мышц вызывает атрофию скелетных мышц посредством пути, в котором участвуют как Gadd45a, так и HDAC4. Am. J. Physiol. Эндокринол.Метаб. 305 , E907 – E915 (2013).
CAS
PubMed
PubMed Central
Статья
Google Scholar
74.
Лу, Дж., Маккинси, Т. А., Никол, Р. Л. и Олсон, Е. Н. Сигнал-зависимая активация фактора транскрипции MEF2 путем диссоциации от гистондеацетилаз. Proc. Natl Acad. Sci. США 97 , 4070–4075 (2000).
CAS
PubMed
Статья
Google Scholar
75.
Galmozzi, A. et al. Ингибирование гистоновых деацетилаз класса I раскрывает митохондриальную сигнатуру и усиливает окислительный метаболизм в скелетных мышцах и жировой ткани. Диабет 62 , 732–742 (2013).
CAS
PubMed
PubMed Central
Статья
Google Scholar
76.
McGee, S. L. & Hargreaves, M. Модификации гистонов и экспрессия метаболических генов в скелетных мышцах. Clin. Exp.Pharmacol. Physiol. 37 , 392–396 (2010).
CAS
PubMed
Статья
Google Scholar
77.
Дэви, Дж. Р. Ингибирование активности гистондеацетилазы бутиратом. J. Nutr. 133 (Дополнение), 2485S – 2493S (2003).
CAS
PubMed
Статья
Google Scholar
78.
Waldecker, M., Kautenburger, T., Daumann, H., Busch, C. & Schrenk, D. Ингибирование активности гистон-деацетилазы короткоцепочечными жирными кислотами и некоторыми метаболитами полифенолов, образующимися в толстой кишке. J. Nutr. Biochem. 19 , 587–593 (2008).
CAS
PubMed
Статья
Google Scholar
79.
Soliman, M. L. & Rosenberger, T. A. Добавка ацетата увеличивает ацетилирование гистонов в головном мозге и подавляет активность и экспрессию гистондеацетилазы. Мол. Клетка. Biochem. 352 , 173–180 (2011).
CAS
PubMed
Статья
Google Scholar
80.
Salminen, A., Kauppinen, A. & Kaarniranta, K. Передача сигналов AMPK / Snf1 регулирует ацетилирование гистонов: влияние на экспрессию генов и эпигенетические функции. Cell. Сигнал. 28 , 887–895 (2016).
CAS
PubMed
Статья
Google Scholar
81.
McGee, S. L. et al. AMP-активированная протеинкиназа регулирует транскрипцию GLUT4 путем фосфорилирования гистондеацетилазы 5. Диабет 57 , 860–867 (2008).
CAS
PubMed
Статья
Google Scholar
82.
Штеффен, Р. П., Маккензи, Дж. Э. и Хэдди, Ф. Дж. Возможная роль ацетата в гиперемии при физической нагрузке в скелетных мышцах собак. Pflugers Arch. 392 , 315–321 (1982).
CAS
PubMed
Статья
Google Scholar
83.
Штеффен, Р. П., Маккензи, Дж. Э., Бокман, Э. Л. и Хэдди, Ф. Дж. Изменения аденозина в тонкой мышце собаки во время физических упражнений и инфузии ацетата. Am. J. Physiol. 244 , h487 – h495 (1983).
CAS
PubMed
Google Scholar
84.
Lin, H. V. et al. Бутират и пропионат защищают от ожирения, вызванного диетой, и регулируют гормоны кишечника с помощью независимых от рецептора 3 свободных жирных кислот механизмов. PLoS ONE 7 , e35240 (2012).
CAS
PubMed
PubMed Central
Статья
Google Scholar
85.
Villanueva-Peñacarrillo, M. L., Alcántara, A. I., Clemente, F., Delgado, E. & Valverde, I. Сильный гликогенный эффект GLP-1 (7-36) амида в скелетных мышцах крысы. Diabetologia 37 , 1163–1166 (1994).
PubMed
Статья
Google Scholar
86.
Villanueva-Peñacarrillo, M. L. et al. Характеристика эффектов GLP-1 на метаболизм глюкозы в скелетных мышцах человека у пациентов с ожирением. Регул. Pept. 168 , 39–44 (2011).
PubMed
Статья
CAS
Google Scholar
87.
Грин, К. Дж., Хенриксен, Т. И., Педерсен, Б. К. и Соломон, Т. П. J. Глюкагоноподобный пептид-1-индуцированный метаболизм глюкозы в дифференцированных мышечных сателлитных клетках человека ослабляется гипергликемией. PLoS ONE 7 , e44284 (2012).
CAS
PubMed
PubMed Central
Статья
Google Scholar
88.
Chai, W., Zhang, X., Barrett, E. J. & Liu, Z. Глюкагоноподобный пептид 1 задействует микрососудистую сеть мышц и улучшает метаболическое действие инсулина в присутствии инсулинорезистентности. Диабет 63 , 2788–2799 (2014).
CAS
PubMed
PubMed Central
Статья
Google Scholar
89.
Subaran, S.C. et al. GLP-1 в физиологических концентрациях задействует микрососуды скелетных и сердечных мышц у здоровых людей. Clin. Sci. (Лондон) 127 , 163–170 (2014).
CAS
Статья
Google Scholar
90.
Dimitriadis, G., Mitrou, P., Lambadiari, V., Maratou, E. & Raptis, S.A. Влияние инсулина на мышечную и жировую ткань. Diabetes Res. Clin. Практик. 93 (Доп.1), S52 – S59 (2011).
CAS
PubMed
Статья
Google Scholar
91.
Абдулла, Х., Смит, К., Атертон, П. Дж. И Идрис, И. Роль инсулина в регуляции синтеза и распада белков скелетных мышц человека: систематический обзор и метаанализ. Диабетология 59 , 44–55 (2016).
CAS
PubMed
Статья
Google Scholar
92.
Ceddia, R. B. Прямая регуляция метаболизма в скелетных мышцах и жировой ткани лептином: последствия для гомеостаза глюкозы и жирных кислот. Внутр. J. Obes. (Лондон) 29 , 1175–1183 (2005).
CAS
Статья
Google Scholar
93.
Yan, J. et al. Микробиота кишечника индуцирует IGF-1 и способствует образованию и росту костей. Proc. Natl Acad. Sci. США 113 , E7554 – E7563 (2016).
CAS
PubMed
Статья
Google Scholar
94.
Scicchitano, B. M., Rizzuto, E. & Musarò, A. Противодействие истощению мышц при старении и нервно-мышечных заболеваниях: критическая роль IGF-1. Старение (Олбани, штат Нью-Йорк) 1 , 451–457 (2009).
CAS
Статья
Google Scholar
95.
Барклай Р. Д., Бурд Н. А., Тайлер К., Тиллин Н. А. и Маккензи Р. В. Роль сигнального каскада IGF-1 в синтезе мышечного белка и анаболической резистентности при старении скелетных мышц. Фронт. Nutr. 6 , 146 (2019).
PubMed
PubMed Central
Статья
CAS
Google Scholar
96.
Thankamony, A. et al. Низкие уровни циркулирующего IGF-1 у здоровых взрослых связаны со снижением функции β-клеток, повышенным внутримиоцеллюлярным липидом и улучшенной утилизацией жира во время голодания. J. Clin. Эндокринол. Метаб. 99 , 2198–2207 (2014).
CAS
PubMed
PubMed Central
Статья
Google Scholar
97.
Маклафлин, Р. Ф., Бертон, Б. С., Дженсен, М. Е., Бейнс, К. Дж. И Вуд, Л. Г. Короткоцепочечные жирные кислоты, пребиотики, синбиотики и системное воспаление: систематический обзор и метаанализ. Am. J. Clin. Nutr. 106 , 930–945 (2017).
CAS
PubMed
Google Scholar
98.
Li, M. et al. Провоспалительное и противовоспалительное действие короткоцепочечных жирных кислот на иммунные и эндотелиальные клетки. Eur.J. Pharmacol. 831 , 52–59 (2018).
CAS
PubMed
Статья
Google Scholar
99.
Londhe, P. & Guttridge, D. C. Воспаление вызвало потерю скелетных мышц. Кость 80 , 131–142 (2015).
CAS
PubMed
PubMed Central
Статья
Google Scholar
100.
Wu, H. & Ballantyne, C. M.Воспаление скелетных мышц и инсулинорезистентность при ожирении. J. Clin. Вкладывать деньги. 127 , 43–54 (2017).
PubMed
PubMed Central
Статья
Google Scholar
101.
Murakami, Y. et al. Дополнительная эпилактоза предотвращает метаболические нарушения за счет разобщения индукции протеина-1 в скелетных мышцах мышей, получавших диету с высоким содержанием жиров. Br. J. Nutr. 114 , 1774–1783 (2015).
CAS
PubMed
Статья
Google Scholar
102.
Bareja, A. et al. Стволовые клетки скелетных мышц человека и мыши: конвергентные и дивергентные механизмы миогенеза. PLoS ONE 9 , e (2014).
PubMed
PubMed Central
Статья
CAS
Google Scholar
103.
Zhu, C.H. et al. Клеточное старение в миобластах человека преодолевается обратной транскриптазой теломеразы человека и циклин-зависимой киназой 4: последствия для старения мышц и терапевтические стратегии при мышечных дистрофиях. Ячейка старения 6 , 515–523 (2007).
CAS
PubMed
Статья
Google Scholar
104.
Bjursell, M. et al. Улучшенный контроль глюкозы и снижение массы жира в организме у мышей с дефицитом рецептора 2 свободных жирных кислот, получавших диету с высоким содержанием жиров. Am. J. Physiol. Эндокринол. Метаб. 300 , E211 – E220 (2011).
CAS
PubMed
Статья
Google Scholar
105.
Bellahcene, M. et al. Самцы мышей, у которых отсутствует рецептор GPR41, связанный с G-белком, имеют низкий расход энергии и повышенное содержание жира в организме. Br. J. Nutr. 109 , 1755–1764 (2013).
CAS
PubMed
Статья
Google Scholar
106.
Kimura, I. et al. Микробиота кишечника подавляет опосредованное инсулином накопление жира через рецептор короткоцепочечных жирных кислот GPR43. Nat. Commun. 4 , 1829 (2013).
PubMed
PubMed Central
Статья
CAS
Google Scholar
107.
Lundquist, F., Tygstrup, N., Winkler, K., Mellemgaard, K. & Munck-Petersen, S. Метаболизм этанола и производство свободного ацетата в печени человека. J. Clin. Вкладывать деньги. 41 , 955–961 (1962).
CAS
PubMed
PubMed Central
Статья
Google Scholar
108.
Steiner, J. L. & Lang, C.H. Нарушение регуляции метаболизма белков скелетных мышц алкоголем. Am. J. Physiol. Эндокринол. Метаб. 308 , E699 – E712 (2015).
PubMed
PubMed Central
Статья
CAS
Google Scholar
109.
Lang, C.H. et al. Алкогольная интоксикация нарушает фосфорилирование S6K1 и S6 в скелетных мышцах независимо от метаболизма этанола. Алкоголь. Clin. Exp.Res. 28 , 1758–1767 (2004).
CAS
PubMed
Статья
Google Scholar
110.
Preedy, V. R., Keating, J. W. & Peters, T. J. Острые эффекты этанола и ацетальдегида на скорость синтеза белка в богатых клетчаткой скелетных мышцах крыс типа I и типа II. Алкоголь Алкоголь. 27 , 241–251 (1992).
CAS
PubMed
Google Scholar
111.
Вич, Р. Л. и Гитомер, В. Л. Медицинские и метаболические последствия введения ацетата натрия. Adv. Enzyme Regul. 27 , 313–343 (1988).
CAS
PubMed
Статья
Google Scholar
112.
Каммингс, Дж. Х., Помаре, Э. У., Бранч, У. Дж., Нейлор, К. П. Э. и Макфарлейн, Г. Т. Короткоцепочечные жирные кислоты в толстой кишке, воротной, печеночной и венозной крови человека. Кишечник 28 , 1221–1227 (1987).
CAS
PubMed
PubMed Central
Статья
Google Scholar
113.
Peters, S. G., Pomare, E. W. & Fisher, C. A. Концентрации короткоцепочечных жирных кислот в воротной и периферической крови после инстилляции лактулозы слепой кишки во время операции. Кишечник 33 , 1249–1252 (1992).
CAS
PubMed
PubMed Central
Статья
Google Scholar
114.
Bloemen, J. G. et al. Обмен короткоцепочечных жирных кислот через кишечник и печень у людей измеряется во время операции. Clin. Nutr. 28 , 657–661 (2009).
CAS
PubMed
Статья
Google Scholar
115.
van der Beek, C.M. et al. Поглощение печенью бутирата, вводимого ректально, предотвращает повышение системных концентраций бутирата у людей. J. Nutr. 145 , 2019–2024 (2015).
PubMed
Статья
CAS
Google Scholar
116.
Neis, E. P. et al. Дистальное и проксимальное высвобождение короткоцепочечных жирных кислот в кишечнике у человека. Кишечник 68 , 764–765 (2019).
CAS
PubMed
Статья
Google Scholar
Что такое метаболизм?
Метаболизм — это термин, который используется для описания всех химических реакций, участвующих в поддержании жизненного состояния клеток и организма.Обмен веществ можно условно разделить на две категории:
Катаболизм — распад молекул для получения энергии
Анаболизм — синтез всех соединений, необходимых клеткам
Метаболизм тесно связан с питанием и доступностью питательных веществ. Биоэнергетика — это термин, который описывает биохимические или метаболические пути, с помощью которых клетка в конечном итоге получает энергию. Образование энергии — один из жизненно важных компонентов обмена веществ.
Изображение предоставлено: VectorMine / Shutterstock.com
Питание, обмен веществ и энергия
Питание — это ключ к метаболизму. Пути метаболизма зависят от питательных веществ, которые они расщепляют, чтобы произвести энергию. Эта энергия, в свою очередь, требуется организму для синтеза таких молекул, как новые белки и нуклеиновые кислоты (ДНК, РНК).
Питательные вещества, связанные с метаболизмом, включают такие факторы, как потребности организма в различных веществах, индивидуальные функции в организме, необходимое количество и уровень, ниже которого ухудшается состояние здоровья.
Основные питательные вещества обеспечивают энергию (калории) и поставляют необходимые химические вещества, которые сам организм не может синтезировать. Пища содержит множество веществ, которые необходимы для построения, содержания и восстановления тканей тела, а также для его эффективного функционирования.
Диета требует основных питательных веществ, таких как углерод, водород, кислород, азот, фосфор, сера и около 20 других неорганических элементов. Основные элементы представлены углеводами, липидами и белком.Кроме того, необходимы витамины, минералы и вода.
Углеводы в обмене веществ
Продукты питания содержат углеводы в трех формах: крахмал, сахар и целлюлозу (клетчатку). Крахмал и сахар являются основными и необходимыми источниками энергии для человека. Волокна увеличивают объем рациона.
Ткани организма зависят от глюкозы во всех сферах деятельности. Углеводы и сахара производят глюкозу в результате пищеварения или метаболизма.
Общая реакция горения глюкозы записывается как:
C 6 H 12 O 6 + 6 O 2 ——> 6 CO 2 + 6 H 2 O + энергия
Большинство людей потребляют около половины своего рациона в виде углеводов.Это происходит из таких продуктов, как рис, пшеница, хлеб, картофель и макаронные изделия.
Белки в обмене веществ
Белки являются основными строителями тканей в организме. Они являются частью каждой клетки тела. Белки помогают в структуре клеток, функциях, образовании гемоглобина для переноса кислорода, ферментах для выполнения жизненно важных реакций и множестве других функций в организме. Белки также жизненно важны для снабжения азотом генетического материала ДНК и РНК и производства энергии.
Белки необходимы для питания, поскольку содержат аминокислоты.Из 20 или более аминокислот человеческий организм не может синтезировать 8, и они называются незаменимыми аминокислотами.
К незаменимым аминокислотам относятся:
Лизин
Триптофан
метионин
лейцин
Изолейцин
Фенилаланин
Валин
Треонин
Продукты с высоким содержанием белка: яйца, молоко, соевые бобы, мясо, овощи и зерновые.
Жир в обмене веществ
Жиры — это концентрированные источники энергии. Они производят в два раза больше энергии, чем углеводы или белки, в пересчете на вес.
Функции жиров включают:
Помогает формировать ячеистую структуру;
Образует защитную подушку и изоляцию вокруг жизненно важных органов;
Способствует усвоению жирорастворимых витаминов,
Обеспечение резервного хранилища энергии
Незаменимые жирные кислоты включают ненасыщенные жирные кислоты, такие как линолевая, линоленовая и арахидоновая кислоты.Их нужно принимать с пищей. Насыщенные жиры, наряду с холестерином, участвуют в артериосклерозе и сердечных заболеваниях.
Минералы и витамины в обмене веществ
Минералы, содержащиеся в пищевых продуктах, не вносят прямого вклада в энергетические потребности, но важны как регуляторы организма и играют роль в метаболических путях организма. В организме человека содержится более 50 элементов. Было обнаружено, что незаменимыми являются около 25 элементов, а это означает, что их дефицит вызывает определенные симптомы дефицита.
Важные минералы включают:
Кальций
Фосфор
Утюг
Натрий
Калий
Хлорид-ионы
Медь
Кобальт
Марганец
Цинк
Магний
Фтор
Йод
Витамины — это незаменимые органические соединения, которые человеческий организм не может синтезировать сам по себе, и поэтому они должны присутствовать в рационе.Витамины, особенно важные для обмена веществ, включают:
Витамин А
B2 (рибофлавин)
Ниацин или никотиновая кислота
Пантотеновая кислота
Изображение предоставлено: Siberian Art / Shutterstock.com
Метаболические пути
Химические реакции метаболизма организованы в метаболические пути. Они позволяют преобразовать основные химические вещества из пищи с помощью последовательности ферментов через ряд этапов в другое химическое вещество.
Ферменты имеют решающее значение для метаболизма, потому что они позволяют организмам запускать желательные реакции, требующие энергии. Эти реакции также связаны с реакциями, высвобождающими энергию. Поскольку ферменты действуют как катализаторы, они позволяют этим реакциям протекать быстро и эффективно. Ферменты также позволяют регулировать метаболические пути в ответ на изменения в клеточной среде или сигналы от других клеток.
Список литературы
Дополнительная литература
Благоприятное влияние на энергетический метаболизм хозяина короткоцепочечных жирных кислот и витаминов, продуцируемых комменсальными и пробиотическими бактериями | Фабрики микробных клеток
1.
Martens JH, Barg H, Warren MJ, Jahn D. Микробное производство витамина B12. Appl Microbiol Biotechnol. 2002. 58: 275–85.
CAS
Статья
Google Scholar
2.
ЛеБлан Дж. Г., Милани С., де Джорджи Г. С., Сесма Ф., ван Синдерен Д., Вентура М. Бактерии как поставщики витаминов для своего хозяина: взгляд на микробиоту кишечника. Curr Opin Biotechnol. 2013; 24: 160–8.
CAS
Статья
Google Scholar
3.
Моровиц М.Дж., Карлайл Е.М., Алверди Дж.С. Вклад кишечных бактерий в питание и обмен веществ у тяжелобольных. Surg Clin North Am. 2011; 91: 771–85 (viii) .
Артикул
Google Scholar
4.
Паттерсон Э., Крайан Дж. Ф., Фицджеральд Г. Ф., Росс Р. П., Динан Т. Г., Стэнтон С. Микробиота кишечника, производимые ими фармакологические препараты и обеспечивающие здоровье. Proc Nutr Soc. 2014; 73: 477–89.
CAS
Статья
Google Scholar
5.
Блаут М., Клавель Т. Метаболическое разнообразие кишечной микробиоты: последствия для здоровья и болезней. J Nutr. 2007; 137: 751С – 5С.
CAS
Google Scholar
6.
Клаус С.П., Цанг Т.М., Ван И, Клорек О., Скорди Е., Мартин Ф.П., Реззи С., Росс А., Кочхар С., Холмс Е., Николсон Дж. Системные многокомпонентные эффекты микробиома кишечника на метаболические фенотипы мышей. Mol Syst Biol. 2008; 4: 219.
Артикул
Google Scholar
7.
Gobbetti M, Cagno RD, De Angelis M. Функциональные микроорганизмы для обеспечения функционального качества пищевых продуктов. Crit Rev Food Sci Nutr. 2010; 50: 716–27.
CAS
Статья
Google Scholar
8.
Бао З., Чи Ю. Оценка in vitro и in vivo ингибирующей активности ферментированного соевого молока (АПФ) ангиотензинпревращающего фермента с помощью штаммов Lactobacillus casei . Curr Microbiol. 2016; 73: 214–9.
CAS
Статья
Google Scholar
9.
Wu Q, Shah NP. Высокое производство гамма-аминомасляной кислоты из молочнокислых бактерий: акцент на Lactobacillus brevis как на функциональную закваску для молочных продуктов. Crit Rev Food Sci Nutr. 2016. DOI: 10.1080 / 10408398.2016.1147418.
10.
Де Ангелис М., Калассо М., Кавалло Н., Ди Каньо Р., Гоббетти М. Функциональная протеомика в пределах рода Lactobacillus. Протеомика. 2016; 16: 946–62.
Артикул
Google Scholar
11.
Milesi MM, McSweeney PL, Hynes ER.Жизнеспособность и вклад в протеолиз дополнительной культуры Lactobacillus plantarum в двух модельных сырных системах: сыр чеддер и мягкий сыр. J Appl Microbiol. 2008; 105: 884–92.
CAS
Статья
Google Scholar
12.
Риццелло К.Г., Лоруссо А., Монтемурро М., Гоббетти М. Использование закваски из муки киноа ( Chenopodium quinoa ) и отобранных автохтонных молочнокислых бактерий для улучшения пищевых, текстурных и сенсорных свойств белого хлеба.Food Microbiol. 2016; 56: 1–13.
CAS
Статья
Google Scholar
13.
Лезер Т.Д., Мольбак Л. Повышение уровня жизни благодаря действию микробов: преимущества микробиоты желудочно-кишечного тракта млекопитающих для хозяина. Environ Microbiol. 2009; 11: 2194–206.
CAS
Статья
Google Scholar
14.
Tremaroli V, Backhed F. Функциональные взаимодействия между кишечной микробиотой и метаболизмом хозяина.Природа. 2012; 489: 242–9.
CAS
Статья
Google Scholar
15.
Иноуэ Д., Цудзимото Г., Кимура И. Регулирование энергетического гомеостаза с помощью GPR41. Фронт-эндокринол (Лозанна). 2014; 5: 81.
Google Scholar
16.
Бугаут М. Возникновение, абсорбция и метаболизм короткоцепочечных жирных кислот в пищеварительном тракте млекопитающих. Comp Biochem Physiol B. 1987; 86: 439–72.
CAS
Статья
Google Scholar
17.
Хавенаар Р. Функции кишечного здоровья микробных метаболитов толстой кишки: обзор. Benef Microbes. 2011; 2: 103–14.
CAS
Статья
Google Scholar
18.
Serpa J, Caiado F, Carvalho T, Torre C, Goncalves LG, Casalou C, Lamosa P, Rodrigues M, Zhu Z, Lam EW, Dias S. Богатая бутиратом микросреда толстой кишки является важным фактором отбора для метаболически адаптированные опухолевые клетки.J Biol Chem. 2010; 285: 39211–23.
CAS
Статья
Google Scholar
19.
den Besten G, van Eunen K, Groen AK, Venema K, Reijngoud DJ, Bakker BM. Роль короткоцепочечных жирных кислот во взаимодействии между диетой, кишечной микробиотой и энергетическим обменом хозяина. J Lipid Res. 2013; 54: 2325–40.
Артикул
Google Scholar
20.
Бергман ЭН. Вклад в энергию летучих жирных кислот из желудочно-кишечного тракта у различных видов.Physiol Rev.1990; 70: 567–90.
CAS
Google Scholar
21.
ден Бестен Г., Ланге К., Хавинга Р., ван Дейк Т.Х., Гердинг А., ван Эйнен К., Мюллер М., Гроен А.К., Хойвельд Г.Дж., Баккер Б.М., Рейнгуд Д.Д. Полученные из кишечника короткоцепочечные жирные кислоты быстро ассимилируются в углеводы и липиды хозяина. Am J Physiol Gastrointest Liver Physiol. 2013; 305: G900–10.
Артикул
Google Scholar
22.
Кимура И., Иноуэ Д., Хирано К., Цудзимото Г. Рецептор SCFA GPR43 и энергетический метаболизм. Фронт-эндокринол (Лозанна). 2014; 5: 85.
Google Scholar
23.
Кимура И., Одзава К., Иноуэ Д., Имамура Т., Кимура К., Маэда Т., Терасава К., Кашихара Д., Хирано К., Тани Т. и др. Микробиота кишечника подавляет опосредованное инсулином накопление жира через рецептор короткоцепочечных жирных кислот GPR43. Nat Commun. 1829; 2013: 4.
Miquel S, Martin R, Rossi O, Bermudez-Humaran LG, Chatel JM, Sokol H, Thomas M, Wells JM, Langella P. Faecalibacterium prausnitzii и здоровье кишечника человека. Curr Opin Microbiol. 2013; 16: 255–61.
CAS
Статья
Google Scholar
26.
Sokol H, Pigneur B, Watterlot L, Lakhdari O, Bermudez-Humaran LG, Gratadoux JJ, Blugeon S, Bridonneau C, Furet JP, Corthier G, et al. Faecalibacterium prausnitzii — это противовоспалительная комменсальная бактерия, выявленная при анализе микробиоты кишечника пациентов с болезнью Крона. Proc Natl Acad Sci USA. 2008. 105: 16731–6.
CAS
Статья
Google Scholar
27.
Pessione E. Вклад молочнокислых бактерий в сложность кишечной микробиоты: свет и тень.Front Cell Infect Microbiol. 2012; 2: 86.
Меймандипур А., Хаир-Беджо М., Шухайми М., Азхар К., Сулеймани А.Ф., Расти Б., Язид А.М. Морфологические изменения желудочно-кишечного тракта из-за неприятного физического воздействия и модулирующей роли Lactobacillus у бройлеров.Br Poult Sci. 2010; 51: 52–9.
van Zanten GC, Knudsen A, Roytio H, Forssten S, Lawther M, Blennow A, Lahtinen SJ, Jakobsen M, Svensson B., Jespersen L.Влияние выбранных синбиотиков на микробный состав и продукцию короткоцепочечных жирных кислот в модельной системе толстой кишки человека. PLoS One. 2012; 7: e47212.
Артикул
Google Scholar
32.
Мацуки Т., Педрон Т., Ренье Б., Мулет С., Хара Т., Сансонетти П.Дж. Остановка пролиферации эпителиальных клеток, индуцированная лактатом и ацетатом из Lactobacillus casei и Bifidobacterium breve . PLoS One. 2013; 8: e63053.
CAS
Статья
Google Scholar
33.
Салазар Н., Бинетти А., Геймонд М., Алонсо А., Гарридо П., Гонсалес дель Рей С., де лос Рейес-Гавилан К. Г., Гонсалес С., Руас-Мадиедо П., де лос-Рейес-Гавилан К. Г.. Безопасность и модуляция кишечной микробиоты штаммами, продуцирующими экзополисахариды, Bifidobacterium animalis IPLA R1 и Bifidobacterium longum IPLA E44, вводимые перорально крысам Wistar. Int J Food Microbiol.2011; 144: 342–51.
Артикул
Google Scholar
34.
Амаретти А., Бернарди Т., Тамбурини Е., Занони С., Ломма М., Маттеуцци Д., Росси М. Кинетика и метаболизм Bifidobacterium adolescentis MB 239, растущего на глюкозе, галактозе, лактозе и галактоолигосахаридах. Appl Environ Microbiol. 2007. 73: 3637–44.
CAS
Статья
Google Scholar
35.
Абдин А.А., Саид Э.М.Экспериментальное исследование язвенного колита как потенциальной мишени для пробиотической терапии Lactobacillus acidophilus с или без «олсалазина». Колит Дж. Крона. 2008; 2: 296–303.
Артикул
Google Scholar
36.
Ван ден Аббеле П., Жерар П., Работ С., Бруно А., Эль-Эйди С., Дерриен М., Клееребезем М., Зоетендал Е. Г., Смидт Н., Верстраете В. и др. Арабиноксиланы и инулин по-разному модулируют микробиоту слизистой и просвета кишечника, а также разложение муцина у гуманизированных крыс.Environ Microbiol. 2011; 13: 2667–80.
Артикул
Google Scholar
37.
Лонсдейл Д. Обзор биохимии, метаболизма и клинических преимуществ тиамина (е) и его производных. Evid Based Complement Alternat Med. 2006; 3: 49–59.
Артикул
Google Scholar
38.
Карденас Н., Лайно Дж. Э., Дельгадо С., Хименес Е., Хуарес-дель-Валле М., Савой-де-Джорджи Г., Сесма Ф., Савой де Джорджи Г., Майо Б., Фернандес Л., Леблан Дж. Г., Родригес Дж. М..Связь между геномом и некоторыми фенотипическими свойствами Lactobacillus fermentum CECT 5716, пробиотического штамма, выделенного из грудного молока. Appl Microbiol Biotechnol. 2015; 99: 4343–53.
CAS
Статья
Google Scholar
39.
LeBlanc JG, Laino JE, Juarez del Valle M, Vannini V, van Sinderen D, Taranto MP, Font de Valdez G, Savoy de Giori G, Sesma F. Производство витаминов группы B молочнокислыми бактериями— текущие знания и потенциальные приложения.J Appl Microbiol. 2011; 111: 1297–309.
CAS
Статья
Google Scholar
40.
Pacheco Da Silva FF, Biscola V, LeBlanc JG, Gombossy de Melo Franco BD. Влияние аборигенных молочнокислых бактерий, выделенных из козьего молока и сыров, на содержание фолиевой кислоты и рибофлавина в ферментированном козьем молоке. LWT-Food Sci Technol. 2016; 71: 155–61.
CAS
Статья
Google Scholar
41.
дель Хуарес Валле М, дель Валле Хуарес, Лаиньо Дж. Э., де Савой Джорджи Дж., Леблан Дж. Дж. Молочнокислые бактерии, продуцирующие рибофлавин, как биотехнологическая стратегия получения биообогащенного соевого молока. Food Res Int. 2014; 62: 1015–9.
Артикул
Google Scholar
42.
Laiño JE, LeBlanc JG, Savoy de Giori G. Производство натуральных фолатов заквасочными культурами молочнокислых бактерий, выделенными из аргентинских йогуртов, приготовленных вручную. Может J Microbiol.2012; 58: 581–8.
Артикул
Google Scholar
43.
Salvucci E, LeBlanc JG, Pérez G. Технологические свойства молочнокислых бактерий, выделенных из сырого зернового материала. LWT-Food Sci Technol. 2016; 70: 185–91.
CAS
Статья
Google Scholar
44.
Resta SC. Влияние пробиотиков и комменсалов на физиологию кишечного эпителия: последствия для обработки питательных веществ.J Physiol. 2009; 587: 4169–74.
CAS
Статья
Google Scholar
45.
Клеребезем М., Бекхорст Дж., Ван Краненбург Р., Моленаар Д., Койперс О.П., Лир Р., Тарчини Р., Петерс С.А., Сандбринк Х.М., Фирс М.В. и др. Полная последовательность генома Lactobacillus plantarum WCFS1. Proc Natl Acad Sci USA. 2003; 100: 1990–5.
CAS
Статья
Google Scholar
46.
Santos F, Wegkamp A, de Vos WM, Smid EJ, Hugenholtz J. Производство высокого уровня фолиевой кислоты в ферментированных пищевых продуктах производителем B12 Lactobacillus reuteri JCM1112. Appl Environ Microbiol. 2008; 74: 3291–4.
CAS
Статья
Google Scholar
47.
Wegkamp A, de Vos WM, Smid EJ. Избыточное производство фолиевой кислоты в Lactobacillus plantarum WCFS1 вызывает резистентность к метотрексату. FEMS Microbiol Lett. 2009; 297: 261–5.
CAS
Статья
Google Scholar
48.
Magnúsdóttir S, Ravcheev D, de Crécy-Lagard V, Thiele I. Систематическая оценка генома биосинтеза витамина B предполагает сотрудничество между кишечными микробами. Fronti Genet. 2015; 6: 148.
Google Scholar
49.
Brestoff JR, Artis D. Комменсальные бактерии на стыке метаболизма хозяина и иммунной системы. Nat Immunol.2013; 14: 676–84.
CAS
Статья
Google Scholar
50.
Beitāne I, Ciproviča I, Gaile Z, Kaķitis A, Dumbrauskas A, Alsins J, Bernhard A. Изменения концентраций тиамина и рибофлавина в молоке, обогащенном пребиотиками и пробиотиками. В: Исследования для развития сельских районов: Материалы международной научной конференции, Елгава, Латвия, 19–22 мая 2006 г. Латвийский сельскохозяйственный университет; 2006 г., стр. 201–204.
51.
Hou JW, Yu RC, Chou CC. Изменения некоторых компонентов соевого молока при ферментации бифидобактериями. Food Res Int. 2000; 33: 393–7.
CAS
Статья
Google Scholar
52.
Кнайфель В., Кауфманн М., Флейшер А., Ульберт Ф. Скрининг коммерчески доступных мезофильных молочных заквасок: биохимические, сенсорные и микробиологические свойства. J Dairy Sci. 1992; 75: 3158–66.
Артикул
Google Scholar
53.
Vogel RF, Pavlovic M, Ehrmann MA, Wiezer A, Liesegang H, Offschanka S, Voget S, Angelov A, Bocker G, Liebl W. Геномный анализ выявил Lactobacillus sanfranciscensis как стабильный элемент в традиционных заквасках. Факт о микробной клетке. 2011; 10 (Приложение 1): S6.
Артикул
Google Scholar
54.
Масуда М., Идем М., Утсуми Х., Нииро Т., Шимамура Ю., Мурата М. Эффективность продукции фолиевой кислоты, витамина B (12) и тиамина молочнокислыми бактериями, выделенными из японских солений.Biosci Biotechnol Biochem. 2012; 76: 2061–7.
CAS
Статья
Google Scholar
55.
Вера Пингиторе Э., Хеберт Э.М., Сесма Ф., Надер-Масиас МЭ. Влияние витаминов и осмолитов на рост и продукцию бактериоцина Lactobacillus salivarius CRL 1328 в среде определенного химического состава. Может J Microbiol. 2009; 55: 304–10.
Артикул
Google Scholar
56.
Бегли Т.П., Даунс Д.М., Илик С.Е., Маклафферти Ф.В., Ван Лун А.П., Тейлор С., Кампобассо Н., Чиу Х.-Дж., Кинсленд С., Реддик Дж. Дж. Биосинтез тиамина у прокариот. Arch Microbiol. 1999; 171: 293–300.
CAS
Статья
Google Scholar
57.
Saulnier DM, Santos F, Roos S, Mistretta TA, Spinler JK, Molenaar D, Teusink B, Versalovic J. Изучение реконструкции метаболических путей и профилирования экспрессии по всему геному в Lactobacillus reuteri для определения функциональных пробиотических свойств .PLoS One. 2011; 6: e18783.
CAS
Статья
Google Scholar
58.
Teusink B, van Enckevort FH, Francke C, Wiersma A, Wegkamp A, Smid EJ, Siezen RJ. Реконструкция in silico метаболических путей Lactobacillus plantarum: сравнение прогнозов потребности в питательных веществах с результатами экспериментов по выращиванию. Appl Environ Microbiol. 2005. 71: 7253–62.
CAS
Статья
Google Scholar
59.
Перкинс Дж. Б., Перо Дж. Биосинтез витаминов. В: Sonenshein A, Hoch J, Losick R, редакторы. Bacillus subtilis и его ближайшие родственники от генов до клеток. Вашингтон: ASM Press; 2002. с. 271–86.
Google Scholar
60.
Берджесс К.М., О’Коннелл-Мазервей М., Сибесма В., Хугенгольц Дж., Ван Синдерен Д. Производство рибофлавина в Lactococcus lactis : потенциал для производства на месте обогащенных витаминами пищевых продуктов.Appl Environ Microbiol. 2004. 70: 5769–77.
Берджесс С.М., Смид Э.Дж., Руттен Дж., Ван Синдерен Д. Общий метод отбора микроорганизмов пищевого качества, продуцирующих избыточно рибофлавин. Факт о микробной клетке. 2006; 5: 24.
Артикул
Google Scholar
63.
Берджесс С.М., Смид Э.Дж., ван Синдерен Д. Избыточное производство бактериальных витаминов B2, B11 и B12: обзор. Int J Food Microbiol. 2009; 133: 1–7.
CAS
Статья
Google Scholar
64.
Capozzi V, Menga V, Digesu AM, De Vita P, van Sinderen D, Cattivelli L, Fares C, Spano G. Биотехнологическое производство обогащенного витамином B2 хлеба и макаронных изделий. J Agric Food Chem 2011, ( In press ).
65.
Capozzi V, Russo P, Dueñas MT, López P, Spano G. Молочнокислые бактерии, продуцирующие витамины группы B: большой потенциал для функциональных зерновых продуктов. Appl Microbiol Biotechnol. 2012; 96: 1383–94.
CAS
Статья
Google Scholar
66.
de Moreno de LeBlan A, Chaves S, Perdigon G. Влияние йогурта на профиль цитокинов с использованием мышиной модели воспаления кишечника. Eur J Inflamm. 2009; 7: 97–109.
Google Scholar
67.
LeBlanc JG, Burgess C, Sesma F, de Giori GS, van Sinderen D. Lactococcus lactis способен улучшать статус рибофлавина у крыс с дефицитом. Br J Nutr. 2005; 94: 262–7.
CAS
Статья
Google Scholar
68.
LeBlanc JG, Burgess C, Sesma F, Savoy de Giori G, van Sinderen D. Употребление молока, ферментированного генетически модифицированными Lactococcus lactis , улучшает статус рибофлавина у крыс с дефицитом. J Dairy Sci. 2005; 88: 3435–42.
CAS
Статья
Google Scholar
69.
ЛеБлан Дж. Г., Руттен Дж., Брюненберг П., Сесма Ф., де Джорджи Г. С., Смид Э. Дж. Новый молочный продукт, ферментированный Propionibacterium freudenreichii , улучшает статус рибофлавина у крыс с дефицитом.Питание. 2006; 22: 645–51.
CAS
Статья
Google Scholar
70.
LeBlanc JG, Savoy de Giori G, Smid EJ, Hugenholtz J, Sesma F. Производство фолиевой кислоты молочнокислыми бактериями и другими пищевыми микроорганизмами. В: Мендес-Вилас А., редактор. Информирование о текущих исследованиях и образовательных темах и тенденциях в прикладной микробиологии, т. 1. Бадахос: Исследовательский центр Formatex; 2007. с. 329–39.
Google Scholar
71.
Sybesma W, Burgess C, Starrenburg M, van Sinderen D, Hugenholtz J. Производство поливитаминов в Lactococcus lactis с использованием метаболической инженерии. Metab Eng. 2004; 6: 109–15.
CAS
Статья
Google Scholar
72.
Sybesma W, LeBlanc JG, Starrenburg M, Sesma F, Savoy G, de Vos WM, Hugenholtz J. Lactococcus lactis , разработанный для производства фолиевой кислоты, в качестве источника биодоступных фолатов. В: Sybesma W, редактор.Метаболическая инженерия производства фолиевой кислоты в молочнокислых бактериях. Нидерланды: Университет Вагенингена; 2003. с. 193–212.
Google Scholar
73.
Sybesma W, Starrenburg M, Kleerebezem M, Mierau I, de Vos WM, Hugenholtz J. Повышенное производство фолиевой кислоты с помощью метаболической инженерии Lactococcus lactis . Appl Environ Microbiol. 2003. 69: 3069–76.
CAS
Статья
Google Scholar
74.
Wegkamp A, Starrenburg M, de Vos WM, Hugenholtz J, Sybesma W. Преобразование Lactobacillus gasseri , потребляющих фолат, в продуцент фолиевой кислоты. Appl Environ Microbiol. 2004; 70: 3146–8.
CAS
Статья
Google Scholar
75.
Laiño JE, Zelaya H, del Valle MJ, de Giori GS, LeBlanc JG. Молоко, ферментированное выбранными штаммами молочнокислых бактерий, способно улучшить фолатный статус грызунов с дефицитом фолиевой кислоты, а также предотвратить дефицит фолиевой кислоты.J Funct Foods. 2015; 17: 22–32.
Артикул
Google Scholar
76.
LeBlanc JG, Sybesma W, Starrenburg M, Sesma F, de Vos WM, de Giori GS, Hugenholtz J. Добавка с модифицированным Lactococcus lactis улучшает статус фолиевой кислоты у крыс с дефицитом. Питание. 2010; 26: 835–41.
CAS
Статья
Google Scholar
77.
Мартин Р., Оливарес М., Марин М.Л., Фернандес Л., Ксаус Дж., Родригес Дж. М..Пробиотический потенциал 3 штаммов лактобацилл, выделенных из грудного молока. J Hum Lact. 2005; 21: 8–17 (тест 18–21, 41) .
Артикул
Google Scholar
78.
Хименес Э., Ланга С., Мартин В., Арройо Р., Мартин Р., Фернандес Л., Родригес Дж. М.. Полная последовательность генома Lactobacillus fermentum CECT 5716, пробиотического штамма, выделенного из грудного молока. J Bacteriol. 2010; 192: 4800.
CAS
Статья
Google Scholar
79.
Gil-Campos M, Lopez MA, Rodriguez-Benitez MV, Romero J, Roncero I., Linares MD, Maldonado J, Lopez-Huertas E, Berwind R, Ritzenthaler KL, et al. Lactobacillus fermentum CECT 5716 безопасен и хорошо переносится детьми в возрасте 1–6 месяцев: рандомизированное контролируемое исследование. Pharmacol Res. 2012; 65: 231–8.
Артикул
Google Scholar
80.
Olivares M, Diaz-Ropero MP, Sierra S, Lara-Villoslada F, Fonolla J, Navas M, Rodriguez JM, Xaus J.Пероральный прием Lactobacillus fermentum CECT5716 усиливает эффекты вакцинации против гриппа. Питание. 2007. 23: 254–60.
CAS
Статья
Google Scholar
81.
Maldonado J, Canabate F, Sempere L, Vela F, Sanchez AR, Narbona E, Lopez-Huertas E, Geerlings A, Valero AD, Olivares M, Lara-Villoslada F. Пробиотик грудного молока Lactobacillus fermentum CECT5716 снижает частоту инфекций желудочно-кишечного тракта и верхних дыхательных путей у младенцев.J Pediatr Gastroenterol Nutr. 2012; 54: 55–61.
Артикул
Google Scholar
82.
Jungersen M, Wind A, Johansen E, Christensen JE, Stuer-Lauridsen B, Eskesen D. Наука, лежащая в основе пробиотического штамма Bifidobacterium animalis subsp. lactis BB-12 ® . Микроорганизмы. 2014; 2: 92–110.
CAS
Статья
Google Scholar
83.
Strozzi GP, Mogna L.Количественное определение фолиевой кислоты в кале человека после введения пробиотических штаммов Bifidobacterium. J Clin Gastroenterol. 2008; 42 (Дополнение 3, часть 2): S179–84.
CAS
Статья
Google Scholar
84.
Помпеи А., Кордиско Л., Амаретти А., Занони С., Маттеуцци Д., Росси М. Производство фолиевой кислоты бифидобактериями как потенциальное пробиотическое свойство. Appl Environ Microbiol. 2007. 73: 179–85.
CAS
Статья
Google Scholar
85.
Pompei A, Cordisco L, Amaretti A, Zanoni S, Raimondi S, Matteuzzi D, Rossi M. Введение фолат-продуцирующих бифидобактерий повышает статус фолиевой кислоты у крыс Wistar. J Nutr. 2007. 137: 2742–6.
CAS
Google Scholar
86.
Giloteaux L, Goodrich JK, Walters WA, Levine SM, Ley RE, Hanson MR. Уменьшение разнообразия и изменение состава микробиома кишечника у людей с миалгическим энцефаломиелитом / синдромом хронической усталости.Микробиом. 2016; 4:30.
Артикул
Google Scholar
87.
Лахан С.Е., Кирхгесснер А. Воспаление кишечника при синдроме хронической усталости. Нутр Метаб (Лондон). 2010; 7: 79.
Артикул
Google Scholar
88.
Laino JE, Juarez del Valle M, Savoy de Giori G, LeBlanc JG. Применимость штамма Lactobacillus amylovorus в качестве сокультуры для естественного биообогащения ферментированного молока фолиевой кислотой.Int J Food Microbiol. 2014; 191С: 10–6.
Артикул
Google Scholar
Модуляция микробных короткоцепочечных жирных кислот на гомеостаз кишечника, метаболизм и психоневрологические расстройства
Alfa MJ, Strang D, Tappia PS, Graham M, Van Domselaar G, Forbes JD, Laminman V, Olson N, DeGagne P, Bray D, Murray BL, Dufault B, Lix LM (2018) Рандомизированное испытание для определения влияния состава крахмала, устойчивого к перевариванию, на микробиом кишечника у людей старшего и среднего возраста.Clin Nutr 37: 797–807. https://doi.org/10.1016/j.clnu.2017.03.025
CAS
Статья
PubMed
Google Scholar
Arpaia N, Campbell C, Fan X, Dikiy S, van der Veeken J, deRoos P, Liu H, Cross JR, Pfeffer K, Coffer PJ, Rudensky AY (2013) Метаболиты, продуцируемые комменсальными бактериями, способствуют периферическому регуляторному T -генерация клеток. Природа 504: 451–455. https://doi.org/10.1038/nature12726
CAS
Статья
PubMed
PubMed Central
Google Scholar
Bajaj JS, Kakiyama G, Savidge T, Takei H, Kassam ZA, Fagan A, Gavis EA, Pandak WM, Nittono H, Hylemon PB, Boonma P, Haag A, Heuman DM, Fuchs M, John B, Sikaroodi M, Gillevet PM (2018) Нарушение состава и функции микробиоты при циррозе, связанное с антибиотиками, восстанавливается с помощью фекальной трансплантации.Гепатология 68: 1549–1558. https://doi.org/10.1002/hep.30037
CAS
Статья
PubMed
Google Scholar
Baxter NT, Schmidt AW, Venkataraman A, Kim KS, Waldron C, Schmidt TM (2019) Динамика микробиоты кишечника человека и короткоцепочечных жирных кислот в ответ на диетические вмешательства с использованием трех ферментируемых волокон. MBio 10: e02566 – e02518. https://doi.org/10.1128/mBio.02566-18
CAS
Статья
PubMed
PubMed Central
Google Scholar
Бегайдарова Р., Юхневич Ю., Бабенко Д., Калиева С., Азизов И., Мулдаева Г., Омаркулов Б. (2015) Определение экспрессии гена PFOR в штаммах G.кишечник с различными ингибирующими концентрациями метронидазола. J Infect Dev Ctries 9: 519–523. https://doi.org/10.3855/jidc.5768
CAS
Статья
PubMed
Google Scholar
Brandhorst S, Choi IY, Wei M, Cheng CW, Sedrakyan S, Navarrete G, Dubeau L, Yap LP, Park R, Vinciguerra M, Di Biase S, Mirzaei H, Mirisola MG, Childress P, Ji L, Groshen S, Penna F, Odetti P, Perin L, Conti PS, Ikeno Y, Kennedy BK, Cohen P, Morgan TE, Dorff TB, Longo VD (2015) Периодическая диета, имитирующая голодание, способствует регенерации нескольких систем и улучшению когнитивных функций , и продолжительность жизни.Cell Metab 22: 86–99. https://doi.org/10.1016/j.cmet.2015.05.012
CAS
Статья
PubMed
PubMed Central
Google Scholar
Budak NH, Aykin E, Seydim AC, Greene AK, Guzel-Seydim ZB (2014) Функциональные свойства уксуса. J Food Sci 79: R757 – R764. https://doi.org/10.1111/1750-3841.12434
CAS
Статья
PubMed
Google Scholar
Byndloss MX, Pernitzsch SR, Bäumler AJ (2018) Здоровые хозяева правят внутри: экологические силы, формирующие микробиоту кишечника.Mucosal Immunol 11: 1299–1305. https://doi.org/10.1038/s41385-018-0010-y
CAS
Статья
PubMed
Google Scholar
Canfora EE, van der Beek CM, Jocken JWE, Goossens GH, Holst JJ, Olde Damink SWM, Lenaerts K, Dejong CHC, Blaak EE (2017) Инфузии смесей короткоцепочечных жирных кислот в толстой кишке способствуют энергетическому метаболизму при избыточном весе / тучные мужчины: рандомизированное перекрестное исследование. Научный журнал 7: 2360. https://doi.org/10.1038/s41598-017-02546-x
Cantó C, Auwerx J (2010) AMP-активированная протеинкиназа и ее нисходящие пути транскрипции. Cell Mol Life Sci 67: 3407–3423. https://doi.org/10.1007/s00018-010-0454-z
Carlson JL, Erickson JM, Hess JM, Gould TJ, Slavin JL (2017) Пребиотические пищевые волокна и здоровье кишечника: сравнение in vitro ферментации бета-глюкана, инулина и ксилоолигосахарида. Питательные вещества 9: E1361. https://doi.org/10.3390/nu61
CAS
Статья
PubMed
Google Scholar
Caron A, Lee S, Elmquist JK, Gautron L (2018) Лептин и перекрестные помехи между мозгом и жиром.Nat Rev Neurosci 19: 153–165. https://doi.org/10.1038/nrn.2018.7
CAS
Статья
PubMed
PubMed Central
Google Scholar
Chang PV, Hao L, Offermanns S, Medzhitov R (2014) Микробный метаболит бутират регулирует функцию кишечных макрофагов посредством ингибирования гистондеацетилазы. Proc Natl Acad Sci U S A 111: 2247–2252. https://doi.org/10.1073/pnas.1322269111
CAS
Статья
PubMed
PubMed Central
Google Scholar
Christl SU, Murgatroyd PR, Gibson GR, Cummings JH (1992) Производство, метаболизм и выделение водорода в толстом кишечнике.Гастроэнтерология 102: 1269–1277
CAS
Статья
Google Scholar
Cussotto S, Sandhu KV, Dinan TG, Cryan JF (2018) Нейроэндокринология оси микробиота-кишечник-мозг: поведенческая перспектива. Фронт нейроэндокринол 51: 80–101. https://doi.org/10.1016/j.yfrne.2018.04.002
CAS
Статья
PubMed
Google Scholar
de Clercq NC, Frissen MN, Groen AK, Nieuwdorp M (2017) Микробиота кишечника и ось кишечник-мозг: новое понимание патофизиологии метаболического синдрома.Psychosom Med 79: 874–879. https://doi.org/10.1097/PSY.0000000000000495
CAS
Статья
PubMed
Google Scholar
De Vadder F, Kovatcheva-Datchary P, Goncalves D, Vinera J, Zitoun C, Duchampt A, Bäckhed F, Mithieux G (2014) Метаболиты, произведенные микробиотой, способствуют метаболическим преимуществам через нервные цепи кишечника и мозга. Cell 156: 84–96. https://doi.org/10.1016/j.cell.2013.12.016
CAS
Статья
PubMed
Google Scholar
De Vadder F, Kovatcheva-Datchary P, Zitoun C, Duchampt A, Bäckhed F, Mithieux G (2016) Сукцинат, произведенный микробиотой, улучшает гомеостаз глюкозы посредством кишечного глюконеогенеза.Cell Metab 24: 151–157. https://doi.org/10.1016/j.cmet.2016.06.013
CAS
Статья
PubMed
Google Scholar
den Besten G, Lange K, Havea R, van Dijk TH, Gerding A, van Eunen K, Müller M, Groen AK, Hooiveld GJ, Bakker BM, Reijngoud DJ (2013a) Короткоцепочечные жирные кислоты кишечного происхождения быстро усваиваются углеводами и липидами хозяина. Am J Physiol Gastrointest Liver Physiol 305: G900 – G910. https://doi.org/10.1152 / ajpgi.00265.2013
CAS
Статья
Google Scholar
den Besten G, van Eunen K, Groen AK, Venema K, Reijngoud DJ, Bakker BM (2013b) Роль короткоцепочечных жирных кислот во взаимодействии между диетой, микробиотой кишечника и энергетическим метаболизмом хозяина. J. Lipid Res., 54: 2325–2340. https://doi.org/10.1194/jlr.R036012
CAS
Статья
Google Scholar
Di Sabatino A, Morera R, Ciccocioppo R, Cazzola P, Gotti S, Tinozzi FP, Tinozzi S, Corazza GR (2005) Пероральный бутират для лечения болезни Крона средней и средней активности.Aliment Pharmacol Ther 22: 789–794
Duncan SH, Barcenilla A, Stewart CS, Pryde SE, Flint HJ (2002) Использование ацетата и бутирилкофермент a (CoA): ацетат-CoA трансфераза в бактериях, продуцирующих бутират, из толстой кишки человека.Appl Environ Microbiol 68: 5186–5190. https://doi.org/10.1128/aem.68.10.5186-5190.2002
CAS
Статья
PubMed
PubMed Central
Google Scholar
Duncan SH, Louis P, Flint HJ (2004) Утилизирующие лактат бактерии, выделенные из человеческих фекалий, которые производят бутират в качестве основного продукта ферментации. Appl Environ Microbiol 70: 5810–5817. https://doi.org/10.1128/AEM.70.10.5810-5817.2004
CAS
Статья
PubMed
PubMed Central
Google Scholar
El Oufir L, Flourié B, Bruley des Varannes S, Barry JL, Cloarec D, Bornet F, Galmiche JP (1996) Взаимосвязь между временем прохождения, продуктами ферментации и флорой, потребляющей водород, у здоровых людей.Gut 38: 870–877
Статья
Google Scholar
Эрни Д., Грабе-де-Ангелис А.Л., Яитин Д., Вигхофер П., Сташевский О., Давид Е., Керен-Шауль Н., Махлаков Т., Якобсхаген К., Бух Т., Швижецек В., Утермолен О., Чун Е., Гарретт В.С., McCoy KD, Diefenbach A, Staeheli P, Stecher B, Amit I, Prinz M (2015) Микробиота хозяина постоянно контролирует созревание и функцию микроглии в ЦНС. Nat Neurosci 18: 965–977. https://doi.org/10.1038/nn.4030
CAS
Статья
PubMed
PubMed Central
Google Scholar
Fachi JL, Felipe JS, Pral LP, da Silva BK, Corrêa RO, de Andrade MCP, da Fonseca DM, Basso PJ, Câmara NOS, de Sales Souza EL E, Dos Santos Martins F, Guima SES, Thomas AM , Setubal JC, Magalhães YT, Forti FL, Candreva T, Rodrigues HG, de Jesus MB, Consonni SR, ADS F, Varga-Weisz P, Vinolo MAR (2019) Бутират защищает мышей от колита, вызванного clostridium difficile, , посредством HIF. -1-зависимый механизм.Cell Rep 27: 750–761.e7. https://doi.org/10.1016/j.celrep.2019.03.054
CAS
Статья
PubMed
Google Scholar
Fernández J, Redondo-Blanco S, Gutiérrez-del-Río I, Miguélez EM, Villar CJ, Lombó F (2016) Ферментация диетических пребиотиков микробиотой толстой кишки в сторону короткоцепочечных жирных кислот и их роль в качестве противовоспалительных и противоопухолевые средства: обзор. J Funct Foods 25: 511–522. https://doi.org/10.1016/j.jff.2016.06.032
CAS
Статья
Google Scholar
Ferreira-Lazarte A, Moreno FJ, Cueva C, Gil-Sánchez I, Villamiel M (2019) Поведение цитрусового пектина во время его желудочно-кишечного переваривания и ферментации в динамическом симуляторе (simgi®). Carbohydr Polym 207: 382–390. https://doi.org/10.1016/j.carbpol.2018.11.088
CAS
Статья
PubMed
Google Scholar
Flint HJ, Scott KP, Louis P, Duncan SH (2012a) Роль кишечной микробиоты в питании и здоровье.Нат Рев Гастроэнтерол Гепатол 9: 577–589. https://doi.org/10.1038/nrgastro.2012.156
CAS
Статья
PubMed
Google Scholar
Flint HJ, Scott KP, Duncan SH, Louis P, Forano E (2012b) Микробное разложение сложных углеводов в кишечнике. Кишечные микробы 3: 289–306. https://doi.org/10.4161/gmic.19897
Артикул
PubMed
PubMed Central
Google Scholar
Gao Z, Yin J, Zhang J, Ward RE, Martin RJ, Lefevre M, Cefalu WT, Ye J (2009) Бутират улучшает чувствительность к инсулину и увеличивает расход энергии у мышей.Диабет 58: 1509–1517. https://doi.org/10.2337/db08-1637
CAS
Статья
PubMed
PubMed Central
Google Scholar
Gao B, Wang R, Peng Y, Li X (2018a) Влияние гомогенного полисахарида из отвара Sijunzi на микробы кишечника человека и короткоцепочечные жирные кислоты in vitro . Дж. Этнофармакол 224: 465–473. https://doi.org/10.1016/j.jep.2018.06.006
CAS
Статья
PubMed
Google Scholar
Gao H, Wen JJ, Hu JL, Nie QX, Chen HH, Xiong T, Nie SP, Xie MY (2018b) Полисахарид из ферментированного Momordica charantia L.с Lactobacillus plantarum NCU116 улучшает диабет 2 типа у крыс. Carbohydr Polym 201: 624–633. https://doi.org/10.1016/j.carbpol.2018.08.075
CAS
Статья
PubMed
Google Scholar
Gill RK, Saksena S, Alrefai WA, Sarwar Z, Goldstein JL, Carroll RE, Ramaswamy K, Dudeja PK (2005) Экспрессия и мембранная локализация изоформ MCT по длине кишечника человека. Am J Phys Cell Phys 289: C846 – C852.https://doi.org/10.1152/ajpcell.00112.2005
CAS
Статья
Google Scholar
Gill PA, van Zelm MC, Muir JG, Gibson PR (2018) Обзорная статья: короткоцепочечные жирные кислоты как потенциальные терапевтические агенты при желудочно-кишечных и воспалительных заболеваниях человека. Алимент Pharmacol Ther 48: 15–34. https://doi.org/10.1111/apt.14689
CAS
Статья
PubMed
Google Scholar
Gomes-Neto JC, Round JL (2018) Микробиота кишечника: новый способ приема витаминов.Нат Рев Гастроэнтерол Гепатол 15: 521–522. https://doi.org/10.1038/s41575-018-0044-3
CAS
Статья
PubMed
Google Scholar
Guo WL, Pan YY, Li L, Li TT, Liu B, Lv XC (2018) Этаноловый экстракт Ganoderma lucidum улучшает нарушения липидного обмена и модулирует состав микробиоты кишечника у крыс, получавших диету с высоким содержанием жиров. Food Funct 9: 3419–3431. https://doi.org/10.1039/c8fo00836a
CAS
Статья
PubMed
Google Scholar
Hasebe T, Ueno N, Musch MW, Nadimpalli A, Kaneko A, Kaifuchi N, Watanabe J, Yamamoto M, Kono T, Inaba Y, Fujiya M, Kohgo Y, Chang EB (2016) Daikenchuto (TU-100 ) формирует архитектуру микробиоты кишечника и увеличивает выработку соединения-метаболита гинсенозида K.Pharmacol Res Perspect 4: e00215. https://doi.org/10.1002/prp2.215
CAS
Статья
PubMed
PubMed Central
Google Scholar
Henning SM, Yang J, Hsu M, Lee RP, Grojean EM, Ly A, Tseng CH, Heber D, Li Z (2018) Полифенолы зеленого и черного чая без кофеина уменьшают прибавку в весе и изменяют популяции и функции микробиома в рационе -индуцированные ожирением мышей. Eur J Nutr 57: 2759–2769. https://doi.org/10.1007/s00394-017-1542-8
CAS
Статья
PubMed
Google Scholar
Henningsson AM, Björck IM, Nyman EM (2002) Комбинации неперевариваемых углеводов влияют на образование короткоцепочечных жирных кислот в заднем кишечнике крыс.J Nutr 132: 3098–3104. https://doi.org/10.1093/jn/131.10.3098
CAS
Статья
PubMed
Google Scholar
Høverstad T, Midtvedt T (1986) Короткоцепочечные жирные кислоты у стерильных мышей и крыс. J Nutr 116: 1772–1776
Артикул
Google Scholar
Ji J, Shu D, Zheng M, Wang J, Luo C, Wang Y, Guo F, Zou X, Lv X, Li Y, Liu T, Qu H (2016) Бутират микробного метаболита способствует поляризации макрофагов M2 и функция.Sci Rep 6: 24838. https://doi.org/10.1038/srep24838
Kaur A, Chen T, Green SJ, Mutlu E, Martin BR, Rumpagaporn P, Patterson JA, Keshavarzian A, Hamaker BR (2019) Физическая недоступность резистентного крахмала сдвигает микробиоту кишечника мышей до бутирогенных фирмикутов.Mol Nutr Food Res 63: e1801012. https://doi.org/10.1002/mnfr.201801012
CAS
Статья
PubMed
Google Scholar
Khamsi R (2016) Метаболизм в уме: новые взгляды на «ось кишечник-мозг» стимулируют коммерческие усилия по его нацеливанию. Nat Med 22: 697–700. https://doi.org/10.1038/nm0716-697
CAS
Статья
PubMed
Google Scholar
Kim M, Qie Y, Park J, Kim CH (2016) Метаболиты кишечных микробов подпитывают ответы антител хозяина.Клеточный микроб-хозяин 20: 202–214. https://doi.org/10.1016/j.chom.2016.07.001
CAS
Статья
PubMed
PubMed Central
Google Scholar
Ким К.С., Сили Р.Дж., Сандовал Д.А. (2018) Передача сигналов от периферии к мозгу, которая регулирует энергетический гомеостаз. Нат Рев Neurosci 19: 185–196. https://doi.org/10.1038/nrn.2018.8
CAS
Статья
PubMed
Google Scholar
Kimura I, Inoue D, Maeda T, Hara T, Ichimura A, Miyauchi S, Kobayashi M, Hirasawa A, Tsujimoto G (2011) Короткоцепочечные жирные кислоты и кетоны напрямую регулируют симпатическую нервную систему через G-протеин. рецептор 41 (GPR41).Proc Natl Acad Sci U S A 108: 8030–8035. https://doi.org/10.1073/pnas.1016088108
Артикул
PubMed
PubMed Central
Google Scholar
Ко А., Де Ваддер Ф, Ковачева-Датчари П., Бекхед Ф. (2016) От пищевых волокон к физиологии хозяина: короткоцепочечные жирные кислоты как ключевые бактериальные метаболиты. Cell 165: 1332–1345. https://doi.org/10.1016/j.cell.2016.05.041
CAS
Статья
PubMed
Google Scholar
Кондо Т., Киши М., Фусими Т., Кага Т. (2009) Уксусная кислота усиливает экспрессию генов ферментов окисления жирных кислот в печени, подавляя накопление жира в организме.J. Agric Food Chem. 57: 5982–5986. https://doi.org/10.1021/jf0c
CAS
Статья
PubMed
Google Scholar
Kong XF, Zhang YZ, Wu X, Yin YL, Tan ZL, Feng Y, Yan FY, Bo MJ, Huang RL, Li TJ (2009) Характеристика ферментации полисахарида китайского ямса и его влияние на микробиоту кишечника крысы. Int J Microbiol 2009: 598152. https://doi.org/10.1155/2009/598152
CAS
Статья
PubMed
PubMed Central
Google Scholar
Lee YS, Kim TY, Kim Y, Lee SH, Kim S, Kang SW, Yang JY, Baek IJ, Sung YH, Park YY, Hwang SW, Eunju O, Kim KS, Liu S, Kamada N, Gao N, Kweon MN (2018) Лактат, полученный из микробиоты, ускоряет развитие эпителия, опосредованное стволовыми клетками кишечника.Клеточный микроб-хозяин 24: 833–846.e6. https://doi.org/10.1016/j.chom.2018.11.002
CAS
Статья
PubMed
Google Scholar
Li J, Zhu X, Chen J, Zhao D, Zhang X, Bi C (2017) Создание нового анаэробного пути в кишечной палочке Escherichia coli для производства пропионата. BMC Biotechnol 17:38. https://doi.org/10.1186/s12896-017-0354-5
CAS
Статья
PubMed
PubMed Central
Google Scholar
Liang Y, Zhang Y, Deng Y, Liang S, He Y, Chen Y, Liu C, Lin C, Han L, Tu G, Yang Q (2018) Отвар Чайху-Шуган-Сан модулирует дисбактериоз кишечных микробов и снимает хроническое метаболическое воспаление у крыс с НАЖБП через путь воспаления NLRP3.Evid Based Complement Alternat Med 2018: 93
. https://doi.org/10.1155/2018/93
Артикул
PubMed
PubMed Central
Google Scholar
Lin HV, Frassetto A, Kowalik EJ Jr, Nawrocki AR, Lu MM, Kosinski JR, Hubert JA, Szeto D, Yao X, Forrest G, Marsh DJ (2012) Бутират и пропионат защищают от ожирения, вызванного диетой, и регулируют гормоны кишечника через 3-независимые механизмы, независимые от рецептора свободных жирных кислот. PLoS One 7: e35240.https://doi.org/10.1371/journal.pone.0035240
CAS
Статья
PubMed
PubMed Central
Google Scholar
Лю З., Лю Т. (2016) Производство акриловой кислоты и пропионовой кислоты путем построения части цикла 3-гидроксипропионат / 4-гидроксибутират из Metallosphaera sedula в Escherichia coli . J Ind Microbiol Biotechnol 43: 1659–1670
CAS
Статья
Google Scholar
Liu J, Yue S, Yang Z, Feng W, Meng X, Wang A, Peng C, Wang C, Yan D (2018) Пероральный гидроксисафлор желтый a снижает ожирение у мышей, регулируя микробиоту кишечника и метаболизм сыворотки.Pharmacol Res 134: 40–50. https://doi.org/10.1016/j.phrs.2018.05.012
CAS
Статья
PubMed
Google Scholar
Louis P, Hold GL, Flint HJ (2014) Микробиота кишечника, бактериальные метаболиты и колоректальный рак. Nat Rev Microbiol 12: 661–672. https://doi.org/10.1038/nrmicro3344
CAS
Статья
PubMed
Google Scholar
Macia L, Tan J, Vieira AT, Leach K, Stanley D, Luong S, Maruya M, Ian McKenzie C, Hijikata A, Wong C, Binge L, Thorburn AN, Chevalier N, Ang C, Marino E, Роберт Р., Офферманн С., Тейшейра М.М., Мур Р.Дж., Флавелл Р.А., Фагарасан С., Маккей С.Р. (2015) Чувствительные к метаболиту рецепторы GPR43 и GPR109A способствуют индуцированному пищевым волокном гомеостазу кишечника посредством регуляции инфламмасомы.Нац Коммуна 6: 6734. https://doi.org/10.1038/ncomms7734
CAS
Статья
PubMed
Google Scholar
Martens EC, Neumann M, Desai MS (2018) Взаимодействие комменсальных и патогенных микроорганизмов со слизистой оболочкой кишечника. Nat Rev Microbiol 16: 457–470. https://doi.org/10.1038/s41579-018-0036-x
CAS
Статья
PubMed
Google Scholar
Макорист А.Л., Миллер Р.Б., Берд А.Р., Кеог Дж.Б., Ноукс М., Топпинг Д.Л., Конлон М.А. (2011) Уровни бутирата в фекалиях сильно различаются у разных людей, но обычно повышаются за счет диеты с высоким содержанием устойчивого крахмала.J Nutr 141: 883–889. https://doi.org/10.3945/jn.110.128504
CAS
Статья
PubMed
Google Scholar
Mei X, Zhang X, Wang Z, Gao Z, Liu G, Hu H, Zou L, Li X (2016) Активность флоризина, повышающая чувствительность к инсулину, связана со снижением липополисахаридов и изменениями микробиоты кишечника при ожирении и типе 2 мыши с диабетом (db / db). J. Agric Food Chem. 64: 7502–7511. https://doi.org/10.1021/acs.jafc.6b03474
CAS
Статья
PubMed
Google Scholar
Metges CC (2000) Вклад микробных аминокислот в аминокислотный гомеостаз хозяина.J Nutr 130: 1857S – 1864S. https://doi.org/10.1093/jn/130.7.1857S
CAS
Статья
PubMed
Google Scholar
Миллер Т.Л., Волин М.Дж. (1996) Пути образования ацетата, пропионата и бутирата фекальной микробной флорой человека. Appl Environ Microbiol 62: 1589–1592
CAS
PubMed
PubMed Central
Google Scholar
Миллс Э.Л., Пирс К.А., Едриховски М.П., Гаррити Р., Винтер С., Видони С., Йонеширо Т., Спинелли Дж. Б., Лу Г.З., Казак Л., Бэнкс А.С., Хейгис М.С., Каджимура С., Мерфи М.П., Гиги С.П., Клиш CB, Chouchani ET (2018) Накопление сукцината контролирует активацию термогенеза жировой ткани.Природа 560: 102–106. https://doi.org/10.1038/s41586-018-0353-2
CAS
Статья
PubMed
Google Scholar
Nagpal R, Shively CA, Appt SA, Register TC, Michalson KT, Vitolins MZ, Yadav H (2018a) Состав микробиома кишечника у нечеловеческих приматов, потребляющих западную или средиземноморскую диету. Передний гайковерт 5:28. https://doi.org/10.3389/fnut.2018.00028
CAS
Статья
PubMed
PubMed Central
Google Scholar
Nagpal R, Wang S, Ahmadi S, Hayes J, Gagliano J, Subashchandrabose S, Kitzman DW, Becton T, Read R, Yadav H (2018b) Пробиотический коктейль человеческого происхождения увеличивает производство короткоцепочечных жирных кислот за счет модуляции мышей и микробиома кишечника человека.Sci Rep 8: 12649. https://doi.org/10.1038/s41598-018-30114-4
CAS
Статья
PubMed
PubMed Central
Google Scholar
Nishitsuji K, Xiao J, Nagatomo R, Umemoto H, Morimoto Y, Akatsu H, Inoue K, Tsuneyama K (2017) Анализ микробиома кишечника и профилей короткоцепочечных жирных кислот в плазме на спонтанной модели метаболизма у мышей. синдром. Sci Rep 7: 15876–15810. https://doi.org/10.1038/s41598-017-16189-5
CAS
Статья
PubMed
PubMed Central
Google Scholar
Nøhr MK, Pedersen MH, Gille A, Egerod KL, Engelstoft MS, Husted AS, Sichlau RM, Grunddal KV, Poulsen SS, Han S, Jones RM, Offermanns S, Schwartz TW (2013) GPR41 / FFAR3 и GPR43 / FFAR2 в качестве сенсоров короткоцепочечных жирных кислот в энтероэндокринных клетках по сравнению с FFAR3 в кишечных нейронах и FFAR2 в энтеросолюбильных лейкоцитах.Эндокринология 154: 3552–3564. https://doi.org/10.1210/en.2013-1142
CAS
Статья
PubMed
Google Scholar
Осадчий В., Мартин С.Р., Майер Е.А. (2019) Ось кишечник-мозг и микробиом: механизмы и клинические последствия. Clin Gastroenterol Hepatol 17: 322–332. https://doi.org/10.1016/j.cgh.2018.10.002
CAS
Статья
PubMed
Google Scholar
Paramsothy S, Nielsen S, Kamm MA, Deshpande NP, Faith JJ, Clemente JC, Paramsothy R, Walsh AJ, van den Bogaerde J, Samuel D, Leong RWL, Connor S, Ng W, Lin E, Borody TJ , Wilkins MR, Colombel JF, Mitchell HM, Kaakoush NO (2019) Специфические бактерии и метаболиты, связанные с ответом на трансплантацию фекальной микробиоты у пациентов с язвенным колитом.Гастроэнтерология 156: 1440–1454.e2. https://doi.org/10.1053/j.gastro.2018.12.001
Артикул
PubMed
Google Scholar
Реймер Р.А. (2019) Установление роли диеты в оси микробиота-болезнь. Nat Rev Gastroenterol Hepatol 16: 86–87. https://doi.org/10.1038/s41575-018-0093-7
Артикул
PubMed
Google Scholar
Rey FE, Faith JJ, Bain J, Muehlbauer MJ, Stevens RD, Newgard CB, Gordon JI (2010) Анализ метаболического потенциала двух кишечных ацетогенов человека in vivo.J Biol Chem 285: 22082–22090. https://doi.org/10.1074/jbc.M110.117713
CAS
Статья
PubMed
PubMed Central
Google Scholar
Rojas OL, Pröbstel AK, Porfilio EA, Wang AA, Charabati M, Sun T, Lee DSW, Galicia G, Ramaglia V, Ward LA, Leung LYT, Najafi G, Khaleghi K, Garcillán B, Li A, Besla R, Naouar I, Cao EY, Chiaranunt P, Burrows K, Robinson HG, Allanach JR, Yam J, Luck H, Campbell DJ, Allman D, Brooks DG, Tomura M, Baumann R, Zamvil SS, Bar-Or A, Horwitz MS, Winer DA, Mortha A, Mackay F, Prat A, Osborne LC, Robbins C, Baranzini SE, Gommerman JL (2019). Рециркулирующие кишечные IgA-продуцирующие клетки регулируют нейровоспаление через IL-10.Cell 177: 492–493. https://doi.org/10.1016/j.cell.2019.03.037
CAS
Статья
PubMed
Google Scholar
Russell WR, Gratz SW, Duncan SH, Holtrop G, Ince J, Scobbie L, Duncan G, Johnstone AM, Lobley GE, Wallace RJ, Duthie GG, Flint HJ (2011) с высоким содержанием белка и пониженным содержанием углеводов — диеты с пониженным содержанием метаболитов способствуют развитию метаболитов, которые могут быть вредными для здоровья толстой кишки. Am J Clin Nutr 93: 1062–7102. https://doi.org/10.3945 / ajcn.110.002188
CAS
Статья
PubMed
Google Scholar
Samuel BS, Shaito A, Motoike T, Rey FE, Backhed F, Manchester JK, Hammer RE, Williams SC, Crowley J, Yanagisawa M, Gordon JI (2008) Воздействие микробиоты кишечника на ожирение хозяина модулируется короткоцепочечный рецептор, связанный с G-белком, связывающий жирные кислоты, Gpr41. Proc Natl Acad Sci U S A 105: 16767–16772. https://doi.org/10.1073/pnas.0808567105
Артикул
PubMed
PubMed Central
Google Scholar
Sandhu KV, Sherwin E, Schellekens H, Stanton C, Dinan TG, Cryan JF (2017) Кормление оси микробиота-кишечник-мозг: диета, микробиом и нейропсихиатрия.Перевод Рез. 179: 223–244. https://doi.org/10.1016/j.trsl.2016.10.002
CAS
Статья
PubMed
Google Scholar
Scarmeas N, Anastasiou CA, Yannakoulia M (2018) Питание и профилактика когнитивных нарушений. Lancet Neurol 17: 1006–1015. https://doi.org/10.1016/S1474-4422(18)30338-7
Артикул
PubMed
Google Scholar
Schneider KM, Albers S, Trautwein C (2018) Роль желчных кислот в системе кишечник-печень.J Hepatol 68: 1083–1085. https://doi.org/10.1016/j.jhep.2017.11.025
Артикул
PubMed
Google Scholar
Scopelliti A, Bauer C, Yu Y, Zhang T, Kruspig B, Murphy DJ, Vidal M, Maddocks ODK, Cordero JB (2019) Нейронное реле опосредует чувствительную к питательным веществам ось кишечника / жирового тела, регулирующую энергетический гомеостаз у взрослых дрозофила. Cell Metab 29: 269–284. https://doi.org/10.1016/j.cmet.2018.09.021
CAS
Статья
PubMed
PubMed Central
Google Scholar
Scott KP, Martin JC, Campbell G, Mayer CD, Flint HJ (2006) Профилирование полногеномной транскрипции выявляет гены, активируемые ростом фукозы в кишечной бактерии «Roseburia inulinivorans».J Bacteriol 188: 4340–4349. https://doi.org/10.1128/JB.00137-06
CAS
Статья
PubMed
PubMed Central
Google Scholar
Шарки К.А., Бек П.Л., Маккей Д.М. (2018) Нейроиммунофизиология кишечника: достижения и новые концепции с упором на эпителий. Нат Рев Гастроэнтерол Гепатол 15: 765–784. https://doi.org/10.1038/s41575-018-0051-4
Singh DP, Singh J, Boparai RK, Zhu J, Mantri S, Khare P, Khardori R, Kondepudi KK, Chopra K, Bishnoi M (2017) Изомальто-олигосахариды, пребиотик, функционально усиливают эффекты зеленого чая против высоких метаболические изменения, вызванные жировой диетой, путем предотвращения дисбактериоза кишечника у мышей. Pharmacol Res 123: 103–113.https://doi.org/10.1016/j.phrs.2017.06.015
CAS
Статья
PubMed
Google Scholar
Sorbara MT, Dubin K, Littmann ER, Moody TU, Fontana E, Seok R, Leiner IM, Taur Y, Peled JU, van den Brink MRM, Litvak Y, Bäumler AJ, Chaubard JL, Pickard AJ, Cross JR , Pamer EG (2019) Ингибирование устойчивых к антибиотикам Enterobacteriaceae за счет опосредованного микробиотой внутриклеточного подкисления. J Exp Med 216: 84–98. https: // doi.org / 10.1084 / jem.20181639
CAS
Статья
PubMed
PubMed Central
Google Scholar
Soty M, Gautier-Stein A, Rajas F, Mithieux G (2017) Передача сигналов глюкозы из кишечника в мозг в энергетическом гомеостазе. Cell Metab 25: 1231–1242. https://doi.org/10.1016/j.cmet.2017.04.032
CAS
Статья
PubMed
Google Scholar
Sun M, Wu W, Chen L, Yang W, Huang X, Ma C, Chen F, Xiao Y, Zhao Y, Ma C, Yao S, Carpio VH, Dann SM, Zhao Q, Liu Z, Cong Y (2018) Короткоцепочечные жирные кислоты, полученные из микробиоты, способствуют выработке IL-10 клетками Th2 для поддержания гомеостаза кишечника.Нац Коммуна 9: 3555. https://doi.org/10.1038/s41467-018-05901-2
CAS
Статья
PubMed
PubMed Central
Google Scholar
Takebe K, Nio J, Morimatsu M, Karaki S, Kuwahara A, Kato I, Iwanaga T (2005) Гистохимическая демонстрация Na (+) — связанного переносчика короткоцепочечных жирных кислот (slc5a8) в кишечнике и почка мыши. Biomed Res 26: 213–221
CAS
Статья
Google Scholar
Tao JH, Duan JA, Jiang S, Guo JM, Qian YY, Qian DW (2016) Одновременное определение шести короткоцепочечных жирных кислот в содержимом толстой кишки мышей с колитом после перорального введения газом полисахаридов из Chrysanthemum morifolium Ramat хроматография с пламенно-ионизационным детектором.J Chromatogr B Anal Technol Biomed Life Sci 1029-1030: 88–94. https://doi.org/10.1016/j.jchromb.2016.07.002
CAS
Статья
Google Scholar
Thaiss CA, Levy M, Korem T, Dohnalová L, Shapiro H, Jaitin DA, David E, Winter DR, Gury-BenAri M, Tatirovsky E, Tuganbaev T, Federici S, Zmora N, Zeevi D, Dori- Бахаш М., Певснер-Фишер М., Картвелишвили Э., Брэндис А., Хармелин А., Шиболет О., Халперн З, Хонда К., Амит I, Сегал Э, Элинав Э. (2016) Программы суточной ритмичности микробиоты содержат колебания транскриптома.Cell 167: 1495–1510.e12. https://doi.org/10.1016/j.cell.2016.11.003
CAS
Статья
PubMed
Google Scholar
Tian L, Bruggeman G, van den Berg M, Borewicz K, Scheurink AJ, Bruininx E, de Vos P, Smidt H, Schols HA, Gruppen H (2017) Влияние пектина на характеристики ферментации, использование углеводов и состав микробного сообщества желудочно-кишечного тракта свиней-отъемышей. Mol Nutr Food Res 61. DOI: https: // doi.org / 10.1002 / mnfr.201600186
Артикул
Google Scholar
Upadhyaya B, McCormack L, Fardin-Kia AR, Juenemann R, Nichenametla S, Clapper J, Specker B, Dey M (2016) Влияние диетического резистентного крахмала типа 4 на микробиоту кишечника человека и иммунометаболические функции. Sci Rep 6: 28797. https://doi.org/10.1038/srep28797
CAS
Статья
PubMed
PubMed Central
Google Scholar
Wan MLY, Forsythe SJ, El-Nezami H (2018) Взаимодействие пробиотиков с патогенами пищевого происхождения: потенциальная альтернатива антибиотикам и будущие проблемы.Crit Rev Food Sci Nutr 5: 1–14. https://doi.org/10.1080/10408398.2018.14
CAS
Статья
Google Scholar
Wang L, Hu L, Yan S, Jiang T, Fang S, Wang G, Zhao J, Zhang H, Chen W. (2017a) Влияние различных олигосахаридов в различных дозах на состав микробиоты кишечника и короткоцепочечных жирные кислоты у мышей при запорах. Food Funct 8: 1966–1978. https://doi.org/10.1039/c7fo00031f
CAS
Статья
PubMed
Google Scholar
Wang LL, Guo HH, Huang S, Feng CL, Han YX, Jiang JD (2017b) Всесторонняя оценка продукции SCFA кишечными бактериями, регулируемыми берберином, с использованием газовой хроматографии в сочетании с полимеразной цепной реакцией.J Chromatogr B Anal Technol Biomed Life Sci 1057: 70–80. https://doi.org/10.1016/j.jchromb.2017.05.004
CAS
Статья
Google Scholar
Wang C, Yang S, Gao L, Wang L, Cao L (2018) Карбоксиметил пахиман (CMP) снижает мукозит кишечника и регулирует микрофлору кишечника у мышей с опухолью CT26, обработанных 5-фторурацилом. Food Funct 9: 2695–2704. https://doi.org/10.1039/c7fo01886j
CAS
Статья
PubMed
Google Scholar
Wang K, Liao M, Zhou N, Bao L, Ma K, Zheng Z, Wang Y, Liu C, Wang W, Wang J, Liu SJ, Liu H (2019) Parabacteroides distasonis снижает ожирение и снижает метаболизм дисфункции из-за выработки сукцината и вторичных желчных кислот.Cell Rep 26: 222–235.e5. https://doi.org/10.1016/j.celrep.2018.12.028
CAS
Статья
PubMed
Google Scholar
Wei X, Tao J, Xiao S, Jiang S, Shang E, Zhu Z, Qian D, Duan J (2018) Xiexin tang улучшает симптомы диабетических крыс 2 типа путем модуляции микробиоты кишечника. Sci Rep 8: 3685. https://doi.org/10.1038/s41598-018-22094-2
CAS
Статья
PubMed
PubMed Central
Google Scholar
Xiao S, Zhang Z, Chen M, Zou J, Jiang S, Qian D, Duan J (2019a) Xiexin tang уменьшает дислипидемию у крыс с ожирением, вызванных диетой с высоким содержанием жиров, за счет повышения уровня короткоцепочечных жирных кислот кишечника, полученных из микробиоты производство и регулировка энергетического обмена.Дж. Этнофармакол 241: 112032. https://doi.org/10.1016/j.jep.2019.112032
Артикул
PubMed
Google Scholar
Xiao S, Liu C, Chen M, Zou J, Zhang Z, Cui X, Jiang S, Shang E, Qian D, Duan J (2019b) Радикс Scutellariae и корневище коптиды улучшают метаболизм гликолипидов у диабетических крыс 2 типа с помощью модулирование микробиоты кишечника и ее метаболитов. Appl Microbiol Biotechnol. https://doi.org/10.1007/s00253-019-10174-w
Артикул
Google Scholar
Yan H, Lu J, Wang Y, Gu W, Yang X, Yu J (2017) Потребление общих сапонинов и полисахаридов из Polygonatum kingianum влияет на микробиоту кишечника у диабетических крыс.Фитомедицина 26: 45–54. https://doi.org/10.1016/j.phymed.2017.01.007
CAS
Статья
PubMed
Google Scholar
Zeng H, Huang C, Lin S, Zheng M, Chen C, Zheng B, Zhang Y (2017) Крахмал, устойчивый к семенам лотоса, регулирует микробиоту кишечника и увеличивает производство короткоцепочечных жирных кислот и всасывание минералов у мышей. J. Agric Food Chem. 65: 9217–9225. https://doi.org/10.1021/acs.jafc.7b02860
CAS
Статья
PubMed
Google Scholar
Zhang X, Zhao Y, Zhang M, Pang X, Xu J, Kang C, Li M, Zhang C, Zhang Z, Zhang Y, Li X, Ning G, Zhao L (2012) Структурные изменения микробиоты кишечника во время опосредованной берберином профилактики ожирения и инсулинорезистентности у крыс, получавших диету с высоким содержанием жиров.PLoS One 7: e42529. https://doi.org/10.1371/journal.pone.0042529
CAS
Статья
PubMed
PubMed Central
Google Scholar
Чжао Ю., Чен Ф, Ву В., Сун М., Билотта А.Дж., Яо С., Сяо Ю., Хуанг Х, Ивз-Пайлес Т.Д., Головко Г., Фофанов Ю., Д’Соуза В., Чжао К., Лю З., Cong Y (2018a) GPR43 опосредует регуляцию SCFA метаболита микробиоты экспрессии антимикробного пептида в эпителиальных клетках кишечника посредством активации mTOR и STAT3.Mucosal Immunol 11: 752–762. https://doi.org/10.1038/mi.2017.118
CAS
Статья
PubMed
PubMed Central
Google Scholar
Zhao L, Zhang F, Ding X, Wu G, Lam YY, Wang X, Fu H, Xue X, Lu C, Ma J, Yu L, Xu C, Ren Z, Xu Y, Xu S, Shen Х, Чжу Х, Ши И, Шен Цюй, Донг В, Лю Р, Лин И, Цзэн И, Ван Х, Чжан Цю, Ван Дж, Ван Л, Ву И, Цзэн Б, Вэй Х, Чжан М, Пэн И, Zhang C (2018b) Кишечные бактерии, избирательно продвигаемые пищевыми волокнами, облегчают диабет 2 типа.Наука 359: 1151–1156. https://doi.org/10.1126/science.aao5774
CAS
Статья
PubMed
Google Scholar
Zhao C, Dong H, Zhang Y, Li Y (2019) Открытие потенциальных генов, вносящих вклад в биосинтез короткоцепочечных жирных кислот и лактата в кишечной микробиоте из систематического исследования E. coli . NPJ Biofilms Microbiomes 5:19. https://doi.org/10.1038/s41522-019-0092-7
CAS
Статья
PubMed
PubMed Central
Google Scholar
Zou J, Chassaing B, Singh V, Pellizzon M, Ricci M, Fythe MD, Kumar MV, Gewirtz AT (2018) Опосредованное клетчаткой питание микробиоты кишечника защищает от ожирения, вызванного диетой, восстанавливая опосредованный IL-22 здоровье толстой кишки.

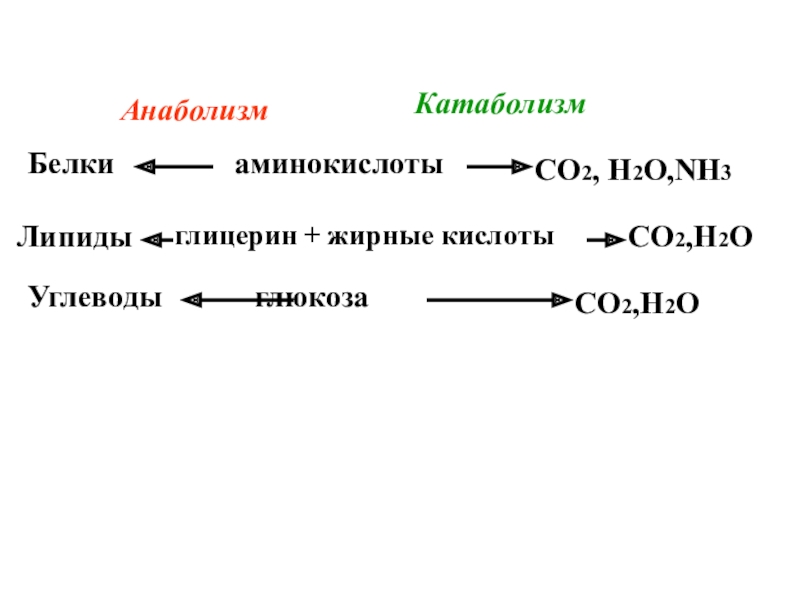 В клетках из них строятся белки тела. Белки входят в состав клеток, участвуют в процессах свертываемости крови, транспортировки газов, входят в состав костей. Они способны к окислению с выделением энергии, которая в дальнейшем будет использоваться организмом.
В клетках из них строятся белки тела. Белки входят в состав клеток, участвуют в процессах свертываемости крови, транспортировки газов, входят в состав костей. Они способны к окислению с выделением энергии, которая в дальнейшем будет использоваться организмом. В результате образуется новый материал для построения клеток и их роста, а так же запасается энергия.
В результате образуется новый материал для построения клеток и их роста, а так же запасается энергия. Например, после еды преобладают анаболические процессы, после сна — катаболические. Более того первая стадия анаболизма является последней стадией катаболизма.
Например, после еды преобладают анаболические процессы, после сна — катаболические. Более того первая стадия анаболизма является последней стадией катаболизма. katabole сбрасывание разрушение) (диссимиляция), совокупность протекающих в живом организме ферментативных реакций расщепления сложных органических веществ (в т. ч. пищевых). В процессе катаболизма происходит освобождение энергии,… … Большой Энциклопедический словарь
katabole сбрасывание разрушение) (диссимиляция), совокупность протекающих в живом организме ферментативных реакций расщепления сложных органических веществ (в т. ч. пищевых). В процессе катаболизма происходит освобождение энергии,… … Большой Энциклопедический словарь А., Лисовенко Л.А., Москва: Изд во ВНИРО, 1995 г.) … Молекулярная биология и генетика. Толковый словарь.
А., Лисовенко Л.А., Москва: Изд во ВНИРО, 1995 г.) … Молекулярная биология и генетика. Толковый словарь. dunwoodypress.com/148/PDF/Biotech Eng Rus.pdf] Тематики биотехнологии EN catabolism … Справочник технического переводчика
dunwoodypress.com/148/PDF/Biotech Eng Rus.pdf] Тематики биотехнологии EN catabolism … Справочник технического переводчика Простыми словами – это генерация клеток и синтез белков и гормонов, благодаря которым происходит рост мышечных волокон, чего добиваются все спортсмены.
Простыми словами – это генерация клеток и синтез белков и гормонов, благодаря которым происходит рост мышечных волокон, чего добиваются все спортсмены. Конституция тела и индивидуальный метаболизм.
Конституция тела и индивидуальный метаболизм.


 Избегать стресса и потрясений, расслабляться.
Избегать стресса и потрясений, расслабляться.



 Поэтому у аэробных организмов после гликолиза (или спиртового брожения) следует третий, завершающий этап энергетического обмена — полное кислородное расщепление, или клеточное дыхание.
Поэтому у аэробных организмов после гликолиза (или спиртового брожения) следует третий, завершающий этап энергетического обмена — полное кислородное расщепление, или клеточное дыхание.

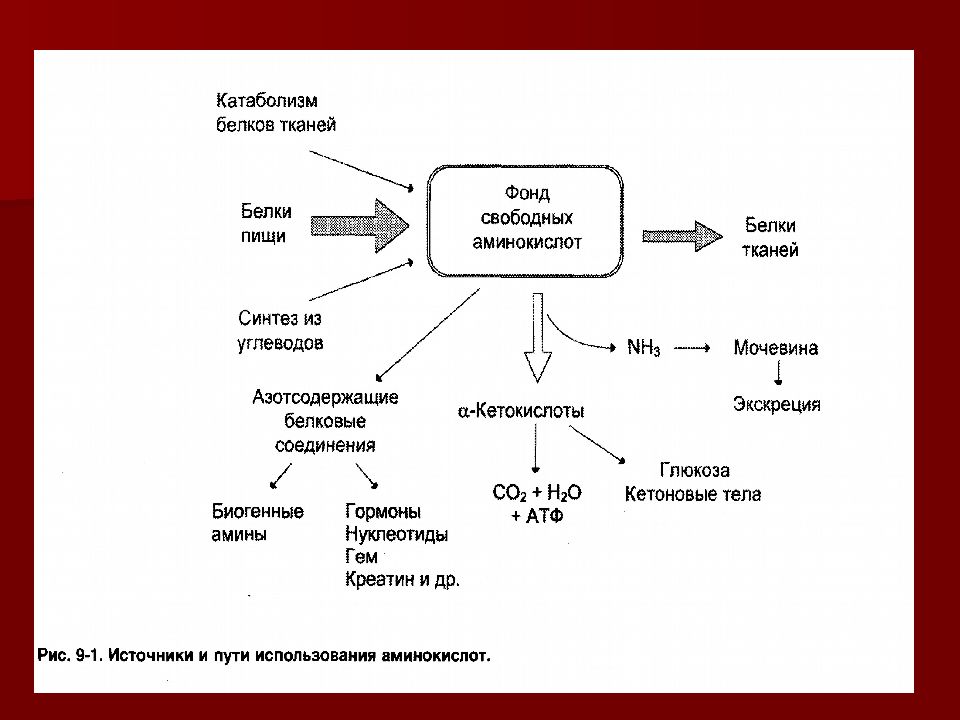

 Л. Билич, В.А. Крыжановский. — 2008
Л. Билич, В.А. Крыжановский. — 2008
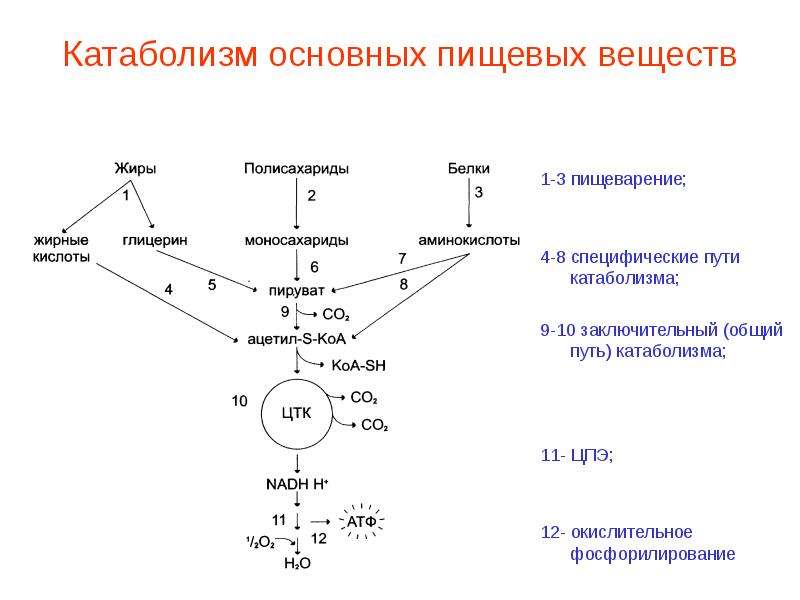
 Кхемогетеротрофамотносятся большинство бактерий, а так
же грибы и животные.
Кхемогетеротрофамотносятся большинство бактерий, а так
же грибы и животные.
 Дальнейшее
превращение С3-углеводов
осуществляется за счет переноса водорода
и фосфорных остатков через ряд органических
кислот с участием специфических
дегидрогеназ. Все реакции этого пути,
за исключением трех, протекающих с
участием гексокиназы, 6-фосфофруктокиназы
и пируваткиназы, полностью обратимы.
На стадии образования пировиноградной
кислоты заканчивается анаэробная фаза
превращения углеводов.
Дальнейшее
превращение С3-углеводов
осуществляется за счет переноса водорода
и фосфорных остатков через ряд органических
кислот с участием специфических
дегидрогеназ. Все реакции этого пути,
за исключением трех, протекающих с
участием гексокиназы, 6-фосфофруктокиназы
и пируваткиназы, полностью обратимы.
На стадии образования пировиноградной
кислоты заканчивается анаэробная фаза
превращения углеводов.

 Образовавшаяся
одним из путей ПВК в клетке подвергается
дальнейшему окислению. Освобождающиеся
углерод и водород удаляются из клетки.
Углерод выделяется в форме CO2,
водород передается на различные
акцепторы. Причем может передаваться
либо ион водорода, либо электрон, поэтому
перенос водорода равноценен переносу
электрона. В зависимости от конечного
акцептора водорода (электрона) различают
аэробное дыхание, анаэробное дыхание
и брожение.
Образовавшаяся
одним из путей ПВК в клетке подвергается
дальнейшему окислению. Освобождающиеся
углерод и водород удаляются из клетки.
Углерод выделяется в форме CO2,
водород передается на различные
акцепторы. Причем может передаваться
либо ион водорода, либо электрон, поэтому
перенос водорода равноценен переносу
электрона. В зависимости от конечного
акцептора водорода (электрона) различают
аэробное дыхание, анаэробное дыхание
и брожение.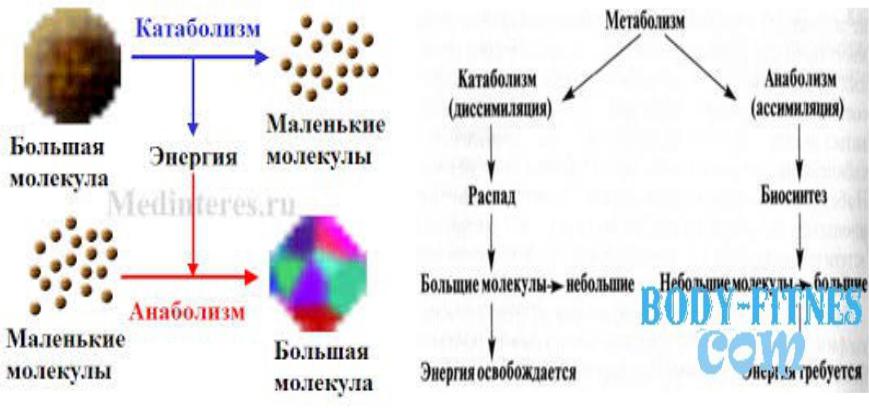 Этот процесс называется анаэробным
дыханием.
Этот процесс называется анаэробным
дыханием. Ацетальдегид, соединяясь
с коферментом одного из окислительных
ферментов – коферментом А (КоА-SH),
образует «активированную уксусную
кислоту» — ацетил-КоА – высокоэнергетическое
соединение.
Ацетальдегид, соединяясь
с коферментом одного из окислительных
ферментов – коферментом А (КоА-SH),
образует «активированную уксусную
кислоту» — ацетил-КоА – высокоэнергетическое
соединение. Оксалоацетат взаимодействует с
ацетил-КоА, и цикл повторяется снова.
Каждая из 10 реакций ЦТК, за исключением
одной, легко обратима. В цикл вступают
два атома углерода в виде ацетил-КоА и
такое же количество атомов углерода
покидают этот цикл в виде СО2.
Оксалоацетат взаимодействует с
ацетил-КоА, и цикл повторяется снова.
Каждая из 10 реакций ЦТК, за исключением
одной, легко обратима. В цикл вступают
два атома углерода в виде ацетил-КоА и
такое же количество атомов углерода
покидают этот цикл в виде СО2.
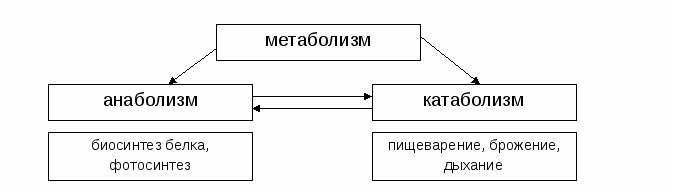
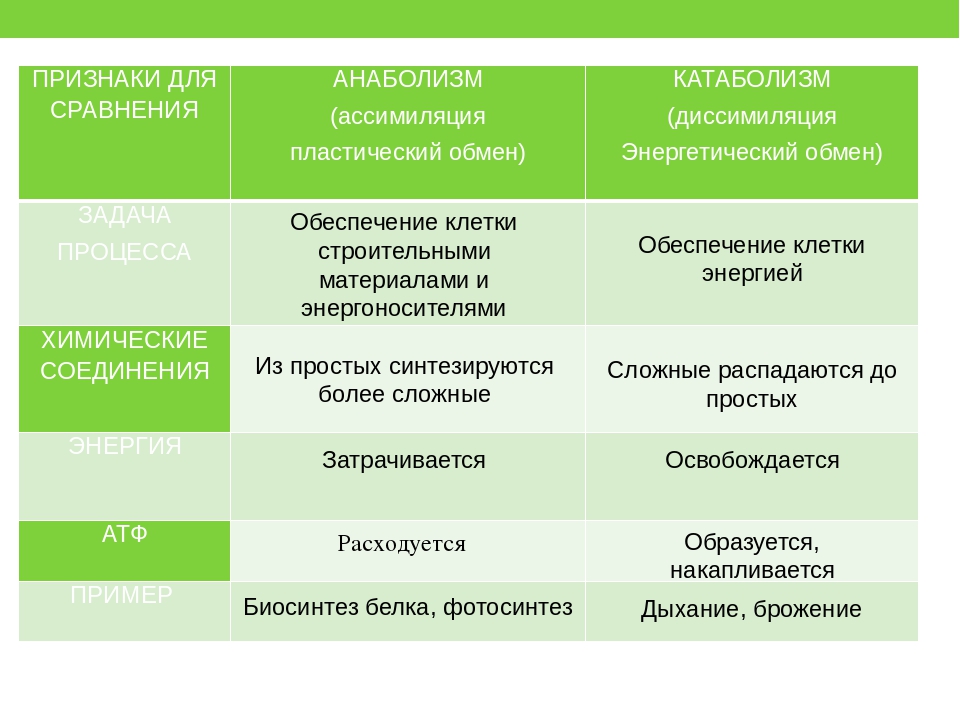 Конечными
продуктами неполного окисления являются
органические кислоты: уксусная, лимонная,
фумаровая, глюконовая и др., которые
аккумулируются в среде. Этот окислительный
процесс используется микроорганизмами
для получения энергии. Однако общий
выход энергии при этом значительно
меньший, чем при полном окислении. Часть
энергии окисляемого исходного субстрата
сохраняется в образующихся органических
кислотах.
Конечными
продуктами неполного окисления являются
органические кислоты: уксусная, лимонная,
фумаровая, глюконовая и др., которые
аккумулируются в среде. Этот окислительный
процесс используется микроорганизмами
для получения энергии. Однако общий
выход энергии при этом значительно
меньший, чем при полном окислении. Часть
энергии окисляемого исходного субстрата
сохраняется в образующихся органических
кислотах.
 Проходит в два этапа: световая фаза (происходит улавливание и фиксация энергии света в АТФ) и темновая (связывание углекислого газа в молекулы глюкзы с затратой энергии АТФ).
Проходит в два этапа: световая фаза (происходит улавливание и фиксация энергии света в АТФ) и темновая (связывание углекислого газа в молекулы глюкзы с затратой энергии АТФ). Конечными продуктами выступают СО2 и Н2О.
Конечными продуктами выступают СО2 и Н2О.