
Анатомия верхней конечности
Области верхней конечности , Анатомия человека : Рисунки
Мышцы; мышечная система , Плечо , Анатомия : Иллюстрации
Лопатка : Мышцы Прикрепление , Атлас анатомии
Плечевой сустав , Плечевой сустав , Анатомия человека : Схема
Поверхностные вены верхней конечности , Анатомия : Иллюстрации: А.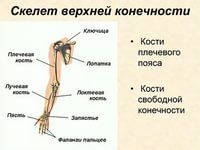
Кисть : Ладонь; ладонная область , Артерии (Иллюстрации: А. Мишо)
Кости кисти , Анатомия : Медицинские иллюстрации
|
|
|
содержание .. 10 11 12 13 14 15 16 17 18 19 20 ..
Скелет конечностей (анатомия человека)
В
строении костей скелета верхней и нижней конечностей много общего. Как в
верхней, так и в нижней конечностях различают скелет пояса и скелет
свободной конечности. Оба отдела состоят из сходных по развитию и
строению костей. В скелете свободной конечности, как верхней, так и
нижней, выделяют три отдела: проксимальный, средний и дистальный. Верхние конечности являются органами хватания, приспособленными для выполнения разнообразных и тонких движений, необходимых во время работы. Рука у человека является органом труда. Нижние конечности человека предназначены для опоры и передвижения. Различие функций конечностей обусловливает определенные отличия в их строении. Кости нижней конечности большего размера, массивнее и имеют большую площадь опоры. Пояс нижней конечности малоподвижный, кости его сращены между собой и соединены с крестцовой частью позвоночника. Пояс верхней конечности подвижный. Кости, его образующие, соединены между собой и с туловищем подвижно посредством суставов и связок. Кости верхней конечности отличаются меньшими размерами.
Развитие костей конечностей (анатомия человека)
Зачатки скелета верхних и нижних конечностей возникают на 6-й неделе
развития в виде выростов мезенхимы из боковых отделов туловища зародыша.
Зачатки скелета верхних конечностей расположены в верхнем отделе
туловища, зачатки костей нижних конечностей — на уровне, поясничных
позвонков. Расширенные концы выростов представляют собой зачатки кисти и
стопы. По мере роста выросты удлиняются и в них последовательно
появляются закладки вначале костей предплечья, затем — плеча. На нижней
конечности вслед за закладкой стопы появляется зачаток голени, позднее —
зачаток бедра. К концу 6-й недели развития формируются зачатки основных
костей конечностей, к 8-й неделе — все части скелета конечностей,
включая мелкие кости кисти и стопы. Первичный скелет верхних и нижних
конечностей зародыша вначале обращен сгибательной поверхностью к
туловищу. В дальнейшем происходит поворот на 90° костей верхней
конечности кнаружи, а нижней — кнутри. Все кости конечностей, за исключением ключицы, проходят три стадии развития: перепончатую, хрящевую и костную. Ключица относится к первичным костям и проходит соединительнотканную, перепончатую и костную стадии развития. Окостенение костей конечностей начинается с ключицы, которая является одной из наиболее рано обызвествляющихся частей скелета. Первые островки окостенения появляются в ней на 6-й неделе эмбрионального развития. В остальных костях конечностей центры окостенения возникают, начиная с 8-й недели внутриутробного развития, и появляются в определенных местах и в определенное время, характерное для каждой кости. Кости, образующие скелет плеча, предплечья, пясти, бедра, голени, плюсны и фаланги пальцев, относятся к трубчатым костям и имеют общие черты развития. Их диафизы развиваются эндохондральным и периостальным путем, эпифизы — эндохондральным путем. Кости поясов развиваются как эндохондральным, так и перихондральным способом, а кости запястья и предплюсны, состоящие преимущественно из губчатого вещества, развиваются эндохондрально. Первый островок окостенения появляется в их хрящевом теле. Одновременно начинается образование кости на периферии за счет надкостницы. К концу внутриутробного развития или в первые месяцы после рождения появляются одна или несколько точек окостенения в эпифизах. За счет прослойки хряща, отделяющего диафиз от эпифиза, трубчатая кость растет в длину, а за счет надкостницы — в толщину. Приблизительно к 25 годам хрящевая прослойка замещается костной тканью, после чего рост кости в длину прекращается.
Наряду с развитием кости в толще диафиза идет резорбция костной ткани,
приводящая к образованию костномозгового канала.
содержание .. 10 11 12 13 14 15 16 17 18 19 20 ..
|
|
|
 Наличие канала
уменьшает вес трубчатых костей и обеспечивает их максимальную прочность.
Наличие канала
уменьшает вес трубчатых костей и обеспечивает их максимальную прочность.Урок 12. Скелет поясов и свободных конечностей.
Тема: Скелет поясов и свободных конечностей. Соединение костей конечностей
Цель: продолжить формирование знаний об особенностях скелета человека, рассмотреть скелет поясов и свободных конечностей, функции, соединение костей, обеспечивающих движение; развивать умение определять кости скелета конечностей.
Ход урока
Организационный момент
Проверка домашнего задания (фронтальный опрос по вопросам)
-
Изучение нового материала
Слайд 2 – В скелете различают пояса конечностей и кости конечностей. Они относятся к добавочному скелету человека.
Слайд 3 – Скелет пояса верхних конечностей состоит из: ключиц (2), лопаточных костей (2), плечевых костей (2), локтевой и лучевой костей (по 2), 16 костей запястья, двадцати фаланг пальцев.
Слайд 4 — Скелет верхних конечностей состоит из трех отделов: плеча, предплечья и кисти. Плечо имеет лишь одну плечевую кость. Ее верхняя часть — шарообразная головка помещается в полушаровидной ямке лопатки. Предплечье образовано двумя костями: локтевой и лучевой.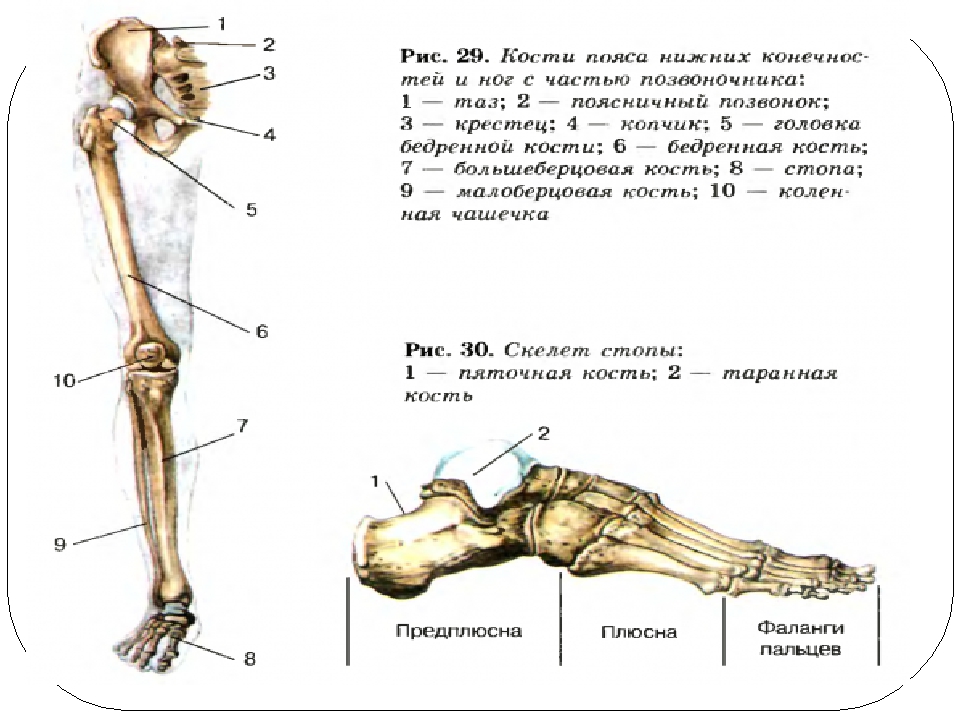
Слайд 5 — В кисти различают 3 отдела: запястье, пястье и пальцы. Скелет запястья состоит из восьми мелких костей. Пять длинных костей пясти составляют скелет ладони и дают опору костям пальцев. Такое строение кисти обеспечивает выполнение разнообразных тончайших движений.
Слайды 6-8 — Пояс верхних конечностей состоит из ключиц и лопаток.
Лопатки — большие плоские кости треугольной формы. Они находятся на задней поверхности грудной клетки и соединены с ребрами и позвоночным столбом только при помощи мышц.
Ключица — слегка изогнутая кость средних размеров. Одним концом она соединена с лопаткой, а другим — с грудиной.
Лопатка лежит на ребрах. При движении руки она меняет свое положение вместе с ключицей.
Попробуйте сделать это движение и убедитесь сами!
Слайд 9 – Благодаря тому, что конечности прикреплены к надежной опоре, они обладают подвижностью во всех направлениях, способны выдерживать большие физические нагрузки.
Слайды 10-11 – Повороты кисти ладонью вверх и вниз возможны потому, что лучевая кость может перемешаться вокруг локтевой. Попробуйте вращать кисть. Дугу описывает большой палец. Попробуйте проделать такой же поворот так, чтобы дугу описывал не большой палец, а мизинец. Это невозможно, так как локтевая кость не может вращаться вокруг лучевой кости.
Слайд 12 – Скелет пояса нижних конечностей состоит из:
1. Тазовых костей
2. Бедренных костей (двух больших бедренных костей и двух малых бедренных костей)
5. Костей стопы
6. Двадцати фаланг пальцев
Он отличается большой прочностью, которая достигается за счет некоторого ограничения подвижности.
Слайды 13-14 – Тазовая кость – плоская, сочленена с крестцом. Кости образуют неподвижное практически неподвижное сочленение. Тазовые кости и мышцы образуют дно брюшной полости, на которое опираются все внутренние органы.
Тазовые кости и мышцы образуют дно брюшной полости, на которое опираются все внутренние органы.
С тазовыми костями сочленяются бедренные кости, образуя прочную арку.
Слайд 15 – Скелет нижних конечностей (30 костей) (пояснения учителя по схеме).
пояс нижних конечностей (таз) = тазовые кости (подвздошная (2) + седалищная (2) + лонная (2))
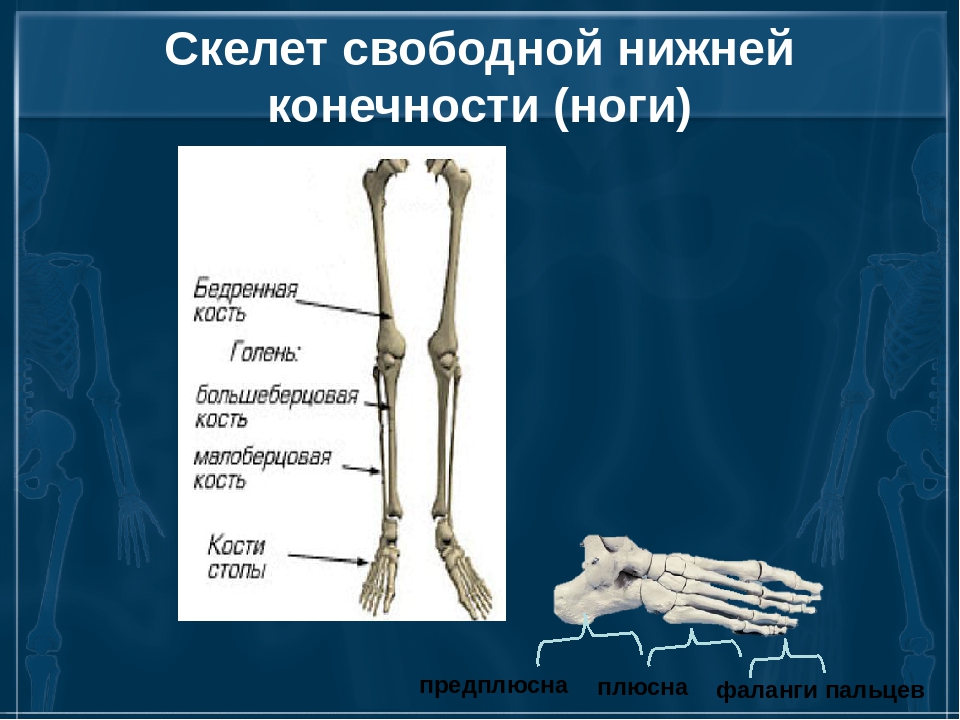
нижняя конечность = бедро (бедренная кость – 1) + надколенник (1) + голень (большая + малая берцовые кости) + кости плюсны (7) + кости предплюсны (5) + кости фаланг пальцев (14)
Слайд 16 – Сочленение таза и бедренной кости
Губчатое вещество кости состоит из пластинок, расположенных по направлениям сил сжатия и растяжения. Этим объясняется её прочность.
Слайд 17 – Большая берцовая кость самая прочная. Она сочленяется с бедром и стопой. Такое строение увеличивает прочность скелета ноги. Малая берцовая кость играет вспомогательную роль.
Слайды 18-19 – Стопу можно сравнить с рессорой, так как предплюсна и плюсна образует своды. Нагрузка приходится на пяточную кость, головки плюсневых костей. Своды поддерживаются и мышцами. Если стопа нормальна, то нагрузка распределяется так, как показано на рисунке.
Слайд 20 — Любое движение связано с перемещением костей, которое осуществляется мышцами.
Слайды 21—23 – Типы соединения костей
Неподвижные и полуподвижные соединения костей.
Между костями черепа образуются швы.
Ребра с грудиной соединяются посредством хряща.
При наклоне, например, вперёд, хрящ между позвонками сжимается спереди и растягивается сзади. За счет этого происходит движение, правда ограниченное.
За счет уплощения межпозвонковых хрящей рост к вечеру уменьшается на 1,5 см. , а к 80 годам – на 5-7 см.
, а к 80 годам – на 5-7 см.
Слайды 24-28 – Подвижные соединения – суставы (пояснения учителя по схеме).
Кости конечностей соединены подвижно: головки одних костей входят во впадины других и скрепляются связками и суставными сумками. Подвижные соединения костей называются суставами.
Найдите на себе следующие суставы!
Подвижность сустава обеспечивается формой суставных поверхностей сочленяющихся костей, суставным хрящом и суставной жидкостью, а прочность – суставной сумкой, связками и более низким давлением внутри сустава по сравнению с давлением наружного воздуха.
Кости могут быть закреплены в суставе, если мышцы прижмут одну кость к другой: суставная жидкость при этом вытиснится в щелевые складки суставной сумки, хрящ суставной головки вдавится в хрящ суставной впадины.
Закрепление изученного материала
Слайды 29-30 – С помощью таблицы давайте уточним особенности строения скелета верхних и нижних конечностей.
Отделы телаОтделы скелета
Кости скелета
Тип костей
Характер соединения костей
Особенности скелета человека
Верхняя конеч
ность
Плечевой пояс
Две лопатки
Две ключицы
Плоские (широкие) Длинные трубчатые
Подвижное
Плечевой пояс подвижный, обеспечивает большую подвижность руки
Свободная конечность:
Плечо
Предплечье
Кисть
Плечевая кость
Локтевая и лучевая —
Кости запястья
пястья
фаланги пальцев
Длинные
трубчатые
короткие
губчатые
короткие
трубчатые
Подвижное
Облегченная конструкция скелета, высокая подвижность в плечевом суставе. Большая подвижность большого пальца и способность его противопоставляться всем остальным
Большая подвижность большого пальца и способность его противопоставляться всем остальным
Нижняя
конеч
ность
Тазовый пояс
Парные кости –
подвздошные,
седалищные,
лобковые
Плоские
Неподвиж ное
Расширенная форма таза, широкораставленные
тазобедренные суставы.
Свободная
Конечность:
Бедро
Голень
Стопа
Бедренная
Большеберцовая малоберцовые
Предплюсны (таранная и пяточная) плюсны фаланги пальцев
Длинные
трубчатые
короткие
губчатые
короткие
трубчатые
Подвижное
Мощное развитие костей ног, выпрямленность в коленном суставе, мощное развитие связок Опорная стопа с мощным большим пальцем. Пружинящий свод стопы.
Устный индивидуальный опрос
Слайд 31 – Ответьте на вопросы:
Какие кости образуют верхнюю конечность?
Чем образован плечевой пояс конечностей?
Какие кости образуют нижнюю конечность?
В чем сходство и различие в строении предплечья и голени?
Слайды 32-33 – Назовите части и функции (работа по рисункам)
Слайд 34 – Дайте обоснованный ответ.
Какое преимущество обеспечивает форма стопы и строение таза для человека в отличие от млекопитающих?
Каковы особенности строения скелета человека, связанные с трудовой деятельностью и прямохождением?
Слайд 35 – Укажите соответствие:
1. Грудная клетка А. Имеет изгибы
Грудная клетка А. Имеет изгибы
2. Стопа Б. Расширена в стороны
3. Позвоночник В. Сводчатая
4. Верхняя конечность Г. Широкий, как чаша
5. Таз Д. Большой палец противопоставляется остальным
Е. Опора для внутренних органов
Ж. Защита внутренних органов
З. Удерживание орудий труда
И. Пружинит, смягчает толчки при движении
Ответы: 1 — Б, Ж 2 – В, И 3 — А, Е 4 — Д, З 5 — Г, Е
Подведение итога урока. Рефлексия
Слайд 36 — Домашнее задание: § 12 прочитать. Провести функциональные пробы, описанные в конце параграфа. Повторить из параграфа 8 материал о мышечных тканях.
К вопросу гомологии в структурной организации костей проксимальных сегментов конечностей человека Текст научной статьи по специальности «Клиническая медицина»
УДК 611.717.4/611.718.4
DOI: 10.21626/vestnik/2019-1/10
К ВОПРОСУ ГОМОЛОГИИ В структурной организации костей ПРОКСИМАЛЬНЫХ СЕГМЕНТОВ КОНЕЧНОСТЕЙ ЧЕЛОВЕКА
© Яшина И.Н.1, Иванов А.В.1, Клочкова С.В.2
1 Курский государственный медицинский университет (КГМУ)
Россия, 305041, Курская область, г. Курск, ул. К. Маркса, д. 3;
2
Первый Московский государственный медицинский университет имени И.М. Сеченова
(Сеченовский Университет)
Россия, 125009, г. Москва, ул. Моховая, д. 11, стр. 10
Москва, ул. Моховая, д. 11, стр. 10
Цель исследования — выявление закономерностей и различий в структурной организации плечевых и бедренных костей, связанных с функциями конечностей с учетом их принадлежности к стороне тела.
Материалы и методы. Проведена остеометрия 35 анатомических структур на 308 костях проксимальных сегментов свободных конечностей людей. С целью учета влияния соматотипа человека на размеры костных структур данные были нормализованы. За единицу измерения был принят поперечный диаметр диафиза на середине длины кости. Полученные результаты были обработаны методами факторного анализа Maximum Maximum Likelihood Factor с вращением Equamax normalized, при этом были сгенерированы четыре выборки, учитывающие принадлежность к поясу конечности и стороне тела.
Результаты. Особенностями структурной организации плечевой кости явилось выделение в качестве базового структурного параметра первого уровня ширины проксимального эпифиза, для бедренной кости сагиттального диаметра головки и межвертельного расстояния. Асимметричность структурной организации костей и вертикальная дифференциация эпифизов контрлатеральных костей определены наличием нестабильных параметров. Асимметрия структурной организации бедренной кости более выражена. Нестабильных параметров проксимального эпифиза правой бедренной кости выделено 4, дистального эпифиза — 1; в структурной организации проксимального эпифиза левой бедренной кости выделен 1, дистального эпифиза — 3. Нестабильных параметров проксимального эпифиза правой плечевой кости выделено 3, дистального эпифиза — 2; в структурной организации эпифизов левой плечевой кости выделено по 6 нестабильных параметров.
Заключение. Основу структурной организации костей проксимальных сегментов конечностей человека независимо от принадлежности к поясам конечностей и стороне тела образуют анатомические образования, отвечающие за трансляцию силовых нагрузок по оси конечности. Функциональная дифференциация конечностей человека отражается в структурной организации костей асимметричной вертикальной дифференциации эпифизов плечевых и бедренных костей.
Функциональная дифференциация конечностей человека отражается в структурной организации костей асимметричной вертикальной дифференциации эпифизов плечевых и бедренных костей.
Ключевые слова: плечевая кость; бедренная кость; структурная организация; факторный анализ.
Яшина Ирина Николаевна — канд. мед. наук, доцент кафедры анатомии человека, КГМУ, г. Курск. ORCID iD: 0000-00016235-2309. E-mail: [email protected] (автор, ответственный за переписку)
Иванов Александр Викторович — д-р мед. наук, профессор, заведующий кафедрой гистологии, цитологии и эмбриологии, КГМУ, г. Курск. ORCID iD: 0000-0002-2412-0475. E-mail: [email protected]
Клочкова Светлана Валерьевна — д-р мед. наук, профессор кафедры анатомии человека, Сеченовский Университет, г. Москва. ORCID iD: 0000-0003-2041-7607. E-mail: [email protected]
Увеличение продолжительности жизни наряду с изменениями спектра двигательной активности современного человека, гиподинамия, ожирение, травмы, инфекционные заболевания и многие другие факторы, воздействующие на опорно-двигательный аппарат, приводят к возрастающей частоте развития дегенеративно-дистрофических заболеваний крупных суставов человека, которые существенно снижают качество жизни человека. Большинство дегенеративно-дистрофических заболеваний опорно-двигательного аппарата носят системный характер и поражают одновременно несколько суставов. При этом исследования патологии опорно-двигательного аппарата связаны с конкретными заболеваниями: поражениями тазобедренного, коленного, плечевого или локте-
вого суставов. При изучении системной организации проксимального эпифиза бедренной кости при коксартрозе [10] мы столкнулись с недостатком информации о структурной организации костей здорового человека, которая необходима для понимания причин и механизмов асимметричной дезорганизации бедренных костей при развитии дегенеративно-дистрофических заболеваний. Развитие современных технологий обработки информации открывает новые возможности для анализа данных, полученных классическими анатомическими методами исследования.
Развитие современных технологий обработки информации открывает новые возможности для анализа данных, полученных классическими анатомическими методами исследования.
Цель настоящего исследования — выявление закономерностей и различий в структурной организации плечевых и бедренных костей, свя-
занных с функциями конечностей с учетом их принадлежности к стороне тела.
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Исследование выполнено на 308 мацериро-ванных плечевых и бедренных костях людей (77 правых плечевых, 77 левых плечевых, 77 бедренных правых и 77 бедренных левых), умерших на рубеже 20-21 веков, не несущих признаков патологии костной ткани, с полным синостозированием эпифизов.
По классической методике остеометрии производились измерения гомологичных структур плечевых и бедренных костей (в таблицах 1-3 и на рисунке 1 представлено описание структур и схемы измерения углов, характеризующих стереометрию костей) с учетом принадлежности к проксимальному эпифизу, диа-физу и дистальному эпифизу.
Для измерения использовались устройство для измерений длинных трубчатых костей [2], цифровой штангенциркуль и транспортир. С целью учета влияния соматотипа человека на размеры костных структур данные, полученные
в ходе остеометрии, были нормализованы. За единицу измерения был принят поперечный диаметр диафиза на середине длины кости.
Для решения поставленных задач на следующем этапе исследования применили метод факторного анализа — Maximum Likelihood Factor с вращением Equamax normalized, при этом были сгенерированы четыре выборки, учитывающие принадлежность к поясу конечности и ла-терализацию кости. Все требования к выборкам для проведения факторного анализа были выполнены. В качестве корреляционной матрицы использовалась матрица корреляции Спирмена. Поскольку методологически алгоритмическая схема реализации метода максимального правдоподобия включает последовательное выделение латентных факторов с объяснением наибольшей доли дисперсии исходных переменных, процесс выделения факторов был прерван на уровне отсутствия изменений в картине факторных нагрузок и их дисперсий при увеличении числа факторов. Статистически значимыми, влияющими на структуру при p>0,05, явились факторные нагрузки со значениями в диапазоне |0,7-1,0|.
Все требования к выборкам для проведения факторного анализа были выполнены. В качестве корреляционной матрицы использовалась матрица корреляции Спирмена. Поскольку методологически алгоритмическая схема реализации метода максимального правдоподобия включает последовательное выделение латентных факторов с объяснением наибольшей доли дисперсии исходных переменных, процесс выделения факторов был прерван на уровне отсутствия изменений в картине факторных нагрузок и их дисперсий при увеличении числа факторов. Статистически значимыми, влияющими на структуру при p>0,05, явились факторные нагрузки со значениями в диапазоне |0,7-1,0|.
Рис. 1. Схема измерения углов, характеризующих стереометрию костей.
Примечание: I — угол наклона диафиза, II — диафизарно-шеечный угол, III — угол скрученности плечевой кости / угол анте/ретроверсии шейки бедра, IV — угол хрящевого края головки плечевой кости.
Fig. 1. The scheme of measurement of angles characterizing the stereometry of bones.
Note: I — angle of the shaft axis, II- shaft-neck angle, III — angle of torsion of humerus / angle of ante-retroversion of the femoral neck, IV — angle of the cartilaginous edge of the humerus head.
Таблица 1
Table 1
Обозначение структур проксимального эпифиза плечевых и бедренных костей человека The osteometric parameters of the proximal epiphysis structures of human humerus and femur
Номер Number Обозначение The osteometric parameters
1 Ширина проксимального эпифиза для плечевой кости — наибольшее расстояние между головкой и большим бугром, для бедренной — наибольшее расстояние между головкой и большим вертелом. The width of the proximal epiphysis for the humerus is the greatest distance between the head and the major tubercle, for the femur is the greatest distance between the head and the major trochanter.
The width of the proximal epiphysis for the humerus is the greatest distance between the head and the major tubercle, for the femur is the greatest distance between the head and the major trochanter.
2 Сагиттальный диаметр головки или передне-задний размер головки, измеренный в горизонтальной плоскости. The sagittal diameter of the head or anterior-posterior head size, measured in the horizontal plane.
3 Вертикальный диаметр головки или верхне-нижний размер головки, измеренный во фронтальной плоскости. The vertical diameter of the head or top-bottom size of the head, measured in the frontal plane.
4 Сагиттальный диаметр шейки или передне-задний размер шейки, измеренный в горизонтальной плоскости. The sagittal diameter of the neck or anterior-posterior neck size, measured in the horizontal plane.
5 Вертикальный диаметр шейки. Расстояние, измеренное между максимально удаленными верхней и нижней точками шейки кости. The vertical diameter of the neck. The maximum distance between the upper and lower points of the bone neck.
6 Задняя длина шейки — расстояние между краем головки бедренной кости и серединой межвертельного гребня.
The back neck length is the distance between the edge of the femoral head and the middle of the intertrochanteric crest.
7 Верхняя длина шейки бедренной кости — расстояние между основанием большого вертела и краем головки бедренной кости сверху.
The upper length of the femoral neck is the distance between the base of the major trochanter and the head from above.
8 Межбугорковое расстояние — расстояние между вершинами большого и малого бугорков плечевой кости.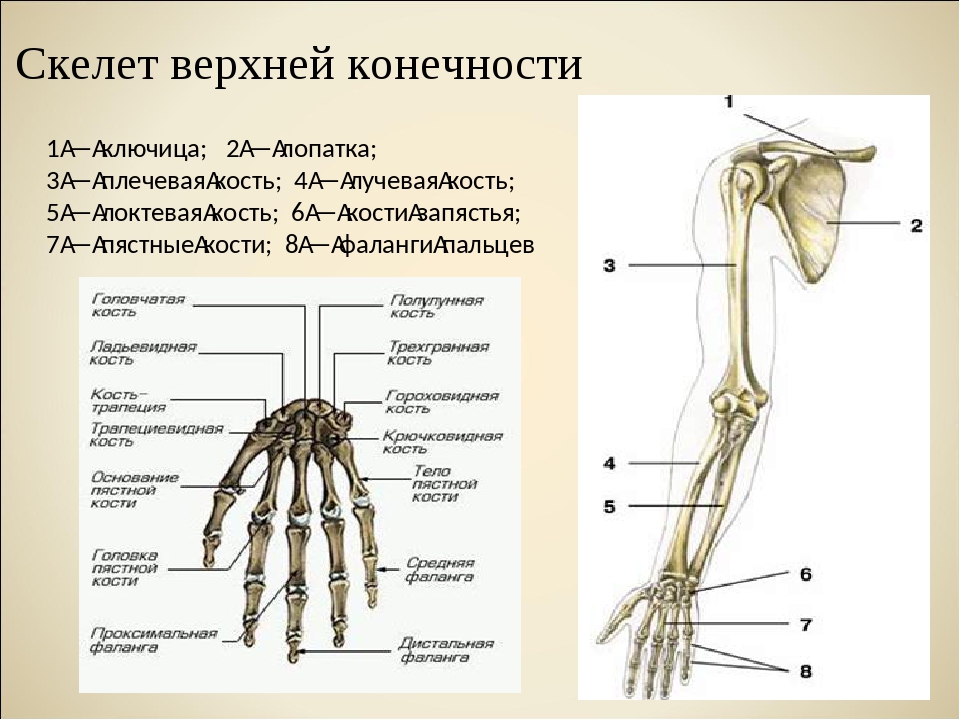 Межвертельное расстояние — расстояние между вершинами большого и малого вертела бедренной кости. The intertubercular distance is the distance between the tops of the major and minor tubercles of the humerus. The intertrochanteric distance is the distance between the peaks of major and minor trochanters.
Межвертельное расстояние — расстояние между вершинами большого и малого вертела бедренной кости. The intertubercular distance is the distance between the tops of the major and minor tubercles of the humerus. The intertrochanteric distance is the distance between the peaks of major and minor trochanters.
9 Ширина межбугорковой борозды. Внутреннее расстояние между гребнями бугорков.
The width of the intertubercular groove. The internal distance between the crests of the tubercles.
10 Глубина межбугорковой борозды — расстояние между дном и касательной линией, проведенной между гребнями бугорков.
The depth of the intertubercular groove is the distance between the bottom and the tangent line drawn between the crests of the tubercles.
11 Диафизарно-шеечный угол. The shaft-neck angle.
12 Угол скрученности кости или анте- (ретро-) версии шейки кости, угол отклонения оси шейки кпереди или кзади от фронтальной плоскости, проведенной по задней поверхности мыщелков.
The angle of torsion of the bone or ante- (retro-) version of the bone neck, the angle of deviation of the neck axis anteriorly or posteriorly from the frontal plane drawn on the back surface of the condyles.
13 Угол хрящевого края головки с диафизом (измеренный по методу В.Г. Властовского).
The angle of the cartilaginous edge of the neck with the shaft (measured according to V.G.Vlastovsky’s method).
Примечание: Серым цветом обозначены ячейки структур, характерных только для одной из костей.
Note: The gray color indicates the structures that are specific to only one of the bones.
Таблица 2
Table 2
Обозначение структур диафиза плечевых и бедренных костей человека The osteometric parameters of shaft structures of human humerus and femur
Номер Number Обозначение The osteometric parameters
14 Наибольшая длина кости, измеренная между максимально удаленными точками на нижней поверхности медиального мыщелка и наивысшей точкой на головке. The maximum bone length measured between the most distant points on the lower surface of the medial condyle and the highest point on the head.
15 Наибольшая длина кости, измеренная между максимально удаленными точками на нижней поверхности медиального мыщелка и наивысшей точкой на большом бугре или большом вертеле. The maximum bone length, measured between the most distant points on the lower surface of the medial condyle and the highest point on a major tubercle or major trochanter.
16 Поперечный диаметр диафиза — расстояние между боковыми точками на середине длины кости во фронтальной плоскости. The transverse diameter of the shaft is the distance between the side points in the middle of the bone length in the frontal plane.
17 Сагиттальный диаметр диафиза — передне-заднее расстояние, измеренное на середине длины кости. The sagittal diameter of the shaft is the anterior-posterior distance measured in the middle of the bone length.
18 Угол наклона оси диафиза по отношению к перпендикуляру, восстановленному от горизонтальной плоскости мыщелков.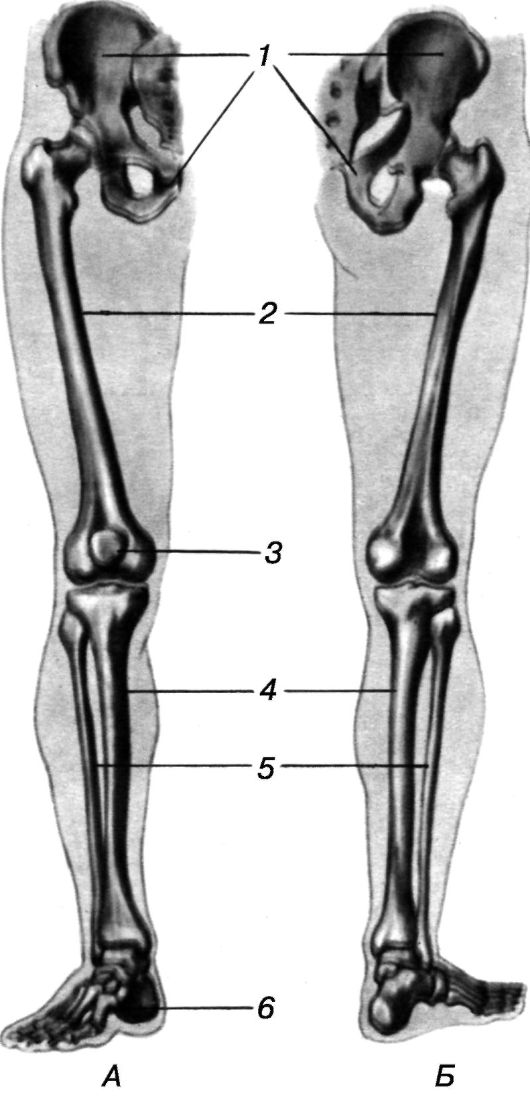 The angle of the axis of the shaft with respect to the perpendicular restored from the horizontal plane of the condyles.
The angle of the axis of the shaft with respect to the perpendicular restored from the horizontal plane of the condyles.
19 Степень изогнутости диафиза или расстояние между передней поверхностью диафиза на середине длины кости и плоскостью, проведенной между задними поверхностями головки и мыщелков, измеренное в сагиттальной плоскости. The degree of curvature of the diaphysis or the distance between the anterior surface of the diaphysis in the middle of the bone length and the plane drawn between the posterior surfaces of the head and the condyles, measured in the sagittal plane.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ИХ ОБСУЖДЕНИЕ
Изменения картины факторных нагрузок на исследуемые параметры плечевых костей контрлатеральных конечностей человека и их дисперсии прекратились при выделении 3 факторов. Для бедренных костей — при выделении 4 факторов.
При анализе факторной структуры костей независимо от принадлежности кости к поясу конечностей и стороне тела были обнаружены постоянно присутствующие параметры. Их мы приняли в качестве стабильных базовых структурных параметров первого уровня (рисунки 2-3). Параметры, характерные для структурной организации костей, относящихся к одному поясу конечностей — либо плечевой, либо бедренной, мы рассматривали в качестве стабильных базовых структурных параметров второго уровня. Параметры, присутствующие в структуре лишь одной из костей — правой плечевой, левой плечевой, правой бедренной или левой бедренной, мы считали системно нестабильными.
В структурной организации плечевой и бедренной костей человека независимо от принадлежности к поясу конечностей и стороне тела определилась группа стабильных базовых параметров первого уровня, состоящая из вертикального диаметра головки, диаметров шейки, длины кости, ширины дистального эпифиза, сагиттального диаметра медиального гребня блока плечевой кости или медиального мыщелка бедренной кости.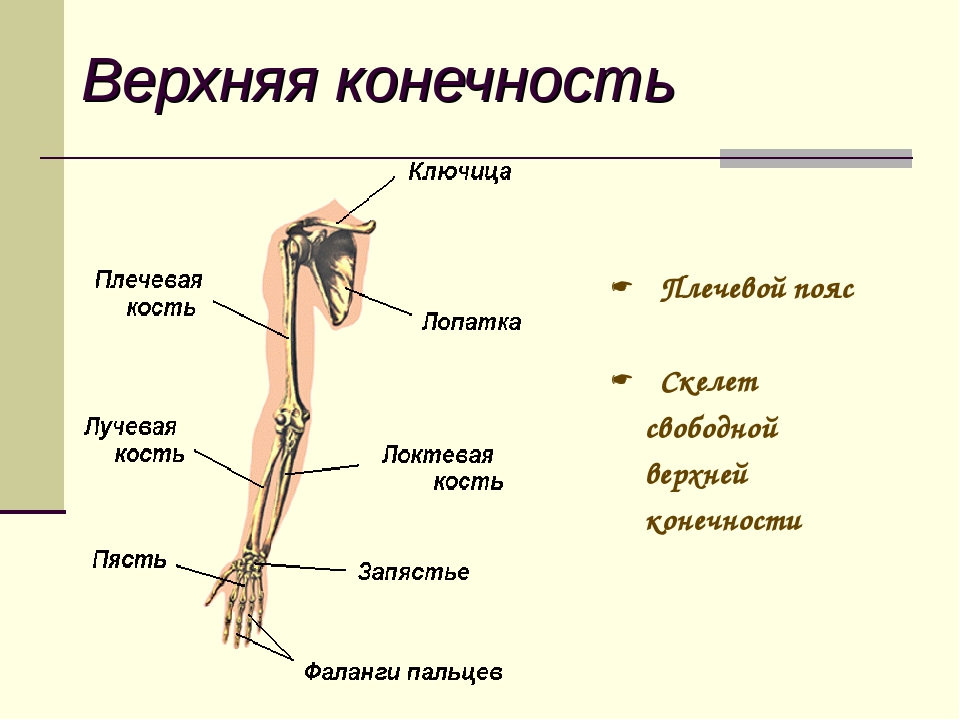 Данные параметры участвуют в передаче веса тела на нижележащие отделы конечности и указывают на прохождение механической оси конечностей через медиальные отделы дистального эпифиза костей [4, 8]. Трансляция веса через диафиз бедренной кости приводит к появлению его изогнутости в сагиттальной плоскости. В связи со снижением опорной роли конечности, для плечевой кости не характерно наличие выраженной изогнутости кпереди. Хотя, ряд исследователей указывают на сагиттальную вытянутость сечения диафиза кости [1]. В любом случае, параметры плечевых и бедренных костей, обеспечивающие трансляцию веса, будут являться стабильными базовыми структурообразующими параметрами первого уровня (рис. 2, 3).
Данные параметры участвуют в передаче веса тела на нижележащие отделы конечности и указывают на прохождение механической оси конечностей через медиальные отделы дистального эпифиза костей [4, 8]. Трансляция веса через диафиз бедренной кости приводит к появлению его изогнутости в сагиттальной плоскости. В связи со снижением опорной роли конечности, для плечевой кости не характерно наличие выраженной изогнутости кпереди. Хотя, ряд исследователей указывают на сагиттальную вытянутость сечения диафиза кости [1]. В любом случае, параметры плечевых и бедренных костей, обеспечивающие трансляцию веса, будут являться стабильными базовыми структурообразующими параметрами первого уровня (рис. 2, 3).
Таблица 3
Table 3
Обозначение структур дистального эпифиза плечевых и бедренных костей человека The osteometric parameters of the distal epiphysis structures of human humerus and femur
Номер Number Обозначение The osteometric parameters
20 Ширина дистального эпифиза — наибольшее расстояние между надмыщелками. The width of the distal epiphysis is the greatest distance between the epicondyles.
21 Ширина локтевой ямки плечевой кости / Ширина межмыщелковой ямки бедренной кости -внутреннее расстояние между краями ямок. The width of the ulnar fossa of humerus / The width of the intercondylar fossa of femur, it is the internal distance between the edges of the pits.
22 Высота локтевой ямки.
The length of the antecubital fossa.
23 Ширина лучевой ямки.
The width of the radial fossa.
24 Ширина венечной ямки.
The width of the coronal fossa.
25 Ширина суставной поверхности для надколенника.
The width of the articular surface for the patella.
26 Сагиттальный диаметр медиального гребня блока плечевой кости или сагиттальный размер медиального мыщелка бедренной кости. The sagittal diameter of the medial crest of the humerus trochlea or the sagittal size of the medial condyle of femur.
27 Сагиттальный диаметр латерального гребня блока плечевой кости или сагиттальный размер латерального мыщелка бедренной кости. The sagittal diameter of the lateral crest of the humerus trochlea or the sagittal size of the lateral condyle of femur.
28 Вертикальный диаметр медиального гребня блока плечевой кости.
The vertical diameter of the medial crest of the humerus trochlea.
29 Вертикальный диаметр латерального гребня блока плечевой кости.
The vertical diameter of the lateral crest of the humerus trochlea.
30 Ширина медиальной части суставной поверхности блока для плечевых костей. Ширина суставной поверхности медиального мыщелка для бедренных костей. The width of the medial part of the articular surface of the humerus trochlea. The width of the articular surface of the medial condyle for femur.
31 Ширина латеральной части суставной поверхности блока для плечевых костей. Ширина суставной поверхности латерального мыщелка для бедренных костей. The width of the lateral part of the articular surface of the humerus trochlea. The width of the articular surface of the lateral condyle for femur.
The width of the lateral part of the articular surface of the humerus trochlea. The width of the articular surface of the lateral condyle for femur.
32 Ширина суставной поверхности головочки, ограниченная латеральным гребнем, измеренная на середине высоты передней поверхности мыщелка.
The width of the articular surface of the head limited by the lateral crest, measured in the middle of the height of the anterior surface of the condyle.
33 Вертикальный диаметр головочки — измеряется между нижней и верхней точками суставной поверхности головочки при виде спереди.
The vertical diameter of the head is measured between the lower and upper points of the articular surface of the head when viewed from the front.
34 Ширина медиального надмыщелка — расстояние, измеренное горизонтально от максимально выступающей точки надмыщелка до края суставной поверхности. The width of the medial epicondyle is the distance measured horizontally from the maximally protruding point of the epicon-dyle to the edge of the articular surface.
35 Ширина латерального надмыщелка — расстояние, измеренное горизонтально от максимально выступающей точки надмыщелка до края суставной поверхности. The width of the lateral epicondyle is the distance measured horizontally from the maximally protruding point of the epicon-dyle to the edge of the articular surface.
Примечание: Серым цветом обозначены ячейки структур, характерных только для одной из костей.
Note: The gray color indicates the structures that are specific to only one of the bones.
Правая плечевая кость Right humerus
Левая плечевая кость Left humerus
Рис. 2. Структурная организация плечевых костей человека.
Примечание: На схеме представлены параметры, имеющие факторные нагрузки с силой |0,7-1,0| при p>0,05. Отрицательным знаком обозначено обратное влияние. Номера параметров соответствуют данным таблиц 1-3.
Fig. 2. Structural organization of human humerus.
Note: the diagram shows the parameters having factor loads with force |0.7-1.0| at p> 0.05. A negative sign indicates the opposite effect. The parameter numbers correspond to the data in tables 1-3.
Правая бедренная кость Левая бедренная кость
Right femur Left femur
Рис. 3. Структурная организация бедренных костей человека.
Примечание: На схеме представлены параметры, имеющие факторные нагрузки с силой |0,7-1,0| при p>0,05. Отрицательным знаком обозначено обратное влияние. Номера параметров соответствуют данным таблиц 1-3.
Fig. 3. The structural organization of the human femur.
Note: the diagram shows the parameters having factor loads with force |0.7-1.0| at p> 0.05. A negative sign indicates the opposite effect. The parameter numbers correspond to the data in tables 1-3.
Особенностью структурной организации плечевой кости независимо от принадлежности к стороне тела явилось выделение ширины проксимального эпифиза в качестве базового структурного параметра первого уровня. Данный параметр, измеренный от медиального края суставной головки до латерального края большого бугорка, отражает функционирование m. pectoralis major и m. supraspinatus, m. infraspinatus, m. teres minor, которые обеспечивают отведение — приведение и вращение руки в плечевом суставе вокруг вертикальной оси.
Особенности структурной организации бедренной кости проявились наличием факторных нагрузок на сагиттальный диаметр головки и межвертельное расстояние. Данные параметры мы отнесли к стабильным базовым структурообразующим параметрам бедренной кости первого уровня. Сферичность головки, подтвержденная факторными нагрузками на вертикальный и сагиттальный диаметры головки бедренной кости, наряду с факторными нагрузками на межвертельное расстояние (место фиксации мышц, обеспечивающих вертикальное положение туловища и вращательные движения в тазобедренном суставе) свидетельствует об увеличении влияния прямохождения при формировании структуры бедренной кости человека.
На уровне дистальных эпифизов структурная организация костей показывает отличия, связанные с функциональной дифференциацией конечностей. В структурной организации плечевой кости человека базовую роль играют параметры, обеспечивающие максимальную амплитуду сгибательно-разгибательных и ротационных движений в локтевом суставе. Это диаметры латерального гребня блока, размеры головочки блока плечевой кости и ширина латерального надмыщелка. Последний является местом прикрепления m. brachioradialis, mm. extensor carpi radialis longus et brevis, mm. extensor digitorum et digiti minimi, m. extensor carpi ulnaris и m. supinator — мышц, обеспечивающих разгибание в нижележащих суставах, супинацию предплечья и кисти в физиологическое состояние [7, 9]. Перечисленные параметры также входят в группу стабильных структурных параметров плечевой кости первого уровня.
Активные сгибательно-разгибательные движения при ходьбе наряду с поддержанием вертикального положения тела на уровне коленного сустава невозможно без симметричного участия обоих мыщелков бедренной кости, что подтверждается факторными нагрузками на латеральный мыщелок бедра. Ранее нами было обнаружено, что механизмы «замыкания» локтевого и коленного суставов [3], как стабилиза-
торы вертикальной оси конечностей животных, у человека на уровне плечевой кости достаточно сильно редуцировались. Это проявилось отсутствием стабильных базовых структурообразующих параметров в организации плечевой кости. Механизм «замыкания» коленного сустава у человека остался сохранным. Стабильными базовыми структурными параметрами бедренной кости второго уровня явились ширина суставной поверхности надколенника и ширина ме-жмыщелковой ямки, показывающие отрицательные факторные нагрузки. Кроме этого, пря-мохождение невозможно без активного участия приводящих мышц. Нижняя порция m. adductor magnus прикрепляется на медиальном надмы-щелке бедра, образуя приводящий бугорок -tuberculum adductorium. M. adductor magnus проходит позади оси тазобедренного сустава, она является его разгибателем, одновременно приводя и супинируя конечность. С точки зрения биомеханики нижняя конечность человека представляет собой многозвеньевой механизм, основу функционирования которого составляет чередующееся расположение мышц-
антагонистов. И если вверху к медиальному надмыщелку прикрепляется разгибатель тазобедренного сустава, то к его нижней поверхности прикрепляется m. gastrocnemius, сгибатель коленного сустава, препятствующая запрокидыванию тела кпереди [4]. Таким образом, медиальный надмыщелок бедренной кости вместе с шириной надколенниковой поверхности и ме-жмыщелковой ямки бедренной кости являются стабильными базовыми параметрами, обеспечивающими вертикальное положение тела при стоянии.
Возникающие в ходе эволюции человека морфофункциональные отличия конечностей проявляются наличием нестабильных параметров в структурной организации плечевой и бедренной костей. Появление таких параметров указывает не только на асимметричность структурной организации сравниваемых костей, характерную для обоих поясов конечностей, но и на наличие вертикальной дифференциации их эпифизов. Исходя из результатов исследования, организация бедренной кости более структурна. Нестабильных параметров проксимального эпифиза правой бедренной кости выделено 4, дистального эпифиза — 1; в структурной организации проксимального эпифиза левой бедренной кости выделен 1, дистального эпифиза — 3. При этом нестабильными параметрами проксимального эпифиза правой бедренной кости оказались ширина проксимального эпифиза, верхняя и нижняя длины шейки и угол антеверсии шейки и головки бедра. Эти параметры обеспечивают передачу веса тела во фронтальной
плоскости и соответствуют прохождению механической оси конечности через головку тазобедренного сустава [5, 6, 8]. Данная особенность структурной организации проксимального эпифиза правой бедренной кости, на наш взгляд, указывает на большее участие правой бедренной кости в реализации опорной функции. Нестабильными параметрами дистального эпифиза левой бедренной кости явились ширина латерального и медиального мыщелка и ширина латерального надмыщелка. Функциональная роль выделенных структур заключается в передаче веса через коленный сустав. Таким образом, в структурной организации бедренной кости максимально ярко проявилось явление вертикального разделения эпифизов контрлатеральных костей по выполняемым функциям.
Структурная организация плечевых костей резко асимметрична, что связано с явлениями функционального доминирования одной из рук. Правая плечевая кость отличается меньшим числом коррелированных нестабильных параметров: в структуре проксимального эпифиза выделено 3 параметра, в структуре дистального — 2. При этом в структуре эпифизов левой плечевой кости нестабильных параметров выделено в два раза больше. При рассмотрении функциональной роли выделенных параметров, обращает на себя внимание факт присутствия на разных уровнях структурной организации обеих плечевых костей параметров, свидетельствующих о различной двигательной активности конечности в плечевом суставе. Это размеры межбугорковой борозды, содержащие длинную головку бицепса, угол хрящевого края головки с диафизом, угол наклона диафиза. Факторные нагрузки на ширину и глубину межбугорковой борозды свидетельствуют о асимметричности выполнения сгибания и разгибания в плечевом суставе. Амплитуда движений правого плечевого сустава в сагиттальной плоскости будет лимитироваться величиной углов, характеризующих стереометрию головки плечевой кости и влияющих на биомеханику двуглавой мышцы плеча. В структуре дистального эпифиза правой кости присутствуют параметры, определяющие объем вращательных движений предплечья вокруг лучевой кости. Факторные нагрузки на межбугорковое расстояние, определяемые в структуре левой плечевой кости, указывают на большую вращательную активность в плечевом суставе, присущую левой плечевой кости. В структуре дистального эпифиза левой плечевой кости большую роль играют параметры, обеспечивающие передачу силовых нагрузок по оси конечности: высота локтевой ямки, ширина суставных поверхностей блока, ширина медиального надмыщелка.
Вывод: независимо от принадлежности к поясам конечностей и стороне тела, основу структурной организации плечевой и бедренной костей человека формируют анатомические образования, участвующие в трансляции силовых нагрузок по осям конечностей. Морфо-функциональная асимметрия контрлатеральных плечевых и бедренных костей человека, а также различия функциональной специализации их эпифизов определяются количеством уровней системной организации и спектром факторных нагрузок.
КОНФЛИКТ ИНТЕРЕСОВ Авторы декларируют отсутствие явных и потенциальных конфликтов интересов, связанных с публикацией настоящей статьи.
ИСТОЧНИКИ ФИНАНСИРОВАНИЯ Авторы заявляют об отсутствии финансирования.
СООТВЕТСТВИЕ ПРИНЦИПАМ ЭТИКИ Исследование костного материала проведено с соблюдением всех этических норм и правил с разрешения РЭК КГМУ, протокол № 5 от 30.06.2018 г.
ЛИТЕРАТУРА/REFERENCES
1. Николенко В.Н., Фомичева О.А. Индивидуальная и типовая анатомическая изменчивость макро-микроскопического строения плечевой кости. Саратовский научно-медицинский журнал. 2007; 3: 29-31. [Nikolenko V.N., Fomicheva O.A. Individual and typical anatomical variability of the macro-microscopical structure of humeral bone. Saratov Journal of Medical Scientific Research. 2007; 3: 29-31. (in Russ.)]
2. Яшина И.Н., Иванов Д.А., Иванов А.В., Колесник А.И., Самаха А.А., авторы; Яшина И.Н., Иванов Д.А., Иванов А.В., Колесник А.И., Сама-ха А.А., патентообладатели. Устройство для антропометрических измерений длинных трубчатых костей. Российская Федерация патент RU 2245101. 27 января 2005 г. [Yashina I.N., Ivanov D.A., Ivanov A.V., Kolesnik A.I., Samakha A.A., inventors; Yashina I.N., Ivanov D.A., Ivanov A.V., Kolesnik A.I., Samakha A.A., assignees. Device for anthropometric measurements of long tubular bones. Russian Federation patent RU 2245101. 2005 Jan 27 (in Russ.)].
3. Яшина И.Н., Клочкова С.В., Иванов А.В., Вагапо-ва В.Ш. Адаптационная гармоничность в строении проксимального эпифиза плечевых костей человека и некоторых животных. Медицинский вестник Башкортостана. 2018; 13(6-78): 51-54. [Yashina I.N., Klochkova S.V., Ivanov A.V., Vagapo-va V.Sh. Adaptive harmony in the structure of the proximal epiphysis of the humerus in humans and some animals. Meditsinskiy vestnik Bashkortostana. 2018; 13(6(78)): 51-54. (in Russ.)]
4. Cronin N.J., Avela J., Finni T., Peltonen J. Differences in contractile behaviour between the soleus and medial gastrocnemius muscles during human walking. J Exp Biol. 2013; 216(Pt 5): 909-914. DOI: 10.1242/jeb.078196.
5. Fong D.T., Chan Y.Y. The Use of Wearable Inertial Motion Sensors in Human Lower Limb Biomechanics Studies: A Systematic Review. Sensors (Basel). 2010; 10(12): 11556-11565. DOI: 10.3390/s101211556.
6. Foucher K.C. Gait abnormalities before and after total hip arthroplasty differ in men and women. J Biomech. 2016; 49(14): 3582-3586.
DOI: 10.1016/j.jbiomech.2016.09.003
7. Gülegyüz M.F., Pietschmann M.F., Michalski S., Eberhard F.M., Crispin A., Schröder C., Mittermüller M.J., Müller P.E. Reference Values of Flexion and Supination in the Elbow Joint of a Cohort without Shoulder
Pathologies. Biomed Res Int. 2017; 2017: 1654796. DOI: 10.1155/2017/1654796.
8. Mu X., Chen J., Du F. Biomechanics analysis of human lower limb during walking for exoskeleton design. Journal of Vibroengineering. 2017; 19(7): 5527553. DOI: 10.21595/jve.2017.18459
9. Roach N., Lieberman D.E., Gill Iii T. J., Palmer W.E. The effect of humeral torsion on rotational range of motion in the shoulder and throwing performance. JAnat. 2012; 220(3): 293-301. DOI: 10.1111/j.1469-7580.2011.01464.x.
10. Samaha A.A., Ivanov A.V., Haddad J.J., Kolesnik A.I., Baydoun S., Arabi M.R., Yashina I.N., Samaha R.A., Ivanov D.A. Asymmetry and structural system analysis of the proximal femur meta-epiphysis: osteoartic-ular anatomical pathology. J Orthop Surg Res. 2008; 3: 11. DOI: 10.1186/1749-799X-3-11
Поступила в редакцию 05.12.2018 Подписана в печать 21.03.2019
Для цитирования: Яшина И.Н., Иванов А.В., Клочкова С.В. К вопросу гомологии в структурной организации костей проксимальных сегментов конечностей человека. Курский научно-практический вестник «Человек и его здоровье». 2019;(1):83-92. DOI: 10.21626/vestnik/2019-1/10.
TO THE QUESTION OF HOMOLOGY IN THE STRUCTURAL ORGANIZATION OF BONES
OF HUMAN LIMB PROXIMAL SEGMENTS
© Yashina I.N.1, IvanovA.V.1, Klochkova S.V.2
1 Kursk State Medical University (KSMU)
3, K. Marx str., Kursk, Kursk region, 305041, Russian Federation
2 I.M. Sechenov First Moscow State Medical University (Sechenov University)
11, building 10, Mokhovaya str, Moscow, 125009, Russian Federation
Objective. The main aim of the research is to discover regularities and specific features of systemic structural organization of the human humeral and femoral bones according to the functions of extremities and their belonging to the side of the body.
Materials and methods. By the classic osteometry with some authors’ modifications 35 anatomical structures on 308 of humerus and femur have been measured. All the bones had no pathological symptoms, with complete synostosis of epiphyses. To access the somatotype effect on the dimensions of human bone structures all the values obtained were normalized. As a unit for measurement normalization the transverse diameter of the shaft in the middle of the bone length was chosen. The values obtained were processed by the methods of factor analysis Maximum Maximum Likelihood Factor with rotation Equamax normalized, and four groups were generated according to the limb girdle and the side of the body.
Results. The asymmetry of the structural organization of bones and vertical differentiation of their epiphyses are determined by the presence of unstable parameters. The asymmetry of the structural organization of the femur is more pronounced. Unstable parameters were found in the proximal epiphysis of the right femur — 4, in the distal epiphysis — 1; in the proximal epiphysis of the left femur — 1, in the distal epiphysis — 3. Unstable parameters of the proximal epiphysis of the right humerus made up 3, distal epiphysis — 2; in the structural organization of the epiphysis of the left humerus 6 unstable parameters were identified.
Conclusion. The basis of the structural organization of the bone proximal segments of the human limbs regardless of belonging to the extremity or the side of the body is formed by anatomical objects which are responsible for conveying the power loads along the axis of the limb. Functional differentiation of human limbs is shown in the structural organization of both bones by asymmetric vertical differentiation of their epiphysis.
Keywords: humerus; femur; structural organization; factor analysis.
Yashina Irina N. — PhD in Medicine, Associate Professor of Human Anatomy Department, KSMU, Kursk, Russian Federation. ORCID iD: 0000-0001-6235-2309. E-mail: [email protected] (correspondence author)
Ivanov Alexander V. — DM, Professor, Head of the Department of Histology, Cytology and Embryology, KSMU, Kursk, Russian Federation. ORCID iD: 0000-0002-2412-0475. E-mail: [email protected]
Klochkova Svetlana V. — DM, Professor of the Department of Human Anatomy, Sechenov University, Moscow, Russian Federation. ORCID iD: 0000-0003-2041-7607. E-mail: [email protected]
CONFLICT OF INTEREST The authors declare the absence of obvious and potential conflicts of interest related to the publication of this article.
SOURCE OF FINANCING The authors state that there is no funding for the study.
CONFORMITY WITH THE PRINCIPLES OF ETHICS The study of bone material was carried out in compliance with all ethical norms and rules and was approved by the Regional Ethical Committee under KSMU (Protocol No. 5 of 30.06.2018).
Received 05.12.2018 Accepted 21.03.2019
For citation: Yashina I.N., Ivanov A.V., Klochkova S.V. To the question of homology in the structural organization of bones of human limb proximal segments. Kurskiy nauchno-prakticheskiy vestnik «Chelovek i ego zdorov’ye» = Kursk Scientific and Practical Bulletin «Man and His Health». 2019;(1):83-92. DOI: 10.21626/vestnik/2019-1/10.
Скелет верхних и нижних конечностей | Биология. Реферат, доклад, сообщение, краткое содержание, лекция, шпаргалка, конспект, ГДЗ, тест
Тема: Костная система
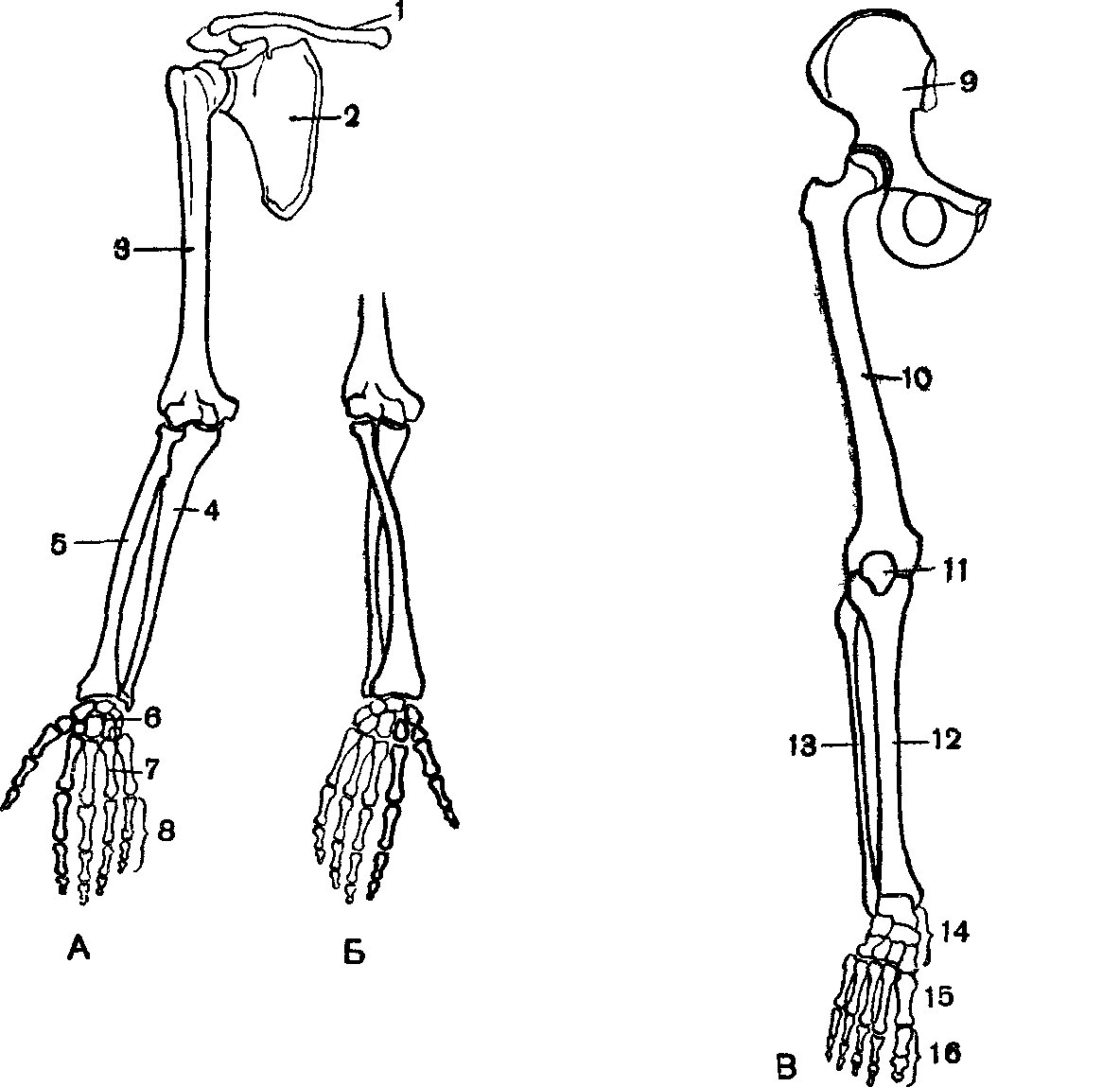
К скелету верхних конечностей относятся плечевой пояс — ключицы и лопатки, свободная верхняя конечность, которая состоит из плечевой кости, костей предплечья — локтевой и лучевой, а также кисти. Последняя состоит из запястья, пястья и костей пальцев (рис. 31, а). Все части кисти соединены между собой многими суставами, что делает её очень подвижной.
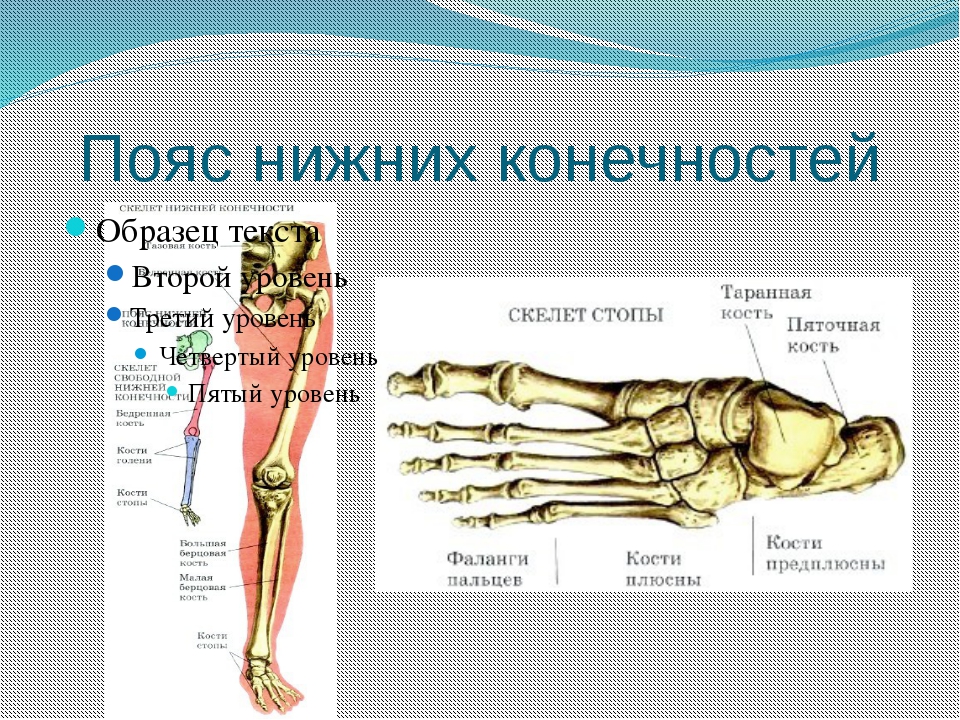
Нижние конечности человека — это единственный орган опоры и передвижения, а поэтому они образованы самыми большими и самыми прочными костями. К скелету нижних конечностей относится тазовый пояс, или таз, который состоит из крестца и двух тазовых костей. Благодаря вертикальному хождению у человека сформировался крепкий и широкий таз. Скелет свободной нижней конечности образован бедренной костью, двумя костями голени — большой берцовой и малой берцовой, а также костями стопы (рис. 31,6). Стопа человека, в отличие от высших позвоночных, имеет свод (рис. 30).
Скелет человека состоит из 206 костей. Материал с сайта http://worldofschool.ru
| Рис. 30. Скелет стопы человека (а) и обезьяны (б). 1 — свод стопы у человека |
| Рис. 31. Скелет кисти (а) и стопы (б): а) 1 — локтевая кость, 2 — лучевая кость, 3-8 — кости запястья, 9 — кости пястья, 10 — фаланги пальцев; б) 11 — пяточная кость, 12-17 — предплюсневые кости, 18— плюсневые кости, 19 — фаланги пальцев |
Биология нижние конечности птицы состоят
Скелет верхних конечностей в рост картинка
Скелет верхних конечностей тест
Верхние и нижние конечности конспект
Пояс свободных верхних конечностей рептилии
Объясните особенности строения и функций скелета верхних конечностей.
Объясните особенности строения и функций скелета нижних конечностей.
Пороки развития
Пороки и дефекты развития конечностей
Пороки развития — стойкие морфологические изменения органа, системы или организма, которые выходят за пределы вариаций их строения и возникают внутриутробно в результате нарушений развития зародыша или после рождения ребенка, как следствие нарушения дальнейшего формирования органов. Пороки развития возникают в результате генных мутаций; хромосомных и геномных мутаций; комбинированного воздействия генных мутаций и факторов внешней по отношению к зародышу среды; тератогенных факторов.
Аномалии, возникающие в результате недостаточности формирования частей конечностей.
Амелия — полное отсутствие конечности (исключая плечевой пояс и таз). Различают верхнюю и нижнюю амелию, в частности отсутствие двух верхних конечностей (абрахия), одной верхней конечности (монобрахия), двух нижних конечностей (апус), одной нижней конечности (монопус).
Фокомелия (тюленеобразные конечности) — отсутствие проксимальных и (или) средних частей конечности и соответствующих суставов (плечевого, тазобедренного). Различают проксимальную, дистальную и полную фокомелию. Проксимальная фокомелия — отсутствие плеча или бедра, дистальная — отсутствие предплечья (радиоульнарная форма) или голени (тибиофибулярная форма), полная фокомелия — отсутствие плеча и предплечья или бедра и голени. Соответственно вполне сформированная кисть или стопа может отходить непосредственно от туловища (полная фокомелия), соединяться с ним посредством сохранившихся костей предплечья, голени (проксимальная форма) или прикрепляться к плечу, бедру (дистальная форма). Фокомелия бывает одно- и двусторонней, иногда в процесс вовлекаются все четыре конечности.
Перомелия — вариант фокомелии, сочетающейся с недоразвитием кистей или стоп. Различают полную (рука или нога отсутствует, соответствующий отдел туловища заканчивается одним рудиментарным пальцем или кожным выступом) и неполную (плечо или бедро недоразвито, заканчивается также одним рудиментарным пальцем или кожным выступом) формы.
Встречаются также лучевая и локтевая косорукость, аплазия большеберцовой кости, аплазия малоберцовой кости, адактилия — отсутствие пальцев, афалангия — отсутствие фаланг, монодактилия — наличие одного пальца на кисти или стопе, ахейрия — отсутствие кисти.
Расщепление кисти (эктродактилия, клешнеобразная кисть, «кисть омара») — аплазия центральных компонентов кисти (пальцев и нередко пястных костей) с бороздой (расщелиной) на месте отсутствующих костей. Выделяют типичные и атипичные формы. Типичная форма характеризуется аплазией III пальца и (нередко) соответствующей пястной кости, а также (иногда) дистального ряда костей запястья, что обусловливает наличие глубокой расщелины. Атипичная расщелина проявляется недоразвитием (реже отсутствием) средних пальцевых компонентов кисти или стопы. При атипичной форме расщелина неглубокая, но широкая; иногда она имеет вид чрезмерно широкого межпальцевого промежутка. Чаще встречается расщепление правой кисти.
Аномалии, возникающие в результате недостаточной дифференцировки частей конечности.
К ним относятся пороки развития лопатки (ладьевидная лопатка, поднятая лопатка), синостозы, синдактилии, брахидактилия, врожденная косолапость, врожденный вывих бедра, артрогрипоз, клинодактилия — укорочение средней фаланги пальцев кистей (чаще V пальца), обычно являющиеся составным компонентом синдромов.
Камптодактилия (кампилодактилия) — сгибательная контрактура проксимальных межфаланговых суставов пальцев кисти. В процесс может вовлекаться любой палец, кроме I. Встречается редко.
Сиреномелия (симподия, симмелия, синдром каудальной регрессии) — слияние нижних конечностей. Слияние может касаться мягких тканей и (чаще) некоторых длинных трубчатых костей, а также сопровождаться гипо- и (или) аплазией отдельных костей конечностей и таза. Стопы могут отсутствовать (sympus apus), бывают сформированы две (sympus dipus) или одна стопа (sympus monopus). Иногда имеется одна рудиментарная стопа с единственным пальцем. Сиреномелия сопровождается аплазией наружных и внутренних половых органов, аплазией мочевой системы, атрезией заднепроходного отверстия и прямой кишки, аплазией одной пупочной артерии.
Аномалии, обусловленные удвоением: полидактилия, диплоподия — удвоение стопы, полимелия — увеличение числа нижних конечностей. Полимелия может быть симметричной и асимметричной, обычно сочетается с пороками, несовместимыми с жизнью.
Аномалии, обусловленные чрезмерным ростом, включают макотородактилию и гигантизм конечности (парциальный гигантизм, односторонняя макросомия, гемигипертрофия), проявляющийся односторонним увеличением относительно пропорционально развитой конечности.
Аномалии, обусловленные недостаточным ростом. В их число входят аномалии, проявляющиеся гипоплазией различных отделов костей конечностей.
Врожденные перетяжки — порок развития амниона в виде тканевых тяжей, проходящих внутри плодовместилища и связывающих между собой плодовую поверхность последа с поверхностью плода, разные точки плодовой поверхности последа и несколько точек поверхности плода.
Генерализованные (системные) скелетные деформации. В их основе лежит нарушение эмбриогенеза соединительной ткани, включая костную ткань. К этой группе относятся хондродисплазии, остеодисплазии.
Клинические проявления пороков и прогноз во многом зависят от того, насколько жизненно важным является пораженный орган, от степени нарушения его функций, а также от присоединившихся осложнений.
Основная часть пороков опорно-двигательного аппарата корригируются хирургическим путем. Применяются различные пластические операции на коже и мягких тканях, так же костно-пластические операции при выраженных деформациях конечностей и позвоночника с применением различных корригирующих систем и аппаратов внешней фиксации, а так же костной пластики различными трансплантатами.
Посттравматические состояния верхних и нижних конечностей
Перелом кости – это травма, которая характеризуется частичным или полным нарушением целостности костной ткани и может стать результатом прямого удара (травматический перелом здоровой кости), когда разрыв костной ткани вызван действием механической силы, превышающей твердость костей. Или перелом может случится в результате заболеваний, связанных с изменением структуры и характеристик костной ткани (спонтанный или патологический перелом). Этот тип переломов может произойти и после незначительной травмы (например, переломы из-за остеопороза). Чаще всего встречаются переломы конечностей. Возможны и переломы других костей: грудная клетка, голова и позвоночник, точнее перелом позвонков.
На основании того, дошло ли при переломе до разрыва кожи и подкожной ткани, и есть ли соприкосновение места перелома с внешней средой различают: открытый перелом (на практике тяжелее подвергается лечению и восстановлению) и закрытый перелом.
По тяжести перелом может быть: сложный – при котором кость повреждена в двух или более местах, и простой перелом или трещина — без разделения сломанной кости на сегменты.
Классификация по действию механической силы при переломе: местный перелом — происходит в месте приложения механической силы (обычно более тяжёлый перелом) и отдаленный перелом — встречается чаще и возникает вдали от места приложения силы, или на дистальных концах конечностей.
Тип и тяжесть перелома зависит от механизма повреждения, мощности и направления силы, действующей на кость, а также от свойств самой кости (прочность и тип кости, часть кости, на которую действует травматическая сила).
В клинической картине различают абсолютные и относительные признаки перелома. Абсолютные признаки: патологическая подвижность, физическое обследование сопровождается сильной болью, явление крепитации костных отломков, деформация продольной оси конечности в месте перелома. Относительные признаки: отёк, боль при давлении, боль при передвижении повреждённой части тела, изменение цвета кожи, мышечный спазм.
Чтобы начался процесс заживления перелома, части сломанной кости должны быть установлены рядом друг с другом так, чтобы они касались. Таким образом создаются условия для нормального функционирования одновременно многих процессов, конечным результатом которых является полное структурное и функциональное восстановление сломанных костей .
В современной медицине применяются два основных метода лечения переломов: консервативный (гипс, иммобилизация) и оперативный (хирургическое), когда части сломанной кости соединяются металлическими направляющими спицами, штифтами и винтами. Выбор метода лечения переломов зависит от состояния пациента и типа перелома. Решение принимается хирургом-ортопедом. Как только состояние больного позволит это, необходимо начать восстановление. Конечной целью применения физиотерапии является наиболее полное восстановление функций пациента.
Физиотерапия в так называемой ранней стадии включает в себя применение электротерапии и магнитотерапии, которые могут проводится через гипс и в присутствии металла в зоне перелома. Для повреждённой конечности значительным является и использование статических упражнений.
Врач физиотерапевт, исходя из того, насколько уже сраслась сломанная кость, клинических анализов, возможных осложнений, и когда состояние пациента и сама травма это позволяют, внимательно и строго индивидуально дозирует и комбинует различные физиотерапевтические процедуры. Всё это для того, чтобы ускорить срастание костей, уменьшить отёк и боль, нормализовать местное кровообращение, воспрепятствовать образованию контрактуры и увеличить подвижность суставов, укрепить мышцы. На поздних этапах реабилитации могут быть применены гидро — кинези терапевтические и все бальнеотерапевтические процедуры (лечебная грязь и минеральная вода). Врач физиотерапевт должен учитывать противопоказания к проведению каждой процедуры в отдельности.
Своевременная, хорошо дозируемая и адекватная физикальная терапия сокращает длительность восстановления пациента после перелома и его быстрому возвращению к нормальной жизнедеятельности.
38.1D: Аппендикулярный скелет человека — Биология LibreTexts
Аппендикулярный скелет поддерживает прикрепление и функции верхних и нижних конечностей человеческого тела.
Задачи обучения
- Описать кости и функции аппендикулярного скелета человека
Ключевые моменты
- Аппендикулярный скелет человека состоит из костей верхних конечностей, нижних конечностей, грудного и тазового пояса.
- Грудной пояс служит точкой прикрепления верхних конечностей к телу.
- Верхняя конечность состоит из руки, предплечья, запястья и кисти.
- Тазовый пояс отвечает за вес тела и за передвижение; он также отвечает за прикрепление нижних конечностей к телу.
- Нижние конечности, включая бедра, ноги и ступни, поддерживают весь вес тела и поглощают возникающие в результате движения силы.
Ключевые термины
- осевой скелет : кости головы и туловища организма
- аппендикуляр : конечность или придаток или относящиеся к ним
- ключица : ключица; выступающая кость в верхней части груди между плечом и шеей
- лопатка : одна из двух больших плоских костей, образующих заднюю часть плеча
- шарнирный : для образования соединения или соединения посредством шарниров
Аппендикулярный скелет человека
Аппендикулярный скелет человека состоит из костей верхних конечностей (которые служат для захвата предметов и манипулирования ими) и нижних конечностей (которые позволяют передвигаться).Он также включает грудной (или плечевой) пояс и тазовый пояс, которые прикрепляют верхние и нижние конечности к телу соответственно.
Рисунок \ (\ PageIndex {1} \): Аппендикулярный скелет : Аппендикулярный скелет состоит из костей грудных конечностей (рука, предплечье, кисть), тазовых конечностей (бедро, нога, ступня), грудного пояса. , и тазовый пояс.Грудной ремень
Кости грудного пояса, обеспечивающие точки прикрепления верхних конечностей к осевому каркасу, состоят из ключицы (или ключицы) спереди, а также лопатки (или лопаток) сзади.Ключицы, S-образные кости, которые позволяют расположить руки на теле, лежат горизонтально через переднюю часть грудной клетки (грудь) чуть выше первого ребра.
Рисунок \ (\ PageIndex {1} \): Грудной пояс : (a) Грудной пояс приматов состоит из ключиц и лопаток. (b) Вид сзади показывает ость лопатки, к которой прикрепляется мышца.Лопатки — это плоские треугольные кости, расположенные в задней части грудного пояса. Они поддерживают мышцы, пересекающие плечевой сустав.Позвоночник проходит через заднюю часть лопатки; это хороший пример костного выступа, который обеспечивает широкую область прикрепления мышц к кости.
Верхние конечности
Верхние конечности содержат 30 костей в трех областях: рука (плечо до локтя), предплечье (локтевая и лучевая кость), а также запястье и кисть. Плечевая кость — самая большая и длинная кость верхней конечности и единственная кость руки. Он сочленяется (соединяется) с лопаткой в плече и с предплечьем в локте.Предплечье, простирающееся от локтя до запястья, состоит из двух костей: локтевой и лучевой. Радиус, расположенный вдоль боковой (большой) стороны предплечья, сочленяется с плечевой костью в локтевом суставе. Локтевая кость, расположенная на медиальной стороне (стороне мизинца) предплечья, длиннее лучевой кости. Он сочленяется с плечевой костью в локте. Лучевая и локтевая кости также соединяются с костями запястья и друг с другом, что у позвоночных обеспечивает различную степень вращения запястья по отношению к длинной оси конечности.Рука состоит из восьми костей запястья (запястья), пяти костей пястной кости (ладони) и 14 костей фаланг (пальцев). Каждый палец состоит из трех фаланг, за исключением большого пальца, у которого их всего две.
Рисунок \ (\ PageIndex {1} \): Верхняя конечность : Верхняя конечность состоит из плечевой кости плеча, лучевой кости и локтевой кости предплечья, восьми костей запястья, пяти костей пястной кости и 14 костей. кости фаланг.Тазовый ремень
Тазовый пояс прикрепляется к нижним конечностям осевого скелета и отвечает за перенос веса тела и за передвижение.Он надежно прикреплен к осевому каркасу прочными связками. Он также имеет глубокие гнезда с прочными связками для надежного прикрепления бедренной кости к телу. Тазовый пояс дополнительно укреплен двумя большими тазобедренными костями. У взрослых тазобедренные кости образуются путем слияния трех пар костей: подвздошной, седалищной и лобковой. Таз соединяется в передней части тела с лонным симфизом и с костями крестца в задней части тела.
Нижние конечности
Нижние конечности состоят из бедра, голени и стопы.Кости нижней конечности — это бедренная кость (бедренная кость), надколенник (коленная чашечка), большеберцовая и малоберцовая кости (кости голени), предплюсны (кости голеностопного сустава), плюсневые кости и фаланги (кости стопы). Кости нижних конечностей толще и прочнее, чем кости верхних конечностей, из-за необходимости выдерживать весь вес тела вместе с силами, возникающими в результате передвижения.
Рисунок \ (\ PageIndex {1} \): Нижняя конечность : Нижняя конечность состоит из бедра (бедра), коленной чашечки (надколенника), голени (голени и малоберцовой кости), лодыжки (предплюсны) и стопы (плюсневые кости и фаланги). ) кости.Бедренная кость или бедренная кость — самая длинная, тяжелая и крепкая кость в теле. Бедренная кость и таз на проксимальном конце образуют тазобедренный сустав. На дистальном конце бедро, голень и надколенник образуют коленный сустав. Коленная чашечка, или коленная чашечка, представляет собой треугольную кость, расположенную перед коленным суставом; он встроен в сухожилие разгибателей бедра (четырехглавой мышцы). Улучшает разгибание колена за счет уменьшения трения. Большеберцовая кость, или большеберцовая кость, представляет собой большую кость ноги, которая расположена непосредственно под коленом.Большеберцовая кость сочленяется с бедренной костью на ее проксимальном конце, с малоберцовой костью и костями предплюсны на ее дистальном конце. Как вторая по величине кость в человеческом теле, она отвечает за передачу веса тела от бедренной кости к стопе. Малоберцовая кость, или кость теленка, проходит параллельно большеберцовой кости и сочленяется с ней. Он не несет веса, но действует как место прикрепления мышц, образуя боковую часть голеностопного сустава.
Заплюсны — это семь костей лодыжки, по которым вес тела передается от большеберцовой и малоберцовой костей к стопе.Плюсны — это пять костей стопы, а фаланги — 14 костей пальцев ног.
Рисунок \ (\ PageIndex {1} \): Стопа и лодыжка : На этом рисунке показаны кости стопы и голеностопного сустава человека, включая плюсневые кости и фаланги.ЛИЦЕНЗИИ И АТРИБУЦИИ
CC ЛИЦЕНЗИОННЫЙ КОНТЕНТ, ПРЕДЫДУЩИЙ РАЗДЕЛ
- Курирование и проверка. Предоставлено : Boundless.com. Лицензия : CC BY-SA: Attribution-ShareAlike
CC ЛИЦЕНЗИОННОЕ СОДЕРЖАНИЕ, СПЕЦИАЛЬНАЯ АТРИБУЦИЯ
- Колледж OpenStax, Биология.17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44783/latest…ol11448/latest . Лицензия : CC BY: Attribution
- Костно-мышечная система. Предоставлено : Википедия. Расположен по адресу : http://en.Wikipedia.org/wiki/Musculoskeletal_System . Лицензия : CC BY-SA: Attribution-ShareAlike
- костно-мышечной системы. Предоставлено : Википедия. Расположен по адресу : en.Wikipedia.org/wiki/musculoskeletal%20system . Лицензия : CC BY-SA: Attribution-ShareAlike
- протез. Предоставлено : Викисловарь. Находится по адресу : en.wiktionary.org/wiki/prosthesis . Лицензия : CC BY-SA: Attribution-ShareAlike
- артрит. Предоставлено : Викисловарь. Расположен по адресу : en.wiktionary.org/wiki/arthritis . Лицензия : CC BY-SA: Attribution-ShareAlike
- Совместное. Предоставлено : Википедия. Расположен по адресу : en.Wikipedia.org/wiki/File:Joint.png . Лицензия : CC BY-SA: Attribution-ShareAlike
- Колледж OpenStax, Введение.17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44783/latest…_38_00_01f.jpg . Лицензия : CC BY: Attribution
- 1105 Передний и задний виды мышц. Предоставлено : Википедия. Расположен по адресу : en.Wikipedia.org/wiki/File:11…of_Muscles.jpg . Лицензия : CC BY: Attribution
- Скелет человека спереди en. Предоставлено : Википедия. Расположен по адресу : en.Wikipedia.org/wiki/File:Hu…n_front_en.svg . Лицензия : Общественное достояние: неизвестно Авторские права
- Колледж OpenStax, Биология. 17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44781/latest…ol11448/latest . Лицензия : CC BY: Attribution
- Колледж OpenStax, Биология.23 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44781/latest…ol11448/latest . Лицензия : CC BY: Attribution
- экзоскелет. Предоставлено : Викисловарь. Расположен по адресу : en.wiktionary.org/wiki/exoskeleton . Лицензия : CC BY-SA: Attribution-ShareAlike
- целом. Предоставлено : Викисловарь. Расположен по адресу : en.wiktionary.org/wiki/coelom . Лицензия : CC BY-SA: Attribution-ShareAlike
- перистальтика. Предоставлено : Викисловарь. Находится по адресу : en.wiktionary.org/wiki/peristalsis . Лицензия : CC BY-SA: Attribution-ShareAlike
- эндоскелет. Предоставлено : Викисловарь. Расположен по адресу : en.wiktionary.org/wiki/endoskeleton . Лицензия : CC BY-SA: Attribution-ShareAlike
- Совместное. Предоставлено : Википедия. Расположен по адресу : en.Wikipedia.org/wiki/File:Joint.png . Лицензия : CC BY-SA: Attribution-ShareAlike
- Колледж OpenStax, Введение.17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44783/latest…_38_00_01f.jpg . Лицензия : CC BY: Attribution
- 1105 Передний и задний виды мышц. Предоставлено : Википедия. Расположен по адресу : en.Wikipedia.org/wiki/File:11…of_Muscles.jpg . Лицензия : CC BY: Attribution
- Скелет человека спереди en. Предоставлено : Википедия. Расположен по адресу : en.Wikipedia.org/wiki/File:Hu…n_front_en.svg . Лицензия : Общественное достояние: неизвестно Авторские права
- Колледж OpenStax, Типы скелетных систем. 17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44781/latest…e_38_01_02.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Типы скелетных систем.17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44781/latest…e_38_01_01.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Типы скелетных систем. 17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44781/latest…e_38_01_03.jpg . Лицензия : CC BY: Attribution
- позвоночник. Предоставлено : Викисловарь. Расположен по адресу : en.wiktionary.org/wiki/vertebral_column . Лицензия : CC BY-SA: Attribution-ShareAlike
- межпозвоночный диск. Предоставлено : Викисловарь. Расположен по адресу : en.wiktionary.org/wiki/intervertebral_disc . Лицензия : CC BY-SA: Attribution-ShareAlike
- Колледж OpenStax, Биология.17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44781/latest…ol11448/latest . Лицензия : CC BY: Attribution
- выпуклый. Предоставлено : Викисловарь. Расположен по адресу : en.wiktionary.org/wiki/convex . Лицензия : CC BY-SA: Attribution-ShareAlike
- косточка. Предоставлено : Викисловарь. Расположен по адресу : en.wiktionary.org/wiki/ossicle . Лицензия : CC BY-SA: Attribution-ShareAlike
- Безграничный. Предоставлено : Безграничное обучение. Расположен по адресу : www.boundless.com//biology/definition/concave . Лицензия : CC BY-SA: Attribution-ShareAlike
- Совместное. Предоставлено : Википедия. Расположен по адресу : en.Wikipedia.org/wiki/File:Joint.png . Лицензия : CC BY-SA: Attribution-ShareAlike
- Колледж OpenStax, Введение. 17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44783/latest…_38_00_01f.jpg . Лицензия : CC BY: Attribution
- 1105 Передний и задний виды мышц. Предоставлено : Википедия. Расположен по адресу : en.Wikipedia.org/wiki/File:1105_Anterior_and_Posterior_Views_of_Muscles.jpg . Лицензия : CC BY: Attribution
- Скелет человека спереди en. Предоставлено : Википедия. Расположен по адресу : en.Wikipedia.org/wiki/File:Human_skeleton_front_en.svg . Лицензия : Общественное достояние: неизвестно Авторские права
- Колледж OpenStax, Типы скелетных систем.17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44781/latest…e_38_01_02.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Типы скелетных систем. 17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44781/latest…e_38_01_01.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Типы скелетных систем.17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44781/latest…e_38_01_03.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Типы скелетных систем. 17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44781/latest…e_38_01_07.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Типы скелетных систем.17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44781/latest…e_38_01_08.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Типы скелетных систем. 17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44781/latest…e_38_01_06.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Типы скелетных систем.17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44781/latest…e_38_01_04.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Типы скелетных систем. 17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44781/latest…e_38_01_05.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Биология.17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44781/latest…ol11448/latest . Лицензия : CC BY: Attribution
- осевой каркас. Предоставлено : Википедия. Расположен по адресу : en.Wikipedia.org/wiki/axial%20skeleton . Лицензия : CC BY-SA: Attribution-ShareAlike
- ключица. Предоставлено : Викисловарь. Расположен по адресу : en.wiktionary.org/wiki/clavicle . Лицензия : CC BY-SA: Attribution-ShareAlike
- лопатка. Предоставлено : Викисловарь. Расположен по адресу : en.wiktionary.org/wiki/scapula . Лицензия : CC BY-SA: Attribution-ShareAlike
- аппендикуляр. Предоставлено : Викисловарь. Расположен по адресу : en.wiktionary.org/wiki/appendicular . Лицензия : CC BY-SA: Attribution-ShareAlike
- членораздельный. Предоставлено : Викисловарь. Находится по адресу : en.wiktionary.org/wiki/articulate . Лицензия : CC BY-SA: Attribution-ShareAlike
- Совместное. Предоставлено : Википедия. Расположен по адресу : en.Wikipedia.org/wiki/File:Joint.png . Лицензия : CC BY-SA: Attribution-ShareAlike
- Колледж OpenStax, Введение. 17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44783/latest…_38_00_01f.jpg . Лицензия : CC BY: Attribution
- 1105 Передний и задний виды мышц. Предоставлено : Википедия. Расположен по адресу : en.Wikipedia.org/wiki/File:1105_Anterior_and_Posterior_Views_of_Muscles.jpg . Лицензия : CC BY: Attribution
- Скелет человека спереди en. Предоставлено : Википедия. Расположен по адресу : en.Wikipedia.org/wiki/File:Human_skeleton_front_en.svg . Лицензия : Общественное достояние: неизвестно Авторские права
- Колледж OpenStax, Типы скелетных систем.17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44781/latest…e_38_01_02.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Типы скелетных систем. 17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44781/latest…e_38_01_01.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Типы скелетных систем.17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44781/latest…e_38_01_03.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Типы скелетных систем. 17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44781/latest…e_38_01_07.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Типы скелетных систем.17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44781/latest…e_38_01_08.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Типы скелетных систем. 17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44781/latest…e_38_01_06.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Типы скелетных систем.17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44781/latest…e_38_01_04.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Типы скелетных систем. 17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44781/latest…e_38_01_05.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Типы скелетных систем.17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44781/latest…e_38_01_09.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Типы скелетных систем. 17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44781/latest…e_38_01_10.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Типы скелетных систем.17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44781/latest…e_38_01_11.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Типы скелетных систем. 17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44781/latest…e_38_01_13.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Типы скелетных систем.17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44781/latest…e_38_01_14.jpg . Лицензия : CC BY: Attribution
Метрические характеристики костей конечностей человека в азиатской и японской популяциях
Реферат
Двенадцать метрических переменных плечевой кости, лучевой кости, бедренной кости и голени были исследованы в 11 выборках мужчин из северо-восточной и восточной Азии.Были рассчитаны вариации между регионами и корреляции между широтой и соответствующими измерениями и индексами, и был проведен анализ главных компонентов для выяснения характеристик костей конечностей человека. Значимая корреляция и предельно значимая корреляция были обнаружены для максимального подквертельного диаметра ( r = 0,662, P = 0,027) и платимерного индекса ( r = -0,583, P = 0,060) с широтой соответственно, что позволяет предположить что бедренная кость у северных азиатов имела широкую и плоскую субвертельную форму.Второй главный компонент анализа главных компонентов показывает, что северо-восточные образцы со сравнительно большой длиной вала и тонкими и плоскими диаметрами вала отличались от южных образцов; второй главный компонент значимо коррелировал с широтой ( r = -0,743, P = 0,009). Расчетное значение F st , равное 0,432–0,336, показывает, что разброс в измерениях костей конечностей по регионам довольно велик, примерно в два или три раза превышающий низкие уровни межрегиональных отклонений (0.078–0.180) при анализе черепных и стоматологических данных. Неоднократно предлагалось, чтобы на морфологию костей конечностей сильнее влияли факторы окружающей среды и питания, чем черты черепа и зубов, но это исследование является первым, подтверждающим это на основе статистического анализа.
Введение
Эволюционные и филогенетические изменения в скелетах конечностей гоминидов, особенно изменения между неандертальцами и современными людьми, исследовались в течение длительного времени (Trinkaus, 1981; Ruff, 2002; Weaver and Steudel-Numbers, 2005; Young et al., 2010). До 1990-х годов морфологические различия, в том числе в пропорциях тела, между неандертальцами и анатомически современными людьми ( Homo sapiens ) объяснялись как одно из свидетельств модели замещения в Европе (Holliday, 1997a, b).
Географические вариации черепных и посткраниальных форм человека были исследованы как отражающие генетическое разнообразие в связи с всемирной миграцией анатомически современных людей из Африки (Relethford, 1994, 2002; Jorde et al., 2000; Холлидей и Рафф, 2001; Manica et al., 2007). Черты черепа и зубов обычно демонстрируют примерно 10–15% общей вариабельности между регионами и поэтому считаются нейтральными генетическими маркерами (Relethford, 2002; Hanihara, 2008).
С другой стороны, считается, что морфология костей конечностей также имеет существенную связь с климатическими, пищевыми и другими условиями окружающей среды (Trinkaus, 1981; Ruff, 1994, 2002). Например, хорошо известно, что люди, живущие в холодном климате, как правило, имеют низкие значения тибиофеморального (голеностопного) индекса (длина большеберцовой кости относительно длины бедра), в то время как для людей в теплом климате верно обратное.
В Японии за последние 100 лет было получено много базовой информации о показателях костей конечностей у людей дзёмон и других доисторических и исторических популяций (например, Коганей, 1893; Кийоно и Хираи, 1928a, b). Во-первых, мы хотели бы дать краткий обзор истории населения Японии. Люди дзёмон были доисторическими жителями Японских островов, которые жили в период с 10 000 до 2300 лет назад (Hanihara, 1991; Habu, 2004). Считалось, что люди культурного периода Яёи, которые жили в северных районах Японии Кюсю и Ямагути с 300 г. до н.э. до 300 г. н.э., находились под значительным влиянием иммигрантов с материковой Азии или их потомков.Основная концепция модели «двойной» структуры для истории японского населения, или теории общего происхождения айну и рюкю, заключается в следующем (Hanihara, 1991): генетический состав современного материкового японца в основном происходит от народа яёй, тогда как и айны в северной части, и рюкюаны на самых южных островах Японии являются потомками народа дзёмон. Недавние генетические исследования, основанные на значительных данных по однонуклеотидному полиморфизму, подтвердили теорию общего происхождения айнов и Рюкю (Koganebuchi et al., 2012; Консорциум генетики населения Японии на архипелаге, 2012 г.).
Ямагути (1982) показал, что и у дзёмон, и у айнов были относительно высокие значения радиоплечевого и тибиофеморального индекса, тогда как у людей яёй и современных японцев с материковой части эти значения были низкими, что подтверждает модель двойной структуры. С другой стороны, у айнов было высокое значение индекса гиперкнемии бедренной кости или плоская субвертельная форма, в то время как у народа Дзёмон вместо этого было высокое значение индекса пилястра (Yamaguchi, 1982).
Примерно за 20 лет до своей публикации 1982 года Ямагути также исследовал морфологию костей конечностей людей из Охотского района (Mitsuhashi and Yamaguchi, 1962a, b), чья культура распространилась от южного острова Сахалин до северо-восточного острова Хоккайдо и Курильских островов с 5-го января. до 12 века нашей эры (Амано, 2003). Охотская культура сформировала значительную морскую инфраструктуру, которая отличалась от инфраструктуры коренного населения Хоккайдо (Hudson, 2004; Naito et al., 2010; Shimoda et al., 2012). Древний ДНК и морфологический анализ выявили, что люди Охотска были тесно связаны с соседними с ними выходцами из северо-восточной Азии (Komesu et al., 2008; Sato et al., 2009, 2010; Kazuta et al., 2011). Подобно северо-восточным азиатам, у охотских людей значения тибиофеморального индекса были намного ниже, чем у других японских популяций (Mitsuhashi, Yamaguchi, 1962a, b).
В 1988–1989 годах последний автор (Х.И.) имел возможность исследовать морфологию костей конечностей северо-восточноазиатских групп из коллекции в России.Кроме того, у авторов была возможность исследовать около 260 останков скелетов Охотска в музее Университета Хоккайдо с 2003 года. Метрические и неметрические данные о черепных и зубных чертах в Охотском районе и северо-восточных азиатских образцах были опубликованы в других местах (Ishida, 1990, 1992, 1993 , 1994, 1995, 1996, 1997; Исида и Додо, 1996; Исида и Кондо, 1999). Недавно Хирофуми Мацумура из Медицинского университета Саппоро, один из авторов, сообщил о метрических характеристиках бедренной кости у жителей Охотска (Matsumura et al., 2010).
В данном исследовании мы хотели бы представить новые метрические данные для костей конечностей у доисторических неолитических байкальских, эквенских и охотских народов железного возраста, а также современных бурят. Используя измерения длинных костей образцов из Северо-Восточной Азии и других доступных образцов из Японии и Микронезии, мы исследовали региональные вариации и оценили корреляционные связи с широтой. Также был проведен анализ основных компонентов для выяснения характеристик костей конечностей человека.
Материалы и методы
В Таблице 1 перечислены 11 выборок населения, использованных в этом исследовании, с указанием географических широт и другой информацией, включая учреждения, в которых они проживают. На рисунке 1 показано расположение образцов.
Таблица 1 Кости конечностей азиатов и японцев, использованные в этом исследовании.| Размер выборки | Период | Широта | Информация | Учреждение | Источник | |
|---|---|---|---|---|---|---|
| Эквен | 65 | 2000 Б.П. | 66 ° с.ш. | Древнее эскимосское кладбище, Россия | МГУ | Настоящее исследование |
| Неолит Байкал | 38 | г.8000–4000 л.н. | 52 ° с.ш. | Восточное и западное побережье Байкала | MAE | Настоящее исследование |
| Буряты | 20 | Последние | 51 ° с.ш. | Восточное побережье Байкала | MAE | Настоящее исследование |
| Охотск | 79 | 400–1200 н.э. | 46 ° с.ш. | Сахалин, Ребун, Хоккайдо | ХУ, СМУ, КУ | Настоящее исследование, Охба (1934), Исида (1994), Мицухаси и Ямагути (1962a, b) |
| Айну | 47 | Раннее Новое время | 42 ° с.ш. | Хоккайдо | UT | Коганей (1893) |
| Современная материковая Япония | 29 | Последние | 35 ° с.ш. | Кинай | КУ | Миямото (1925), Хираи и Табата (1928) |
| Джомон Йошиго | 80 | 4000–2300 л.н. | 34 ° с.ш. | Айти | КУ | Охба (1935), Исидзава (1931) |
| Джомон Цукумо | 24 | 3000–2300 л. Н. | 34 ° с.ш. | Окаяма | КУ | Киёно и Хираи (1928a, b) |
| Яёи | 128 | 2300–1700 л.н. | 33 ° с.ш. | Северный Кюсю, Симанэ, Ямагути | KY | Кафедра анатомии Университета Кюсю (1988) |
| Житель острова Рюкю | 84 | 1600–1900 гг. | 26 ° с.ш. | Ранний современный Кумедзима, Окинава | OPAC | Fukumine et al.(2001) |
| Микронезия | 42 | Раннее Новое время | 13 ° с.ш. | Гуам | BM | Исида (1993) |
МГУ: МГУ, Москва, Россия.
МАЭ: Музей антропологии и этнографии, Санкт-Петербург, Россия.
HU: Университет Хоккайдо, Саппоро, Япония.
SMU: Медицинский университет Саппоро, Саппоро, Япония.
KU: Университет Киото, Киото, Япония.
UT: Токийский университет, Токио, Япония.
KY: Университет Кюсю, Фукуока, Япония.
OPAC: Археологический центр префектуры Окинава, Нишихара, Япония.
BM: Музей Б. П. Бишопа, Гонолулу, Гавайи, США.
фигура 1Расположение образцов, использованных в данном исследовании.
Скелетные материалы Эквена взяты с древнего кладбища арктических народов (Арутюнов, Сергеев, 1975). Образцы неолитического Байкала происходят как с восточного, так и с западного побережья озера Байкал. Недавняя бурятская серия была собрана на восточном побережье озера Байкал.Семь наборов скелетных материалов, состоящих из представителей племен Охотск, Хоккайдо, айнов, Дзёмон Ёсиго, Дзёмон Цукумо, современных материковых японцев, яёй и рюкю, происходят с Японских островов. Микронезийская серия представляет собой образец скелетов чаморро с Марианских островов.
Мы исследовали для сравнения только мужские образцы из-за отсутствия женской бурятской коллекции. Пол оценивали по морфологическому исследованию целых скелетов (White, 2000). В данном исследовании первый автор (М.К.) измерил охотские образцы, а последний автор (Х.I.) собраны данные по эквенским, неолитическим байкальским и бурятским образцам. Кости конечностей, состоящие из плечевой, лучевой, бедренной и большеберцовой костей (12 метрических переменных), были измерены в основном по методикам Мартина, перечисленным в Приложениях 1 и 2 (Knussmann, 1988). Однако для субвертельных диаметров бедренной кости мы использовали не сагиттальный и поперечный диаметры, а максимальный и минимальный (Koganei, 1893). Поперечные диаметры диафиза большеберцовой кости были взяты на месте питательного отверстия в соответствии с определением, данным Валлуа (Olivier, 1960).В принципе, мы измерили кости с правой стороны. Однако в случаях, когда максимальная длина кости на правой стороне была недоступна, для измерения вместо этого использовалась левая сторона. Другие данные по образцам были процитированы из литературы, перечисленной в Таблице 1. Однако сагиттальный и поперечный субвертельный диаметры бедренной кости были приняты в образцах Яёи и Рюкю. Поэтому для сопоставления сравнительных данных максимальный и минимальный диаметры были оценены с помощью множественных методов регрессии с использованием имеющихся сагиттальных и поперечных подвертельных диаметров.
Мы рассчитали базовую статистику для каждой выборки. Затем был проведен дисперсионный анализ для оценки групповых различий (Snedecor and Cochran, 1980). Когда между образцами были обнаружены существенные различия, был проведен тест Стьюдента t , чтобы определить, значительно ли отличается каждая группа от среднего значения всех других групп после поправки Бонферрони. Затем были рассчитаны коэффициенты ранговой корреляции Спирмена между измерениями и широтой для оценки связи между морфологией скелета конечностей и климатическими условиями.Анализ главных компонентов проводился с 12 переменными для характеристики структуры костей конечностей. Оценки Z отдельных наборов данных использовались для анализа, чтобы смягчить эффекты, связанные с размером (Howells, 1989; Brace and Hunt, 1990). Кроме того, коэффициенты ранговой корреляции Спирмена были вычислены между оценками основных компонентов и широтами.
Наконец, для сравнения дисперсии внутри и между географическими группами использовалась модификация статистики Райта F st , разработанная Релетфордом и Бланжеро (1990), Релетфордом (1994) и Релетфордом и Харпендингом (1994).
Результаты
Основные статистические данные измерений костей конечностей среди 11 выборок населения перечислены в Приложениях 1 и 2. Дисперсионный анализ показал, что все измерения и индексы значительно различаются между образцами. На рис. 2 показаны графики соответствующих измерений, в которых популяции расположены с севера (слева) на юг (справа). Среднее значение группы, отмеченной звездочкой (*), значительно выше, чем общее среднее значение всех других групп, в то время как знак решетки (#) указывает на значительно более низкое значение, чем другие, на основе соответствующих тестов t после коррекции Бонферрони.
фигура 2Сравнение размеров костей конечностей среди 11 групп. Звездочки (* P <0,05, ** P <0,01) указывают на более высокую значимость, чем общее среднее значение для всех других групп, в то время как решетки (# P <0,05, ## P <0,01) указывают на то, что более низкая значимость, чем у других, на основе соответствующих тестов t после поправки Бонферрони. Верхнее и нижнее поля прямоугольных диаграмм представляют 75-й и 25-й процентили соответственно.Медиана — это линия, разделяющая прямоугольник пополам. Байкал: неолитические байкальцы; Япония: современный континентальный японский язык; Цукумо: люди Дзёмон Цукумо; Ёсиго: люди Дзёмон Ёсиго; Рюкю: жители острова Рюкю.
Рисунок 2 (продолжение)Сравнение размеров костей конечностей среди 11 групп. Звездочки (* P <0,05, ** P <0,01) указывают на более высокую значимость, чем общее среднее значение для всех других групп, в то время как хеш-метки (# P <0,05, ## P <0.01) указывают на более низкую значимость, чем другие, на основании соответствующих тестов t после поправки Бонферрони. Верхнее и нижнее поля прямоугольных диаграмм представляют 75-й и 25-й процентили соответственно. Медиана — это линия, разделяющая прямоугольник пополам. Байкал: неолитические байкальцы; Япония: современный континентальный японский язык; Цукумо: люди Дзёмон Цукумо; Ёсиго: люди Дзёмон Ёсиго; Рюкю: жители острова Рюкю.
Рисунок 2 (продолжение)Сравнение размеров костей конечностей среди 11 групп.Звездочки (* P <0,05, ** P <0,01) указывают на более высокую значимость, чем общее среднее значение для всех других групп, в то время как решетки (# P <0,05, ## P <0,01) указывают на то, что более низкая значимость, чем у других, на основе соответствующих тестов t после поправки Бонферрони. Верхнее и нижнее поля прямоугольных диаграмм представляют 75-й и 25-й процентили соответственно. Медиана — это линия, разделяющая прямоугольник пополам. Байкал: неолитические байкальцы; Япония: современный континентальный японский язык; Цукумо: люди Дзёмон Цукумо; Ёсиго: люди Дзёмон Ёсиго; Рюкю: жители острова Рюкю.
По продольным размерам верхней конечности обитатели Японских островов, как правило, ниже ростом. Жители островов Рюкю демонстрируют исключительно малые значения плечевой и лучевой длин. Однако у охотских людей были довольно крупные кости верхних конечностей, как у неолитических байкальцев и бурят. Радиоплечевые индексы (R1: h2) у народа дзёмон оказались заметно высокими, превышая 80 («dolichokerisch» на немецком языке), как сообщалось ранее (Yamaguchi, 1989; Temple et al., 2008). Было обнаружено, что микронезийские образцы имеют длинную и толстую плечевую кость.
Микронезийские образцы также имели длинные кости нижних конечностей, в то время как нижние конечности жителей острова Рюкю были самыми короткими. Показатели большеберцовой кости (T1: F1) в популяциях дзёмон и айнов Хоккайдо были выше, превышая 82. У бурят только более плоская морфология средней части диафиза (F6: F7) бедра. С другой стороны, бедра неолитических байкальских и охотских людей имели плоскую суб-вертельную форму (F10 ′: F9 ′) из-за большого максимального суб-вертельного диаметра в целом, в то время как бедренная кость микронезийской серии оказалась более плоской. округлый.Что касается большеберцовой кости, то у неолитических народов Байкала, Хоккайдо айнов и Дзёмон были плоские стержни (T9a ′: T8a).
Затем мы вычислили коэффициенты корреляции между широтой и средними значениями измерений или показателей костей конечностей. Существенная корреляция была обнаружена только для максимального подквертельного диаметра ( r = 0,662, P = 0,027), а незначительно значимая корреляция была также обнаружена для платимерного индекса ( r = -0,583, P = 0.060), как показано на рисунке 3. Это указывало на то, что бедренная кость у северных азиатов имела тенденцию иметь широкую и плоскую субвертельную форму.
Рисунок 3Корреляция с широтой. Среди измерений и индексов значимая корреляция наблюдалась только для максимального подквертельного диаметра (F9 ‘) ( r = 0,662, P = 0,027), но незначительно значимая корреляция была обнаружена для платимерного индекса (F10′: F9 ‘ ) ( r = -0,583, P = 0,0597).
Результаты анализа главных компонентов 12 метрических переменных показаны в Таблице 2 и на Рисунке 4. На первый и второй главные компоненты приходилось 55,83% и 10,64% общей дисперсии, соответственно. Первый главный компонент можно интерпретировать как компонент размера, особенно с точки зрения длины. Второй главный компонент можно интерпретировать как относительный размер общих диаметров диафиза по сравнению с общей продольной длиной, максимальным подвертельным диаметром и поперечным диаметром диафиза бедренной кости.Образцы микронезийцев и бурят со сравнительно длинными конечностями показали более высокие баллы по первому главному компоненту, что привело к правой позиции на графике на Рисунке 4. Это сильно контрастирует с коротконогими островитянами Рюкю, показанными в крайнем левом положении. Вторая ось главных компонентов на графике также показывает, что северо-восточные образцы со сравнительно длинным, тонким и плоским валом примерно расположены вокруг нижней области, тогда как южные образцы расположены в верхней области.Только второй главный компонент был достоверно коррелирован с широтой (рис. 5), а коэффициент корреляции ( r = -0,743, P = 0,009) был больше, чем у двух бедренных переменных.
Таблица 2 Анализ основных компонентов при измерениях костей конечностей| PC1 | PC2 | PC3 | PC4 | |
|---|---|---|---|---|
| h2. Максимальная длина плечевой кости | 0,783 | −0.403 | -0,190 | -0,208 |
| H5. Максимальный средний диаметр стержня плечевой кости | 0,730 | 0,398 | 0,161 | 0,023 |
| H6. Минимальный средний диаметр стержня плечевой кости | 0,710 | 0,398 | 0,091 | -0,206 |
| R1. Радиальная максимальная длина | 0,750 | -0,345 | -0,249 | -0,049 |
| F1.Максимальная длина бедра | 0,833 | -0,252 | -0,329 | -0,030 |
| F6. Диаметр среднего стержня бедренной кости сагиттальный | 0,737 | 0,172 | -0,092 | 0,596 |
| F7. Диаметр среднего стержня бедренной кости поперечный | 0,701 | -0,153 | 0,586 | -0,072 |
| F9 ′. Бедренный максимальный подвертельный диаметр | 0,649 | −0.294 | 0,595 | -0,103 |
| F10 ′. Бедренный минимальный подвертельный диаметр | 0,820 | 0,145 | 0,124 | 0,248 |
| T1. Латеральный мыщелок большеберцовой кости-лодыжка длина | 0,830 | -0,318 | -0,225 | 0,053 |
| T8a. Сагиттальный диаметр большеберцовой кости у питательного отверстия | 0,790 | 0,270 | -0,087 | 0.014 |
| T9a ′. Поперечный диаметр большеберцовой кости в области питательного отверстия | 0,595 | 0,530 | -0,247 | -0,381 |
| Собственное значение | 6,700 | 1,277 | 1,081 | 0,669 |
| % VAR | 55,83% | 10,64% | 9,01% | 5,58% |
Анализ главных компонентов проводился с помощью корреляционной матрицы, полученной из стандартизованных ( Z -баллов) значений.
Рисунок 4Результаты анализа главных компонентов на основе 12 метрических переменных. Вверху: двумерные отношения между первым и вторым основными компонентами. Внизу: собственные векторы в первой и второй главных компонентах. Сокращения приведены на Рисунке 2 и в Приложениях 1 и 2. Сплошной стержень: диаметр вала; открытый стержень: длина вала.
Рисунок 5.Значительная корреляция была обнаружена для второй главной компоненты с широтой ( r = −0.743, P = 0,009).
Предполагая, что выборки, использованные в этом исследовании, имели одинаковый эффективный размер популяции, межрегиональные различия были оценены с использованием значений F st . В таблице 3 показаны расчетные значения F st ( h 2 = 0,4–0,6) для измерений костей конечностей и другие значения F st . Результат оценки F st , равный 0,432–0,336, показывает, что разброс размеров костей конечностей по регионам довольно велик, примерно в два или три раза превышающий низкие уровни межрегиональных вариаций (0.078–0.180) при анализе черепных и стоматологических данных (Haneji et al., 2007; Toma et al., 2007; Komesu et al., 2008; Ishida et al., 2009).
Таблица 3 Сравнение значений F st , полученных на основе метрических и неметрических признаков скелета| Данные | ф ул | Образец | Каталожный номер |
|---|---|---|---|
| Размеры костей конечностей | 0,336–0,432 | Северо-Восточная Азия, Восточная Азия, Микронезия | Настоящее исследование, Наследственность 0.4–0,6 |
| Краниометрия | 0,153 | Северо-Восточная Азия, Восточная Азия | Ishida et al. (2009), Наследственность 0,55 |
| Неметрические черты черепа | 0,078 | Северо-Восточная Азия, Восточная Азия, Восточная Европа | Komesu et al. (2008), Наследственность 0,50 |
| Стоматологические измерения | 0,180 | Северо-Восточная Азия, Восточная Азия, Юго-Восточная Азия, острова Тихого океана | Toma et al.(2007), Наследственность 0,55 |
| Неметрическая дентальная вариация | 0,163 | Восточная Азия, Западная Азия | Haneji et al. (2007), Наследственность 0,55 |
Обсуждение
Подвертельная формаВ этом исследовании мы обнаружили новую корреляцию между субвертельной формой бедренной кости (особенно F9 ‘) и широтой использованных образцов. Это означает, что население, живущее в более северных районах, имеет более плоские субвертельные бедра.Однако вызваны ли эти различия генетическими или эпигенетическими (т. Е. Средовыми) факторами или обоими?
Люди восточноазиатского происхождения обладают более плоской субвертельной формой бедренной кости, чем европейцы и африканцы (Gill, 2001; Wescott, 2005). Это исследование также показало, что азиаты, кроме микронезийцев, имеют более низкие значения платимерного индекса (ниже 80), что подтверждает предыдущие результаты и предполагает, что некоторая популяционная разница (или генетический вклад) связана с субвертельной формой.
Недавно Кестерке (2008) сообщил, что предполагаемая наследуемость ( h 2 ) субвертельной формы бедренной кости составляла 0,47 ± 0,24 в образцах Papio hamadryas , что указывает на почти равный вклад генетических и негенетических факторов. к общей дисперсии.
В этом свете более плоская субвертельная форма у народов Северо-Восточной Азии также может быть объяснена не только генетическими, но и негенетическими факторами. Результаты анализа главных компонентов показали, что у северных народов был относительно длинный ствол с плоской подвертельной областью (рис. 5).Предполагается, что у северных народов было большое туловище в целом в соответствии с экологическими правилами (Ruff, 2002). Это могло вызвать большую срединно-латеральную изгибающую силу на диафизе верхней бедренной кости (Ruff, 2002), что, возможно, привело бы к сравнительно плоской подквертельной диафизе. Однако, поскольку в этом исследовании не измерялись ширина двубрюшной кости и диаметр головки бедренной кости, мы не можем оценивать эту гипотезу в дальнейшем.
У народов Северной Азии, как правило, был более низкий индекс платикнемии (T9a ‘: T8a) или более плоский диафиз большеберцовой кости, хотя не было значительной корреляции между индексом платикнемии и широтой ( r = -0.464, P = 0,150). Поскольку предполагается, что устойчивость диафиза соответствует наземной подвижности (Stock, 2006), плоский стержень бедренной и большеберцовой кости может отражать это.
Результаты этого исследования могут также внести свой вклад в давнюю проблему, заключающуюся в том, что у айнов на Хоккайдо была плоская субвертельная форма бедренной кости (74,2), в то время как у людей Дзёмон на Хонсю (76,1–78,9) была менее плоская субвертельная форма (Ямагути). , 1982). Кроме того, мы ранее исследовали индексы диафиза верхней бедренной кости среди популяций дзёмон из Хоккайдо и Тохоку, север Японии.Результаты показали, что платимерный индекс составлял 74,8 у людей Хоккайдо Дзёмон (Исида и др., 1987, 1992) и 80,8–80,9 у людей Тохоку Дзёмон (Ямагути, 1983; Баба, 1988). Это региональное различие повышает вероятность того, что плоская субвертельная форма бедренной кости у жителей Хоккайдо айнов отражает сохранение черты народа Хоккайдо Дзёмон.
Пропорция внутри конечностейКак упоминалось выше, индекс голени обычно выше в районах с холодным климатом или на высоких широтах (Holliday and Hilton, 2010).Хотя дисперсионный анализ выявил значительную разницу между образцами, сравниваемыми в этом исследовании, индекс голени (T1: F1) существенно не коррелировал с широтой, отчасти потому, что в это исследование были включены народы дзёмон и айны (рис. 2c). У народов Дзёмон и Айну очень высокие индексы бедра среди жителей Восточной Азии, тогда как такие индексы низки у людей яёй, современных континентальных японцев и жителей островов Рюкю с тех же островов Японии (Yamaguchi, 1989; Temple et al., 2008; Fukase et al., 2012). Однако даже когда ряд Дзёмон был исключен из анализа, корреляция не была значимой.
Дисперсионный анализ показал значительную разницу в плечевом индексе (R1: h2) между группами, но также не показал корреляции с широтой (рис. 2а). Как уже сообщал Ямагути (1982), люди дзёмон имеют исключительно большой радиус относительно плечевой кости (81,1–81,5). В то время как северо-восточные народы имеют более низкие плечевые индексы, у современных континентальных японцев и островитян Рюкю также низкие по сравнению с другими группами, например, бедренный индекс.
Причины, по которым эти два индекса не коррелируют с широтой в данном исследовании, могут быть следующими. Во-первых, сложная история популяции на Японских островах, а именно «модель двойной структуры» (Hanihara, 1991), возможно, ослабила корреляцию между пропорциями внутри лимба и широтой. То есть пропорции интралимбов у японских народов были осложнены недавней миграцией (около 2000 г. до н.э.) народа яёй (Yamaguchi, 1982; Temple et al., 2008). Вторая причина — это особенность народа дзёмон.Хотя это только одна часть «модели двойной структуры», она очень важна. Гипотеза южного или северного происхождения народа дзёмон обсуждалась на основе морфологических и генетических исследований (Turner, 1987, 1989, 1990; Matsumura, Hudson, 2005; Cavalli-Sforza et al., 1988; Kozintsev, 1990; Omoto). и Saitou, 1997; Ishida and Kondo, 1999; Bannai et al., 1999; Tokunaga et al., 2001; Tajima et al., 2004; Tanaka et al., 2004; Fukumine et al., 2006; Hammer et al. , 2006; Комесу и др., 2008; Ханихара и Исида, 2009 г .; Adachi et al., 2011), но этот вопрос остается нерешенным. Однако, поскольку длинные дистальные сегменты в пропорциях интралимбов у народа Дзёмон стабильны от северного Хоккайдо до южных островов Рюкю (Fukase et al., 2012), стабильность может уменьшаться с увеличением склона между севером и югом в пропорциях внутри лимба в этом регионе. изучение. В качестве последней причины недавние жители островов Рюкю показывают очень низкие плечевые и голеностопные индексы (76,0 и 79,9 соответственно), которые сопоставимы с таковыми у жителей Северо-Восточной Азии.Поскольку у них очень короткие кости конечностей, это явление можно частично объяснить с помощью аллометрии (Holliday and Ruff, 2001; Sylvester et al., 2008; Auerbach and Sylvester, 2011).
Ф улРасчетное значение F st , равное 0,432–0,336, показывает, что разброс размеров костей конечностей по регионам довольно велик. Это показывает, что параметры измерения костей конечностей не являются нейтральными и могут быть выбраны до или после рождения. Это контрастирует с чертами черепа и зубов, которые демонстрируют низкие межрегиональные различия (0.078–0.180) и рассматриваются как нейтральные генетические маркеры (Relethford, 1991, 1994, 2002; Relethford, Harpending, 1994, 1995; Relethford et al., 1997; Powell and Neves, 1999; Hanihara and Ishida, 2005; Hanihara, 2006). ; Haneji et al., 2007).
С другой стороны, Релетфорд (2002) сообщил, что цвет кожи очень сильно различается между регионами (0,88), что указывает на закономерность естественного отбора. Таким образом, межрегиональная вариация размеров костей конечностей (0,336–0,432) является промежуточной между вариацией цвета кожи (0.88) и краниодентальных признаков (0,078–0,180). Кроме того, поскольку уровень отражения кожи сильно коррелирует с широтой ( r 2 = 0,85, из Relethford, 1997), разумно, чтобы только второй главный компонент (таблица 2, рисунок 4) или субвертельная форма бедра может быть коррелирован с широтой в этом исследовании ( r 2 = 0,44–0,55). Неоднократно предлагалось, чтобы на морфологию костей конечностей сильнее влияли факторы окружающей среды и питания, чем черты черепа и зубов (Trinkaus, 1981; Ruff, 1994, 2002), но это исследование является первым, подтверждающим это на основе статистический анализ.
Благодарности
Мы хотим поблагодарить следующих людей за разрешение изучать находящиеся под их опекой материалы скелета: I.I. Гохмана и А.Г. Козинцева, Институт этнографии им. Петербургский филиал, г. Санкт-Петербург; покойный академик В. Алексеев, Институт археологии, Москва; и Т. Алексеева, МГУ, Москва, Россия. Б. Ямагути, Национальный музей природы и науки, Токио, любезно предоставил индивидуальные данные о костях конечностей из памятника Омисаки, Япония.
Это исследование было частично поддержано грантом на научные исследования (№ 22370087) Японского общества содействия науке. Мы глубоко признательны двум анонимным рецензентам за их ценные комментарии.
Ссылки
- Адачи, Н., Шинода, К., Умецу, К., Китано, Т., Мацумура, Х., Фудзияма, Р., Савада, Дж., И Танака, М. (2011) Митохондриальные Анализ ДНК скелетов Хоккайдо Дзёмон: остатки архаичных материнских линий на юго-западной окраине бывшей Берингии.Американский журнал физической антропологии, 146 : 346–360.
- Амано Т. (2003) Что такое охотская культура? В: Номура Т., Утагава Ю. (ред.), Эпи-Дзёмон, Охотская культура. Хоккайдо Синбунша, Саппоро, стр. 110–133 (на японском языке).
- Арутюнов С.А., Сергеев Д.А. (1975) Проблемы этнической истории Берингоморья: Эквенский могильник. Наука, Москва, с. 240 с.
- Ауэрбах, Б.М., Сильвестр А.Д. (2011) Аллометрия и очевидные парадоксы пропорций конечностей человека: влияние на коэффициенты масштабирования. Американский журнал физической антропологии, 144 : 382–391.
- Баба Х. (1988) Кость конечности. В: Ракушечник Санганджи. Отчет музея префектуры Фукусима 17. Фукусима, стр. 443–480 (на японском языке).
- Bannai, M., Ohashi, J., Harihara, S., Takahashi, Y., Juji, T., Omoto, K., and Tokunaga, K. (1999) Анализ гаплотипов генов HLA у айнов ( из Хоккайдо, северная Япония) поддерживает предположение, что они происходят от верхнепалеолитических популяций Восточной Азии.Тканевые антигены, 55 : 128–139.
- Брейс, К.Л., и Хант, К.Д. (1990) Нерасовый черепно-лицевой взгляд на человеческие вариации: от A (Австралия) до Z (унив). Американский журнал физической антропологии, 82 : 341–360.
- Кавалли-Сфорца, Л.Л., Пьяцца, А., Меноцци, П., и Маунтин, Дж. (1988) Реконструкция эволюции человека: объединение генетических, археологических и лингвистических данных. Труды Национальной академии наук США, 85 : 6002–6006.
- Кафедра анатомии Университета Кюсю (1988) Генезис японского населения и культуры. Рокко-Шуппан, Токио (на японском языке).
- Фукасе, Х., Вакебе, Т., Цурумото, Т., Сайки, К., Фудзита, М., и Исида, Х. (2012) Географические изменения формы тела доисторических самцов дзёмон на Японском архипелаге: экогеографические последствия. Американский журнал физической антропологии, 149 : 125–135.
- Фукумине, Т., Дои, Н., Исида, Х., Зукеран, К., Сэнсуи, С., Сасо, А., и Хига, Т. (2001) Человеческие скелеты из могил Яччи-но-Гама и Канджинбару. В: Археологический центр префектуры Окинава (ред.), Яччи-но-Гама, могилы Кандзинбару. Отчет об исследовании археологического центра префектуры Окинава, № 6, Окинава, стр. 345–385 (на японском языке).
- Фукумине, Т., Ханихара, Т., Нишиме, А., и Исида, Х. (2006) Неметрические черепные вариации ранних современных человеческих скелетных останков из Кумедзима, Окинава и населения островов Рюкю.Антропологическая наука, 114 : 141–151.
- Gill, G.W. (2001) Расовые вариации проксимального и дистального отдела бедренной кости: наследуемость и судебно-медицинская ценность. Journal of Forensic Sciences, 46 : 791–799.
- Хабу, Дж. (2004) Древний Дзёмон Японии. Издательство Кембриджского университета, Беркли, Калифорния.
- Молот, М.Ф., Карафет, Т.М., Парек, Х., Омото, К., Харихара, С., Стоункинг, М., и Хораи, С. (2006) Двойное происхождение японцев: общая основа для охотников-собирателей и фермерские Y-хромосомы.Журнал генетики человека, 51 : 47–58.
- Ханеджи, К., Ханихара, Т., Сунакава, Х., Тома, Т., и Исида, Х. (2007) Неметрические стоматологические вариации на островах Сакисима, Окинава, Япония: сравнительное исследование Сакисимы и других соседние популяции. Антропологическая наука, 115 : 35–45.
- Ханихара, К. (1991) Модель двойной структуры для истории населения Японии. Japan Review, 2 : 1–33.
- Ханихара, Т. (2006) Интерпретация черепно-лицевых изменений и диверсификация жителей Восточной и Юго-Восточной Азии.В: Oxenham, M. и Tayles, N. (ред.), Биоархеология Юго-Восточной Азии. Издательство Кембриджского университета, Кембридж, стр. 91–111.
- Ханихара, Т. (2008) Морфологическая изменчивость основных человеческих популяций на основе неметрических стоматологических черт. Американский журнал физической антропологии, 136 : 169–182.
- Ханихара, Т., и Исида, Х. (2005) Метрические стоматологические вариации основных популяций человека. Американский журнал физической антропологии, 128 : 287–298.
- Ханихара, Т., и Ishida, H. (2009) Региональные различия в черепно-лицевом разнообразии и истории населения Jomon Japan. Американский журнал физической антропологии, 139 : 311–322.
- Хираи Т. и Табата Т. (1928) Антропологические исследования скелета современных японцев. IV. Часть. Нижняя конечность. №1. О бедренной кости, надколеннике, голени и малоберцовой кости. Журнал антропологического общества Японии, 43 (1-е приложение): 1–82 (на японском языке).
- Холлидей, Т.W. (1997a) Пропорции тела в Европе позднего плейстоцена и современное происхождение человека. Журнал эволюции человека, 32 : 423–448.
- Холлидей, T.W. (1997b) Посткраниальные доказательства адаптации к холоду неандертальцев Европы. Американский журнал физической антропологии, 104 : 245–258.
- Холлидей, Т.В., и Хилтон, С.Е. (2010) Пропорции тела циркумполярных народов, о чем свидетельствуют данные скелетов: Ипютак и Тигара (Пойнт-Хоуп) в сравнении с инуитами на острове Кадьяк. Американский журнал физической антропологии, 142 : 287–302.
- Холлидей, Т.В., и Рафф, С.Б. (2001) Относительные вариации в длине проксимальных и дистальных сегментов конечностей человека. Американский журнал физической антропологии, 116 : 26–33.
- Howells, W.W. (1989) Формы черепа и карта. Краниометрические анализы в дисперсии современного человека. Документы Музея археологии и этнологии Пибоди, Vol. 79 , Музей Пибоди, Кембридж, Массачусетс, стр. 189.
- Хадсон, М.Дж. (2004) Перспективные реальности изменений: интеграция мировой системы и Охотская культура на Хоккайдо.Журнал антропологической археологии, 23 : 290–308.
- Исида, Х. (1990) Морфология черепа нескольких этнических групп из бассейна Амура и Сахалина. Журнал Антропологического общества Японии, 98 : 137–148.
- Исида, Х. (1992) Плоскостность лицевых скелетов у сибирских и других околотихоокеанских популяций. Zeitshrift für Morphologie und Anthropologie, 79 : 53–67.
- Исида, Х. (1993) Популяционное сходство перуанцев с сибиряками и североамериканцами: неметрический краниальный подход.Антропологическая наука, 101 : 47–63.
- Исида, Х. (1994) Морфология скелета охотников на острове Сахалин. Антропологическая наука, 102 : 257–269.
- Исида, Х. (1995) Неметрические черепные вариации популяций Северо-Восточной Азии и их популяционное сходство. Антропологическая наука, 103 : 385–401.
- Исида, Х. (1996) Метрические и неметрические черепные вариации доисторических охотников. Антропологическая наука, 104 : 233–258.
- Исида, Х. (1997) Краниометрическая изменчивость популяций Северо-Восточной Азии. Homo, 48 : 106–124.
- Исида, Х., Додо, Ю. (1996) Морфология черепа у сибиряков и жителей Восточной Азии. В: Akazawa, T., and Szathmary, E.J.E. (ред.), Доисторические монголоидные расселения. Oxford University Press, Oxford, pp. 113–124.
- Исида, Х., Кондо, О. (1999) Неметрические черепные вариации айнов и соседних популяций человека. Перспективы в биологии человека, 4 : 127–138.
- Исида, Х., Додо, Ю. и Ямагути, Б. (1987) Скелеты ракушечника Такасаго. Антропологическое исследование доисторических курганов на берегу вулканической бухты № 2. Кафедра анатомии (2), Медицинский колледж Саппоро, Саппоро, стр. 107–185.
- Исида, Х., Додо, Ю., Ханихара, Т., Охшима, Н., Кида, М., и Мацумура, Х. (1992) Остатки скелетов Дзёмон из поселения Котан-Онсэн, Якумо, Хоккайдо. В: Сайт Котан-Онсэн, Совет по образованию Якумо, Якумо, стр. 403–421.
- Исида, Х., Ханихара Т., Кондо О. и Фукумине Т. (2009) История краниометрической дивергенции японского населения. Антропологическая наука, 117 : 147–156.
- Исисава, М. (1931) Антропологические исследования скелета людей каменного века Ёсико. III. Часть. Нижняя конечность. На бедре, надколеннике, голени и малоберцовой кости. Журнал Антропологического общества Японии, 46 (1-е приложение): 1–192 (на японском языке).
- Консорциум по генетике населения Японии на японском архипелаге (2012 г.) История популяций людей на Японском архипелаге, выведенная на основе общегеномных данных SNP с особым упором на популяцию айнов и рюкюань.Журнал генетики человека, 57 : 787–795.
- Jorde, L.B., Watkins, W.S., Bamshad, M.J., Dixon, M.E., Ricker, C.E., and Seielstad, M.T. (2000) Распределение генетического разнообразия человека: сравнение митохондриальных, аутосомных данных и данных по Y-хромосоме. Американский журнал генетики человека, 66 : 979–988.
- Kazuta, H., Sato, T., Dodo, Y., Matsumura, H., Amano, T., Ishida, H., and Masuda, R. (2011) Частоты генотипов гена ABCC11 в 2000 г. –3000-летние человеческие кости из стоянок Эпи-Дзёмон и Дзёмон на Хоккайдо, Япония.Антропологическая наука, 119 : 81–86.
- Кестерке, М.Дж. (2008) Наследуемость субвертельной формы бедренной кости (платимерный индекс) в Papio hamadryas и ее значение для посткраниальной изменчивости и эволюции гоминидов и современного человека. Магистерская диссертация на кафедре антропологии Университета Вайоминга.
- Kiyono, K., and Hirai, T. (1928a) Anthropologische Untersuchungen über das skelett der Tsukumo-Steinzeitmenschen, III. Teil. Die Oberen Extremitaten. Журнал Антропологического общества Японии, 43 (Дополнение.): 179–285.
- Kiyono, K., and Hirai, T. (1928b) Anthropologische Untersuchungen über das skelett der Tsukumo-Steinzeitmenschen, IV. Хвост. Die Unteren Extremitaten. № 1. Uber die Femur, die Patella, die Tibia und die Fibula. Журнал Антропологического общества Японии, 43 (Дополнение): 303–390.
- Knussmann, R. (1988) Martin / Knussmann-Anthropologie: Handbuch der vergleichenden Biologie des Menschen, Bd. I. Густав Фишер Верлаг, Штутгарт.
- Коганебучи, К., Кацумура, Т., Накагоме, С., Исида, Х., Кавамура, С., и Оота, Х. Азиатский консорциум архивных репозиториев ДНК (2012) Аутосомные и Y-хромосомные STR-маркеры показывают тесную взаимосвязь между Хоккайдо айну и Островитяне Рюкю. Антропологическая наука, 120 : 199–208.
- Koganei, Y. (1893) Beiträge zur physischen Anthropologie der Aino, I. Untersuchungen am Skelet. Mittheilungen aus der Medicinischen Facultät der Kaiserlich-Japanischen Universität, Bd. 2. Kaiserlichen Universität, Токио (на немецком языке).
- Комесу, А., Ханихара, Т., Амано, Т., Оно, Х., Йонеда, М., Додо, Ю., Фукумин, Т., и Исида, Х. (2008) Неметрические черепные вариации у человека скелетные останки, связанные с охотской культурой. Антропологическая наука, 116 : 33–47.
- Козинцев А. (1990) Айны, японцы, их предки и соседи: краниоскопические данные. Журнал Антропологического общества Японии, 98 : 247–267.
- Manica, A., Amos, W., Balloux, F., and Hanihara, T. (2007) Влияние узких мест древних популяций на фенотипические вариации человека.Nature, 448 : 346–349.
- Мацумура, Х. и Хадсон, М. (2005) Стоматологические взгляды на историю населения Юго-Восточной Азии. Американский журнал физической антропологии, 127 : 182–209.
- Мацумура, Х., Хадсон, М.Дж., Кавамура, К., и Кашива, Т. (2010) Морфометрические особенности бедренных костей у жителей Охотска. Антропологическая наука (японская серия), 118 : 69–82.
- Мицухаси К. и Ямагути Б. (1962a) Антропологические исследования останков человека из Омисаки, Хоккайдо.II. Бедренная, большеберцовая и малоберцовая кости. Sapporo Medical Journal, 21 : 23–32 (на японском языке с резюме на английском языке).
- Мицухаси К. и Ямагути Б. (1962b) Антропологические исследования останков человека из Омисаки, Хоккайдо. IV. Плечевая, лучевая и локтевая кости. Sapporo Medical Journal, 22 : 288–294 (на японском языке с резюме на английском языке).
- Миямото, Х. (1925) Антропологические исследования скелета современных японцев. II. Верхняя конечность. Журнал антропологического общества Японии, 40 : 219–305 (на японском языке).
- Наито, Ю.И., Чикараиси, Ю., Окоучи, Н., Мукаи, Х., Шибата, Ю., Хонч, Н.В., Додо, Ю., Исида, Х., Амано, Т., Оно, Х., и Йонеда, М. (2010) Реконструкция питания охотской культуры на Хоккайдо, Япония, на основе изотопного состава азота в аминокислотах: значение для коррекции воздействия морских резервуаров радиоуглерода на кости человека. Радиоуглерод, 52 : 671–681.
- Охба, Х. (1934) О верхних конечностях человеческих костей из курганов Сусуя, Сахалин.Журнал Антропологического общества Японии, 49 : 389–401, 493–504. (на японском языке)
- Охба, Х. (1935) Антропологические исследования скелета людей каменного века Ёсико из пров. Микава, Япония. Часть IV. верхняя конечность. №1. Плечевой пояс. Журнал антропологического общества Японии, 50 (9-е приложение): 99–169 (на японском языке).
- Оливье, Г., (1960) Pratique anthropologieque. Vigot Freres, Париж.
- Омото, К., и Сайто, Н.(1997) Генетическое происхождение японцев: частичная поддержка гипотезы двойственной структуры. Американский журнал физической антропологии, 102 : 437–446.
- Пауэлл, Дж. Ф. и Невес, В. А. (1999) Черепно-лицевая морфология первых американцев: образец и процесс в заселении Нового Света. Ежегодник физической антропологии, 42 : 153–188.
- Relethford, J.H. (1991) Генетический дрейф и антропометрические вариации в Ирландии. Биология человека, 63 : 155–165.
- Relethford, J.H. (1994) Краниометрические вариации среди современных человеческих популяций. Американский журнал физической антропологии, 95 : 53–62.
- Relethford, J.H. (1997) Полушарные различия в цвете кожи человека. Американский журнал физической антропологии, 104 : 449–457.
- Relethford, J.H. (2002) Распределение глобального генетического разнообразия человека на основе краниометрии и цвета кожи. Американский журнал физической антропологии, 118 : 393–398.
- Релетфорд, Дж. Х. и Бланжеро, Дж. (1990) Обнаружение дифференциального потока генов по образцам количественной изменчивости. Биология человека, 62 : 5–25.
- Релетфорд, Дж. Х., и Харпендинг, Х. С. (1994) Краниометрические вариации, генетическая теория и современное происхождение человека. Американский журнал физической антропологии, 95 : 249–270.
- Релетфорд, Дж. Х., и Харпендинг, Х. С. (1995) Древние различия в численности населения могут имитировать недавнее африканское происхождение современного человека.Текущая антропология, 36 : 667–674.
- Relethford, J.H., Crawford, M.H., and Blangero, J. (1997) Генетический дрейф и поток генов в Ирландии после голода. Биология человека, 69 : 443–465.
- Рафф, К. Б. (1994) Морфологическая адаптация к климату современных и ископаемых гоминидов. Американский журнал физической антропологии, 37 : 65–107.
- Ruff, C.B. (2002) Изменение размера и формы человеческого тела. Ежегодный обзор антропологии, 31 : 211–232.
- Сато, Т., Амано, Т., Оно, Х., Исида, Х., Кодера, Х., Мацумура, Х., Йонеда, М., и Масуда, Р. (2009) Гаплогруппировка митохондриальной ДНК Охотцы на основе анализа древней ДНК: промежуточного звена генного потока от жителей континентального Сахалина к айнам. Антропологическая наука, 117 : 171–180.
- Сато, Т., Казута, Х., Амано, Т., Оно, Х., Исида, Х., Кодера, Х., Мацумура, Х., Йонеда, М., Додо, Ю. и Масуда, R. (2010) Полиморфизмы и частоты аллелей гена группы крови ABO среди народов Дзёмон, Эпи-Дзёмон и Охотска на Хоккайдо, северная Япония, выявленные с помощью анализа древней ДНК.Журнал генетики человека, 55 : 691–696.
- Симода, Ю., Нагаока, Т., Моромизато, К., Сунагава, М., Ханихара, Т., Йонеда, М., Хирата, К., Оно, Х., Амано, Т., Фукумине, Т. ., и Исида, Х. (2012) Дегенеративные изменения позвоночника у людей из Охотской культуры и двух древних групп из Канто и Окинавы, Япония. Антропологическая наука, 120 : 1–21.
- Снедекор, Г.В., Кокран, В.Г. (1980) Статистические методы, 7-е изд. Издательство государственного университета Айовы, Эймс.
- Сток, J.T. (2006) Посткраниальная устойчивость охотников-собирателей относительно моделей мобильности, адаптации к климату и отбора для экономии тканей. Американский журнал физической антропологии, 131 : 194–204.
- Сильвестр А.Д., Крамер П.А., Юнгерс В.Л. (2008) Современные люди не (совсем) изометричны. Американский журнал физической антропологии, 137 : 371–383.
- Таджима, А., Хаями, М., Джуджи, Т., Мацуо, М., Марзуки, С., Омото, К., и Хораи, С.(2004) Генетическое происхождение айнов установлено на основе комбинированного анализа ДНК материнской и отцовской линий. Журнал генетики человека, 49 : 187–193.
- Танака, М., Кабрера, В.М., Гонсалес, А.М., Ларруга, Д.М., Такеясу, Т., Фуку, Н., Ли-Джун, Г., Хиросе, Р., Фудзита, Ю., Курата, М. , Шинода, К., Умецу, К., Ямада, Ю., Осида, Ю., Сато, Ю., Хаттори, Н., Мизуно, Ю., Танака, Ю., Кавамори, Р., Шамото-Нагай, М., Маруяма В., Симоката Х., Судзуки Р. и Шимодаира Х. (2004) Вариации митохондриального генома в Восточной Азии и население Японии.Genome Research, 14 : 1832–1850.
- Temple, D.H., Auerbach, B.M., Nakatsukasa, M., Sciilli, P.W., and Larsen, C.S. (2008) Различия в пропорциях конечностей между собирателями дзёмон и земледельцами яёи из доисторической Японии. Американский журнал физической антропологии, 137 : 164–174.
- Tokunaga, K., Ohashi, J., Bannai, M., and Juji, T. (2001) Генетическая связь между азиатами и коренными американцами: данные по генам и гаплотипам HLA. Иммунология человека, 62 : 1001–1008.
- Тома Т., Ханихара Т., Сунакава Х., Ханеджи К. и Исида Х. (2007) Метрическое стоматологическое разнообразие жителей островов Рюкю: сравнительное исследование среди населения Рюкю и других азиатских народов. Антропологическая наука, 115 : 119–131.
- Тринкаус, Э. (1981) Пропорции конечностей неандертальца и адаптация к холоду. В: Стрингер К. (ред.), Аспекты эволюции человека. Тейлор и Фрэнсис, Лондон, стр. 187–224.
- Тернер, C.G.II (1987) История популяции Восточной Азии в позднем плейстоцене и голоцене на основе дентальной изменчивости.Американский журнал физической антропологии, 73 : 305–321.
- Тернер, C.G.II (1989) Зубы и предыстория в Азии. Scientific American, 260 : 70–77.
- Тернер, C.G.II (1990) Основные особенности Sundadonty и Sinodonty, включая предположения о восточноазиатской микроэволюции, истории популяции и позднеплейстоценовых отношениях с австралийскими аборигенами. Американский журнал физической антропологии, 82 : 295–317.
- Уивер, Т.Д., и Штудель-Числа, К.(2005) Объясняет ли климат или мобильность различия в пропорциях тела неандертальцев и их последователей в верхнем палеолите? Эволюционная антропология, 14 : 218–223.
- Уэскотт, Д.Дж. (2005) Популяционные различия в субвертельной форме бедренной кости. Journal of Forensic Sciences, 50 : 286–293.
- Уайт, Т. Д. (2000) Человеческая остеология, 2-е изд. Academic Press, Сан-Диего, Калифорния.
- Ямагути Б. (1982) Обзор остеологических характеристик популяции дзёмон в доисторической Японии.Журнал Антропологического общества Японии, 90 (Дополнение): 77–90.
- Ямагути, Б. (1983) Посткраниальные кости человеческих скелетных останков периода Дзёмон из кургана ракушек Эбисима (Кайтори) в Ханаидзуми, префектура Иватэ. Воспоминания Национального музея науки, 16 : 201–222.
- Ямагути Б. (1989) Пропорции сегментов конечностей в человеческих останках периода Дзёмон. Бюллетень Национального музея науки, Токио, сер. Д, 15 : 41–48.
- Янг Н.М., Вагнер Г.П. и Халлгримссон Б. (2010) Развитие и эволюция конечностей человека. Труды Национальной академии наук США, 107 : 3400–3405.
(PDF) Фундаментальные отношения и логарифмическая периодичность в костях конечностей человека
Buikstra JE, Ubelaker DH. (1994) Стандарты сбора данных
из останков человеческого скелета. Отчет № 44. Файетвилл:
Arkansas Archaeological Survey Press.
Byers S, Akoshima K, Curran B (1989) Определение роста взрослых
по длине плюсны.Am J Phys Anthropol 79,
275–279.
Dahn RD, Fallon JF (2000) Межпальцевое регулирование идентификации пальцев
и гомеотическое преобразование с помощью модулированного сигнала BMP-
ing. Science 289, 438–441.
Dekanty A, Milan M (2011) Взаимодействие между морфогенами
и ростом ткани. EMBO Rep 12, 1003–1010.
Доган А., Услу М., Айдынлиоглу А. и др. (2007) Морфометрическое исследование
плюсневых костей и фаланг человека. Клин Анат 20,
209–214.
Эшак Г.А., Ахмед Х.М., Абдель Гавад Э.А. (2011) Определение пола
нации по длине и объему костей рук с использованием компьютерной томографии multidetec-
: исследование на египетском народе.
J Forensic Leg Med 18, 246–252.
Гундерсон Х.Дж., Бендстен Т.Ф., Корбо Л. и др. (1988) Некоторые новые,
простых и эффективных стереологических методов и их использование в
патологических исследованиях и диагностике. APMIS 96, 379–394.
Hahn SB, Park HW, Park HJ и др.(2003) Удлинение нижней конечности —
у карликов Тернера. Yonsei Med J 44, 502–507.
Hamilton R, Dunsmuir RA (2002) Радиографическая оценка
относительной длины костей пальцев руки человека
. J Hand Surg Br 27, 546–548.
Hauser R, Smoli
nski J, Gos T (2005) Оценка роста по
на основе измерений бедренной кости. Forensic Sci Int 147,
185–190.
Хокман Д., Кретекос С.Дж., Мейсон М.К. и др.(2008) Вторая волна
экспрессии Sonic hedgehog во время развития
конечности летучей мыши. Proc Natl Acad Sci U S. A 105, 16982–16987.
Хатчисон А.Л., Хатчисон Р.Л. (2010) Фибоначчи, Литтлер и рука
: краткий обзор. Хэнд (Нью-Йорк) 5, 364–368.
Kyriakidis I, Papaioannidou P, Pantelidou V, et al. (2010) Цифра
Соотношениеи отношение к инфаркту миокарда у греческих мужчин и
женщин. Генд Мед 7, 628–636.
Ландис Дж. Р., Кох Г. Г. (1977) Измерение согласия наблюдателя —
для категориальных данных.Биометрия 33, 159–174.
Littler JW (1973) Об адаптивности руки человека (со ссылкой на
на равноугольную кривую). Рука 5, 187–191.
Lowrance EW, Latimer HB (1967) Вес и изменчивость компонентов позвоночника человека
. Анат Рек. 159,83–88.
Mall G, Hubig M, B €
uttner A, et al. (2001) Определение пола и оценка роста длинных костей руки
. Foren-
sic Sci Int 117,23–30.
Мэннинг Дж. Т., Скатт Д., Уилсон Дж. И др. (1998) Отношение длины 2-го пальца к
: показатель количества и концентрации сперматозоидов —
единиц тестостерона, лютеинизирующего гормона и эстрогена.
Hum Reprod 13, 300–304.
Meinhardt H (1992) Формирование паттернов в биологии: сравнение
моделей и экспериментов. Rep Prog Phys 55, 797–849.
Mu ~
noz JI, Li ~
nares-Iglesias M, Su
arez-Pe ~
naranda JM, et al.(2001)
Оценка роста по рентгенологически определенной длине
длины кости в выборке из испанской популяции. J Forensic Sci 46,
363–366.
Odaci E, Sahin B, Sonmez OF, et al. (2003) Быстрая оценка объема тела позвонка
: комбинация принципа Cavalieri
и изображений компьютерной томографии. Eur J Radiol 48,
316–326.
Ориоли IM, Castilla EE, Barbosa-Neto JG (1986) Preva-
коэффициентов лентности для скелетных дисплазий.J Med Genet 23,
328–332.
Парк А.Е., Фернандес Дж. Дж., Шмеддерс К. и др. (2003) Последовательность Фибоначчи
: Связь с человеческой рукой. J Hand Surg 28A,
157–160.
Pelin IC, Duyar I (2003) Оценка роста по длине большеберцовой кости: сравнение методов
. J Forensic Sci 48, 708–712.
Рахман А.А., Лофатананон А., Стюарт-Браун С. и др. (2011)
Рисунок руки указывает на риск рака простаты. Br J Cancer 104,
175–177.
Резник Д. (1994) Возрождение рациональной морфологии: очистить философию биологии структурализма. Acta Biotheor 42,
1–14.
Richardson MK, Oelschl €
ager HH (2002) Время, закономерности и гетерохрония:
ipper. Evol Dev 4, 435–444.
Ричардсон М.К., Джеффри Дж.Э., Табин С.Дж. (2004) Проксимодистальный пат-
граница конечности: выводы из эволюционной морфологии.
Evol Dev 6,1–5.
Рикеттс Р.М. (1982) Биологическое значение божественной части
и ряда Фибоначчи. Am J Orthod 81, 351–370.
Sears KE, Behringer RR, Rasweiler JJ IV и др. (2006) Разработка —
полета летучей мыши: морфологическая и молекулярная эволюция
пальцев крыльев летучей мыши. Proc Natl Acad Sci U S A 103, 6581–6586.
Тривелла Г.П., Бригадой Ф., Альдегери Р. (1996) Удлинение ноги в
карликах Тернера. J Bone Joint Surg Br 78, 290–293.
Ulmer H, Kelleher CC, Dunser MW (2009) Джордж Клуни, цветочная машина
, кардиолог, и фи, золотое сечение. BMJ
339, b4745.
Verheyden JM, Sun X (2008) Fgf / Gremlin ингибирующая обратная связь
Петлязапускает прекращение отрастания зачатка конечности. Nature 454,
638–641.
Язар Ф, Имре Н, Баттал Б и др. (2012) Есть ли связь
между дистальными параметрами бедренной кости и ее высотой и шириной
? Хирургическая радиология Анат 34, 125–132.
Чжу Дж., Чжан Ю.Т., Альбер М.С. и др. (2010) Формирование паттерна голых костей —
: основная регуляторная сеть различной геометрии
воспроизводит основные черты развития конечностей
и эволюции позвоночных. PLoS ONE 5, e10892.
© 2013 Анатомическое общество
Фундаментальные соотношения в костях конечностей человека, A. Pietak et al. 537
Костяные стены из человеческих конечностей и черепов, обнаруженные под церковью в Бельгии
Археологи недавно обнаружили стены, построенные из жуткого материала: человеческих костей, включая разбитые черепа, при раскопках на территории церкви в Генте, Бельгия.
К концу раскопок археологи обнаружили девять стен, построенных в основном из бедер и голеней взрослых людей. Промежуточные зоны были заполнены черепами, многие из которых были фрагментированы, по словам Рубена Уилларта, голландской компании Restoration & Archeology / Decoration, которая нашла стены во время раскопок перед строительством нового центра для посетителей в соборе.
Эти ужасные строения, вероятно, были работой людей, которые сотни лет назад расчистили старое кладбище, чтобы освободить место для новых тел или ремонта церкви, сказал археолог Яник де Гриз, сотрудник Ruben Willaert и проект раскопок. лидер.
Связанный: На фотографиях: Кладбище воинов железного века
«При расчистке кладбища скелеты нельзя просто выбросить», — сказал де Гриз Live Science в электронном письме. «Поскольку верующие верили в воскресение тела, кости считались самой важной частью».
Охрана человеческих останков была настолько важна, что иногда каменные дома «строились у стен городских кладбищ, чтобы хранить черепа и длинные кости в том, что называется склепом», — сказал де Гриз.
Более мелкие кости, такие как позвонки, ребра, руки и ноги, не проникали в стену. (Изображение предоставлено: Copyright Ruben Willaert bvba)Костяные стены были обнаружены на северной стороне собора Святого Бавона, ранее известного как церковь Святого Иоанна Крестителя или Святого Яна. Радиоуглеродное датирование костей предполагает, что они датируются во второй половине 15 века, но стены, вероятно, были построены позже, в 17 или начале 18 веков, сказал де Гриз.
Исторические документы подтверждают эти даты. Один источник отмечает, что кладбище церкви было очищено в первой половине 16 века и снова после 1784 года, когда кладбище перестало принимать новые тела.
Независимо от даты, эти стены — единственная в своем роде находка.
«У нас нет сравнения в Бельгии», — сказал де Гриз. Большинство исторических кладбищ представляют собой большие ямы или слои, заполненные рыхлыми человеческими костями. «Мы никогда не видели таких структур, как стены, которые намеренно построены из человеческих костей», — сказал де Гриз.
Кто бы ни строил стены, должно быть, спешил, потому что они не удосужились собрать мелкие или хрупкие кости, такие как позвонки, ребра или кости с рук или ног. Любопытно, что археологи также не обнаружили никаких костей рук.
«Стены состоят только из костей нижних конечностей», — сказал де Гриз. «В настоящий момент мы все еще изучаем, какая идея вызвала это. Это только практическая вещь (очень компактное нагромождение костей) или есть еще религиозное / духовное измерение?»
Хотя есть кости и взрослых мужчин, и женщин, детские кости, похоже, отсутствуют на стенах, что противоречит известной продолжительности жизни с того периода, когда дети часто умирали от болезней.
«Но [это] типично для костей из расчищенных гробниц», — говорится в заявлении Рубена Уилларта (переведено с голландского языка с помощью Google Translate). «Детские кости маленькие и хрупкие, поэтому их не собирали».
На данный момент кости будут доставлены в новый дом в Гентском университете, где они будут изучены в рамках подробной инвентаризации, сказал де Гриз.
Первоначально опубликовано на Live Science .
ПРЕДЛОЖЕНИЕ: Сэкономьте минимум 53% с нашей последней скидкой на журнал!
Благодаря впечатляющим вырезанным иллюстрациям, показывающим, как все устроено, и умопомрачительным фотографиям самых вдохновляющих зрелищ в мире, How It Works представляет собой вершину увлекательного, фактического развлечения для основной аудитории, стремящейся не отставать от новейших технологий и самых впечатляющих явлений в мире. планета и за ее пределами.Написанный и представленный в стиле, который делает даже самые сложные предметы интересными и легкими для понимания, How It Works нравится читателям всех возрастов.
Посмотреть сделку
Развитие и эволюция человеческих конечностей
Abstract
Длинные ноги и короткие руки людей являются отличительными чертами приматов, результатом отбора, действующего в противоположных направлениях на каждую конечность на разных этапах нашей эволюционной истории. Этот мозаичный узор бросает вызов нашему пониманию взаимосвязи развития и эволюционируемости, потому что конечности серийно гомологичны, а генетические корреляции должны действовать как существенное ограничение на их независимую эволюцию.Здесь мы тестируем развивающуюся модель ковариации конечностей у антропоидных приматов и демонстрируем, что как люди, так и обезьяны демонстрируют значительно меньшую интеграцию между конечностями по сравнению с четвероногими обезьянами. Этот результат указывает на то, что ископаемые гоминины, вероятно, избежали ограничений на независимые вариации конечностей за счет редукции генетической плейотропии у обезьяноподобного последнего общего предка (LCA). Это критическое изменение в интеграции среди гоминоидов, которое отражается в макроэволюционных различиях в несоответствии между длинами конечностей, облегчило выбор пропорций конечностей современного человека и демонстрирует, как развитие помогает формировать эволюционные изменения.
Конечности — это последовательно повторяющиеся гомологичные структуры (1, 2), такие как зубы (3) и позвонки (4). Важно отметить, что событие древней дупликации, которое дало начало будущей задней конечности, реплицировало как анатомическую структуру грудного придатка, так и генетическую архитектуру, ответственную за индукцию его развития, формирование паттерна и рост (1, 5, 6). Это общее происхождение конечностей имеет важные последствия для их эволюции (7, 8). В частности, теория количественной генетики предсказывает, что генетические корреляции заставят конечности развиваться параллельно (9–11), особенно между гомологичными сегментами: i.е., стилопод, зевгопод и автопод (рис. 1 A ). Эта генетическая интеграция, которая отражается в паттернах фенотипической ковариации (12), влияет на вариабельность, доступную для отбора, и, следовательно, на независимую эволюционируемость конечностей (11, 13). В то время как высокие корреляции помогают гарантировать, что одинаковые пропорции конечностей масштабируются по размеру тела, такая же интеграция действует как существенное ограничение на эволюционность, ограничивая фенотипы, доступные для отбора (7, 8, 13). Таким образом, предполагаемый примитивный паттерн сильной интеграции между гомологичными модулями развития может быть значительным предубеждением для вариаций конечностей и эволюции в сторону масштабных вариантов (рис.1 B ).
Рис. 1.( A ) Общая генетическая архитектура конечностей, что демонстрируется их сходным паттерном Hox (серый, низкая экспрессия; черный, первичная экспрессия) (6), отражает их последовательную гомологию и предполагает иерархическую ковариационная структура конечностей, распределенная между конечностями и внутри них (7, 8). Показаны теоретические онтогенетические и функциональные модули конечности человека: стилопод (плечевая и бедренная кость), зевгопод (лучевая / локтевая, большеберцовая / малоберцовая) и аутопод (руки, ступни и пальцы).( B ) Ковариация между модулями развития конечностей в ответ на отбор определяет фенотипическое пространство (серые эллипсы) и независимую эволюционируемость конечностей. Когда корреляции низкие, фенотипическое пространство распределяется более равномерно (, верхний левый угол, ). Когда корреляции высоки, это пространство будет иметь тенденцию к лицам, различающимся по размеру, но с похожими пропорциями ( внизу справа ). Модель предсказывает, что мозаичная эволюция пропорций конечностей современного человека потребовала сокращения интеграции.( C ) Люди (черный круг) имеют относительные пропорции конечностей, отличные от обезьян (серые круги) и четвероногих обезьян (белые круги). Чистым направлением эволюционных изменений было увеличение относительной длины ног и меньшее уменьшение относительной длины рук, и оно приблизительно ортогонально межвидовой аллометрии относительных пропорций четвероногих конечностей (пунктирная линия). Данные основаны на исх. 19. ( D ) Селекция гомининов происходила по крайней мере в две фазы: ( и ) у ранних гомининов, таких как австралопитеки [ Au.afarensis (AL-288-1) и BOU-12/1, серый, реконструированный] относительная длина ног увеличилась с меньшими изменениями относительной длины руки (18, 20, 55, 56) [примечание: оценочный IMI Ar. ramidus сопоставим с Au. afarensis (25)] и ( ii ) у H. ergaster (KNM-WT15000) относительная длина предплечья уменьшилась, а длина ног еще больше увеличилась (18, 20, 21). Этот мозаичный узор указывает на независимую изменчивость конечностей и снижение интеграции. Расположение окаменелостей основано на опубликованных описаниях и оценках (18, 20, 21, 55, 56).
Люди представляют собой проблему для этой модели, потому что относительные пропорции наших конечностей, которые, как считается, отражают независимую адаптацию к двуногому движению (14–16), выносливости, бегу (17), ловкости рук и использованию инструментов (15, 18) , отличают нас от всех других приматов (14–16, 19) (Рис. 1 C ). Важно отметить, что летопись окаменелостей гомининов указывает на то, что эти селективные факторы, вероятно, различались с точки зрения времени, функциональной направленности и направленного воздействия на отдельные элементы каждой конечности (18, 20) (рис.1 D ). Таким образом, наши отличительные пропорции конечностей и связанные с ними адаптации эволюционировали через историю отбора, которая расходится по направлению и, что более важно, противоречит наследственным генетическим корреляциям из-за последовательной гомологии. Учитывая ограничения развития на независимые вариации конечностей, как мы можем согласовать прогнозы, полученные на основе общей генетической архитектуры конечностей, с историей эволюции гомининов?
Возможное решение этой проблемы состоит в том, чтобы предположить, что в какой-то момент эволюции приматов интеграция между гомологичными модулями передних и задних конечностей была уменьшена, что привело к более высокой степени вариационной независимости размера и формы конечностей.Наша модель предсказывает, что выбор независимой функции конечностей должен привести к изменениям ковариационной структуры конечностей. В частности, предполагается, что люди и обезьяны будут демонстрировать пониженные фенотипические корреляции между передними и задними конечностями по сравнению с четвероногими. Если уменьшение интеграции между передними и задними конечностями является продуктом отбора для более независимой функции конечностей, то время любого изменения интеграции имеет значение для реконструкции предкового позиционного поведения.Более того, поскольку интеграция связана с независимой эволюционирующей способностью конечностей, наша модель предсказывает, что в линиях с более слабой интеграцией эволюционное неравенство в пропорциях конечностей будет относительно выше, тогда как в линиях с более сильной интеграцией диспропорции в пропорциях конечностей будут относительно ниже.
Мы проверили эти прогнозы, сравнив людей с выборкой человекообразных обезьян (шимпанзе, гориллы и гиббоны) и четвероногих обезьян Старого и Нового Света (макаки и листовые обезьяны, а также белки и совы, соответственно) (методы и таблица S1 ).Для каждого вида мы вычислили парные фенотипические корреляции между длинами отдельных элементов конечностей стилопода (плечевая и бедренная кость), зевгопода (лучевая и большеберцовая кость) и аутопода (пястная кость III и плюсневая кость III). Мы также использовали метод для учета артефактов в оценках корреляции, которые ранее не признавались в сравнительных исследованиях. В частности, мы скорректировали различия в величине фенотипических корреляций из-за изменчивости выборки населения (SI Text, Figs, S1 – S3).Этот метод позволил нам сделать вывод о существенных различиях в интеграции с ранее невозможной степенью уверенности. Из этих данных мы сделали выводы о моделях модульности с частичными корреляциями и оценили интеграцию между конечностями и внутри них, используя анализ структуры собственных значений корреляционной матрицы и дисперсии.
Результаты
Наш анализ показывает, что люди, обезьяны и обезьяны имеют общую модульную структуру, в которой гомологичные по развитию элементы передних и задних конечностей демонстрируют повышенные частичные корреляции по сравнению с негомологичными элементами, несмотря на миллионы лет эволюции, резкие изменения пропорций конечностей. , и узкоспециализированные приспособления (рис.2 A и таблицы S2 – S10). Напротив, внутриконечностные корреляции между функциональными модулями более вариабельны, вероятно, потому, что они обладают меньшим количеством плейотропных взаимодействий и, таким образом, более свободны для ответа на отбор для скоординированной вариации (например, для передвижения). Люди демонстрируют сильные частичные корреляции соседних элементов задних конечностей, согласующиеся с отбором для двуногого передвижения, тогда как поддерживающие обезьяны демонстрируют более последовательную модульность между элементами передних конечностей, а четвероногие обезьяны Старого и Нового Света демонстрируют значительные частичные корреляции как в пределах задних конечностей, так и в негомологичных элементах.
Рис. 2.( A ) Антропоидные приматы демонстрируют последовательную модульность развития конечностей. Модульность внутри конечностей отражает различные функциональные сигналы (например, двуногие задние конечности у людей и подвешивание передних конечностей у обезьян). Модули (частичные корреляции P <0,05) показаны в виде прямоугольников между элементами. Модули заштрихованы относительно силы расчетной преобразованной корреляции Fisher- z . Показан расчетный коэффициент корреляции Пирсона.(MC — пястная кость; MT — плюсневая; R — лучевая; T — большеберцовая; H — плечевая; F — бедренная). Виды расположены по филогенетическому родству, как показано на Bottom . ( B ) Люди значительно менее интегрированы по сравнению с четвероногими обезьянами и похожи на обезьян, что указывает на то, что сокращение интеграции и более независимо эволюционирующие конечности характерны как для ископаемых гомининов, так и для гоминоидов. Коробчатые диаграммы показывают нижний и верхний квартили, медианное значение дисперсии повторных собственных значений (VE) (10000 повторов).Усы указывают на доверительный интервал 95% оценки. Пунктирные линии и заштрихованные прямоугольники показывают средний VE для гоминоидов (= 1,79), церкопитекоидов (= 3,32) и цебоидов (= 2,90) и 95% доверительный интервал, соответственно.
Сохранение модульности между конечностями убедительно подтверждает предсказание нашей модели о том, что совместное развитие стимулирует вариативную интеграцию конечностей, но только усугубляет загадку того, как расходящиеся пропорции развивались мозаичным образом. Однако сравнение структуры собственных значений и средних корреляций подтверждает гипотезу о том, что интеграция снижается у видов с функционально независимыми конечностями.И люди, и обезьяны значительно менее интегрированы, чем четвероногие обезьяны (снижение на 34–38%) (Рис. 2), и значительно различаются по тому, насколько сильно элементы конечностей коррелируют по большинству разделов матрикса (Таблица 1). Учитывая общие различия в интеграции людей и обезьян по сравнению с обезьянами, мы делаем вывод из этих результатов, что диссоциация гомологичных модулей передних и задних конечностей произошла до эволюции пропорций конечностей современного человека, а не одновременно с ними.
Таблица 1.Среднее значение Фишера- z преобразованные корреляции и дисперсия собственных значений (VE)
Сравнение пропорций между конечностями живых антропоидных приматов подтверждает предсказание о том, что интеграция имеет макроэволюционные эффекты на модель дивергенции конечностей. Анализ данных по всем живым родам обезьян показывает, что пропорции конечностей гоминоидов значительно более изменчивы по сравнению с церкопитекоидами и цебоидами (критерий Левена, P <0,05) (рис. 3 A и B и таблицы S11 и S12). .Этот результат, несмотря на существенную таксономическую, экологическую и функциональную диверсификацию у обезьян, без учета ограничений развития может увеличивать разнообразие пропорций конечностей. Более того, эта закономерность не может быть полностью объяснена более поздней диверсификацией на уровне видов, потому что, хотя излучения гиббонов, макак, генонов и тамаринов имеют сопоставимую временную глубину и количество видов, гиббоны значительно более изменчивы в пропорциях конечностей, чем любая из этих четвероногих линий (Тест Левена, P <0.05; (Рис.3 C и Таблица S13). На основании этих результатов мы заключаем, что уменьшение интеграции конечностей и большая независимая эволюционируемость конечностей характерны для людей и обезьян и отражают эволюцию адаптации (й), которая подчеркивает функционально расходящееся использование конечностей. Напротив, низкое разнообразие пропорций конечностей у четвероногих таксонов, вероятно, отражает сдерживающий эффект сильной интеграции на возможность эволюции пропорций между конечностями в сторону масштабных вариантов.
Рис.3.Таксоны гоминоидов имеют более изменчивые пропорции между конечностями по сравнению с церкопитекоидными и цебоидными обезьянами, что предсказывается их уменьшенной интеграцией (все сравнения: P <0,05, критерий Левена). Каждая точка представляет собой значение одного вида. Черные ящики показывают среднее значение для каждой группы. ( A ) Сравнение относительной длины рук и ног для видов со смешанной функцией из каждого суперсемейства (Таблица S11). ( B ) Сравнение между подвешивающими обезьянами и четвероногими видами обезьян по межмембральному индексу (таблица S12).( C ) Сравнение излучений приматов на сопоставимых временных глубинах и численности видов (Таблица S13).
Обсуждение
Уменьшение интеграции между конечностями у предков обеспечивает контекст развития как для мозаичной эволюции пропорций конечностей гомининов, так и для интерпретации позиционного поведения (я), характеризующего последнего общего предка (LCA) людей и обезьян. У ранних гомининов снижение интеграции конечностей облегчило бы эволюцию относительно более длинной ноги, что является отличительным признаком выбора для двуногости, но также позволило бы пропорциям передних конечностей остаться относительно неизменными.Более поздние сокращения длины предплечья у Homo демонстрируют аналогичные некоррелированные изменения с длиной задней конечности (20, 21), таким образом, летопись окаменелостей предполагает, что передняя конечность оставалась относительно независимой от задней конечности в то время. Эта некоррелированная закономерность показательна, потому что, хотя более длинная нога положительно влияет на длину шага и энергетическую эффективность двуногости (22), длинные руки более адаптивны в древесных контекстах, часто связанных с ранними гомининами (например, как адаптация к вертикальному лазанию или подвешиванию). ) (20, 23), но они увеличивают дистальные нагрузки и могут выступать в качестве энергетической нагрузки при шаговом двуногом беге, беге на выносливость (17) или при тонких манипуляциях и использовании инструментов, обнаруженных у более поздних гомининов.Редукция интеграции способствовала бы независимым изменениям между конечностями, которые в противном случае были бы коррелированными и потенциально неадаптивными, и поэтому, вероятно, сыграли важную роль в диссоциированном и мозаичном процессе, посредством которого эволюционировали пропорции конечностей современного человека.
Утверждается, что происхождение человеческого двуногого поведения — ортоградное и поддерживающее позиционное поведение, присущее всем обезьянам (например, ссылка 24), но точный преадаптивный режим (ы), характерный для LCA, все еще является источником значительных споров ( е.г., исх. 23–30). Наиболее скупая реконструкция представленных здесь результатов поддерживает предковый морфотип, в котором позиционное поведение диссоциировано на более независимые функции конечностей. Африканские обезьяны являются наиболее логичной моделью для этого морфотипа, потому что, помимо их тесной филогенетической близости, их смесь адаптаций конечностей отражает выборочный компромисс для позиционного поведения, начиная от наземного четвероногого и подвешенного кормления до ограниченного двуногого поведения (23).Соответствующий им паттерн пониженной интеграции, который согласуется с повышенной вариабельностью в других посткраниальных структурах (например, ссылки 31, 32), подразумевает, что африканская обезьяноподобная LCA имела бы относительно более некоррелированные вариации, на которые отбор мог действовать независимо. Напротив, LCA, похожая на четвероногую обезьяну (например, ссылка 25), потребует нескольких эпизодов независимой эволюции расходящихся пропорций и функции конечностей у обезьян и будет означать LCA, конечности и посткраниум которой фундаментально более ограничены для совместного развития.По этой причине LCA, подобная африканской обезьяне, которая была свободна от некоторых ограничений высокой интеграции, имела бы больший потенциал для эволюции в сторону человеческого состояния, чем более интегрированная LCA на четвероногих, несмотря на то, что обезьяны занимали фенотипическое пространство, противоположное этому. людей. В этом контексте эволюционная траектория от обезьяноподобного LCA к человеку, которая «пересекает» пропорции четвероногих конечностей (рис. 1 C и D ), предполагает, что более обезьяноподобные пропорции конечностей у ранних гомининов (например.g., Ardipithecus ramidus , Australopithecus afarensis ) (25) являются предсказуемыми промежуточными результатами мозаичного отбора на двуногость из более эволюционирующей обезьяноподобной LCA, а не общих примитивных черт из ограниченной LCA на четвероногих.
Эти результаты дополнительно указывают на то, что интеграция имеет широкие макроэволюционные последствия для моделей диверсификации пропорций конечностей, что видно по большему разнообразию пропорций конечностей у живых обезьян по сравнению с четвероногими обезьянами.Этот эффект интеграции на долгосрочные паттерны эволюционного разнообразия может помочь объяснить посткраниальную изменчивость и мозаичную эволюцию ископаемых обезьян (например, ссылки 33–37). Наши результаты предполагают одну возможность того, что изменчивость может отражать эволюционное «экспериментирование» в посткраниальном плане тела, которое стало возможным благодаря эффекту отбора для функциональной диссоциации на независимую эволюционируемость конечностей. Таким образом, тенденция к диссоциации функции конечностей могла быть ключевой адаптацией ранних гоминоидов, которая помогла сформировать их более позднюю посткраниальную эволюционную радиацию.Напротив, сильная интеграция, по-видимому, смещает обезьян в сторону масштабных вариантов со сходной пропорцией конечностей, что ограничивает их посткраниальную диверсификацию. Летопись окаменелостей церкопитекоидов согласуется с этой интерпретацией, потому что она не указывает на серьезные изменения в плане тела или пропорциях конечностей (14, 38), достаточных для того, чтобы можно было разумно заключить, что таксоны ископаемых предков являются близкими посткраниальными аналогами их живых потомков. Однако, учитывая большое количество живых видов обезьян Старого Света по сравнению с обезьянами, очевидно, что это ограничение не повлияло на их эволюционный успех.
Хотя конечной причиной этого паттерна интеграции является дивергентный отбор по длине элементов в каждой конечности, ближайший механизм, вероятно, находится во влиянии этого отбора на генетическую плейотропию. В этом случае отбор, связанный с независимой функцией конечностей у обезьян и людей, вероятно, действовал, чтобы уменьшить или ограничить эффекты генетических влияний, общих между гомологичными элементами конечностей. Другие возможности, такие как различия в интеграции, вторично связанные с вариациями самой функции, менее вероятны, потому что привычная активность не оказывает значительного влияния на структуру или величину ковариации длины конечности, по крайней мере, в экспериментальных условиях (39).С другой стороны, вероятность того, что снижение плейотропии между конечностями обеспечивает механизм снижения интеграции, убедительно подтверждается открытием генетических элементов, которые конкретно изменяют корреляции между количественными признаками (40), предполагая, что степень интеграции является как наследственной, так и наследственной. эволюционируемый. Следовательно, источник этих наблюдаемых различий, вероятно, находится в эволюции генов, которые уникальным образом экспрессируются либо в конечностях, либо в отборе для изменения регуляции времени, местоположения и функции генов, общих для обеих конечностей.Таким образом, выяснение того, что это за механизмы и как они вносят вклад в дифференциацию передних и задних конечностей приматов, будет иметь решающее значение для реконструкции того, как человеческие конечности реагировали на отбор на выполнение различных функций.
Таким образом, сравнение фенотипических корреляций у людей и антропоидных приматов подтверждает важную роль общей генетической архитектуры и развития в структурировании паттерна дивергенции конечностей. В более общем плане, связывая микро- и макроэволюционные процессы в количественной структуре, эти результаты служат моделью для эволюции индивидуации и диверсификации частей других последовательных гомологов [e.g., пальцы (41), зубы (3), позвонки (4), придатки насекомых (42), пятна бабочек (43) и т. д.], которая применима как для растений, так и для животных. Кроме того, это показывает, что, поскольку интеграция в процессе развития развивается в ответ на отбор, паттерны изменений могут помочь информировать интерпретации временных рамок и давления отбора, вовлеченного в морфологическую диверсификацию, особенно в отношении летописи окаменелостей. Дальнейшая работа, направленная на определение и тестирование этих проблем (например, ссылки 44, 45), в конечном итоге поможет определить, как эволюционные модификации систем развития ограничивают или облегчают эволюционные изменения.
Методы
Данные.
Выборка состоит из взрослых самцов и самок Homo sapiens sapiens (современные люди, n = 133), Pan troglodytes troglodytes (обычный центральный шимпанзе, n = 43), Gorilla gorilla gorilla (западная равнинная горилла, n = 62), Hylobates lar (белорукий гиббон, n = 63), Macaca mulatta (макака-резус, n = 176), Trachypithecus cristatus (серебристый лист обезьяна, n = 59), Saimiri sciureus (обезьяна-белка, n = 102) и Aotus trivirgatus (обезьяна-сова, n = 74) (Таблица S1).Для каждого вида мы собрали данные о максимальной измеренной длине каждого из следующих элементов конечностей: плечевой кости, лучевой кости, III пястной кости, бедренной кости, голени и III плюсневой кости. Различия в средних значениях между полами, популяциями или этнической принадлежностью были удалены путем центрирования данных (то есть различия в средних значениях были добавлены к индивидуальным значениям из группы с более низким средним).
Модульность.
Матрицы частичной корреляции были рассчитаны на основе матриц корреляции видов (46) (таблицы S2 – S10).Частичные корреляции измеряют корреляцию двух переменных, обусловленную всеми другими переменными, то есть корреляцию двух переменных независимо от информации из других переменных в корреляционной матрице. Значимость условной независимости двух переменных оценивалась с помощью теоретико-информационной меры, известной как отклонение исключения краев и распределения χ 2 (46). Существенные частные корреляции показаны в виде модульных графиков между элементами конечностей.
Интеграция.
Мы использовали две дополнительные меры: ( i ) дисперсию собственных значений (VE) (47, 48), которая получается путем вычисления собственных значений матриц корреляции оценок видов и их дисперсии популяций; и ( ii ) средние преобразованные корреляции Fisher- z для пяти наборов теоретических функциональных модулей и модулей развития [гомологичных (стилопод, зевгопод, автопод), негомологичных (т.е. почти гомологичных), передних конечностей, задних конечностей и всего] . Чтобы исправить артефакты дисперсии в корреляциях, мы использовали метод повторной выборки.Во-первых, для каждого вида мы провели повторную выборку каждого центрированного набора данных с заменой (10000 повторов), и для каждого повтора мы рассчитали матрицу корреляции, дисперсию популяции [средний коэффициент вариации (CV) для всех признаков], VE, среднее значение Фишера. z преобразованные корреляции и матричная корреляция ( r m ) с исходным центрированным набором данных. Во-вторых, используя только те реплики, у которых r m > 0,95 (т. Е. Корреляционная структура, статистически не отличимая от идентичности), мы оценили лог-линейную связь между каждой из этих переменных и средним CV признака (рис.S4). В-третьих, используя эту предполагаемую взаимосвязь, мы оценили корреляции между видами при общей дисперсии популяций. Матрицы корреляции и оценки VE стабильны ( r m > 0,90) для значений CV 3,5–5,5% (т. Е. В пределах 95% доверительного интервала для оценок CV всех видов). Таким образом, мы сообщаем оценочное значение для VE и средних преобразованных корреляций Fisher- z при единственном значении CV (= 4%) с 95% доверительными интервалами, оцененными путем повторной выборки (10 000 повторов).Все процедуры выполнялись с использованием программного обеспечения PopTools 3.8 (49) или R (50).
Несоответствие пропорций конечностей.
Мы использовали оценки пропорций конечностей, рассчитанные на основе этого набора данных и других опубликованных источников (14, 16, 19, 51–53) (таблицы S11 и S12). Данные Schultz (19) рассчитываются как максимальная длина конечности от плеча / бедра до кончика пальца или подошвы стопы, деленная на длину туловища, что отличается от более часто используемого интермембрального индекса (IMI) [т.е., (длина плечевой кости + длина лучевой кости) / (длина бедренной кости + длина большеберцовой кости) × 100] (14, 16). Расхождение рассчитывалось как абсолютное значение среднего отклонения от среднего значения по линии, а значимость оценивалась с помощью теста Левена (54).
Благодарности
Мы благодарим Джуди Чупаско (Гарвардский университет, Музей сравнительной зоологии, Кембридж, Массачусетс), Джин Тернквист (Университет Пуэрто-Рико, Кайо-Сантьяго), Лаймана Джеллема (Кливлендский музей естественной истории, Кливленд, Огайо) и Ричарду Торингтону (Полевой музей естественной истории, Чикаго) за предоставление доступа к находящимся у них образцам.Мы также благодарим Д. Э. Либермана, Р. С. Маркучио, М. Л. Зельдича, М. Э. Штайпера, К. П. Ролиана и М. Д. Роуза за неопубликованные данные и / или комментарии к рукописи. Мы благодарим двух анонимных рецензентов за их полезные комментарии. Это исследование было поддержано грантом Alberta Ingenuity Grant 200300516 (для Нью-Йорка) и грантом Национального совета по науке и технике № 238992-06 (для B.H.).
Сноски
- 1 Кому следует направлять корреспонденцию. Электронная почта: nathan.m.young {at} Gmail.com.
Автор: N.M.Y. спланированное исследование; N.M.Y. проведенное исследование; N.M.Y. и Б. внесены новые реагенты / аналитические инструменты; N.M.Y., G.W. и B.H. проанализированные данные; и N.M.Y., G.W. и B.H. написал газету.
↵ * Для этой статьи с прямым представлением был назначен редактор.
Авторы заявляют об отсутствии конфликта интересов.
Эта статья содержит вспомогательную информацию на сайте www.pnas.org/cgi/content/full/0
- 6107/DCSupplemental.
Что влияет на изменение роста скелета в костях конечностей человека? — Подкаст Arch and Anth
В этом выпуске Ан-Ди Йим (Университет Иллинойса в Урбана-Шампейн) рассказывает нам о своем докторском исследовании, посвященном различным экологическим и генетическим факторам, которые способствуют росту скелета и формированию скелета до взрослого человека.
Ан-Ди фотографирует череп (не показан) в рамках своей судебно-медицинской экспертизы.
Что такое подход «популяционной генетики» и как антропологи могут использовать его для количественной оценки степени, в которой различные эволюционные влияния влияют на рост скелета? Почему у некоторых детей и подростков конечности короче или длиннее, чем у других во всем мире, и проявляют ли скелеты молодых людей во всем мире эти тенденции с самого рождения? Как Ан-Ди объединяет данные из судебно-медицинских образцов, баз данных генома и археологических материалов из разных популяций мира в своем анализе?
Ан-Ди ведет раскопки во время своей полевой судебно-медицинской экспертизы в Сомали.
Мы также говорим о преподавании биологической антропологии, а также о предыдущей работе Ан-Ди по интернированию и выходу в поле практики судебной антропологии в реальных ситуациях. Будучи оба из Восточной Азии, Майкл делится своими впечатлениями от биологической антропологии в Гонконге, в то время как Ан-Ди рассказывает о тайваньской антропологии, оба с прицелом на улучшение представления образцов восточной и юго-восточной Азии в будущей антропологической работе.
Виртуальное 3D-сканирование бедренных и большеберцовых костей человека.
Послушайте, что говорит Ан-Ди, нажав «Воспроизвести» в эпизоде ниже. Вы также можете подписаться на нашу RSS-ленту или найти нас в Spotify, Stitcher, iTunes или в любом другом месте, где вы обычно найдете подкасты!
Показать примечания
точек на скелете Ан-Ди наблюдал за тем, где жертвы судебно-медицинской экспертизы, скорее всего, будут расчленены, что помогло развеять миф о том, что те, кто совершает такие преступления, должны быть образованы, иметь медицинскую подготовку.
Если у вас есть отзывы или вопросы к Ан-Ди, вы можете найти его в Twitter и на его веб-сайте.
Вы также можете найти Майкла в Twitter и Instagram.
Подкаст Arch and Anth находится в Twitter и Instagram, и у него есть страница в Facebook.
Если вам понравился этот эпизод, и вы хотите внести свой вклад в шоу, посетите страницу Patreon, чтобы узнать, как это сделать.
Интерактивный атлас анатомии человека
Анатомия артерий и костей нижней конечности на основе 3D-снимков и ангиограммы (ангиографии).
Эта часть интерактивного атласа анатомии человеческого тела посвящена артериальной сосудистой сети тазового пояса, таза, бедра, колена, ноги и ступни, а также изучению костей и суставов. Он включает в себя трехмерную реконструкцию костей и артерий из ангиоКТ с введением контрастного вещества (йода), ангиографический снимок, коррелированный с трехмерным изображением, и цифровую рентгенографию скелета нижней конечности.
Кость и артерия тазового пояса: анатомия на основе медицинских изображений (3D-изображения и ангиограмма, адаптированные на основе компьютерной томографии)
Подробная процедура этого интерактивного атласа анатомии нижних конечностей:
Мы выбрали нормальное ангиоКТ (компьютерная томография) обследование нижних конечностей: регистрация была сделана с помощью спиральной мультидетекторной КТ (MDCT) после инъекции йодированного контраста.Осевые изображения были подвергнуты последующей обработке с использованием программного обеспечения для последующей обработки на рабочей станции для мультимодальной реконструкции и трехмерного изображения кости и кровеносных сосудов, затем было создано аналогичное ангиографическое изображение.
Ангиография (ангиограмма), адаптированная из ангиоКТ, показывающая все помеченные тазовые артерии: внутреннюю и внешнюю артерию, половую артерию, ягодичную артерию, бедренную артерию…
Изображения DICOM были экспортированы, а затем интегрированы с помощью Adobe Animate для создания этого модуля анатомии. Анатомические структуры были помечены на трехмерном изображении.
Рентгенография скелета: все анатомические структуры (кости и суставы) тазового пояса, отмеченные на рентгенографическом снимке (X-Rays)
Мы также выбрали непатологические цифровые рентгеновские снимки: таза, бедра, колена, голеностопного сустава и стопы, с маркировкой структур скелета и суставов.
Нормальная артериография голени: изображение, адаптированное из MDCT (MIP), показывающее подколенную, малоберцовую, переднюю большеберцовую артерию и заднюю большеберцовую артерию
Все анатомические структуры Terminologia Anatomica переведены на французский, английский, испанский, португальский, русский, итальянский, немецкий, польский, корейский, японский и китайский языки.Этот модуль можно использовать как медицинский словарь.
Рентгенография тазобедренного сустава: анатомия всего тела человека на основе медицинских изображений: бедренная кость, тазобедренная кость, вертлужная впадина, головка бедренной кости, гребень подвздошной кости …
Визуализация периферических сосудов нижней конечности:
Переднезадняя рентгенология (рентген) коленного сустава: анатомия бедренной кости, надколенника, большеберцовой кости, малоберцовой кости, латерального и медиального мыщелка, межмыщелкового бугорка…
Мы предпочли использовать ангиоКТ нижней конечности, а не цифровую артериографию, потому что это позволяет пользователю делать корреляцию между ангиографическими изображениями артерий с трехмерными структурами.
Вкладка 3D / ангиографии может использоваться для выбора этих двух видов обзора.
рентгенологический атлас нижней конечности: рентгенограмма колена (вид сбоку), показывающая суставы (бедренно-пателлярный сустав, бедренно-большеберцовый сустав, большеберцовый сустав) и кости
Поля обзора — это те поля зрения, которые обычно используются для ангиографии: общее видение всей артериальной сосудистой сети (от аорты до стопы), а затем 3 конкретных поля зрения: подвздошная, бедренная и треножная артерии
Мы разделили метки артерий на разные группы: аорта и ее ветви, внутренняя подвздошная артерия и ее ветви, наружная подвздошная артерия и ее ветви.Это уменьшает количество меток, отображаемых на каждом изображении, и обеспечивает лучший образовательный подход.
Были помечены только артерии, видимые на трехмерном изображении, это объясняет, почему на ангиографическом изображении некоторые артерии видны только под определенными углами поворота.
Этот модуль разработан для учителей анатомии, включая сосудистых заболеваний, студентов-медиков и студентов парамедицинского профиля (медсестер, радиологов), а также практикующих специалистов в области сосудистой медицины, ультразвукового и допплеровского обследования, сосудистой хирургии, интервенционной радиологии, кардиологии, ангиологии и др. эндокринология.Он облегчает исследование артериальных заболеваний нижних конечностей (атерома, атеросклероз), эмболизаций в экстренных случаях, баллонной ангиопластики или стентов, сосудистого обходного анастомоза, гинекологической эмболизации (миома матки, первичные послеродовые кровотечения (примечание: пациент мужчина, поэтому имеются некоторые отличия от сосудистой сети женского таза)
Визуализация скелета нижней конечности
Также были помечены суставы и кости скелета таза, бедра, бедра, колена, ноги, лодыжки и стопы.В конце этого модуля обратите внимание, что традиционная анатомическая рентгенология может быть полезна в обычной практике, поэтому вы можете найти некоторые маркированные цифровые рентгенограммы.
Рентгенография (рентген) голеностопного сустава: анатомия на виде спереди, показывающая большеберцовую, малоберцовую, таранную кость, латеральную и медиальную лодыжку.
Радиоанатомия голеностопного сустава: рентгенологическое исследование голеностопного сустава (вид сбоку) с анатомическими структурами, обозначенными как пяточная кость, таранная кость, ладьевидная кость, голеностопный сустав.
Длинные кости и суставы представлены таким образом, чтобы можно было проследить каждую структуру на всем ее протяжении.
Пришлось использовать группы анатомических структур: крестец, тазик, бедро, надколенник, большеберцовая кость, малоберцовая кость, кости стоп (предплюсна и плюсна). По техническим причинам метки костных структур появляются на ангиографических изображениях, а некоторые кости не видны; пожалуйста, простите нас за эту проблему.
Суставы тоже были промаркированы, хотя их легче изучить на таких обследованиях, как КТ или МРТ артрография.
Превосходный рентгеновский снимок стопы со всеми отмеченными анатомическими структурами (кости, суставы)
Этот модуль менее специфичен, чем модули визуализации срезов, поэтому он полезен для всех, кто интересуется анатомией скелета и сосудов и патологией нижних конечностей:
- Студенты-медики, изучающие медицинские степени и степени бакалавра (MBBS, MBChB, MD, DO, MDCM, BMed и т. Д.) Или экзамен на получение медицинской лицензии в США
- Медсестры, санитары, вспомогательные медсестры, фельдшеры, дипломированные медсестры или лицензированные медсестры
- Волонтеры больницы
- Скорая помощь и парамедики
- Техники, техники по флеботомии, хирургические технологи и техники, обученные работе с таким оборудованием, как рентгеновские аппараты
- Практикующие врачи
- Провизоры
- Медицинские технологи и физиотерапевты, работающие с медперсоналом
- Учителя анатомии или профессора медицинских университетов, медицинских школ или медицинского факультета
- Стажеры или ассистенты, хирурги-ортопеды
- Ревматологи, ортопеды
- Любой врач, который хочет объяснить костно-мышечной патологии пациенту: перелом, растяжение связок, артрит
Боковой вид стопы на рентгенограмме (рентгенограмме): плюсневые кости, фаланга, кубовидная, ладьевидная, клиновидные кости…
Библиография:
- Атлас анатомии человека, профессиональное издание Фрэнка Х. Неттера · Издатель: Saunders; 4 издание (13 июля 2006 г.) Язык: Английский · ISBN-10: 1416036997 · ISBN-13: 978-1416036999
- Terminologia Anatomica: Международная анатомическая терминология, разработанная FCAT · Издатель: Thieme Medical Publishers; Двуязычное издание (1 января 2000 г.) · Язык: Латинский / английский · ISBN-10: 0865778086 · ISBN-13: 978-0865778085
- Диагностическая и хирургическая визуализация: анатомия скелетно-мышечной системы (международное издание): опубликовано Amirsys · Издатель: Lippincott Williams & Wilkins; 1 издание (1 декабря 2006 г.
