
расчет калорийности и БЖУ в сутки для похудения
Добрый день, Дамы и Господа – читатели!
Полагаю, все уже уяснили, что для здорового рациона очень важно потреблять «качественные» макроэлементы. Для тех, кто забыл: макроэлементами называются органические вещества, непосредственно участвующие в жизнедеятельности организма, — это белки, жиры и углеводы. Давайте ещё раз вспомним, как правильно рассчитывать норму БЖУ для себя.
На заметку! Я уже поднимала тему КБЖУ, вот тут и тут. А в конце статьи будет несколько лайфаков о том, как упростить процесс подсчетов.
Норма белков, жиров и углеводов
Оговорюсь сразу, что говорим мы только о качественной еде. Если речь идет, к примеру, об углеводах, то автономно подразумеваются сложные углеводы, которые медленно перерабатываются нашим организмом. Пшеничные булки – тоже углеводы и дневную норму ими тоже можно набрать. Но, скорее всего, эти булки постепенно превратят Вас самих в булку.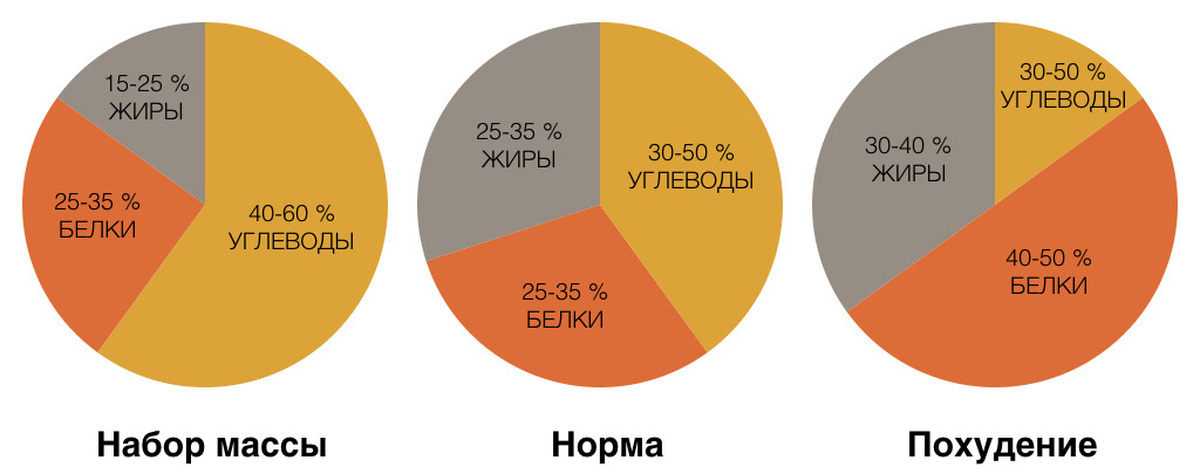
Итак, существует общепринятое соотношение БЖУ, при котором человеческий организм чувствует себя хорошо и не испытывает метаболического стресса.
1). Это стандартная усредненная норма БЖУ (показатели можно варьировать в пределах 10%):
Такого соотношения следует придерживаться в случаях, если Вы хотите оставаться в той форме, в которой сейчас находитесь. То есть, если худеть или набирать вес не нужно.
2). Если требуется похудеть, то соотношение немного изменяется:
3). А для массонабора нужно увеличить объемы потребляемой пищи, выбрав 1-ое или 2-ое соотношение (опять же в зависимости от того, устраивают ли Вас параметры тела).
Расчёт индивидуальной нормы БЖУ
Для каждого человека есть индивидуальная норма макроэлементов, требуемая для стабильного метаболизма. Чтобы считать себя полноценным ПП-шником, нужно высчитать эту норму.
ШАГ 1
Начнем с простого: сколько калорий нужно потреблять в сутки? Рассчитать проще всего в онлайн-калькуляторе ниже (будь то стандартная норма, или для похудения).
Для достижения конкретного результата обязательно проконсультируйтесь с тренером или диетологом. Калькулятор не учитывает множество нюансов и состояние вашего здоровья.
| Пол: МУЖЖЕН | Возраст: 101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960616263646566676869707172737475767778798081828384858687888990919293949596979899100 | Вес: 30313233343536373839404142434445464748495051525354555657585960616263646566676869707172737475767778798081828384858687888990919293949596979899100101102103104105106107108109110111112113114115116117118119120121122123124125126127128129130131132133134135136137138139140141142143144145146147148149150151152153154155156157158159160161162163164165166167168169170171172173174175176177178179180181182183184185186187188189190191192193194195196197198199200 | Рост: 110111112113114115116117118119120121122123124125126127128129130131132133134135136137138139140141142143144145146147148149150151152153154155156157158159160161162163164165166167168169170171172173174175176177178179180181182183184185186187188189190191192193194195196197198199200201202203204205206207208209210211212213214215216217218219220 | |
| Уровень физической активности: Сидячая работаТруд не физический, но много хожу (езжу)Преимущественно физический трудТяжелый физический трудГружу вагоны с цементом круглые сутки Ваша цель: Хочу похудетьХочу набрать мышечную массуХочу улучшить фигуру не меняя вес Количество тренировок в неделю: | ||||
ШАГ 2
Собираем все имеющиеся показатели. Теперь можно рассчитать количество БЖУ, которое требуется в сутки худеющему человеку (рассматриваем худеющих потому что таковых, как правило, больше:)).
Берем процентное соотношение БЖУ в сутки:
| Белки | 40-45% |
| Жиры | 15-20% |
| Углеводы | 30-40% |
Наша задача – перевести это соотношение в калории и граммы, которыми мы сможем оперировать при формировании рациона. Для этого нужно знать, сколько ккал в 1 грамме каждого макроэлемента.
| 4 ккал | |
| 1 гр жиров | 9 ккал |
| 1 гр углеводов | 4 ккал |
Высчитываем, сколько калорий приходится на каждый процентный показатель макроэлементов, то есть:
- Сколько калорий составят 45% белков от Ксут – это Кбел
- Сколько калорий составят 20% жиров от Ксут – это Кжиры
- Сколько калорий составят 40% углеводов от Ксут – это Куглев
Теперь суточные калории от белков, жиров и углеводов переводим в граммы:
- Кбел ÷ 4 ккал = Кол-во белковсут
- Кжиры ÷ 9 ккал = Кол-во жировсут
- Куглев ÷ 4 ккал = Кол-во углеводовсут.

Вот и всё! Получили норму БЖУ и в граммах, и в калориях на сутки. Что, не любите формулы? Тогда читаем дальше – посмотрим пример рассчета.
Пример расчета нормы БЖУ
- Скажем, мы рассчитали в калькуляторе свою суточную норму калорий для похудения Ксут – 1340 ккал.
- Переведем теперь суточные проценты БЖУ при похудении в калории с помощью Ксут:
- 45% белков от 1340 ккал = 603 ккал;
- 20% жиров от 1340 ккал = 264 ккал;
- 40% углеводов от 1340 ккал = 536 ккал.
- Переведем суточные калории для БЖУ в граммы:
- Белки: 603 ккал ÷ 4 ккал = 150,7 гр;
- Жиры: 264 ккал ÷ 9 ккал = 29 гр;
- Углеводы: 536 ккал ÷ 4 ккал = 134 гр;
То есть, для похудения человеку, рассматриваемому в примере, требуется в сутки 1340 ккал, которые обеспечиваются 150.7 граммами белка, 29 гр жиров и 134 гр углеводов.
Лайфхаки по подсчетам и следованию норме БЖУ
- Пользуйтесь калькуляторами калорий.

- Ведите дневник питания. Можете записывать сразу в приложение на телефоне или в бумажный блокнот. Но в конце дня подведите итог и посчитайте – сколько и чего Вы съели.
- Делайте перерывы: слишком строгие рамки, со словами: «О, Боже! Вчера я сжевала яблоко и не взвесила его! Как я узнаю, сколько калорий и БЖУ теперь съела?!», приводят к стрессам и психологическим нарушениям. Живите проще.
- Создайте список универсальных простых (и немножко сложных) блюд, подсчитайте для них КБЖУ. Это Вам очень поможет, когда на приготовление блюд и их пищевой анализ нет времени, сил, желания.
- Придумайте рацион на неделю. Уделите на выходных 15 минут и продумайте недельное или, хотя бы, трехдневное меню. Серьезно, планирование очень хороший помощник!
- Исходя из пункта 8 (↑) можно составить и список продуктов, которые тоже рекомендую покупать заранее на 3 дня вперед.
 Так у Вас всегда будет «сырье», из которое можно будет сотворить что-то «годное-полезное».
Так у Вас всегда будет «сырье», из которое можно будет сотворить что-то «годное-полезное».
На данный момент это всё. А начинаю новую «похудательную кампанию», ибо я, как говорит бабушка, «немножко раздобрела». Стресс, наверное. Пойду в FatSecret.
Баланс БЖУ
Расчет с нуля – процедура сложная, в нее входит такое количество факторов, что просчитывать соотношение бжу каждый раз слишком долго. Для быстрых вычислений созданы специальные формулы. По этим формулам достаточно просто вычислить не средние, а точные показатели необходимой суточной калорийности.Формула подсчета Миффлин-Сан Жеора
Она создана в 2005 году диетологами из США, отдельно для мужчин и женщин. Для высокой точности расчетов применяется коэффициент А – показатель физической активности человека. Для простоты используется пять основных значений:
· 1.2 — физическая активность стремится к нулю. Офисные работники и другие представители профессий, не требующих физической активности, не занимающиеся спортом даже в минимальных количествах.
· 1,375 — минимальная активность. Либо работа с небольшой физической активностью, либо тренировки минимальной эффективности трижды в неделю.
· 1,55 — средняя активность. Работа со средней нагрузкой или спортивные тренировки трижды и чаще в неделю длительностью от 30 минут до часа.
· 1,7 — высокая активность. Тяжелый физический труд или ежедневные интенсивные тренировки.
· 1,9 – экстремально высокая активность. Профессиональные спортсмены, рабочие, занимающиеся тяжелым физическим трудом высокой интенсивности.
Мужчины:
· (10 x масса человека (кг) + 6.25 x рост (см) – 5 x возраст (г) + 5) x A;
Женщины:
· (10 x вес (кг) + 6.25 x рост (см) – 5 x возраст (г) — 161) x A.
Для расчета вес в килограммах умножается на 10, к нему прибавляется рост в сантиметрах, умноженный на 6.25, и из суммы вычитается возраст в годах, умноженный на 5. Результат умножаем на заранее определенный коэффициент активности.
Единственный недостаток такой схемы – некоторая узость группы, к которой его можно применить. По ней не рекомендуется рассчитывать рацион для людей младше 14 или старше 80 лет.
По ней не рекомендуется рассчитывать рацион для людей младше 14 или старше 80 лет.
Формула Харриса-Бенедикта
В своем изначальном виде метод расчета появился еще в 1919 году, однако поздней был существенно скорректирован и переработан. Он также опирается на коэффициент А и служит для расчета рациона по отдельности мужчин и женщин.
Мужчины:
· (88.362 + (13.397 x вес (кг)) + (4.799 x рост (см)) — (5.677 x возраст (г))) * А;
Женщины:
· (447.593 + (9.247 x вес (кг)) + (3.098 x рост (см)) — (4.330 x возраст (г))) * А.
После того, как необходимое количество калорий определено, можно переходить к расчету БЖУ.
Какой Должен Быть Баланс Бжу В Процентах
По формуле сбалансированного питания процентное соотношение белков, жиров, углеводов должно быть следующим: 14%: 30%: 56%. Mar 23, 2015 — Считается за норму соотношение БЖУ 1:1:4 соответственно. Тела, то вы должны уметь верно рассчитать свое БЖУ с учетом тренировок и вашего веса, в противном случае все ваши старания могут быть напрасными. В процентах от калорийности 1320 ккал/сут БЖУ 149/36/100. May 31, 2017 — Соотношение БЖУ в процентах будет выглядеть так: 16,6, 16,6 и 66,6%. Примерно 70% жиров должны быть природными (например,. Баланс БЖУ — это правильное соотношение белков, жиров и углеводов в питании. Для здорового человека оптимальным считается соотношение 1:1:4. Не забываем, что это соотношение в калориях, а не в грамма.
Баланс БЖУ — это правильное соотношение белков, жиров и углеводов в питании. Для здорового человека оптимальным считается соотношение 1:1:4. Не забываем, что это соотношение в калориях, а не в граммах. 1 г белка — это 4 ккал, 1 г углевода — тоже 1 ккал, а вот 1 г жира — это 9 ккал. Каким витамином богат лимон. Поэтому при соотношении белков и жиров 1:1 белков будет в граммах больше приблизительно в 2 раза, чем жиров. Если придерживаться соотношения БЖУ 1:1:4 и при этом не выходить за калорийность, необходимую для поддержания текущего веса, векс расти не будет, но и уменьшаться тоже. Салат мимоза со сливочным маслом слои. Если нужно похудеть, то есть использовать как энергию имеющийся в организме жир, предлагаются другие соотношения.
Одно из самых популярных, известное как Zone Diet 30:30:40. Для более быстрогго сброса веса, а так же для тех, кто занимается спортом, есть и такое соотношение 40:20:40, то есть больше калорий из белка, меньше калорий из жира. Соотношение 35:20:45 тоже поможет похудеть, не потеряв большое количество мышечной массы.
Не забываем, что для того, чтобы потерять лишний жир, нужно уменьшить калорийность рациона, создав в нем дефицит калорий. В среднем такой дефицит составляет 500 ккал. Для спортсменов — бодибилдеров популярно соотношение 50:20:30 — большой перевес в сторону белка. Учитывая то, что калорийность рациона спортсменов высока, съесть столько белка (половина калорийности) из обычной пищи проблематично.
Поэтому недостающий белок спортмены добирают белковыми коктейлями (протеинами). Это обеспечивает им необходимый баланс БЖУ.
БЖУ: нормы здорового питания В поисках точной информации о дневной калорийности питания и цифрах БЖУ, необходимых мужчинам и женщинам для похудения или для набора мышечной массы, легко столкнуться с большим количеством разнящихся мнений. Многие сайты предлагают свои рекомендации или даже готовые калькуляторы для расчета суточных норм, совершенно не вдаваясь в детали и не объясняя, откуда берутся цифры. Ситуация усложняется и тем, что универсальных норм питания просто не существует — все зависит от конкретного человека, его целей и уровня физической активности. В данном материале мы суммировали последнюю научную информацию и представили не просто общие данные, а подробные таблицы по нормам употребления белков, жиров и углеводов с учетом пола человека и веса его тела. Что такое калории и как их правильно считать?
Существуют ли программы для определения? Суточная норма углеводов Несмотря на то, что и строгое ограничение углеводов в рационе — один из наиболее эффективных способов быстрого похудения и снижения веса, подобные диеты нельзя считать «нормальным» режимом повседневного питания. Для оптимального здоровья необходимо не просто максимально урезать суточную норму потребления углеводов, а делить их на «плохие» и «хорошие». Секрет состоит в том, что углеводы с высоким (сахар, белая мука и прочие «пустые калории») приводят к набору лишнего жира — в отличии от углеводов с низким ГИ (различные овощи и цельнозерновые крупы). Помимо всего прочего, продукты с низким гликемическим индексом обычно содержат клетчатку, помогающую не только оптимизировать пищеварение, но и худеть быстрее.
Нормы употребления углеводов в день: Норма при весе 50 кг Норма при весе 60 кг Норма при весе 70 кг Норма при весе 80 кг Мужчины Для похудения 160 г 165 г 175 г 185 г Для поддержания веса 215 г 230 г 250 г 260 г Для набора мышц 275 г 290 г 300 г 320 г Женщины Для похудения 120 г 150 г 170 г 150 г Для поддержания веса 150 г 190 г 200 г 220 г Для набора мышц 200 г 245 г 260 г 240 г. Суточная норма белка Мнение, что белок должен являться основной рациона для роста мышц ошибочно — повышенная калорийность питания и достаточное количество правильных углеводов намного важнее для набора массы. Последние научные исследования и рекомендации говорят о суточной норме 1.5-2.5 г белка на килограмм сухого веса тела для спортсменов, тренирующихся с целью увеличения мышц (1). При этом несмотря на то, что превышение вышеобозначенной дневной нормы потребления белка не приводит к добавочному эффекту роста мышц, оно может быть весьма полезно для похудения или сжигания жира — FitSeven уже писал о том, что в отличии от калорий углеводов и жиров, организму существенно сложнее. Нормы употребления белков в день: Норма при весе 50 кг Норма при весе 60 кг Норма при весе 70 кг Норма при весе 80 кг Мужчины Для похудения 165 г 170 г 175 г 185 г Для поддержания веса 145 г 155 г 165 г 175 г Для набора мышц 180 г 190 г 200 г 210 г Женщины Для похудения 140 г 150 г 165 г 175 г Для поддержания веса 115 г 125 г 135 г 145 г Для набора мышц 155 г 165 г 175 г 185 г Суточная норма жиров Необходимо помнить о том, что жиры являются важнейшим элементом здорового питания человека, без которого невозможен нормальный метаболизм и обмен веществ. Роль играет и то, что вреден как переизбыток жиров в рационе, приводящий к набору лишнего веса, так и их недостаток, снижающий не только выработку гормонов, но и иммунитет. При этом особую важность имеют.
Дробное питание: что такое, и как считать БЖУ
Марина Николаева |
Дробное питание — способ по-новому взглянуть на свой рацион и привычки. Это не диета и не лечебная программа. Это возможность не срываться на внезапные перекусы сладким, ведь вы практически не испытываете чувство голода, а также осознанно выбирать продукты, контролировать сколько и что едите, придерживаясь нормы калорий и макронутриентов.
Дробное питание избавляет от проблемы переедания. Подумайте, насколько часто вам не удается нормально позавтракать и пообедать, поэтому вся калорийность приходится на ужин, когда наконец-то есть время поесть. С небольшими порциями, распределенными в течение дня, такой проблемы не возникает.
Главное, что питание небольшими порциями практически не меняет привычный рацион. Вы создаете расписание, которое приучает организм работать, как часы.
Набор здорового питания на весь день сервиса SoloFood.ruЧто такое дробное питаниеГлавный принцип дробного питания — есть часто, но маленькими порциями. Как правило, получается 5–6 приемов пищи каждые 3–4 часа. Неважно, когда вы просыпаетесь, просто поделите день на равные промежутки времени.
Расписание на день может быть таким:
- 08:00 — Завтрак
- 11:00 — Второй завтрак
- 14:00 — Обед
- 17:00 — Полдник
- 20:00 — Легкий ужин
- 23:00 — Кефир или травяной чай
Общее количество калорий, которое вам необходимо в день, равномерно делится между приемами пищи. Например, суточная норма в 2000 ккал разбивается на шесть перекусов по 330 ккал, а не на три приема пищи по 660 ккал.
Такое дробление помогает контролировать калорийность рациона, ведь небольшой объем легче просчитать, а также приучает к маленьким порциям. В результате, можно плавно снизить суточную калорийность, что способствует похудению.
При условно трехразовом питании, мы тоже делаем перекусы. Но чаще всего это быстрые углеводы из печенья, шоколада или энергетических напитков, которые только прибавляют калорийность рациону, но не приносят пользу.
При дробном питании на первую половину дня следует планировать более калорийные продукты. Исследования показывают, что у тех, кто привык плотно завтракать, уровень глюкозы в крови в среднем ниже, чем у тех, кто привык плотно обедать. Поэтому в первый и второй завтрак необходимо включать медленные углеводы и полезные жиры.
Во второй половине дня выбирайте овощи, несладкие фрукты и легкие молочные продукты.
Кому подходит дробное питаниеДробное питание подходит практически всем, только если нет особых противопоказаний по состоянию здоровья.
При проблемах с желудочно-кишечным трактом питание небольшими порциями даже рекомендовано — снижается нагрузка на пищеварительную систему, еда быстрее переваривается и не оставляет чувство тяжести.
Для тех, кто ведет активный образ жизни и часто тренируется, дробное питание легко соотносится с режимом дня. Проще подобрать продукты до и после тренировки, а также подготовить организм к нагрузкам. Об основах фитнес-питания рассказываем в статье «Как пользоваться спортивной программой питания».
Плюсы дробного питания
Главные правила дробного питания:
- Составить расписание на 5–6 приемов пищи.
- Соблюдать примерно одинаковое количество калорий в каждый прием.
- Придерживаться размера порции равной ладони.
- Не допускать перерывов в еде более 4 часов.
Главное преимущество дробного питания — ощущение постоянной сытости. Вы создаете суточный режим, где нет места перееданию. А питание по режиму, как и режим сна, благоприятно воздействует на здоровье.
При дробном питании в организм поступают небольшие порции пищи. Пищеварительная система не перегружается и еда полностью переваривается, оставляя ощущение легкости. Кроме того, питательные вещества и витамины усваиваются быстрее.
Стабильный уровень сахараЧерез 3 часа после еды уровень сахара в крови снижается. Через 4 часа переваривается все, что вы съели до этого, а через 5 часов уровень сахара совсем опускается и вы готовы съесть что угодно. Если начинаете день с завтрака, а затем продолжаете принимать пищу каждые 3–4 часа, то обеспечиваете организм и мозг постоянным потоком питательных веществ.
Длительные перерывы между едой вызывают резкие скачки уровня сахара в крови. Это приводит к спонтанным перекусам сладким или фастфудом, перееданию и набору веса.
Питание небольшими порциями поддерживает уровень сахара на одном уровне, организм наполнен энергией и вы не успеваете проголодаться так, чтобы срочно перекусить печеньем, чипсами или шоколадкой.
Ощущение сытости и подъем энергииДробное питание приучает к маленьким порциям и помогает плавно снизить суточную калорийность. В результате вы худеете, не испытывая дискомфорта.
Чем больше времени проходит между приемами пищи, тем активнее вырабатывается гормон грелин и мы испытываем голод. Грелин отвечает за жировые отложения, поэтому чем дольше длится чувство голода, тем больше вырабатывается ферментов, которые способствуют отложению жира и мешают его сжиганию.
Второй аспект голода — переедание. Как только мы добрались до еды, то готовы съесть все и даже попросить добавки. Ведь мозг не сразу осознает сытость.
При дробном питании чувство голода не успевает возникнуть. Это означает, что вероятность переесть снижается до минимума. Ощущая постоянную сытость, мы бодры. И, главное, после небольших порций еды не тянет в сон, как после плотного обеда.
В статье «Интервальное голодание: почему это вредно» подробнее рассказываем про опасность длительного голода.
Дробное питание помогает плавно снизить калорийность рационаи худеть, не испытывая дискомфортаСнижение веса
Между количеством приемов пищи и весом существует прямая зависимость. Как показывают исследования, люди с избыточным весом часто пропускают приемы пищи, а с нормальным питаются 3–4 раза в день.
Но влияет ли дробное питание на скорейшее снижение веса? В течение шести месяцев ученые сравнивали вес людей в двух группах. Одна группа питалась три раза в день, вторая шесть раз, съедая по 100 ккал каждые 2–3 часа. Участникам каждой группы установили индивидуальный лимит по калориям, с учетом их желания снизить вес. В результате, участники обеих групп снизили вес. Разница в частоте питания не повлияла на цифры, но участники, которые питались чаще, отметили что практически не испытывали чувство голода.
Такой же результат показывают исследования Кембриджского университета и Университета Оттавы — частое питание небольшими порциями не ускоряет метаболизм и не способствует скорейшему снижению веса. В вопросе похудения решающую роль играет общая калорийность рациона. Но дробное питание помогает контролировать калорийность и плавно приучать себя к меньшим порциям. В результате, вы снижаете суточную калорийность рациона и худеете, не испытывая дискомфорта. В статье «Похудение без спорта: будет ли результат» мы собрали советы, как кроме уменьшения калорийности, можно успешно снижать вес.
Какие недостатки у дробного питанияПри переходе на дробное питание необходимо четко ответить себе на три вопроса: что есть, когда есть и сколько есть. Поэтому основной недостаток дробного питания — сложность реализации.
Потребуется изменить привычное расписание, составить верное меню на день с учетом калорийности, а также быть готовым к тому, что в течение рабочего дня придется отвлекаться на 2–3 перекуса. Причем, это будут не привычные шоколадки или печенье, а небольшие блюда или фрукты и овощи. При дробном питании важно соблюдать питьевой режим, к чему также бывает сложно привыкнуть.
Планирование меню на неделю вперед помогает решить задачу. Вы выбираете, что взять на основные приемы пищи, а что останется на перекусы. Для этого необходимо рассчитать ежедневную калорийность рациона и норму белков, жиров и углеводов.
Диетологи сервиса доставки сбалансированного питания SOLO помогают составить программу дробного питания с учетом ваших пожеланий — сбросить или набрать вес или оставить все, как есть.
На чувство сытости, мозговую активность, физическое здоровье и гормональный баланс влияет сбалансированное поступление макронутриентов — белков, жиров и углеводовЗачем соблюдать баланс белков, жиров и углеводовКлючевой вопрос в дробном питании — что есть и как узнать калорийность каждого приема пищи. Калорийность продуктов может быть одинакова, но влияние на организм различается. На чувство сытости, мозговую активность, физическое здоровье и гормональный баланс влияет сбалансированное поступление макронутриентов — белков, жиров и углеводов (БЖУ).
Приведем пример:
Большая тарелка брокколи (340 г) содержит 100 ккал и дает нам 8 г клетчатки. Половина пончика с глазурью также содержит 100 ккал, которые получаются из насыщенных жиров и простых углеводов. Энергетическая ценность одинаковая, а влияние на организм разное. Съев в один прием пищи тарелку брокколи, вы насытитесь на несколько часов и получите порцию клетчатки. Половинка пончика лишь на короткое время утолит голод и, вероятнее всего, вы скоро доедите и вторую.
Поэтому каждая небольшая порция еды должна содержать полезную комбинацию макронутриентов: белок из постного мяса или молочных продуктов, сложные углеводы из круп, бобовых или овощей, а также полезные жиры из растительных масел или жирной рыбы. Избегайте простых углеводов, переработанных продуктов и вредных жиров. Они быстро утоляют голод, но не приносят пользы организму.
Каждый из макронутриентов выполняет свою функцию в организме:
- Белок дает чувство сытости и избавляет от потери мышечной массы. Мясо, рыба, птица, яйца и молочные продукты — источники животного белка. Растительный белок содержится в сое, зерновых и бобовых.
- Углеводы — главный источник энергии для организма. Углеводы важны для правильной работы мозга и влияют на наше настроение, память и даже принятие решений.
Выбирайте сложные углеводы, которые медленно усваиваются, поддерживают чувство сытости и не приводят к скачкам уровня сахара в крови.
Полезные углеводы содержатся в цельных зернах, крупах, бобовых, овощах и фруктах.
- Жиры — основной источник калорий и строительный материал для клеточных мембран и нервных оболочек. Жиры помогают усваивать витамины, регулируют термообмен, поддерживают баланс гормонов и сохраняют репродуктивное здоровье.
К хорошим жирам относятся мононенасыщенные и полиненасыщенные, которые содержатся в растительных маслах, орехах, авокадо, жирной рыбе, семенах чиа.
«Вредными» жирами считаются насыщенные жирные кислоты и трансжиры. Подробнее о жирах читайте в нашей «Польза жиров для здоровья».
В каждый прием пищи комбинируйте продукты: молочные с фруктами, бобовые с цельными зернами, овощи с постным мясом или рыбой. Пробуйте новые сочетания, чтобы сделать меню разнообразным. Например, омлет со шпинатом и кальмаром, кинотто с морепродуктами или лосось с киноа и зелеными бобами.
Сбалансированный рацион обеспечивает организм необходимыми веществами для правильной работы. Соблюдая баланс, вы не страдаете от нехватки углеводов и не становитесь слабым из-за белкового голодания. Кроме того, изменяя соотношение БЖУ можно быстрее сбросить вес или нарастить мышечную массу.
Как рассчитать суточную норму калорий и макронутриентовСуточная норма калорий показывает какое количество энергии требуется организму для поддержания обмена веществ и правильного функционирования.
Калорийность и норма БЖУ зависят от личных параметров и уровня активности. Для сбалансированного питания и поддержания веса рекомендуется соотношение: 20% белков, 30% жиров и 50% углеводов.
Процентное соотношение может варьироваться в зависимости от ваших целей. Для тех, кто хочет похудеть компания SOLO предлагает программу Solo Slim с таким распределением нутриентов: 30% белков, 30% жиров и 40% углеводов, при этом решающую роль играет общая калорийность — от 850 до 1600 ккал в сутки.
Попробуйте рассчитать свою норму калорий и БЖУ за три шага:
Шаг 1Узнайте показатель Основного обмена веществ или базального метаболизма, basal metabolic rate (BMR). Результат покажет сколько калорий организм тратит в состоянии покоя для поддержания процессов жизнедеятельности. Показатель зависит от возраста, роста и веса, поэтому для каждого будет свой.
Для расчета BMR воспользуйтесь онлайн калькулятором или формулой Миффлина-Джеора:
- Для мужчин: BMR = 5 + (10 × вес в кг) + (6,25 × рост в см) − (5 × возраст в годах).
- Для женщин: BMR = (10 × вес в кг) + (6,25 × рост в см) − (5 × возраст в годах) − 161.
Полученный результат — это количество калорий, которое вам необходимо ежедневно потреблять, чтобы не возникло проблем со здоровьем.
Шаг 2Дневная норма калорий зависит от уровня физической активности. Умножьте ваш показатель на один из коэффициентов активности, которые разработала Всемирная организация здравоохранения:
- 1,53 — сидячий образ жизни без занятий спортом.
- 1,76 — умеренно активный образ жизни с легкими нагрузками и прогулками.
- 2,25 — активный образ жизни: спортсмены и люди с интенсивной физической нагрузкой не менее двух часов в день.
Полученный результат — ваша дневная норма калорий.
Теперь определитесь с целью и скорректируйте калорийность рациона:
- Поддерживать вес — количество калорий равно получившемуся результату.
- Снизить вес — создайте дефицит и уменьшите полученный результат на 10–30%.
- Набрать вес — добавьте к результату 10–30%.
Определите дневную норму БЖУ, распределив калорийность в соответствии с вашим планом питания. Затем переведите процентное соотношение БЖУ в граммы, учитывая содержание килокалорий в 1 грамме нутриента.
| Макронутриент | Энергетическая ценность на один грамм | Норма в сутки на 1 кг веса |
| Белки | 4 ккал | 1,2–2 г |
| Жиры | 9 ккал | 0,8–1,1 г |
| Углеводы | 4 ккал | 2–6 г |
Пример расчета
Ваша суточная норма: 1500 ккал
Соотношение БЖУ: 30% белков, 30% жиров и 40% углеводов
Переведем эти данные в калории и граммы:
40% углеводов = 600 ккал = 150 г
30% белков = 450 ккал = 112 г
30% жиров = 450 ккал = 50 г
Калорийность и содержание микронутриентов в продуктах проверяйте в специальных таблицах, их множество в интернете, или читайте данные на упаковках. Например, 100 г куриной грудки содержит 31 г белка, а в чашке приготовленного киноа — 8 г. В одном авокадо 15 г жиров, а в одной чашке риса около 54 г углеводов.
Распределяйте БЖУ в зависимости от образа жизни. Если ведете активные тренировки, то потребуется увеличить долю белка и снизить жиры. В службе здорового питания SOLO мы предлагаем особую программу Solo Prive Sport с шестиразовым питанием для тех, кто тренируется не менее шести часов в неделю. Чередование белковых и углеводных дней позволяет эффективно сжигать жир и укрепляет мышцы.
Как перейти на дробное питаниеОсобой подготовки не требуется. Когда вы знаете суточную норму калорий и БЖУ, то можно смело переходить на дробное питание. Сохраняйте три основных приема пищи и добавьте 2–3 правильных перекуса.
Несколько советов, как сделать переход более комфортным:
- Выбирайте маленькие тарелки. Первое время так будет спокойнее воспринимать размер порций. На маленькой тарелке объем еды кажется больше.
- Постепенно изменяйте размер порций. Если привыкли есть 1–2 раза в день большими порциями, то уменьшите сначала наполовину. В результате придете к тому, что порция на один прием будет равняться ладони, а объем жидкости — стакану.
- Запоминайте порции на глаз. Положите в измерительный стакан порцию овощей, риса или творога. Затем переложите на пустую тарелку. Так вы визуально запомните объем.
- В кафе или ресторане заказывайте половину порции.
- Поешьте, если очень хочется. Лучше уменьшите порцию вдвое, но не терпите сильный голод.
- Берите на работу порционные контейнеры. Так вы не собьетесь с графика и всегда будете знать, чем перекусить.
- Купите на выходных фрукты и овощи, йогурт, полезные батончики-мюсли, орехи и крупы.
Дробное питание позволяет соединять в небольших порциях разные продукты и не чувствовать тяжести после еды. Вы удивитесь, насколько неожиданными могут быть сочетания и разнообразным ваше питание.
При составлении меню следуйте правилам:
- В первой половине дня отдавайте предпочтение углеводам и жирам: каша с фруктами, мед, цельнозерновой хлеб и масло.
- На обед и ужин сочетайте белковые продукты с некрахмалистыми овощами или крупами: супы-пюре, рыба или птица с булгуром, котлетки с рататуем. Здесь есть одно правило: если вы стремитесь снизить вес, то исключите крупы на обед. Если хотите сохранить вес или крайне активны в течение дня, то оставляйте.
- Для второго завтрака и полдника подходит сочетание белка с клетчаткой: фрукты с творожным сыром, цельнозерновые хлебцы с чайной ложкой урбеча, сырые овощи с хумусом или дзадзики.
Дробный рацион подходит всем, кто стремится к здоровому и сбалансированному питанию. В небольших порциях все на своих местах: белки, полезные жиры и медленные углеводы, которые можно получить из орехов, фруктов, молочных продуктов, мяса и круп. Сезонные овощи и фрукты вносят разнообразие в рацион.
Также это палочка-выручалочка для тех, кто с трудом справляется с вредными перекусами днем и склонен наедаться на ночь.
Питание небольшими порциями учит есть осознанно, уделяя внимание качеству еды и ее влиянию на организм.
Подпишитесь на рассылку
БЖУ в цифрах: подробные таблицы для здорового питания: aleks070565 — LiveJournal
Дневные нормы употребления углеводов, белков и жиров — ключевой момент многих научных исследований. В поисках точной информации о суточной калорийности питания и нормах БЖУ, необходимых для похудения или для набора мышечной массы, диетологи совершили множество экспериментов. Однако выяснилось, что единых и однозначных норм и рекомендаций употребления углеводов просто не существует. Несмотря на то, что многие сайты предлагают свои рекомендации или даже готовые калькуляторы для расчета суточных норм, зачастую цифры определяются ошибочно.
Универсальных суточных норм питания в граммах просто невозможно установить — в конечном итоге, все зависит как от гликемического индекса съедаемой еды, так и от метаболизма человека, его спортивной подготовки и уровня физической активности. Помимо всего прочего, таблицы калорийности продуктов также подразумевают погрешность в 10-30% от указываемых цифр — что ставит под вопрос всю теорию подсчета калорий.
Ситуация усложняется и тем, что универсальных норм питания просто не существует — все зависит от конкретного человека, его целей и уровня физической активности. В данном материале просуммирована последняя научная информация и представлены не просто общие данные, а подробные таблицы по нормам употребления белков, жиров и углеводов с учетом пола человека и веса его тела.
Суточная норма углеводов
Несмотря на то, что безуглеводная диета и строгое ограничение углеводов в рационе — один из наиболее эффективных способов быстрого похудения и снижения веса, подобные диеты нельзя считать «нормальным» режимом повседневного питания. Для оптимального здоровья необходимо не просто максимально урезать суточную норму потребления углеводов, а делить их на «плохие» и «хорошие».
Секрет состоит в том, что углеводы с высоким гликемическим индексом (сахар, белая мука и прочие «пустые калории») приводят к набору лишнего жира — в отличии от углеводов с низким ГИ (различные овощи и цельнозерновые крупы). Помимо всего прочего, продукты с низким гликемическим индексом обычно содержат клетчатку, помогающую не только оптимизировать пищеварение, но и худеть быстрее.
Нормы употребления углеводов в день:
| Норма при весе 50 кг | Норма при весе 60 кг | Норма при весе 70 кг | Норма при весе 80 кг | |
| Мужчины | ||||
| Для похудения | 160 г | 165 г | 175 г | 185 г |
| Для поддержания веса | 215 г | 230 г | 250 г | 260 г |
| Для набора мышц | 275 г | 290 г | 300 г | 320 г |
| Женщины | ||||
| Для похудения | 120 г | 150 г | 170 г | 150 г |
| Для поддержания веса | 150 г | 190 г | 200 г | 220 г |
| Для набора мышц | 200 г | 245 г | 260 г | 240 г |
Суточная норма белка
Мнение, что белок должен являться основной рациона для роста мышц ошибочно — повышенная калорийность питания и достаточное количество правильных углеводов намного важнее для набора массы. Последние научные исследования и рекомендации говорят о суточной норме 1.5-2.5 г белка на килограмм сухого веса тела для спортсменов, тренирующихся с целью увеличения мышц.
При этом несмотря на то, что превышение вышеобозначенной дневной нормы потребления белка не приводит к добавочному эффекту роста мышц, оно может быть весьма полезно для похудения или сжигания жира — в отличии от калорий углеводов и жиров, организму существенно сложнее конвертировать калории протеинов в жировые запасы.
Нормы употребления белков в день:
| Норма при весе 50 кг | Норма при весе 60 кг | Норма при весе 70 кг | Норма при весе 80 кг | |
| Мужчины | ||||
| Для похудения | 165 г | 170 г | 175 г | 185 г |
| Для поддержания веса | 145 г | 155 г | 165 г | 175 г |
| Для набора мышц | 180 г | 190 г | 200 г | 210 г |
| Женщины | ||||
| Для похудения | 140 г | 150 г | 165 г | 175 г |
| Для поддержания веса | 115 г | 125 г | 135 г | 145 г |
| Для набора мышц | 155 г | 165 г | 175 г | 185 г |
Суточная норма жиров
Необходимо помнить о том, что жиры являются важнейшим элементом здорового питания человека, без которого невозможен нормальный метаболизм и обмен веществ. Роль играет и то, что вреден как переизбыток жиров в рационе, приводящий к набору лишнего веса, так и их недостаток, снижающий не только выработку гормонов, но и иммунитет. При этом особую важность имеют Омега-3 жирные кислоты.
Интересно и то, что наиболее полезные для здоровья и поддержания веса режимы питания — традиционные Палеодиета и Средиземноморская диета — подразумевают долю достаточно весомую долю жира в рационе (порядка 35-50% от всех дневных калорий). Однако нужно помнить о том, что источником этих жиров должны быть преимущественно растительные масла, а не животный жир.
Нормы употребления жиров в день:
| Норма при весе 50 кг | Норма при весе 60 кг | Норма при весе 70 кг | Норма при весе 80 кг | |
| Мужчины | ||||
| Для похудения | 40 г | 40 г | 40 г | 40 г |
| Для поддержания веса | 55 г | 60 г | 60 г | 65 г |
| Для набора мышц | 70 г | 70 г | 75 г | 80 г |
| Женщины | ||||
| Для похудения | 30 г | 35 г | 35 г | 40 г |
| Для поддержания веса | 45 г | 50 г | 50 г | 55 г |
| Для набора мышц | 60 г | 60 г | 65 г | 70 г |
Определение нормы калорий
С одной стороны, определение дневной потребности в калориях — основа составления режима питания для похудения или для набора мышечной массы. С другой стороны, рассчитанная даже по самой точной формуле цифра будет сильно приблизительной, поскольку в любой метод математического определения суточной нормы калорий заложена существенная погрешность в 300-500 ккал.
Роль играет и то, что потребности реального человека в суточных калориях всегда варьируется ото дня к дню, поскольку организм всегда подстраиваться под различные внешние факторы, замедляя или ускоряя обмен веществ. По сути, в большинстве случаев для определения примерной нормы калорий достаточно умножить вес тела в килограммах на коэффициент 35 (т.е. 2625 ккал для 75 кг).
Таблица БЖУ: Нормы для похудения
| Суточные нормы для похудения | Процентное соотношение | Расчет для мужчины весом 75 кг и 20% жира |
| Калории | Минус 20% от нормы | 2000 ккал |
| Белки | 25-35% | 125 – 175 г / 500-700 ккал |
| Жиры | 20-40% | 45 – 90 г / 400 – 800 ккал |
| Углеводы | 25-50% | 125 – 250 г / 500 – 1000 ккал |
Напомним, что для эффективного похудения необходимо сокращение суточной нормы калорий примерно на 15-20%. В расчетах важно учитывать то, что дневная норма калорий рассчитывается лишь по сухому весу тела, то есть без учета жировой массы. Доля жиров в рационе — 20-40% от всех калорий, предпочтение необходимо отдавать растительным, а не животным жирам.
Важно и то, что рекомендуемая диетологами дневная норма углеводов для похудения во многом зависит от типа телосложения человека и его пола. Мужчинам с крупной костью для похудения необходимо минимум углеводов с низким гликемическим индексом, тогда как женщины с быстрым метаболизмом могут есть больше углеводов, тратя энергию на жиросжигающие тренировки.
Таблица БЖУ: Нормы для набора массы
| Суточные нормы для набора мышц | Процентное соотношение | Расчет для мужчины весом 70 кг и 10% жира |
| Калории | Плюс 20% к норме | 3000 ккал |
| Белки | 20-25% | 150 – 190 г / 600 – 750 ккал |
| Жиры | 30-35% | 100 – 115 г / 900 – 1050 ккал |
| Углеводы | 50-60% | 375 – 450 г / 1500 – 1800 ккал |
По сути, при набора мышечной массы питание играет более значимую роль, чем сами силовые тренировки. Для роста количество суточных калорий должно быть увеличено примерно на 15-20%. Углеводы должны употребляться по максимальной границе нормы, однако белки и жиры — в умеренном количестве (в противном случае организм может снизить выработку тестостерона).
Отметим и то, что важным является время употребления нутриентов — например, значительная часть углеводов должна съедаться в период «углеводного окна», длящийся 2-3 часа после силовой тренировки). В этом случае организм будет запасать углеводы в виде гликогена в мышцах (в буквальном смысле увеличивая их визуальный объем), а вовсе не в жировых запасах.
***
Суточные нормы углеводов, белков и жиров: резюме
- Потребности конкретного человека в калориях варьируется ото дня к дню, поскольку организм всегда подстраиваться под различные внешние факторы, замедляя или ускоряя обмен веществ.
- При определении суточной потребности в белках, жирах и углеводах необходимо учитывать не только основную цель диеты (поддержание веса, похудение или набор мышц), но и делать поправку на возраст и общий уровень активности человека.
- Онлайн калькуляторы калорий с большой долей вероятности покажут лишь случайную и ошибочную цифру, поскольку при расчете не учитывается множество вторичных факторов.
- Ключевую роль играют не просто нормы суточной нормы потребления нутриентов, а то, какие именно продукты питания за скрывается за рекомендованной цифрой. Как углеводы, так жиры, не одинаковы — одни из них полезны для здоровья, а другие вредны.
Что такое БЖУ и как его рассчитать
В полноценном питании, как и в жизни, важно соблюдать баланс. Для разных процессов, происходящих в нашем теле, нужны разные вещества. Нехватка одного из важных элементов моментально отразиться на вашем самочувствии и состоянии тела. Ломкие волосы и тонкие ногти, проблемы с кожей и пищеварением, воспаленные суставы и медленное восстановление после тренировок, судороги, плохая концентрация и многое другое — все это свидетельствует о том, что вам явно чего-то не хватает.
Для устранения дисбаланса важно понимать, как работает наш организм, и как именно должен выглядеть рацион среднестатистического человека. Если вы активно занимаетесь спортом, меню будет отличаться, особенно в дни тяжелых тренировок. Хороший тренер объяснит вам, что такое БЖУ и как его правильно рассчитать в зависимости от нагрузок.
Что такое БЖУ
БЖУ или БЖУК (белки, жиры, углеводы, калории) — это соотношение белков, жиров и углеводов в пределах вашего дневного рациона по калорийности. Он может отличаться в зависимости от ваших целей и особенностей организма (метаболизм, соотношение жировой ткани к мышечной).
Если вы хотите похудеть, то именно расчет сбалансированного питания поможет сделать это без вреда для здоровья.
Если вы хотите набрать мышечную массу, расчет баланса БЖУ приблизит вас к цели быстрее и без побочных эффектов, которых в этом процессе не меньше, чем в похудении. Кто хотя бы один раз пытался правильно набрать мышечную массу (акцент на слове «мышечная»), тот прекрасно понимает, что иногда сделать это даже сложнее, чем похудеть.
читайте такжеКак рассчитать БЖУ
Как мы уже говорили выше, БЖУ будет зависеть от ваших целей и особенностей функционирование организма. Если брать взрослого среднестатистического человека, то для сбалансированного дневного рациона соотношение БЖУ (формула сбалансированного питания) выглядит так: 1:1:4 — одна часть белков, одна часть жиров и четыре части углеводов. Как всегда, гарнира должно быть много.
Для спортсменов этот баланс будет выглядеть иначе — 1:0,8:4 или 1:0,7:4. В их рационе уменьшена часть жиров, так как во время высоких физических нагрузок может возникать кислородный голод, а на сжигание жиров требуется больше энергии, чем на преобразование в энергию углеводов.
Для расчета количества БЖУ в граммах можно пользоваться усредненными данными. Для взрослого человека со средним уровнем физической активности на один килограмм массы тела необходимо 1,2 граммов белка, 1 грамм жира и 2 грамма углеводов. Это количество можно корректировать в зависимости от целей и потребностей организма.
Если же говорить о процентном соотношении, то ваш рацион может выглядеть так, как на картинке ниже:
Для тех, кто серьезно занимается спортом, выведение баланса будет сложнее, так как в этом случае нужно учитывать свое телосложение (эктоморф, мезоморф и эндоморф), цель и физическую активность. Например, белок помогает восстановлению мышц, так как является строительным материалом. Поэтому при повышенных физических нагрузках нужно увеличиваться его порции: силовые тренировки — 1,2—1,6 г на килограмм веса, тренировки на выносливость — 1,4—1,8 г на килограмм веса.
Углеводы являются основным видом топлива для организма, и при повышенной физической активности их количество их потребления на один килограмм массы тела также растет — до 9 грамм при высоких нагрузках! На всякий случай напоминаем, что это эти граммы нужно измерять не тортиками и другими привлекательными быстрыми углеводами, а чем-то более полезным — цельнозерновыми крупами, фруктами и овощами.
Кроме определения соотношения БЖУ важно знать свою дневную норму калорий. Для этого можно воспользоваться формулами Харриса — Бенедикта или Миффлина — Сан Жеора.
Формула Харриса — Бенедикта:
Мужчины: 88,362 + (13,397 × вес [кг]) + (4,799 × рост [см]) − (5,677 × возраст [лет]).
Женщины: 447,593 + (9,247 × вес [кг]) + (3,098 × рост [см]) − (4,33 × возраст [лет]).
Формула Миффлина — Сан Жеора:
Мужчины: 5 + (10 × вес [кг]) + (6,25 × рост [см]) − (5 × возраст [лет]).
Женщины: (10 × вес [кг]) + (6,25 × рост [см]) − (5 × возраст [лет]) − 161.
Для облегчения страданий можно воспользоваться онлайновыми калькуляторами, а не высчитывать данные самостоятельно.
Калькуляторы: musclegain.ru, gym-training.com, goodlooker.ru, beregifiguru.ru, dailyfitnessblog.ru.
Подписывайся на нашу страничку в Instagram и не пропускай самые полезные видеоматериалы от Beauty HUB!
читайте такжеДневная норма белков, жиров и углеводов (БЖУ)
В предыдущей статье, мы рассчитали суточную норму калорий. Следующим шагом на пути к стройной фигуре является определение такого показателя, как дневная норма белков, жиров и углеводов (БЖУ). От баланса этих макронутриентов напрямую зависит наше состояние здоровья и внешний вид. Каждый продукт в нашем рационе имеет различное содержание БЖУ. Их грамотно подобранное соотношение является залогом правильного питания.
Раньше классической пропорцией было принято считать соотношение 1:1:4 (белки, жиры и углеводы соответственно). В энергетической ценности это можно выделить следующим образом:
✔️ 1 грамм белка = 4 ккал
✔️ 1 грамм жира = 9 ккал
✔️ 1 грамм углеводов = 4 ккал
Сегодня диетологи пришли к выводу, что не существует универсальной формулы, которая подходила бы без исключения каждому. Эти показатели должны постоянно корректироваться в зависимости от возраста, состояния здоровья и активности человека. Единственное остается неизменным – в рационе обязательно должны присутствовать все три компонента. Отказ от одного из них чреват большими проблемами со здоровьем.
Показатели БЖУ вместе с суточной нормой калорий являются основными критериями, на которых должно строится питание. Для начала, давайте вспомним биологические функции этих макронутриентов в организме человека. Их значение сложно переоценить!
Белки
Белки являются строительным материалом для нашего тела, активно участвуют в процессе метаболизма и поддерживают иммунитет. Их недостаток может привести к ослаблению защитной функции организма, отставанию в физическом развитии и проблемам с памятью. Опасен и переизбыток белка. Создается нагрузка на почки, страдает микрофлора кишечника, появляется раздражительность и депрессия.
Основным источником являются продукты животного происхождения.
Жиры
Жиры насыщают организм энергией, защищают внутренние органы от повреждений и участвуют в синтезе гормонов. Их нехватка сопровождается быстрой утомляемостью, приводит к заболеваниям кожи и проблемам со зрением. При избытке жиров повышается риск атеросклероза, повышается давление и возрастает нагрузка на печень, почки.
Преобладают в маслах растительного происхождения, орехах, рыбе и молочных продуктах.
Углеводы
Углеводы-главные поставщики энергии для физической и умственной деятельности нашего организма. Благодаря их работе происходит полноценное функционирование сердца, нервной системы и всех обменных процессов. Недостаток углеводов понижает функцию щитовидной железы, вызывает головные боли, уменьшает трудоспособность. Избыток может привести к ожирению, сахарному диабету и потере памяти.
Содержатся в большом количестве в корнеплодах, бобовых, крупах, овощах и хлебе грубого помола.
В зависимости от целей, пропорция БЖУ рассчитывается в процентном соотношении от суточной нормы калорий.
Благодаря индивидуальному расчету КБЖУ, вы обеспечите организм всем необходимым для правильного развития и функционирования. Обучаясь питаться грамотно, привыкая к осознанному потреблению мы совершаем большой вклад в наше здоровье и будущее.
Наш канал на ЯндексДзен.
487
Следует ли стоимостным инвесторам выбирать акции Beazer Homes (BZH)?
Стоимостное инвестирование — один из самых популярных способов найти отличные акции в любой рыночной среде. В конце концов, кто бы не захотел найти акции, которые либо незаметны и привлекают внимание, либо предлагают соблазнительные скидки по сравнению со справедливой стоимостью?
Один из способов найти эти компании — взглянуть на несколько ключевых показателей и финансовых коэффициентов, многие из которых имеют решающее значение в процессе выбора ценных бумаг.Давайте включим в это уравнение акции Beazer Homes USA, Inc. BZH и выясним, является ли это хорошим выбором для инвесторов, ориентированных на стоимость, прямо сейчас или инвесторам, подписавшимся на эту методологию, следует поискать лучшие предложения в другом месте:
PE Отношение
Ключевым показателем, на который всегда обращают внимание инвесторы, является отношение цены к прибыли, или сокращенно PE. Это показывает нам, сколько инвесторы готовы платить за каждый доллар прибыли от данной акции, и, безусловно, является одним из самых популярных финансовых коэффициентов в мире.Лучше всего использовать коэффициент PE, чтобы сравнить текущий коэффициент PE акции с: а) тем, где это соотношение было в прошлом; б) как он соотносится со средним показателем по отрасли / сектору; и c) как он соотносится с рынком в целом.
На этом фронте Beazer Homes имеет конечный коэффициент PE за двенадцать месяцев, равный 7,62, как вы можете видеть на диаграмме ниже:
Этот уровень на самом деле довольно выгодно отличается от рынка в целом, поскольку PE для S&P 500 сравнивается в около 18,54. Если мы сосредоточимся на долгосрочном тренде PE акций, текущий уровень ставит текущий коэффициент PE Beazer Homes немного ниже его средней точки (которая составляет 8.85) за последние пять лет.
Кроме того, PE акций выгодно отличается от показателя PE сектора Zacks Construction за двенадцать месяцев, который составляет 16,75. По крайней мере, это указывает на то, что акции сейчас относительно недооценены по сравнению с аналогами.
Мы также должны отметить, что у Beazer Homes форвардный коэффициент PE (цена относительно прибыли этого года) составляет 8,75, так что справедливо ожидать увеличения стоимости акций компании в ближайшем будущем.
Коэффициент P / S
Другой ключевой показатель, на который следует обратить внимание, — это соотношение цена / объем продаж. Этот подход сравнивает цену данной акции с ее общим объемом продаж, при этом более низкое значение обычно считается лучшим. Некоторым людям этот показатель нравится больше, чем другие, ориентированные на ценность, потому что он учитывает продажи, то есть чем-то, что гораздо труднее манипулировать бухгалтерскими уловками, чем прибылью.
Прямо сейчас у Beazer Homes соотношение P / S составляет около 0,21. Это значительно ниже, чем средний показатель S&P 500, равный 3.27 прямо сейчас. Кроме того, как мы видим на графике ниже, это несколько ниже максимумов для этой акции, в частности, за последние несколько лет.
История продолжается
Во всяком случае, это говорит о некотором уровне недооцененной торговли — по крайней мере, по сравнению с историческими нормами.
Общий прогноз стоимости
В целом, Beazer Homes в настоящее время имеет оценку качества стоимости B, что позволяет ему войти в топ 40% всех акций, которые мы рассматриваем в этом обзоре. Это делает BZH надежным выбором для стоимостных инвесторов, и некоторые другие его ключевые показатели также делают это довольно очевидным.
Например, коэффициент P / CF (еще один отличный показатель стоимости) составляет 6,11, что немного лучше, чем в среднем по отрасли 7,42. Ясно, что BZH — это надежный выбор на ценовом фронте с разных точек зрения.
Как насчет запасов в целом?
Хотя Beazer Homes может быть хорошим выбором для стоимостных инвесторов, есть много других факторов, которые следует учитывать, прежде чем инвестировать в это имя. В частности, стоит отметить, что компания имеет оценку роста F и оценку Momentum B.Это дает BZH оценку VGM — или ее всеобъемлющую фундаментальную оценку — D. (Подробнее о рейтингах стиля Закса можно узнать здесь >> )
Между тем, последние оценки прибыли компании в лучшем случае неоднозначны. В текущем году за последние шестьдесят дней одна оценка снизилась по сравнению с отсутствием более высокой, в то время как в оценке за весь год наблюдалась аналогичная тенденция в тот же период времени.
Это оказало заметное влияние на консенсус-оценку, поскольку консенсус-оценка за текущий квартал упала на 10.9% за последние два месяца, тогда как годовая оценка выросла на 6,3%. Вы можете увидеть тенденцию консенсус-оценки и недавнее движение цен для акций на графике ниже:
Beazer Homes USA, Inc. Цена и прогноз
Beazer Homes USA, Inc. Цена и прогноз
Beazer Homes USA , Inc. график консенсуса цен | Beazer Homes USA, Inc. Котировка
Эта смешанная тенденция объясняет, почему акции имеют только 3-й рейтинг Zacks («Держать»), и почему мы ждем от компании прямых результатов в ближайшем будущем.
Bottom Line
Beazer Homes — отличный выбор для стоимостных инвесторов, поскольку на этом фронте сложно превзойти его невероятную статистику. Более того, сильное положение в отрасли (12% из более чем 250 отраслей) дополнительно поддерживает потенциал роста акций. Тем не менее, имея 3-й ранг Zacks, эта компания в целом не вызывает особого восторга. Однако за последний год этот сектор явно отставал от рынка в целом, как вы можете видеть ниже:
Таким образом, стоимостные инвесторы могут сначала подождать, пока оценки, настроения аналитиков и отраслевые тенденции станут благоприятными в этом названии, но как только это произойдет, эта акция может стать отличным выбором.
7 лучших акций на следующие 30 дней
Только что выпущено: эксперты выделяют 7 элитных акций из текущего списка 220 Zacks, занимающих первое место среди сильных покупок. Они считают эти тикеры «наиболее вероятными для раннего скачка цен».
С 1988 г. полный список опередил рынок более чем в 2 раза со средним приростом + 24,6% в год. Так что не забудьте сразу обратить внимание на эти 7 отобранных вручную.
Посмотреть их сейчас >>
Хотите получить последние рекомендации от Zacks Investment Research? Сегодня вы можете скачать 7 лучших акций на следующие 30 дней.Щелкните, чтобы получить этот бесплатный отчет
Beazer Homes USA, Inc. (BZH): Бесплатный отчет по анализу запасов
Чтобы прочитать эту статью на Zacks.com, щелкните здесь.
Zacks Investment Research
ai-optim / lab3.md at master · brain-bzh / ai-optim · GitHub
Lab Session 3
Целью этой третьей лабораторной сессии является выполнение экспериментов с использованием методов обрезки.
Часть 1
Pytorch предоставляет библиотеку, предназначенную для облегчения сокращения нейронных сетей: Учебное пособие по сокращению.Обратите внимание на разницу между функциями сокращения (например, prune.random_unstructured ) и функцией prune.remove .
Например, когда он применяется к весам, применение функций отсечения к модулю создает дубликат исходных весов ( weight_orig ) и связанную маску ( weight_mask ). Это меняет структуру модуля.
weight_orig становится torch.nn.parameter.Parameter, и поэтому, если вы тренируете модель, именно этот тензор будет изменен. weight — это простой атрибут, который вычисляется во время прямого прохода путем применения weight_mask к weight_orig .
Если вы хотите навсегда применить обрезку и вернуться к исходной структуре вашей модели, вы должны применить prune.remove к модулю. Он воссоздает вес в качестве параметра с содержанием weight_orig для неотрезанных весов и нулей для усеченных весов.
В примере из Руководства по сокращению рассматривается очень простая сеть.Ваш будет более сложным. Первым шагом должно быть извлечение модулей, которые нужно обрезать, чтобы их обрезать. Выполните итерации по (torch.nn.Module.modules) [https://pytorch.org/docs/stable/generated/torch.nn.Module.html#torch.nn.Module.modules], чтобы извлечь сверточные и линейные слои. Затем примените обрезку.
Цель сегодняшнего сеанса — применить полученные ранее знания для реализации метода отсечения. Вы можете выбрать любой из методов, которые мы изучили в курсе 3, но, вероятно, следующие четыре являются наиболее простыми для реализации:
- Global Pruning, без переобучения: просто удалите веса с наименьшей нормой l1, измерьте точность для различных соотношений обрезки
- Изучение весов и соединений для эффективных нейронных сетей: применить повторное обучение после первого глобального сокращения
- Фильтры обрезки для эффективных ConvNets: разные соотношения обрезки на разных слоях
- ThiNet: то же, но на основе норм карт характеристик
Есть несколько способов обрезки, будьте новаторски! Разные соотношения на разных слоях, разные критерии обрезки, разные способы тонкой настройки… Играть !
Часть 2 — Объединение всех методов на MiniCIFAR, CIFAR10 и CIFAR100
Теперь ваша очередь объединить все, что мы видели до сих пор, чтобы начать выполнение некоторых интересных сравнений с использованием трех наборов данных MiniCIFAR, CIFAR10 и CIFAR100.
Рассмотрите различные факторы, которые могут повлиять на общий объем памяти, необходимый для хранения сетевых параметров, а также карт функций / активаций.
Ключевой вопрос, который нас интересует:
Какая максимальная достижимая точность при минимальном объеме памяти?
Подготовьте презентацию для занятия 4, подробно описав вашу методологию и исследования для ответа на этот вопрос.У вас будет 10 минут на выступление, а затем 5 минут на вопросы. Удачи !
Beazer Homes USA Inc (BZH) Стенограмма телефонного звонка за 2 квартал 2019 года
Источник изображения: Пестрый дурак.
Beazer Homes USA Inc (NYSE: BZH)
Отчет о прибылях и убытках за 2 квартал 2019 г.
May. 02 января 2019 г., 17:00 ET
В комплекте:
- Подготовленные примечания
- Вопросы и ответы
- Обзвон участников
Подготовлено примечаний:
Оператор
Добрый день, и добро пожаловать на телеконференцию Beazer Homes по доходам за квартал, закончившийся 31 марта 2019 года.Сегодняшний звонок записывается, и его повтор будет доступен на сайте компании сегодня же. Кроме того, слайды PowerPoint, которые будут сопровождать эту телеконференцию, доступны в разделе по связям с инвесторами на веб-сайте Компании www.beazer.com.
Сейчас я передам вызов Дэвиду Голдбергу, вице-президенту и казначею.
Дэвид Голдберг — Вице-президент и казначей
Спасибо, Мишель. Добрый день и добро пожаловать на телеконференцию Beazer Homes, посвященную нашим результатам за второй квартал 2019 финансового года.
Прежде чем мы начнем, вы должны знать, что во время этой телеконференции мы сделаем прогнозные заявления. Такие заявления связаны с известными и неизвестными рисками, неопределенностями и другими факторами, которые описаны в наших документах SEC, включая нашу форму 10-Q за квартал, что может привести к тому, что фактические результаты будут существенно отличаться от наших прогнозов.
Любые прогнозные заявления действительны только на дату, когда это заявление сделано. И мы не берем на себя никаких обязательств по обновлению или пересмотру каких-либо прогнозных заявлений, будь то в результате новой информации, будущих событий или иным образом.Время от времени появляются новые факторы, и все эти факторы просто невозможно предсказать. Сегодня ко мне присоединяются Аллан Меррилл, наш президент и главный исполнительный директор; и Боб Саломон, наш исполнительный вице-президент и главный финансовый директор.
Во время нашего сегодняшнего звонка Аллан рассмотрит основные моменты второго квартала и расскажет об обесценении, которое мы понесли в этом квартале. Боб более подробно расскажет о наших результатах за второй квартал, а также о наших ожиданиях на третий квартал 2019 финансового года.Затем я вернусь, чтобы предоставить более подробную информацию о наших расходах на землю, счет сообщества и наших обновленных приоритетах распределения капитала, после чего Аллан подведет итоги.
После наших подготовленных замечаний мы ответим на вопросы в оставшееся время.
Сейчас я передам вызов Аллану.
Аллан П. Меррилл — Президент и главный исполнительный директор
Спасибо, Дэвид, и спасибо, что присоединились к нам сегодня днем. Наши результаты за второй квартал оказались на уровне или выше ожиданий, которые мы выразили в отчете о прибылях и убытках за прошлый квартал.Это отразило лучшие условия для продаж на наших рынках, а также пользу от нашей быстрой реакции на снижение спроса, которое мы испытали в первом квартале. Улучшение условий продаж стало результатом сочетания более низких ставок по ипотечным кредитам, ускорения роста заработной платы и более конкурентоспособных цен на жилье. В то же время наша стратегия обеспечения исключительной ценности по доступной цене привела к изменениям в нашем продукте и ценах, которые повысили доступность.
Обнадеживает то, что последовательные улучшения продаж, которые произошли в течение квартала, продолжались и в апреле.Что касается операционных и финансовых показателей, то во втором квартале мы сделали то, что обещали. Заказы снизились меньше, чем мы ожидали, поскольку растущее количество участников сообщества компенсировало снижение темпов продаж. Продажи на сообщество в месяц неплохо выросли по сравнению с первым кварталом и отразили здоровые темпы в соответствии с нашими результатами за второй квартал последних лет.
ВыручкаHome Building также превзошла наши ожидания благодаря большему количеству закрытий. Наша операционная маржа была лучше, чем мы ожидали, поскольку мы извлекли выгоду из более высокой валовой прибыли и более низких коммерческих и административных расходов в процентах от выручки.И, наконец, расходы на землю были почти такими же, как и в прошлом году, как мы и предполагали. Что касается распределения капитала, мы вернули инвесторам более 10 миллионов долларов в течение квартала, включая 7,5 миллионов долларов в виде обратного выкупа акций и 5 миллионов долларов в виде погашения долга. Напомним, наш Совет директоров одобрил программу обратного выкупа акций на сумму 50 миллионов долларов.
На сегодняшний день мы завершили строительство примерно на 24 миллиона долларов. Мы также заявляли, что к концу года наши выплаты по погашению долга превысят объемы выкупа акций, и этого мы по-прежнему ожидаем.Короче говоря, в операционном и финансовом плане мы продолжаем реализовывать нашу стратегию сбалансированного роста, нацеленную на повышение доходности и уменьшение заемных средств. Конечно, другой важной новостью второго квартала стало обесценение некоторых калифорнийских активов. Это неденежное обесценение составило 148 млн долларов до налогообложения и 107 млн долларов после налогообложения.
Обесценение относится к 15 активам, все из которых в настоящее время или ранее классифицировались как земли, предназначенные для будущего строительства.14 из этих активов были приобретены до 2007 года. Обесценение произошло по двум категориям. Первый связан с девятью проектами, которые находятся в стадии разработки и / или активно продаются. Активы, попавшие в эту категорию, составили 109 миллионов долларов от обесценения до налогообложения и были распределены по всей Южной Калифорнии. Вторая категория относилась к шести общинам, которые мы решили продать и которые все еще классифицировались как земли, предназначенные для будущего строительства. Обесценение этих активов до налогообложения составило 39 миллионов долларов, разделенных между Северной и Южной Калифорнией.
После этих корректировок у нас не осталось земли, предназначенной для будущего строительства в Калифорнии. Мы много раз обсуждали основные характеристики наших земельных активов. Мы постоянно заявляли, что, когда эти активы начнут сдавать дома, их маржа будет ниже средней по Компании. Тем не менее, по мере улучшения рыночных условий за последние несколько лет мы постепенно активировали их, что позволило нам начать возвращать ранее связанный капитал.
Как уже широко отмечалось, спрос на новые дома значительно замедлился за последние три месяца календарного 2018 года. Это особенно характерно для Калифорнии, где доступность является особенно сложной. Оценив основные причины замедления и наблюдая за реакцией конкурентов на рынке, мы пришли к выводу, что нам необходимо снизить цены для достижения приемлемого темпа продаж. Меры по ценообразованию, которые мы предприняли, привели нас к более широкому рынку и стимулировали более высокий уровень спроса для тех сообществ, которые открыты для продажи.С точки зрения бухгалтерского учета, такое снижение цен привело к тому, что некоторые активы оказались ниже линии обесценения.
Слайд 7 — иллюстрация того, как работает этот процесс ослабления. Самый простой способ подумать об этой линии обесценения — добавить ли недисконтированный будущий денежный поток нашего сообщества к его текущей балансовой стоимости. Если да, то даже если маржа ниже рыночных норм, актив находится выше линии обесценения, и корректировка бухгалтерского учета не допускается. Однако, если эти денежные потоки составляют сумму ниже его текущей балансовой стоимости, актив должен быть обесценен.Снижение цен, которое мы предприняли для согласования с рынком, привело к тому, что совокупные денежные потоки по обесцененным активам оказались ниже линии. На этом этапе бухгалтерский учет прост.
Мы должны отразить убыток по разнице между приведенной стоимостью тех же денежных потоков и текущей балансовой стоимостью. Приведенная стоимость этих денежных потоков составила в среднем чуть менее 50% от предыдущей балансовой стоимости, но значительно варьировалась в зависимости от продолжительности существования каждого сообщества. Вторая категория обесценения связана с активами, которые мы решили продать, а не развивать.Это решение было основано на рыночных условиях для каждого актива, нашей долгосрочной стратегии в Калифорнии и нашем намерении улучшить рентабельность инвестиций. В настоящее время эти активы отражаются по ожидаемой стоимости реализации.
С учетом этого обесценения мы можем ускорить возврат денежных средств, что позволит нам перераспределить капитал таким образом, чтобы принести пользу акционерам. Будь то новые сообщества, сокращение долга или обратный выкуп акций, этот капитал можно использовать лучше. А при более низкой основе затрат эти сообщества будут генерировать более высокую валовую прибыль, что приведет к более высоким EBITDA и EPS в следующие несколько лет.
На этом этапе я передам вызов Бобу, чтобы он подробно рассмотрел результаты.
Роберт Л. Саломон — Исполнительный вице-президент, главный финансовый директор и главный бухгалтер
Спасибо, Аллан, и добрый день всем. Если посмотреть на результаты нашего второго квартала по сравнению с предыдущим годом, количество заказов на новое жилье снизилось на 4,8% до 1 598. При продажах на сообщество в месяц 3.3. Доходы от жилищного строительства упали на 5% до 421 миллиона долларов США из-за снижения количества закрытий на 10%, что частично компенсировалось увеличением ASP на 7% до 371 000 долларов США.Коэффициент конверсии невыполненных заказов превысил 74%, увеличившись почти на 800 базисных пунктов, так как количество домов, проданных и закрытых в этом квартале, увеличилось.
Наше среднее количество сообществ выросло до 163, что на 12 сообществ больше, чем в предыдущем году, и мы вошли в квартал с 166 активными сообществами. Наша валовая маржа за второй квартал без учета амортизированных процентных ставок и отказа от обесценения составила 19,8%, что превзошло наши ожидания, главным образом в связи с единовременными выплатами, которые добавили к нашим результатам около 60 базисных пунктов.Доля SG&A в общей выручке составила 12,7%, что на 10 базисных пунктов больше. В результате скорректированный показатель EBITDA составил 33 миллиона долларов. Наконец, благодаря обесценению некоторых активов в Калифорнии, наш чистый убыток от продолжающейся деятельности составил 101 миллион долларов. Без учета этого обесценения мы получили операционную прибыль.
Теперь перейдем к нашим ожиданиям на третий квартал. Наши продажи сильно выросли в апреле, и мы продолжаем использовать это улучшение для увеличения прибыльности.Соответственно, мы ожидаем, что заказы вырастут на 5-10% в годовом исчислении, что предполагает аналогичный рост нашего среднего числа сообществ. Говоря о валовой прибыли, исходя из того, что мы закрыли до сих пор в третьем квартале, и маржи в портфеле заказов, мы ожидаем, что валовая прибыль составит около 19%. Конечно, сочетание наших дополнительных продаж, завершившихся в этом квартале, может повлиять на это на четверть пункта в любом направлении. Тем не менее, валовая прибыль в третьем квартале должна представлять собой минимум за год. Поскольку мы работаем за счет повышенных стимулов, введенных ранее в этом году, и потому, что дома, которые будут построены, будут представлять более высокий процент закрытия в четвертом квартале.
Наш ASP должен быть выше $ 375 000 по сравнению с предыдущим годом и вторым кварталом. Мы ожидаем, что коэффициент конверсии невыполненных заказов превысит 60%. Общие и административные расходы должны быть ниже прошлогодних в долларовом выражении. И, наконец, денежная составляющая расходов на землю должна быть выше 100 миллионов долларов.
Сейчас я передам слово Дэвиду.
Дэвид Голдберг — Вице-президент и казначей
Спасибо, Боб. В этом квартале мы потратили 140 миллионов долларов на землю и застройку относительно стабильно по сравнению с предыдущим годом, после изменения нашей стратегии в отношении наших давних активов в Калифорнии, теперь мы сократили баланс нашей земли, предназначенной для будущего развития, до менее чем 29 миллионов долларов. .Это ниже пикового уровня в 420 миллионов долларов в 2009 году, и, как отметил Аллан, у нас больше нет земли, предназначенной для будущего строительства в Калифорнии.
Во второй половине этого финансового года мы ожидаем, что наши расходы на землю снизятся примерно на 100–150 млн долларов по сравнению с предыдущим годом, что обусловлено двумя факторами. Во-первых, мы продолжаем повышать эффективность нашего баланса за счет расширения использования опционов и предотвращения крупных оптовых закупок. Во-вторых, наши расходы на землю во второй половине прошлого года включали приобретение нами венчурных домов.
На слайде 11 мы обрисовываем компоненты нашего ожидаемого роста количества сообществ в ближайшие кварталы. Расходы на землю за последние 18 месяцев позволили нам рассчитывать на растущее количество жителей. В большинстве этих новых сообществ ASP будут ниже нашей текущей средней цены продажи. Мы по-прежнему ожидаем завершения 2019 финансового года с более чем 170 активными сообществами, хотя прогнозировать точные квартальные тенденции сложно. Целью нашей стратегии сбалансированного роста является повышение рентабельности наших активов за счет повышения EBITDA без неэффективного увеличения нашей базы активов.
Вы можете увидеть успех, которого мы достигли до сих пор, в верхней части слайда 12. В предыдущих кварталах мы обсуждали продолжающееся улучшение, которое мы ожидали в ROA, поскольку неприбыльные активы на нашем балансе начали приносить доход. Хотя обесценение, которое мы приняли в этом квартале, снизило балансовую стоимость этих активов, основная история не изменилась. Мы ожидаем, что в течение следующих 12 месяцев практически все бывшие земельные активы, которые сегодня не приносят доход, начнут приносить доход, что будет способствовать повышению доходности.
Мы добились значительного прогресса в выполнении наших программ выкупа акций и долговых обязательств после их объявления в ноябре. После завершения нашего ASR в первом квартале мы продолжили выкуп акций, в результате чего общий выкуп составил более 24 миллионов долларов США или 2,2 миллиона акций, что составляет более 6% компании. Кроме того, в течение второго квартала мы начали обратный выкуп долгов и до 31 марта погасили более 5 миллионов долларов наших непогашенных облигаций на 2023 и 2025 годы.
За последние 11 лет мы добились значительных успехов в улучшении нашей левериджа, сократив нашу общую задолженность более чем на 500 миллионов долларов. С 2015 финансового года наш чистый долг по отношению к скорректированному показателю EBITDA снизился до 6,2x, и мы нацелены на соотношение чистого долга к скорректированному показателю EBITDA на минимальных уровнях (ph) за счет повышения нашей прибыльности, в том числе за счет монетизации ранее не приносящего дохода капитала и дальнейшего сокращения долг.
На этом позвольте мне передать звонок Аллану для заключения.
Аллан П. Меррилл — Президент и главный исполнительный директор
Спасибо, Дэвид. Нас обнадеживает возобновление более здоровых тенденций спроса этой весной, и мы по-прежнему уверены, что наша стратегия сбалансированного роста подходит для этих условий. В течение следующего квартала наша деятельность в поддержку этой стратегии будет включать в себя увеличение количества наших сообществ за счет домов по более доступным ценам, расширение нашей платформы и технологий выбора ипотеки, а также расширение зоны нашего присутствия.Что касается распределения капитала, мы продолжим инвестировать в наш бизнес, ускорить обратный выкуп долгов и выкупить акции. Я хочу поблагодарить нашу команду за их постоянные усилия. Я уверен, что у нас есть люди, стратегия и ресурсы для реализации нашего плана в ближайшие годы.
И на этом я передам звонок оператору, чтобы он ответил на вопросы.
Вопросы и ответы:
Оператор
Спасибо.В этот раз мы хотели бы начать секцию вопросов и ответов конференции. (Инструкции оператора) Наш первый вопрос будет от Джея МакКэнлесса из Wedbush. Ваша линия теперь открыта.
Джей МакКанлесс — Wedbush Securities — Аналитик
Привет, добрый день. Спасибо, что ответили на мои вопросы. Я предполагаю, что первый у меня был с шестью активами, которые вы, ребята, хотите продать. Есть ли у вас что-либо из этого по контракту, когда эти деньги попадут на баланс?
Дэвид Голдберг — Вице-президент и казначей
Нет, Джей, ни с одним из них еще не заключен контракт.Решение, которое мы только что приняли в этом квартале, но конвертировать его в наличные деньги будет для нас первоочередной задачей, но я не могу назвать точные сроки на данный момент.
Джей МакКанлесс — Wedbush Securities — Аналитик
Хорошо. Я предполагаю, что, может быть, это просто замедление темпов продаж или некоторое снижение цен на этих рынках. Не могли бы вы рассказать о некоторых факторах? Во-первых, вы должны сделать это в этом квартале.
Дэвид Голдберг — Вице-президент и казначей
Конечно.Так что мы не сообщаем никаких новостей, вспоминая и указывая на то, что период октябрь, ноябрь, декабрь был довольно неравномерным. И это хороший эвфемизм для жесткого катания на санях с точки зрения продаж новых домов, особенно в Калифорнии. И это действительно привело нас к глубокому изучению каждого из наших сообществ, того, что делают наши конкуренты, структурных проблем на рынке. И я думаю, что все мы выиграли от повышения цен на этом рынке за последние несколько лет. И мы поняли, что находимся в точке, где все закончилось.Доступность имеет первостепенное значение, и в любом контексте роста цен на жилье мы просто слишком высоки, чтобы поддерживать приемлемый уровень продаж. Итак, мы приняли решение о перезагрузке тех активных сообществ и тех, которые мы делаем активными, но это также дало нам возможность, Джей, подумать о географии и рынках Северной и Южной Калифорнии, где находится наш могущественный союзник. Что у нас хорошо получается? На какие субрынки мы действительно хотим ориентироваться? Какие продукты? Какие профили покупателей?
И, как я отмечал в своих комментариях, 14 из 15 активов, с которыми мы имели дело, были в бухгалтерском учете более 12 лет.Итак, мы сказали, что у нас есть эти активы, мы действительно собираемся поехать в эти места, чтобы построить эти активы, или пришло время признать, что у нас есть целенаправленная стратегия, мы можем построить растущий бизнес в Калифорнии, но давайте построим его на наших условиях в тех местах, где мы хотим быть. И на самом деле все это как бы соединилось, и мы сказали, что пришло время, спусковым крючком явно было снижение цен и скорости в том первом финансовом квартале, но это привело к более глубокому анализу, который мы сделали.
Джей МакКанлесс — Wedbush Securities — Аналитик
И я был впечатлен, увидев, что валовая прибыль, похоже, будет немного увеличиваться последовательно от 2 до 3, что не соответствует графику, я думаю, это то, что мы видели у некоторых из ваших конкурентов. Может быть, вы можете рассказать об уровнях дисконтирования во втором квартале этого года по сравнению с прошлым годом? А как двигались наши скидки по мере того, как вы перешли в апрель?
Дэвид Голдберг — Вице-президент и казначей
Итак, пара вещей, просто пояснение.Наша валовая прибыль за второй квартал составила 19,8%, мы признаем, что у нас были некоторые преимущества в этом отношении, и на самом деле она была больше примерно 19,2%. Мы сказали, что исходя из отставания и того, что мы уже закрыли за квартал, 19% выглядит примерно правильно. Мы все еще в разгаре весеннего сезона продаж. У нас есть спецификации, мы их будем продавать. В зависимости от сочетания проданных и закрытых спецификаций, мы думаем, что маржа в третьем квартале будет колебаться около этих 19%, и мы дали взятый в скобки диапазон на 25 базисных пунктов в сторону увеличения или уменьшения около 19%.Итак, я бы сказал, что будет справедливо характеризовать некоторую плоскостность от Q3 или Q2 до Q3.
Позвольте мне перейти к более структурной части вашего вопроса. Если вы думаете, а не если вы думаете, если мы говорим о том, что произошло с точки зрения продаж, январь был лучше декабря в том смысле, что он был меньше по сравнению с прошлым годом. И февраль был лучше, чем январь, потому что он был почти, но не совсем неизменным по сравнению с прошлым годом. Март был без изменений по сравнению с аналогичным периодом прошлого года, а апрель, откровенно говоря, вырос на двузначные числа.Итак, чего вы разумно ожидали, и на самом деле произошло то, что торговая среда проявила себя как сокращение стимулов за этот период времени. Но мы разберемся. Мы работали во втором квартале, а в третьем мы будем иметь дело с некоторыми из усиленных стимулов, которые мы выдвинули в январе и феврале для стимулирования активности. Я думаю, что мы в значительной степени проработали это в третьем квартале, но это своего рода динамика того, что происходит со стимулами.
Джей МакКанлесс — Wedbush Securities — Аналитик
Ага. Спасибо за улучшение валовой прибыли.
Дэвид Голдберг — Вице-президент и казначей
Да, Джей, чтобы прояснить ситуацию, и я хочу убедиться, что мы все это как следует. Мы также сказали, что третий квартал будет падением рентабельности. Теперь мы видим улучшения в четвертом квартале, и, возможно, именно в этом и была путаница, но ясно, что в третьем квартале, как сказал Аллан, и в четвертом квартале некоторые улучшения, когда мы направимся в следующий год.
Аллан П. Меррилл — Президент и главный исполнительный директор
Спасибо, Дэвид. И я думаю, единственный другой вопрос, который у меня был, был ли вы, ребята, можете поговорить, я думаю, что вы, Аллан, вы сказали, что рост в апреле двузначный. Было ли это — можете ли вы поговорить об этом подробнее? А также мы слышали от других строителей, что активный взрослый покупатель внезапно ожил в марте и апреле. Так что, если вы, ребята, можете прокомментировать эти две вещи, и на этом все для меня.
Ну, на самом деле у меня нет для вас комментария по дифференциации сегментов.Я бы сказал, что мы увидели силу нашего присутствия в каждой ценовой категории и на каждом рынке, поскольку февраль превратился в март, а март — в апрель. Поэтому я бы не стал пытаться различать так много конкретных профилей покупателей, которых не было в нашем опыте.
Джей МакКанлесс — Wedbush Securities — Аналитик
Хорошо. И тогда заказы на апрель по сравнению с прошлым годом?
Да. Так что они выросли больше, чем подсчет сообщества. У нас был фактический рост темпов в течение одного месяца, но я думаю, что Боб правильно определил, что мы хотим сделать перед лицом этой силы — восстановить маржу.Итак, один месяц, один месяц полезен, но мы увеличили количество заказов на 5% или 10%, потому что мы собираемся использовать ту среду, в которой мы живем лучше, мы ее уже использовали. Мы использовали его в апреле и продолжим делать это в мае, чтобы увеличить рентабельность. И я заинтересован в улучшении прибыльности и порядка, а не только одного или другого.
Звучит хорошо. Спасибо, что ответили на мои вопросы.
Дэвид Голдберг — Вице-президент и казначей
Спасибо, Джей.
Наш следующий вопрос будет исходить от Майкла Рехо из J.P. Morgan. Ваша линия теперь открыта.
Мэгги МакГрат — Дж. П. Морган — аналитик
Привет. На самом деле это Мэгги для Майка. Просто чтобы продолжить последний вопрос. Вы сказали, что заказы на протяжении всего квартала росли лучше, а апрель вырос на двузначные цифры, и что вы видели силу на каждом ценовом уровне на этом новом рынке, но есть ли какие-то конкретные рынки, которые вы хотели бы назвать или привлечь больше ?
Дэвид Голдберг — Вице-президент и казначей
Я уверен.Я имею в виду, что было несколько мест, которые для нас, и позвольте мне, я отвечу на ваш вопрос довольно прямо, но позвольте мне просто убедиться, что запись ясна. Каждый раз, когда мы или любой другой строитель говорим о силе рынка, это всегда немного опасно, потому что вы можете смотреть на макроданные на рынке и говорить: ну, ну, это не то, что говорят другие. Так что часто сочетание сообществ между годом и годом может немного искажать. И я приведу вам небольшой пример. Наше подразделение с самым высоким темпом продаж во втором квартале — Лас-Вегас.
Мы действительно гордимся тем, что они сделали в больших количествах, но знаете что, темпы их роста снизились на 20% по сравнению с прошлым годом. Такова история, которая была неудачной, или история, лучшая для нас в стране, и поэтому я просто хочу быть осторожным, когда мы комментируем отдельные рынки. Это область большой силы. Мы увидели силы в Средней Атлантике и в Роли, чтобы выбрать несколько примеров мест, где в годовом исчислении мы наблюдали хороший рост продаж.
Мэгги МакГрат — Дж.П. Морган — Аналитик
Хорошо. Спасибо.
Оператор
Наш следующий вопрос будет задан Аланом Ратнером из Zelman & Associates. Ваша линия теперь открыта.
Алан Ратнер — Zelman & Associates — Аналитик
Привет, ребята. Добрый день. Спасибо, что ответили на мои вопросы.
Дэвид Голдберг — Вице-президент и казначей
Привет, Алан. Конечно.
Алан Ратнер — Zelman & Associates — Аналитик
Итак, по проектам с ограниченными возможностями это звучит так, как будто они были в списках уже какое-то время.Были ли они ранее ослаблены во время экономического спада или это было только первое обвинение, которое вы взяли на них?
Дэвид Голдберг — Вице-президент и казначей
Я бы немного неохотно отвечал на него окончательно. У одного или нескольких из них могло быть обесценение, но они не были обесценены с 2007 или 2008 года, потому что, как только они перешли в категорию земель, находящихся в собственности, где оценка заключалась в том, что они могут быть восстановлены, они ежегодно тестировались на обесценение в том же способ.Так что разумно предположить, что они в основном остались нетронутыми.
Алан Ратнер — Zelman & Associates — Аналитик
Понятно, хорошо. Так что нам не обязательно экстраполировать это на другие активы в Калифорнии, которые могли быть куплены за последние несколько лет, как указание на то, что там может быть какой-то риск?
Дэвид Голдберг — Вице-президент и казначей
Я согласен с вами, вам не следует делать такую экстраполяцию.Вот почему мы осторожно указывали срок действия этих активов.
Алан Ратнер — Zelman & Associates — Аналитик
И я предполагаю, что в более широком смысле, когда я знаю, что ведро земли, предназначенное для будущего развития, невелико с точки зрения компании. Но я думаю, что вы, ребята, и другие в прошлом имели своего рода… в прошлом сказали, что данный показатель или количество сообществ, у которых были потенциальные индикаторы нарушений. Так что мне просто любопытно, не могли бы вы просто поговорить о компании.Есть ли другие активы, другие сообщества, где, возможно, они приближаются к тому выступу, через который вы нас как бы провели?
Дэвид Голдберг — Вице-президент и казначей
Итак, пара вещей, и мы с Бобом разделим это на части. Я думаю, вы задаете вопрос о сообществах, за которыми мы внимательно следим, и у нас есть для этого процесс. На земле находится такое небольшое количество сообществ, что их на самом деле нет с точки зрения обесценения.Они там, потому что есть причина, по которой разработка этого актива не входит в нашу стратегию. Это может быть связано с правами; это может быть связано с инфраструктурой; это может быть связано с продуктом или профилем покупателя. Так что, как правило, существует множество причин, по которым у нас нет текущего намерения развиваться. И те, что были в другом ведре, на котором приземлились, содержали ведро стоимостью до 29 миллионов долларов, и это действительно соответствует намерению. Я думаю, что ваш вопрос касается остальной части бизнеса, и Боб может справиться с работой лучше, чем я, рассказывая о нашем процессе с этими активами.
Роберт Л. Саломон — Исполнительный вице-президент, главный финансовый директор и главный бухгалтер
Да, у нас был процесс списка наблюдения, при котором сообщества, у которых есть фактическая или ожидаемая маржа ниже определенного уровня, включаются в список наблюдения. И когда вы — когда вы получите наш 10-Q, вы заметите, что у нас было около 13 сообществ, которые мы рассмотрели, и девять из них были повреждены. А остальные три были проверены без учета математических или иных причин, по которым они не попали в ту категорию, в которой мы должны были проводить анализ недисконтированного или дисконтированного денежного потока.
Алан Ратнер — Zelman & Associates — Аналитик
Это очень полезно. Если бы я мог прокрасться сюда, в Калифорнию. Я думаю, Аллан, может быть, просто прокомментируй, ты упомянул, что собираешься переработать капитал в сделки, которые сегодня имеют больше смысла, но каковы твои долгосрочные обязательства перед Калифорнией? Мы много слышали об этом только из общего окружения. И это, безусловно, дорогостоящая область для вас, ребята, требует больших вложений.Так это — все еще тот рынок, на котором вы, ребята, привержены своей долгосрочной перспективе?
Аллан П. Меррилл — Президент и главный исполнительный директор
Я рад, что вы спросили, потому что есть много людей, которые слушают этот звонок, и некоторые из них — инвесторы, некоторые — сотрудники, а некоторые — продавцы земли. Так что я очень ценю, что вы спросили об этом. Мы воспользовались шансом или возможностью в ходе этой оценки, чтобы задать действительно фундаментальные вопросы о том, почему мы живем в Калифорнии.И если да, то в какой степени и на каких субрынках. И мы пришли к выводу, что дисбаланс спроса настолько велик, что это рынок, который оправдывает наш капитал.
Теперь ваши оценки нашей шкалы относительно других означают, что мы должны выбирать наши точки. Мы должны знать, на каких субрынках, какие типы продуктов, какие профили покупателей мы хотим преуспеть, и я думаю, что мы можем это сделать. Наши рыночные амбиции заключаются в том, чтобы вложить минимум 200 миллионов долларов и, возможно, больше через циклы. И я думаю, что у нас может быть 1000 единиц бизнеса.Сейчас нет, но у нас очень сильное присутствие на рынке Сакраменто, и мы собираемся быть там еще долгое время. У нас есть самый крупный генеральный план в Сакраменто, который будет запущен в этом году. В Южной Калифорнии мы находимся на строительной площадке I215, I10, I5 в Оушенсайд. Нам нравятся эти субрынки. В Южной Калифорнии есть целая куча недвижимости, которая не является нашим профилем покупателя, не является нашим аппетитом, и я думаю, что дисциплинированность в том, чтобы знать, кто мы, дает нам действительно хороший шанс, чтобы Калифорния была важной и высокой рентабельностью бизнеса с активами для нас. .
Алан Ратнер — Zelman & Associates — Аналитик
Отлично. Большое спасибо за это. Удачи.
Дэвид Голдберг — Вице-президент и казначей
Спасибо, Алан.
Оператор
(Инструкции для оператора) Наш следующий вопрос будет исходить от Алекса Бэррона из Исследовательского центра жилищного строительства. Ваша линия теперь открыта.
Алекс Баррон — Центр жилищных исследований — Аналитик
Да, спасибо.Итак, я напомнил, что вы упомянули эти активы, которые (неразборчиво) в Калифорнии, но я надеялся получить немного больше информации о том, где эти сообщества с более высокой ценой продвигаются вверх по уровню, или вы можете прокомментировать более конкретные места?
Дэвид Голдберг — Вице-президент и казначей
Что ж, Алекс, мы не говорим об отдельных активах в этом контексте. Мы сказали о сообществах и категории девяти, что они были разбросаны по Южной Калифорнии.И они были в разных типах продуктов, в разных местах, и снова всем, кроме одного, было больше 12 лет. Так что я бы не сказал, что они действительно отражали какую-то конкретную стратегию, но я думаю, что у них есть общая черта — это локации B. И что мы осознали, так это то, что мы знаем, что такое локация А, и мы сохранили локации А и Б, а остальные собираемся продавать.
Алекс Баррон — Центр жилищных исследований — Аналитик
Ага.И это будет моим следующим вопросом. Итак, забегая вперед, когда вы сказали, что собираетесь выбирать свои споты, что вы, ребята, — я думаю, вы определяете, чем больше ваши стратегии будут продвигаться в Калифорнии? Что ты собираешься там делать?
Дэвид Голдберг — Вице-президент и казначей
Ага. Мы — разработчик EVAP, и мне очень жаль, что я использую это аббревиатуру: «Исключительная ценность и доступная цена». Мы не собираемся опускаться до самой низкой цены.Есть вещи, которые мы хотим привнести в дом, которые, по нашему мнению, важны с точки зрения качества строительства и материалов, а также долгосрочной пригодности для жизни, комфорта и энергии. Мы просто не идем на компромисс, и это означает, что мы не можем быть абсолютно низкой ценой. Но мы думаем, что решения, которые мы приняли в отношении того, что мы включаем, имеют очевидную ценность для покупателей.
Сказав это, мы определенно не гонимся за рынком первой, второй и третьей переездов, потому что есть другие строители, для которых это действительно их опыт.Итак, мы находимся в этом первом профиле покупателя, который продвигается вверх, с огромным количеством одноэтажных продуктов, просто за то, что они стоят. Это ориентировано на ценность, но не конкурирует исключительно на основе цены. И есть несколько почтовых индексов, несколько школьных округов, где у нас хорошая ДНК. Мы очень давно в Калифорнии. У нас есть наследие множества успешных сообществ. И мы действительно изучили, в чем у нас хорошо получается, и этим мы продолжим заниматься.
Алекс Баррон — Центр жилищных исследований — Аналитик
Понятно.А потом мне показалось, что я слышал, как вы сказали, что в следующем квартале вы ожидаете, что маржа будет низкой, как будто я не знаю, пропустил ли я, например, если вы дадите какие-то базисные пункты, или она примерно на пару пунктов ниже, или как вы дать хоть какое-то представление о возможном диапазоне прибылей в этом квартале?
Роберт Л. Саломон — Исполнительный вице-президент, главный финансовый директор и главный бухгалтер
Ага. Алекс, в сценарии мы сказали, что нас будет около 19, но поскольку мы продаем дополнительные спецификации в этом квартале, которые могут колебаться на 25 базисных пунктов вверх или вниз, но это будет минимум после ожидаемого года. рентабельность также улучшится в четвертом квартале.
И Алекс, чтобы убедиться, что вы слышали, мы действительно повысили прибыль на 19 пунктов, валовую прибыль на 8% в этом квартале, но в нем были некоторые единовременные статьи. Так что более регулярно около 19,2. То, о чем мы говорили, было относительно ровным 19, и тогда этот диапазон является нашим ориентиром.
Алекс Баррон — Центр жилищных исследований — Аналитик
Хорошо. Я ценю это. Спасибо.
Дэвид Голдберг — Вице-президент и казначей
Спасибо, Алекс.
Оператор
В настоящее время у нас нет дополнительных вопросов.
Дэвид Голдберг — Вице-президент и казначей
Хорошо, хорошо. Что ж, мы хотим поблагодарить всех, кто принял участие в объявлении о прибылях и убытках за второй квартал. Мы с нетерпением ждем возможности поговорить с вами примерно через 90 дней. Спасибо.
Оператор
На этом сегодняшняя конференция завершается. Большое спасибо за ваше участие в сегодняшней телеконференции.В это время все участники могут отключиться.
Продолжительность: 33 минуты
Участники звонка:
Дэвид Голдберг — Вице-президент и казначей
Аллан П. Меррилл — Президент и главный исполнительный директор
Роберт Л. Саломон — Исполнительный вице-президент, главный финансовый директор и главный бухгалтер
Джей МакКанлесс — Wedbush Securities — Аналитик
Мэгги МакГрат — Дж.П. Морган — Аналитик
Алан Ратнер — Zelman & Associates — Аналитик
Алекс Баррон — Центр жилищных исследований — Аналитик
Подробнее Анализ БЖ
Расшифровка стенограммы предоставлена AlphaStreet
Эта статья представляет собой стенограмму этого конференц-звонка, подготовленного для The Motley Fool. Несмотря на то, что мы стремимся к лучшему, в этой расшифровке могут быть ошибки, упущения или неточности.Как и в случае со всеми нашими статьями, The Motley Fool не несет никакой ответственности за использование вами этого контента, и мы настоятельно рекомендуем вам провести собственное исследование, в том числе прослушать звонок самостоятельно и прочитать документы SEC компании. Пожалуйста, ознакомьтесь с нашими положениями и условиями для получения дополнительной информации, включая наши Обязательные заявления об отказе от ответственности с заглавной буквы.
Эта статья представляет мнение автора, который может не согласиться с «официальной» позицией рекомендаций премиальной консультационной службы Motley Fool.Мы разношерстные! Ставка под сомнение по поводу инвестиционного тезиса — даже нашего собственного — помогает всем нам критически относиться к инвестированию и принимать решения, которые помогают нам стать умнее, счастливее и богаче.
Эндорибонуклеаза, функционально связанная с перинуклеарными мРНП, ассоциациями контроля качества с комплексами ядерных пор
Abstract
Экспорт ядерной мРНК является критическим этапом в экспрессии эукариотических генов, которая в дрожжах связана со сборкой и надзором за котранскрипционные медиаторные рибонуклеопротеиновые частицы (мРНП).Было описано несколько систем наблюдения, которые отслеживают биогенез и экспорт ядерных мРНП, но механизм, с помощью которого распознаются и устраняются неподходящие мРНП, остается плохо изученным. Здесь мы сообщаем, что консервативный белок PIN-домена Swt1 представляет собой РНК-эндонуклеазу, которая участвует в контроле качества ядерных мРНП и может связываться с комплексом ядерных пор (NPC). Swt1 проявлял эндорибонуклеазную активность in vitro, которая ингибировалась точечной мутацией в предсказанном каталитическом сайте.Swt1 не обладал четкой специфичностью последовательности, но демонстрировал сильное предпочтение одноцепочечным областям. Генетические взаимодействия были обнаружены между Swt1 и комплексами THO / TREX и TREX-2, а также с компонентами перинуклеарной системы наблюдения мРНП, Mlp1, Nup60 и Esc1. Ингибирование нуклеазной активности Swt1 увеличивало уровни и цитоплазматическую утечку несплайсированной аберрантной пре-мРНК и индуцировало устойчивое ядерное накопление РНК поли (A) + в штаммах mlp1 Δ и esc1 Δ.Сверхэкспрессия Swt1 также вызывала сильное накопление РНК поли (А) + в ядре. Swt1 обычно распределяется по ядру и цитоплазме, но концентрируется в комплексах ядерных пор (NPCs) у мутанта nup133 Δ, что вызывает кластеризацию NPC и дефекты в экспорте мРНП. Данные предполагают, что эндорибонуклеаза Swt1 может временно рекрутироваться в NPCs, чтобы инициировать деградацию дефектных пре-мРНП или мРНП, захваченных на периферии ядра, чтобы избежать их цитоплазматического экспорта и трансляции.
Сведения об авторе
Ядерный экспорт матричной РНК (мРНК) является важным этапом экспрессии эукариотических генов. Недавно синтезированные мРНК-предшественники обрабатываются во время синтеза, упаковываются в частицы рибонуклеопротеина-мессенджера (мРНП) и транспортируются через комплекс ядерных пор в цитоплазму. Чтобы избежать ядерного экспорта аберрантных транскриптов и их трансляции в цитоплазму, качество ядерных мРНП контролируется несколькими системами наблюдения. Здесь мы показываем, что консервативный белок Swt1 представляет собой РНК-эндорибонуклеазу, фермент, разрушающий РНК, который становится незаменимым при мутации факторов, участвующих в сборке котранскрипции мРНП и контроле качества мРНП.Мы обнаружили, что неактивный Swt1 увеличивает уровни и утечку цитоплазмы аберрантной необработанной мРНК предшественника. Более того, Swt1 накапливается в комплексах ядерных пор в мутанте с кластеризацией пор nup133 Δ. Таким образом, мы предполагаем, что эндорибонуклеаза Swt1 может временно рекрутироваться на периферию ядра, чтобы инициировать деградацию дефектных, захваченных порой пре-мРНП, чтобы предотвратить их несоответствующий цитоплазматический экспорт.
Образец цитирования: Skružný M, Schneider C, Rácz A, Weng J, Tollervey D, Hurt E (2009) Эндорибонуклеаза, функционально связанная с перинуклеарными мРНП, связанными с контролем качества с комплексами ядерных пор.PLoS Biol 7 (1): e1000008. https://doi.org/10.1371/journal.pbio.1000008
Академический редактор: Анита Корбетт, Университет Эмори, Соединенные Штаты Америки
Поступила: 17 июня 2008 г .; Принята к печати: 20 ноября 2008 г .; Опубликовано: 6 января 2009 г.
Авторские права: © 2009 Skružný et al. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Финансирование: MS была получателем исследовательской стипендии Александра фон Гумбольдта, CS была поддержана долгосрочной стипендией Научной программы Human Frontiers, а EH является получателем грантов от Deutsche Forschungsgemeinschaft (SFB 638 / B3) и Fonds der Chemischen Industrie.
Конкурирующие интересы: Авторы заявили, что никаких конкурирующих интересов не существует.
Сокращения: GFP, зеленый флуоресцентный белок; мРНП, мессенджер рибонуклеопротеиновая частица; NPC, ядерно-поровый комплекс; ОТ-ПЦР, полимеразная цепная реакция с обратной транскрипцией; сл синтетическая летальность; ТРЕКС, транскрипция-экспорт
Введение
Ядерный экспорт информационной РНК (мРНК) является фундаментальным процессом экспрессии эукариотических генов.В дрожжах вновь синтезированные пре-мРНК котранскрипционно собираются в частицы рибонуклеопротеина-мессенджера (мРНП) с помощью комплексов экспорта-транскрипции (TREX). Комплексы THO / TREX (Tho2-Hpr1-Mft1-Thp2-Tex1-Sub2-Yra1) и TREX-2 (Sac3-Thp1-Sus1-Cdc31) связывают транскрипцию мРНК с привлечением РНК-связывающих белков (например, Yra1, Npl3, и Nab2), а также консервативный рецептор экспорта мРНК Mex67-Mtr2 (TAP-p15 у человека) [1,2]. В отсутствие компонентов TREX или TREX-2 возникающие мРНК неправильно собираются в мРНП, что может привести к гибридизации с ДНК-матрицей (образование R-петель), дефектному полиаденилированию и накоплению на 3′-конце гена [ 3–5].В результате в ядре накапливаются некомпетентные к экспорту мРНП [6,7]. Во время созревания и транспорта ядерные mRNPs контролируются системами наблюдения для предотвращения экспорта и последующей трансляции аберрантных мРНК (rev. [8,9]). У мутантов TREX несовершенные мРНП могут быть нацелены и разрушены ядерной экзосомой вместе с комплексом полиаденилирования TRAMP [10].
Важной задачей контроля качества ядерной мРНК является предотвращение экспорта интрон-содержащих пре-мРНК в цитоплазму.Экспорт несплайсированных пре-мРНК редко встречается в клетках дикого типа, тогда как значительная утечка пре-мРНК наблюдается в штаммах с дефектами ранних факторов сплайсинга или для транскриптов, мутировавших в 5′-сайте сплайсинга или в последовательности точки ветвления [11 –13]. Утечка пре-мРНК, лишенных каких-либо явных дефектов сплайсинга, наблюдалась в клетках, лишенных Mlp1 (гомолог человеческого TPR) или Nup60, двух структурных компонентов нуклеоплазматической корзины комплекса ядерных пор (NPC) [14]. Это указывает на то, что Nup60-Mlp1 избирательно улавливает содержащие интроны пре-мРНК и, возможно, др. Аберрантно процессируемые мРНК, чтобы предотвратить их выход из ядра [14,15].Совсем недавно было показано, что Pml39, партнер связывания Mlp1, и Esc1, белок периферии ядра, участвующий в правильной локализации комплекса Nup60-Mlp1, вносят вклад в функцию Mlp1-Nup60 [16,17]. Однако механизм распознавания и удаления аберрантных транскриптов остается неясным.
Белок дрожжевого PIN-домена Swt1 / Yor166c (синтетический летальный с TREX 1) был идентифицирован при скрининге на синтетическую летальность с субъединицей TREX Hpr1, функционально взаимодействует с комплексом TREX и необходим для оптимальной скорости транскрипции [18].Модуль PIN (PilT N-конец) имеет длину приблизительно 130 остатков и имеет складку, аналогичную 5′-экзонуклеазам и эндонуклеазам FLAP [19,20]. Типичными чертами являются четыре инвариантных кислотных остатка, расположенные в отрицательно заряженной поверхностной щели, образующие предсказанный каталитический центр [21]. Эти остатки, вероятно, хелатируют двухвалентный ион и вместе с двумя соседними гидроксильными остатками участвуют в нуклеофильной атаке на фосфодиэфирную связь субстрата. PIN-домен человеческого SMG6 проявляет эндонуклеазную активность in vitro и необходим для его функции при нонсенс-опосредованном распаде мРНК (NMD) [22].Геном дрожжей кодирует восемь белков, содержащих домен PIN, включая Nob1 и Utp24 / Fcf1, которые помогают в эндонуклеолитическом расщеплении предшественников рРНК во время биогенеза рибосомы [23,24] и субъединицы Rrp44 / Dis3 экзосомы (C. Schneider и D. Толлервей, неопубликованные данные). Однако было продемонстрировано, что ни один из этих дрожжевых белков не функционирует как эндонуклеаза.
Здесь мы сообщаем, что эволюционно консервативный белок Swt1 проявляет РНК-эндонуклеазную активность, которая зависит от домена PIN.Swt1 необходим в клетках, нарушенных для образования котранскрипционных мРНП или контроля качества перинуклеарных мРНП, и может связываться с NPC. Эти данные показывают важную роль эндонуклеолитического расщепления в процессе наблюдения ядерной мРНК.
Результаты
SWT1 Функционально перекрывается с факторами, участвующими в сборке котранскрипционных мРНП и контроле качества перинуклеарных мРНПЧтобы идентифицировать факторы, которые взаимодействуют с TREX-2, комплексом экспорта транскрипции, который функционирует на внутренней стороне NPC, мы выполнили синтетический скрининг летальности (sl) с штаммом sac3 Δ.Среди кандидатов мы идентифицировали мутантный аллель гена SWT1 / YOR166 . Генетическое взаимодействие было подтверждено отсутствием роста гаплоидного штамма sac3 Δ swt1 Δ, полученного в результате потери плазмиды (фигура S1). Другие компоненты TREX-2 Thp1 и Sus1 были либо sl ( thp1 Δ), либо демонстрировали сильное синергетическое взаимодействие ( sus1 Δ) с swt1 Δ, демонстрируя тесное функциональное взаимодействие между Swt1 и комплексом TREX-2 (Таблица 1 и Рисунок S1).
Чтобы расширить эти исследования, мы проверили, перекрывается ли SWT1 функционально с другими факторами, участвующими в биогенезе и обмене мРНП. Сильные генетические взаимодействия наблюдались между swt1 Δ и компонентами THO / TREX ( tho2 Δ, hpr1 Δ, mft1 Δ, thp2 Δ и sub2–85 ) в соответствии с предыдущими выводами [18] . Напротив, не было обнаружено никаких генетических взаимодействий между swt1 Δ и рецептором экспорта мРНК Mex67 или адаптером Yra1 [18].Кроме того, мы обнаружили sl-взаимодействие между swt1Δ и NPL3 , которое кодирует мРНК-связывающий белок с множеством ролей в биогенезе мРНП [25,26] (Таблица 1 и Рисунок S1).
Эти исследования также выявили генетические взаимодействия между SWT1 и MLP1 , NUP60 и ESC1 (Таблица 1 и Рисунок S1). Mlp1 и Nup60 являются факторами, ассоциированными с NPC, которые, как полагают, составляют ядерную корзину NPC и участвуют в контроле качества mRNP перед ядерным экспортом [14].Esc1 является белком внутренней стороны ядерной мембраны, недавно было показано, что он влияет на ассоциацию комплекса Mlp1-Nup60 с NPC [17].
Несколько факторов, участвующих в различных этапах производства, экспорта и надзора мРНП, не взаимодействовали генетически с swt1Δ , включая субъединицы ядерной экзосомы (Rrp6 и Rrp44 / Dis3), компоненты комплекса TRAMP (Trf4 и Mtr4), Rat1 5′-экзонуклеаза и факторы, участвующие в обмене цитоплазматической мРНК (проверенные факторы см. В таблице 1).Эти генетические анализы показывают, что Swt1 специфически взаимодействует с факторами, необходимыми для котранскрипционной сборки и перинуклеарного контроля качества ядерных мРНП.
Swt1 обладает эндорибонуклеазной активностью in vitro
PIN-домен Swt1, который расположен в центральной области (остатки 130–262), является эволюционно консервативным и содержит все четыре консервативных кислотных остатка, участвующих в катализе (Рисунок S2). Поэтому мы проверили, обладает ли очищенный из дрожжей Swt1 нуклеазной активностью in vitro.Мы создали меченый FLAG Swt1 дикого типа и мутант Swt1, которые, как ожидается, будут каталитически неактивными (Swt1-D135N; Asp 135, как предполагается, является остатком каталитического центра). Они были экспрессированы в дрожжах и очищены с помощью аффинной хроматографии (рис. 1А). Способность этих белков расщеплять одноцепочечную РНК in vitro оценивали с использованием 5′- или 3′-радиоактивно меченных олиго (A) 30 субстратов. Swt1 дикого типа эффективно расщепляет как 5′-, так и 3′-меченные олиго (A) 30 РНК. Напротив, мутантный белок Swt1-D135N не проявлял активности в отношении 3′-меченого субстрата РНК и только очень слабую активность в отношении 5′-меченного зонда (рис. 1B).Остаточная активность на 5′-меченном субстрате может быть связана с загрязняющей 3′-5′-экзонуклеазой, что часто наблюдается в анализах экзонуклеаз (C. Schneider, неопубликованные данные). Отсутствие деградации 3′-меченного субстрата за счет этой активности, вероятно, отражает защиту за счет присутствия 3′-фосфата на [ 32 P] pCp-меченой РНК. Эксперименты с течением времени также показали, что Swt1 дикого типа разлагает как 5′-, так и 3′-меченные субстраты с очень сходной кинетикой (рис. 1C), что согласуется с активностью эндорибонуклеазы.
Рисунок 1. Swt1 обладает эндонуклеолитической активностью в отношении одноцепочечной РНК In vitro
(A) Очищенные белки Swt1 дикого типа и мутантные белки Swt1-D135N, используемые для измерения нуклеазной активности in vitro. Указанные меченые FLAG белки Swt1 были очищены аффинно из дрожжей и проанализированы с помощью SDS-PAGE и окрашивания Кумасси. M, маркер молекулярной массы.
(B) Белок Swt1 дикого типа или мутантный белок Swt1-D135N (по 0,5 мкг каждый) инкубировали с 5 ‘- [ 32 P] -конец, меченным (левая панель) или 3′ — [ 32 P] — меченый концом (правая панель) олиго (A) 30 РНК в течение 0, 60 и 120 мин при 30 ° C.Продукты реакции разделяли на денатурирующем геле 12% полиакриламид / 8 М мочевина и визуализировали с помощью авторадиографии. Зонды РНК, инкубированные без белка, служили отрицательными контролями, обозначенными отрицательным знаком (-). Звездочка (*) обозначает низкую активность контаминирующей экзонуклеазы дрожжей.
(C) Динамика активности РНКазы Swt1 на 5′- [ 32 P] -конце, меченном (левая панель) или 3′- [ 32 P] -конце, меченном (правая панель) олиго (A) 30 РНК. Анализ выполняли, как описано в (B), и аликвоты отбирали в указанные моменты времени.
(D) Swt1 дикого типа или мутантный Swt1-D135N инкубировали с 52-нуклеотидной РНК, меченной 5 ‘- [ 32 P] -концом, со стабильным концевым стержнем в течение 0, 30 и 60 минут при 30 ° C. , и реакцию анализировали, как указано в (B). ОН, частичный щелочной гидролиз РНК-зонда, используемого в качестве лестницы молекулярной массы. Положения стержневых остатков обозначены синим цветом на правой стороне геля и на вторичной структуре РНК, рассчитанной с помощью MFOLD.
https://doi.org/10.1371 / journal.pbio.1000008.g001
Чтобы оценить, функционирует ли Swt1 как эндонуклеаза, использовали структурированный субстрат РНК из 52 нуклеотидов (нт), который содержит стабильную основу (от 5.8S рРНК мыши), которая закрывает 5 ‘- и 3’-концы РНК (рис. 1D). Оставшаяся часть РНК (нуклеотиды 11–43), как предполагается, будет по крайней мере частично одноцепочечной. Swt1 дикого типа, но не мутант Swt1-D135N, эффективно эндонуклеолитически расщеплял структурированную РНК из 52 нуклеотидов с явным предпочтением одноцепочечных областей субстрата РНК (рис. 1D).Взятые вместе, данные показывают, что Swt1 проявляет in vitro эндонуклеазную активность без четкой специфичности последовательности, но с предпочтением одноцепочечной РНК.
Делеция
SWT1 индуцирует накопление ядерного поли (A) + РНК и усиливает утечку цитоплазмы пре-мРНК, дефектных по сплайсингу, в mlp1 Δ и nup60 Δ мутантовДля дальнейшего исследования роли Swt1 in vivo в контексте экспорта мРНК, связанной с транскрипцией, и надзора за ядерной мРНП, мы выполнили анализы экспорта мРНК и утечки пре-мРНК с жизнеспособными двойными мутантами (Рисунок S1), несущими swt1 Δ вместе с thp2 Δ, mft1 Δ (THO / TREX), sus1 Δ (TREX-2), nup60 Δ, mlp1 Δ и esc1 Δ (перинуклеарный контроль качества мРНП).Делеция SWT1 значительно усиливала накопление ядерного поли (A) + РНК по сравнению с одиночными мутантами thp2 Δ, mft1 Δ, sus1 Δ и nup60 Δ (рис. ). Более того, swt1 Δ индуцировал накопление в ядре РНК poly (A) + в мутантах mlp1 Δ и esc1 Δ, что не было обнаружено у единичных мутантов (рис. 2A; см. Также [27,28] ).
Рисунок 2. SWT1 участвует в контроле качества ядерных мРНП с помощью Mlp1-Nup60
(A) Нарушение SWT1 индуцирует или усиливает ядерное накопление поли (A) + и SSA1 РНК в mlΔp1 esc1 Δ или nup60 Δ, соответственно. Указанные штаммы с одним и двумя мутантами выращивали при 30 ° C ( nup60 Δ) или при изменении температуры до 37 ° C в течение 90 мин ( mlp1 Δ и esc1 Δ), а локализация поли (A) + РНК и SSA1 РНК анализировали гибридизацией in situ с использованием Cy3-oligo (dT) и SSA1 -специфических зондов, соответственно.ДНК окрашивали DAPI. Процент клеток, показывающих ядерное накопление поли (A) + РНК или SSA1 мРНК, соответственно, был рассчитан для n > 150 клеток и указан под каждой панелью ( mlp1 Δ и esc1 Δ штаммов). В случае штамма nup60 Δ показан процент клеток со значительно усиленным ядерным сигналом поли (А) + (более чем в 1,5 раза выше средней интенсивности ядерного сигнала nup60 Δ).
(B) Делеция SWT1 вызывает утечку репортерных пре-мРНК в цитоплазму и синергетически усиливает утечку пре-мРНК клеток mlp1 Δ и nup60 Δ клеток. Утечка репортеров пре-мРНК lacZ , которые вызывают активность β-галактозидазы, анализировали, как описано в разделе «Материалы и методы». Отношения между активностями β-галактозидазы, полученными из репортерной пре-мРНК и безинтронной контрольной мРНК, были рассчитаны и показаны относительно отношения соответствующего штамма дикого типа.Показаны результаты четырех независимых экспериментов. Планки погрешностей указывают на стандартные отклонения.
https://doi.org/10.1371/journal.pbio.1000008.g002
Чтобы оценить, отражает ли накопление поли (A) + РНК накопление ядерной мРНК, мы протестировали специфический транскрипт, тепловой шок МРНК SSA1 (рис. 2А). Эта мРНК явно накапливалась в ядре двойного мутанта mpl1Δ swt1 Δ, но не в соответствующих одиночных мутантах.Напротив, никакого ядерного накопления не наблюдалось при анализе ядерного экспорта тРНК (неопубликованные данные). Чтобы количественно определить, увеличились ли уровни транскрипта SSA1 в двойном мутанте mlp1 Δ swt1 Δ, мы измерили количество мРНК SSA1 до и после теплового шока с помощью полимеразной цепной реакции с обратной транскрипцией в реальном времени ( ОТ-ПЦР). Уровни SSA1 были слегка увеличены у одиночных мутантов, но синергетическое увеличение наблюдалось у двойного мутанта mlp1 Δ swt1 Δ по сравнению со штаммом дикого типа (фиг. S4).
Чтобы проверить, приводят ли двойные мутанты swt1Δ и mlp1Δ или nup60Δ к синергическим дефектам в надзоре за пре-мРНП, мы отслеживали утечку цитоплазмы репортеров пре-мРНК [11,12,14]. Утечка репортера с интактными сигналами сплайсинга (pJCR1) не наблюдалась в штамме swt1Δ , тогда как штамм nup60Δ показал повышенную утечку несплицированной пре-мРНК, которая синергетически усиливалась в штамме swt1 Δ nup60 Δ двойной мутант (рис. 2В).Затем мы проверили, проявляют ли репортеры пре-мРНК с мутацией в 5′-сайте сплайсинга (mut5’SS) или в сайте точки ветвления (mutBP) повышенную утечку цитоплазмы в клетках swt1 Δ. Примечательно, что утечка репортера mutBP была увеличена примерно в 4 раза в одинарных мутантах swt1 Δ или mlp1Δ относительно дикого типа (рис. 2В; см. Также [14,16]). Утечка цитоплазмы обоих мутантных репортеров пре-мРНК снова была сильно синергетически усилена в двойном мутанте swt1Δ mlp1Δ (рис. 2В).
Мы пришли к выводу, что потеря Swt1 ведет к увеличению утечки цитоплазмы дефектных по сплайсингу пре-мРНК, что может синергетически усиливаться мутациями в комплексе Mlp1-Nup60. Это указывает на то, что Swt1 и Mlp1-Nup60 играют разные, но частично совпадающие роли в надзоре ядерных мРНП. На первый взгляд кажется противоречивым, что повышенная утечка пре-мРНК сопровождается накоплением в ядре поли (A) + РНК. Однако эти два эффекта не обязательно исключают друг друга.Когда контроль качества ядер серьезно нарушен (например, в клетках mlp1 Δ swt1 Δ или nup60 Δ swt1 Δ), большее количество аберрантных мРНП может накапливаться в ядре и, следовательно, иметь более высокий шанс утечки. .
Эндонуклеазный домен необходим для функции Swt1 на физиологических уровнях экспрессии
В предыдущем исследовании [18] было показано, что Swt1, лишенный PIN (эндонуклеазного) домена, дополняет летальность swt1Δ в мутантных штаммах THO.Однако Swt1 является белком с низким содержанием (см. Ниже), и конструкции были сверхэкспрессированы с относительно сильного промотора NOP1 . Чтобы оценить, является ли эндонуклеазная активность Swt1 существенной для комплементации фенотипов swt1Δ при физиологических уровнях экспрессии, мы проанализировали каталитически неактивный мутант Swt1-D135N, экспрессируемый с эндогенного промотора (рис. 3А). Swt1-D135N и Swt1 дикого типа экспрессировались на аналогичных уровнях (фиг. 3B), но Swt1-D135N не смог компенсировать дефекты роста swt1Δ в любой комбинации двойных мутантов (фиг. 3A и неопубликованные данные).Более того, Swt1-D135N не спасает накопление ядерной мРНК, вызванное swt1Δ на фоне мутаций mlp1 Δ, nup60 Δ, sus1 Δ или thp2 Δ (рис. 3C) и неопубликованные данные. . Наконец, утечка из ядра репортера пре-мРНК (mutBP) в клетки, экспрессирующие Swt1-D135N (рис. 3D), была неотличима от утечки, наблюдаемой в штамме swt1 Δ (рис. 2В). Подобные фенотипы потери функции наблюдались для усеченного мутанта swt1 , лишенного всего PIN-домена (Swt1ΔPIN) (рисунок S5 и неопубликованные данные).
Рисунок 3. Каталитически неактивный мутант swt1-D135N Напоминает нулевую мутацию swt1 In vivo
(A) Мутант swt1-D135N не дополняет синтетически летальные / усиленные фенотипы указанного Δ1 swt штаммы с двойными мутантами. Штаммы трансформировали диким типом SWT1 или мутантом swt1-D135N , и трансформанты наносили пятнами в 10-кратных серийных разведениях на указанные планшеты. Штаммы инкубировали в течение 5 дней при 30 ° C ( tho2 Δ swt1 Δ) или в течение 3 дней при 36 ° C ( thp2 Δ swt1 Δ и nup60 Δ swt1 Δ).
(B) Уровни экспрессии белков Swt1 дикого типа и мутантных белков сходны. Лизаты цельных клеток, полученные из штамма swt1 Δ или штамма swt1 Δ, экспрессирующего Swt1 или Swt1-D135N дикого типа, анализировали с помощью SDS-PAGE и вестерн-блоттинга с использованием поликлональных антител против Swt1. Обнаружение белка Arc1 поликлональным антителом против Arc1 использовали в качестве контроля загрузки.
(C) Мутант Swt1-D135N не может устранить дефект экспорта поли (A) + штамма mlp1 Δ swt1 Δ. mlp1 Δ swt1 Δ Клетки, экспрессирующие Swt1 или Swt1-D135N дикого типа, анализировали на локализацию поли (A) + РНК, как показано на рисунке 2A, или окрашивали DAPI для обнаружения ДНК.
(D) Swt1-D135N-экспрессирующие клетки демонстрируют фенотип утечки ядерной пре-мРНК. Штаммы swt1 Δ и mlp1 Δ swt1 Δ трансформировали указанными плазмидами и анализировали на утечку репортера пре-мРНК lacZ (mutBP), как показано на фиг. 2B.Показаны результаты четырех независимых экспериментов. Планки погрешностей указывают на стандартные отклонения.
(E) Swt1-D135N клетки накапливают повышенные уровни мутированной пре-мРНК lacZ . Тотальную РНК выделяли из swt1-D135N и изогенного штамма дикого типа, экспрессирующего содержащий интрон репортерный транскрипт lacZ (норма . , для нормального) или содержащий интрон транскрипт lacZ , мутировавший в ветви сайт точечного сплайсинга (mut ., для мутировавших). Уровни lacZ количественно определяли с помощью ОТ-ПЦР в реальном времени с использованием мРНК GAL1 в качестве контрольного транскрипта. Показаны результаты четырех независимых экспериментов. Планки погрешностей указывают стандартные ошибки среднего.
(F) Сверхэкспрессия Swt1 дикого типа, но не сверхэкспрессия каталитически неактивного мутанта Swt1-D135N, токсична для клеток. Swt1 Δ клетки, содержащие GAL1 :: SWT1 , GAL1 :: swt1-D135N или пустую плазмиду, наносили пятнами в 10-кратных серийных разведениях на планшеты, содержащие либо галактозу, либо глюкозу, и инкубировали в течение 3 дней при 30 ° C.Показаны только два последних разведения чашки с глюкозой.
(G) Сверхэкспрессия Swt1 дикого типа, но не сверхэкспрессия каталитически неактивного мутанта Swt1-D135N, вызывает сильное ядерное накопление поли (A) + РНК. Те же штаммы, что и в (F), выращивали в среде, содержащей рафинозу, до фазы ранней логарифмической активности, а затем индуцировали 2% галактозой в течение 5 часов. Локализацию РНК поли (А) + анализировали путем гибридизации in situ с использованием зонда Cy3-oligo (dT). ДНК окрашивали DAPI.Показан процент клеток, показывающих накопление ядерного сигнала поли (А) + ( n > 150 клеток).
https://doi.org/10.1371/journal.pbio.1000008.g003
Однако Swt1-D135N, Swt1ΔPIN и другие конструкции с частичной делецией могут дополнять swt1Δ для роста фона tho2Δ при сверхэкспрессии в условиях контроль сильного промотора ADh2 (рисунок S5). Напротив, другие фенотипы swt1Δ , включая утечку ядерной пре-мРНК и накопление ядерной РНК poly (A) + в штаммах mlp1Δ swt1Δ , не могли быть подавлены сверхэкспрессией Swt1-D135N или Swt1ΔPIN мутантов из ADh2. Промотор (рисунок 3D и неопубликованные данные).Таким образом, при физиологических уровнях экспрессии эндонуклеазная активность Swt1 важна для его функции, тогда как потребность в каталитической активности может быть частично устранена сверхэкспрессией неактивного белка Swt1.
Swt1-D135N Клетки накапливают повышенные уровни аберрантной пре-мРНКЧтобы оценить возможную роль эндонуклеазной активности Swt1 в деградации дефектных мРНП, мы определили, стабилизируется ли «аберрантный» транскрипт в клетках swt1-D135N по сравнению с изогенными клетками дикого типа.В качестве аномального транскрипта мы использовали интрон-содержащую конструкцию lacZ (экспрессируемую из плазмиды mutBP; см. Выше), которую использовали в анализах утечки пре-мРНК. Это таит в себе мутацию в сайте точки ветвления сплайсинга, которая эффективно блокирует сплайсинг пре-мРНК [12]. Содержащий интрон дикого типа транскрипт lacZ , кодируемый плазмидой pJCR51, использовали в качестве контрольной пре-мРНК. Накопление аберрантного транскрипта было существенно повышено в клетках swt1-D135N по сравнению со штаммом дикого типа (mut.столбцы на фиг. 3E), тогда как уровни нормально процессируемой мРНК lacZ не изменились между штаммами swt1-D135N и диким типом (нормальные столбцы на фиг. 3E). Эти результаты предполагают, что эндорибонуклеазная активность Swt1 участвует в селективном устранении дефектных транскриптов мРНК.
Сверхэкспрессия Swt1 токсична и вызывает сильное ядерное накопление пре-мРНК
Поскольку Swt1 участвует в надзоре за ядерными мРНП, его экспрессия и / или нацеливание могут регулироваться.Количественный вестерн-блот-анализ с использованием антител против Swt1 показал, что in vivo уровни эндонуклеазы Swt1 очень низкие (~ 200–300 молекул на клетку) (неопубликованные данные; см. Также [18]), что может быть важно для предотвращения несоответствующей деградации РНК. В крупномасштабном анализе сообщалось, что Yor166 / Swt1 токсичен после сверхэкспрессии системой Tet-off [29]. В соответствии с этим открытием, доминантно-летальный фенотип был индуцирован сверхэкспрессией Swt1 дикого типа, но не каталитически неактивного мутанта Swt1-D135N с промотора GAL1 (рис. 3F).Большинство сверхэкспрессирующих Swt1 клеток показали сильное ядерное накопление как поли (A) + РНК (рис. 3E), так и мРНК SSA1 , тогда как тРНК обычно экспортировалась (рис. S3B и неопубликованные данные). Примечательно, что при анализе локализации репортерных конструкций lacZ , содержащих интрон / без интрона, накопление в ядре репортера пре-мРНК, содержащего интрон, было сильнее, чем для мРНК без интрона (рис. S3C), что согласуется с дефектом в пре-мРНК. надзор за мРНК.Эти анализы демонстрируют, что сверхэкспрессия каталитически активного Swt1 токсична и мешает экспорту мРНК и / или надзору пре-мРНК.
Swt1 ассоциирует с комплексами ядерных пор в
nup133 Δ клеткахСубклеточное фракционирование ранее показало, что меченный TAP Swt1 расположен в ядре [18]. Точное расположение Swt1 не может быть определено, однако, потому что количество зеленого флуоресцентного белка (GFP) или иммуно-меченого Swt1, экспрессируемого с эндогенного промотора, было слишком низким для визуализации (неопубликованные данные и [18]).Присоединение нескольких тегов GFP может позволить обнаруживать белки с низким содержанием (N. Daigle, A. Bancaud и J. Ellenberg, неопубликованные данные). Таким образом, мы создали функциональную конструкцию 4xGFP-Swt1, экспрессируемую с эндогенного промотора SWT1 , которая дополняла штаммы с двойной делецией swt1 Δ mlp1 Δ, swt1 Δ nup60 Δ или Δ17 swt1 Δ (Рисунок S6 и неопубликованные данные). При анализе в штамме swt1 Δ, 4xGFP-Swt1 был локализован как в ядре, так и в цитоплазме с небольшим ядерным обогащением в некоторых клетках (рис. 4A).При экспрессии в мутанте nup133 Δ с кластеризацией ядерных пор [30], 4xGFP-Swt1 проявлял значительную совместную кластеризацию с NPC на периферии ядра, которая терялась при комплементации штамма nup133 Δ плазмидным производным NUP133 ( Рисунок 4A). Когда конструкция 4xGFP-Swt1, лишенная каталитического PIN-домена (4xGFP-Swt1ΔPIN), экспрессировалась в клетках swt1 Δ, она концентрировалась в нескольких точках. Они всегда были близки к NPC, как показала совместная визуализация Nup120-mCherry (фиг. 4B), и 4xGFP-Swt1ΔPIN, также связанный с NPC в nup133 Δ клетках.Нацеливание Swt1ΔPIN на NPC не зависело от Mlp1, Nup60 или Esc1. Вместо этого 4xGFP-Swt1ΔPIN показал более отчетливое и пунктированное окрашивание ядерной оболочки в штаммах mlp1 Δ, nup60 Δ и esc1 Δ, чем в соответствующих изогенных штаммах дикого типа (рисунок 4B и неопубликованные данные). Нуклеоцитоплазматическая локализация полноразмерного 4xGFP-Swt1 в клетках mlp1 Δ, nup60 Δ и esc1 Δ не изменилась (неопубликованные данные).Вместе с генетическими данными эти наблюдения показывают, что Swt1 взаимодействует с NPC.
Рисунок 4. Swt1 ассоциирует с комплексами ядерных пор в нуклеопориновом мутанте nup133 Δ
(A) 4xGFP-Swt1 (зеленый) или (B) 4xGFP-Swt1ΔPIN (зеленый) коэкспрессировались с Nup120-mCherry (маркер для Nup120-mCherry). ; красный) в указанных штаммах ( swt1 Δ, nup133 Δ swt1 Δ, nup133 Δ swt1 Δ + NUP133 [экспрессируется из центромерной плазмиды] Δ1 Δ1 , и nup60 Δ swt1 Δ) и анализировали с помощью флуоресцентной микроскопии.Клетки также просматривались оптикой Номарского.
https://doi.org/10.1371/journal.pbio.1000008.g004
Discussion
В этом исследовании мы показываем, что Swt1 представляет собой РНК-эндонуклеазу, которая может связываться с NPCs и участвует в системах, которые контролируют качество ядерных мРНП перед экспортом в цитоплазму. Эндонуклеазная активность PIN-домена Swt1 имеет решающее значение для его функции in vivo на физиологических (то есть низких) уровнях экспрессии. Сильно сверхэкспрессированные конструкции Swt1, лишенные PIN-домена, могли выполнять некоторые из функций Swt1 in vivo, тогда как сверхэкспрессия активного Swt1 была летальной.Это указывает на то, что Swt1 функционирует и как нуклеаза, и как белковый компонент комплексов наблюдения мРНП.
Чтобы избежать цитоплазматической трансляции дефектных мРНП в потенциально вредные белки, системы биогенеза и наблюдения мРНП взаимодействуют в ядре. Эндорибонуклеазная активность Swt1 важна, когда функция комплексов, котранскрипционно формирующих и проверяющих ядерные мРНП, нарушена, это указывает на то, что Swt1 может участвовать в деградации компонента мРНК аберрантных ядерных мРНП.Эндонуклеазная активность может быть важной, потому что транскрипционно захваченные мРНП, экспортно-некомпетентные мРНП или пре-мРНП, неспособные к сплайсингу, лишены всех доступных концов РНК, которые могли бы способствовать деградации ядерной 5′-экзонуклеазой Rat1 или 3′-экзонуклеазной активностью экзосомы. . Эндонуклеазное расщепление дефектной мРНК и мРНП с помощью Swt1 может генерировать свободные концы мРНК для дальнейшей экзонуклеолитической деградации. В соответствии с этой моделью, аберрантная репортерная конструкция пре-мРНК, которая не может быть сплайсирована, избирательно накапливалась в каталитически неактивном мутанте Swt1-D135N, и потеря Swt1 из штамма mlp1Δ стабилизировала ограниченную ядерной реакцией мРНК SSA1 .
Высокие уровни Swt1 были токсичными и индуцировали сильное ядерное накопление поли (A) + РНК. Среди нескольких возможностей, которые могут объяснить этот фенотип, сверхэкспрессия Swt1 может вызывать ошибочное расщепление РНК в правильно сформированных комплексах RNP. По нашим оценкам, количество молекул Swt1 в клетке составляет всего около 200–300 копий, и этот низкий уровень может быть важным для подавления расщепления нецелевой РНК. Накопление в ядре интрон-содержащей репортерной пре-мРНК, индуцированной сверхэкспрессией Swt1, было сильнее, чем для соответствующей безинтронной пре-мРНК, что согласуется с дефектом в наблюдении за пре-мРНК.Примечательно, что сверхэкспрессия Mlp1, но не делеция его гена, также вызывает накопление в ядре интрон-содержащих пре-мРНК [14,28], что согласуется с моделью, согласно которой Swt1 и Mlp1 функционируют вместе в этом пути.
Наши данные согласуются с ранее предполагаемой ролью Swt1 в транскрипции мРНК с помощью TREX [18]. Комплексы TREX / TREX-2 необходимы для котранскрипционной сборки мРНП, и нарушение их функции приводит к образованию аберрантных структур мРНП, которые препятствуют транскрипции RNAPII [3–5,9].Мы прогнозируем, что эти дефектные частицы мРНП представляют собой важные субстраты для деградации эндонуклеаз с помощью Swt1. В соответствии с этой моделью, дефектные и некомпетентные к экспорту мРНП, сформированные в мутантах TREX, накапливаются вместе с NPCs [9,31], где также может быть обнаружен Swt1.
В клетках дикого типа Swt1 распределялся между ядром и цитоплазмой и, следовательно, потенциально мог детектировать образование дефектных пре-мРНП на всем пути биогенеза мРНП. Однако Swt1 накапливался в NPC, когда они были кластеризованы в мутанте nup133 Δ [30].Эта локализация Swt1 в NPC не может быть коррелирована с нарушением экспорта мРНК ( nup133, Δ клетки также обнаруживают дефект экспорта мРНК), поскольку 4xGFP-Swt1 не накапливался в ядерной оболочке у мутантного экспортного мРНК mex67-5 (неопубликовано). данные). Более того, Swt1, лишенный своего эндонуклеазного домена, стал концентрироваться в перинуклеарных пятнах даже в клетках с нормальным набором нуклеопоринов. Мы предполагаем, что Swt1 временно рекрутируется в NPC через взаимодействия с нуклеопоринами и / или с экспортно-некомпетентными пре-мРНП, заблокированными на NPC, и функционирует там, чтобы предотвратить несоответствующий экспорт пре-мРНК.
Анализ ядерной деградации дефектных РНК у дрожжей в основном сосредоточен на активности 5′- и 3′-экзонуклеаз. Эта работа демонстрирует, что расщепление эндонуклеазой также может играть важную роль в надзоре за ядерной РНК.
Материалы и методы
Штаммы дрожжей, плазмиды и генетические манипуляции.
Штаммы дрожжей, использованные в этом исследовании, перечислены в таблице S1; плазмидные конструкции описаны в таблице S2. Сайт-направленный мутагенез гена SWT1 для создания мутанта swt1-D135N был выполнен с помощью гибридной ПЦР [32].Скрининг синтетической летальности со штаммом sac3 Δ проводили, как описано ранее [33].
Анализ нуклеазной активности in vitro.
Для измерения активности нуклеазы Swt1 in vitro, белок FLAG-Swt1 дикого типа и мутантный белок D135N были экспрессированы из плазмид pADh211-FLAG- SWT1 или pADh211-FLAG- swt1 — D135N ( ADh2). и очищают аффинной хроматографией против FLAG. Вкратце, собирали 4 мкл дрожжевой культуры с оптической плотностью при 600 нм (OD 600 ) приблизительно 4.0, ресуспендировали в буфере FLAG (50 мМ Трис-HCl [pH 7,5], 200 мМ NaCl, 0,5 мМ DTT, 0,05% NP-40) до конечного объема 50 мл, лизировали взбиванием шариков (Pulverissete 6; Fritsch) и инкубировали с 120 мкл агарозы anti-FLAG M2 (Sigma) в течение 120 мин при 4 ° C. Суспензию анти-FLAG промывали 750 объемами буфера FLAG, и связанный материал элюировали пептидом FLAG в соответствии с инструкциями производителя (Sigma) с последующим концентрированием и заменой буфера.
Анализы нуклеазной активности очищенных белков на 5′-конце или 3′-конце радиоактивно меченых РНК проводили при 30 ° C в 15 мМ Трис-HCl (pH 7.6), 75 мМ NaCl, 2 мМ DTT, 100 мкг / мл БСА, 0,8 Ед / мкл РНКазина, 4,5% глицерина, 0,05% NP-40, 0,3 мкМ тРНК Escherichia coli и 7,5 мМ MnCl 2 . Перед добавлением 20–30 фмоль меченого субстрата РНК 20 или 30 мкл реакционной смеси, содержащей 10 пмоль белка, предварительно инкубировали в течение 15 мин при 30 ° C. Затем отбирали аликвоты по 4 мкл в моменты времени, указанные на фиг. 1, и реакцию останавливали добавлением буфера для загрузки РНК, содержащего формамид. Продукты реакции разделяли на денатурирующем геле 12% полиакриламид / 8 М мочевина и визуализировали с помощью авторадиографии.5 ‘- [ 32 P] — и 3′ — [ 32 P] -меченные субстраты РНК (Dharmacon) метили и очищали в геле, как описано [34].
Анализ утечки пре-мРНК.
По крайней мере, четыре независимых эксперимента были выполнены для каждого штамма, содержащего указанный репортер утечки пре-мРНК lacZ (pJCR1, pmut5’SS и pmutBP) или безинтронный контроль lacZ (pLGSD5). Трансформанты объединяли и выращивали в течение ночи в селективной среде раффинозы. Клетки разбавляли, выращивали до середины логарифмической фазы и экспрессию репортера индуцировали 2% галактозой в течение 150 мин.Анализы β-галактозидазы проводили в трех экземплярах для каждого образца в соответствии с опубликованным протоколом [11] с модификациями, описанными в [16].
Анализ уровней РНК
SSA1 и lacZ .Для индукции экспрессии SSA1 к экспоненциально выросшим культурам применяли тепловой шок (42 ° C в течение 15 мин) путем добавления равного объема предварительно нагретой среды. Экспрессию репортеров пре-мРНК lacZ (pmutBP или pJCR51) индуцировали добавлением галактозы (конечная концентрация 2%) к культурам в ранней логарифмической фазе и их последующей инкубацией в минимальной среде галактоза-раффиноза в течение 6 часов.Клетки лизировали стеклянными шариками и встряхиванием, и общую РНК выделяли с помощью набора RNeasy Mini Kit (QIAGEN). РНК подвергали обратной транскрипции с использованием набора для обратной транскрипции QuatiTect (QIAGEN), а уровни специфического транскрипта РНК измеряли с помощью количественной ПЦР в реальном времени с использованием системы ABI Prism 7000 (Applied Biosystems) и ABsolute QPCR ROX Mix (ABgene). Каждый образец кДНК, а также контроли без обратной транскриптазы, амплифицировали в трех экземплярах с наборами праймеров и зондов, описанными в таблице S3.Уровни SSA1 и lacZ РНК были нормализованы до уровней мРНК RPB4 и GAL1 соответственно, которые, как было обнаружено, не изменились в испытанных условиях.
Разное.
Флуоресцентная гибридизация in situ (FISH) с использованием олиго (dT), SSA1, транскрипт-, тРНК- и lacZ -специфических Cy3-меченных зондов выполняли, как описано [33,35,14]. Локализацию слияний 4xGFP-Swt1, 4xGFP-Swt1ΔPIN и Nup120-mCherry анализировали в Zeiss Axio Imager.Микроскоп Z1 с программным обеспечением AxioVision 4.6.3. Экспрессию Swt1 анализировали иммуноблоттингом с использованием кроличьей поликлональной сыворотки против Swt1, созданной против рекомбинантного Swt1 (PickCell Laboratories), и конъюгата против кроличьей HRP в качестве вторичного антитела (BIO-RAD). Экспрессию вариантов белка Swt1, меченных FLAG, анализировали с помощью конъюгата анти-FLAG-HRP (Sigma).
Вспомогательная информация
Рисунок S1. Генетические взаимодействия между
SWT1 и факторами сборки котранскрипционных мРНП и контролем качества перинуклеарных мРНПУказанные штаммы дикого типа, одиночные и двойные мутанты были обнаружены в 10-кратных серийных разведениях на 5′-фтороротовой кислоте (FOA ) -содержащие чашки в случае синтетического летального взаимодействия (левая панель) и на пластинах с синтетической декстрозой (SDC) -Leu после перетасовки 5′-FOA в случае синергетически усиленного дефекта роста (средняя и правая панели).Планшеты инкубировали в течение 5 дней при 30 ° C (левая панель) или 3 дня при 30 ° C или 36 ° C (средняя и правая панели).
https://doi.org/10.1371/journal.pbio.1000008.sg001
(2,23 МБ PDF)
Рисунок S2. Сравнение последовательностей PIN-доменов белков Swt1 и SMG6 / EST1A
Последовательности PIN-доменов белков Swt1 из Saccharomyces cerevisiae (Swt1; Yor166c), Schizosaccharomyces pombe (SpSwt1; Spcc24b10.15p), Dro206mGe и Homo sapiens (HsSwt1; C1orf26) были выровнены с PIN-доменами Drosophila и человеческих белков SMG6 / EST1A (DmSmg6, HsSmg6) с помощью ClustalW.Звездочки указывают на консервативные кислотные остатки, участвующие в нуклеазной активности PIN-домена. Указаны положения соответствующих остатков Swt1.
https://doi.org/10.1371/journal.pbio.1000008.sg002
(88 КБ PDF)
Рисунок S3. Накопление ядерной мРНК после делеции и сверхэкспрессии Swt1
(A) Делеция SWT1 усиливает накопление в ядре поли (A) + РНК мутантов TREX-2 и TREX. Указанные штаммы с одним и двумя мутантами были переведены на температуру 37 ° C на 90 мин, и локализация поли (A) + РНК была проанализирована гибридизацией in situ с использованием зонда Cy3-oligo (dT).ДНК окрашивали DAPI. Показан процент клеток, показывающих значительно усиленный ядерный сигнал поли (А) + (более 1,5-кратный средней интенсивности ядерного сигнала соответствующего одиночного мутантного штамма) ( n > 150 клеток).
(B) Сверхэкспрессия Swt1 запускает накопление в ядре мРНК SSA1 . Swt1 Δ клетки или swt1 Δ клетки, экспрессирующие GAL1 :: SWT1 , выращивали в селективной среде раффинозы до фазы раннего логарифма, индуцированной 2% галактозой в течение 5 часов, и была обнаружена локализация мРНК SSA1 гибридизацией in situ с использованием специфичного для SSA1 , Cy3-меченного зонда.ДНК окрашивали DAPI. Был рассчитан процент клеток, демонстрирующих накопление в ядре сигнала SSA1 ( n > 150 клеток), и он показан ниже.
(C) Сверхэкспрессия Swt1 вызывает накопление в ядре репортеров мРНК lacZ . Безинтронная (pLGSD5) или содержащая интрон (pJCR51) мРНК lacZ была экспрессирована в штамме swt1 Δ или GAL1 :: SWT1 дикого типа, и ее распределение анализировали, как в (B), с использованием штамма lacZ -специфический зонд.В каждом случае было исследовано не менее 300 клеток, и указан процент клеток с ядерным накоплением сигнала lacZ .
https://doi.org/10.1371/journal.pbio.1000008.sg003
(2,30 МБ PDF)
Рисунок S4.
mlp1Δ swt1Δ Двойное разрушение приводит к повышенным клеточным уровням SSA1 Транскрипт после теплового шокаОбщая РНК была выделена из указанных культур в средней логарифмической фазе до (25 ° C) или после теплового шока до 42 ° C в течение 15 мин.Уровни SSA1 количественно определяли с помощью анализа RT-PCR в реальном времени с использованием мРНК RPB4 в качестве контрольного транскрипта. Показаны репрезентативные результаты двух независимых экспериментов.
https://doi.org/10.1371/journal.pbio.1000008.sg004
(65 КБ PDF)
Рисунок S5. Анализ делеции Swt1
(A) Схема конструкций Swt1, используемых для анализа делеции. Цифры указывают положение соответствующей аминокислоты вдоль белка Swt1.
(B) Анализ удаления Swt1.Последовательность перетасовки tho2Δ swt1Δ трансформировали указанными делеционными конструкциями SWT1 , экспрессировавшимися из эндогенного SWT1 или сильного промотора ADh2 , и способность к комплементации тестировали путем роста на пластинах 5′-FOA при 30 ° C для 5 дн.
https://doi.org/10.1371/journal.pbio.1000008.sg005
(515 КБ PDF)
Рисунок S6. Характеристика слитых белков 4xGFP-Swt1 и 4xGFP-Swt1ΔPIN
(A) Слияние 4xGFP-Swt1 дополняет синтетически усиленные фенотипы штаммов mlp Δ swt1 Δ и nup60 Δ Δ мутантных штаммов swt.Штаммы трансформировали плазмидой YCplac111-FLAG-4xGFP- SWT1 , YCplac111-FLAG- SWT1 или пустой плазмидой, и трансформанты наносили пятнами в 10-кратных серийных разведениях на планшеты SDC-Leu. Штаммы инкубировали 5 дней при указанных температурах.
(B) Сравнение уровней экспрессии белков Swt1, Swt1ΔPIN, 4xGFP-Swt1 и 4xGFP-Swt1ΔPIN. Лизаты цельных клеток получали из штаммов swt1 Δ или mlp Δ swt1 Δ, экспрессирующих указанные конструкции из центромерной плазмиды YCplac111, и анализировали с помощью SDS-PAGE и вестерн-блоттинга с использованием моноклональных антител против FLAG.Звездочка обозначает белок в клеточном лизате, перекрестно реагирующий с конъюгатом анти-FLAG.
https://doi.org/10.1371/journal.pbio.1000008.sg006
(457 КБ PDF)
Благодарности
Выражаем признательность Р. Кунце за отличную техническую помощь. Мы хотим поблагодарить А. Корбетта, Н. Дейгла, А. Банко, Дж. Элленберга, О. Гадала, В. Гали, С. Грунда, Г. Хартцога, Д. Кресслера, П. Стелтера и Ф. Штутца за дрожжи. штаммы, плазмиды и реагенты.
Вклад авторов
MS, CS, DT и EH разработали и разработали эксперименты.MS, CS, AR и JW проводили эксперименты. MS, CS, JW, DT и EH проанализировали данные. MS, CS, DT и EH написали статью.
Ссылки
- 1. Köhler A, Hurt E (2007) Экспорт РНК из ядра в цитоплазму. Nat Rev Mol Cell Biol 8: 761–773.
- 2. Rougemaille M, Villa T, Gudipati RK, Libri D (2008) Путешествие мРНК в цитоплазму: требуется одежда. Biol Cell 100: 327–342.
- 3. Huertas P, Aguilera A (2003) ДНК, образованная котранскрипцией: гибриды РНК опосредуют нарушение элонгации транскрипции и рекомбинацию, связанную с транскрипцией.Mol Cell 12: 711–721.
- 4. Gonzales-Aguilera C, Tous C, Gomez-Gonzales B, Huertas P, Luna R и др. (2008) Комплекс THP1-SAC3-SUS1-CDC31 работает в экспорте мРНК элонгации транскрипции, предотвращая РНК-опосредованную нестабильность генома. Mol Biol Cell 10: 4310–4318.
- 5. Сагес К., Шмид М., Олесен Дж. Р., Гази М.А., Ку X и др. (2008) Надзор за ядерной мРНК у мутантов THO / sub2 запускается неэффективным полиаденилированием. Mol Cell 31: 91–103.
- 6.Strässer K, Masuda S, Mason P, Pfannstiel J, Oppizzi M и др. (2002) TREX — это консервативный комплекс, связывающий транскрипцию с экспортом информационной РНК. Природа 417: 304–308.
- 7. Фишер Т., Стрэссер К., Рац А., Родригес-Наварро С., Оппицци М. и др. (2002) Механизм экспорта мРНК требует, чтобы новый комплекс Sac3p-Thp1p стыковался с нуклеоплазматическим входом в ядерные поры. EMBO J 21: 5843–5852.
- 8. Fasken MB, Corbett AH (2005) Процесс или смерть: контроль качества в биогенезе мРНК.Nat Struct Mol Biol 12: 482–488.
- 9. Villa T, Rougemaille M, Libri D (2008) Ядерный контроль качества рибонуклеопротеинов РНК-полимеразы II в дрожжах: изменение баланса для формирования транскриптома. Biochim Biophys Acta 1779: 524–531.
- 10. Rougemaille M, Gudipati RK, Olesen JR, Thomsen R, Seraphin B и др. (2007) Рассмотрение механизмов надзора за ядерной мРНК у мутантов комплекса THO / sub2. EMBO J 26: 2317–2326.
- 11. Legrain P, Rosbash M (1989) Некоторые цис- и транс-действующие мутанты для сплайсинга пре-мРНК-мишени в цитоплазму.Cell 57: 573–583.
- 12. Rain JC, Legrain P (1997) Обязательство in vivo сплайсинга в дрожжах включает нуклеотид, расположенный выше консервативной последовательности сайта разветвления, и белок Mud2. EMBO J 16: 1759–1771.
- 13. Rutz B, Séraphin B (2000) Двойная роль BBP / ScSF1 в удерживании и сплайсинге ядерной пре-мРНК. EMBO J 19: 1873–1886.
- 14. Galy V, Gadal O, Fromont-Racine M, Romano A, Jacquier A, Nehrbass U (2004) Ядерное удержание несплицированных мРНК в дрожжах опосредуется перинуклеарным Mlp1.Ячейка 116: 63–73.
- 15. Vinciguerra P, Iglesias N, Camblong J, Zenklusen D, Stutz F (2005) Перинуклеарные белки Mlp подавляют экспрессию генов в ответ на дефект в экспорте мРНК. EMBO J 24: 813–823.
- 16. Palancade B, Zuccolo M, Loeillet S, Nicolas A, Doye V (2005) Pml39, новый белок ядерной периферии, необходимый для удержания в ядре неправильных рибонуклеарных частиц-мессенджеров. Mol Biol Cell 16: 5258–5268.
- 17. Lewis A, Felberbaum R, Hochstrasser M (2007) Белок ядерной оболочки, связывающий сборку корзины ядерных пор, регуляцию протеазы SUMO и наблюдение за мРНК.J Cell Biol 178: 813–822.
- 18. Röther S, Clausing E, Kieser A, Strässer K (2006) Swt1, новый дрожжевой белок, участвует в транскрипции. J Biol Chem 281: 36518–36525.
- 19. Клиссолд П.М., Ponting CP (2000) ПИН-домены в нонсенс-опосредованном распаде мРНК и РНКи. Curr Biol 10: R888–890.
- 20. Anantharaman V, Aravind L (2006) Домены NYN: новый предсказал РНКазы со складкой, подобной домену PIN. РНК Биол 3: 18–27.
- 21.Jeyakanthan J, Inagaki E, Kuroishi C, Tahirov TH (2005) Структура PIN-домена белка PH0500 из Pyrococcus horikoshii. Acta Crystallograph Sect F Struct Biol Cryst Commun 61: 463–468.
- 22. Glavan F, Behm-Ansmant I, Izaurralde E, Conti E (2006) Структуры PIN-доменов SMG6 и SMG5 выявляют нуклеазу в комплексе наблюдения за мРНК. EMBO J 25: 5117–5125.
- 23. Fatica A, Tollervey D, Dlakic M (2004) PIN-домен Nob1p необходим для расщепления D-сайта в пре-рРНК 20S.РНК 10: 1698–1701.
- 24. Bleichert F, Granneman S, Osheim YN, Beyer AL, Baserga SJ (2006) Белок домена PINc Utp24, предполагаемая нуклеаза, необходим для ранних стадий расщепления при созревании 18S рРНК. Proc Natl Acad Sci U S A 103: 9464–9469.
- 25. Lei EP, Krebber H, Silver PA (2001) Информационные РНК рекрутируются для ядерного экспорта во время транскрипции. Genes Dev 15: 1771–1782.
- 26. Gilbert W, Guthrie C (2004) Ядерная фосфатаза Glc7p способствует экспорту мРНК, облегчая ассоциацию Mex67p с мРНК.Mol Cell 13: 201–212.
- 27. Strambio-de-Castillia C, Blobel G, Rout MP (1999) Белки, соединяющие комплекс ядерной поры с внутренним пространством ядра. J Cell Biol 144: 839–55.
- 28. Косова Б., Панте Н., Ролленхаген С., Подтелейников А., Манн М. и др. (2000) Mlp2p, компонент внутриядерных филаментов, прикрепленных к ядерным порам, ассоциируется с Nic96p. J Biol Chem 275: 343–350.
- 29. Бойер Дж., Бадис Дж., Фэрхед С., Талла Е., Хантрей Ф. и др.(2004) Крупномасштабное исследование ингибирования роста, вызванного сверхэкспрессией геномных фрагментов в Saccharomyces cerevisiae. Геном Биол 5: R72.
- 30. Doye V, Wepf R, Hurt EC (1994) Новый белок ядерной поры Nup133p с различными ролями в транспорте поли (A) + РНК и распределении ядерных пор. EMBO J 13: 6062–6075.
- 31. Rougemaille M, Dieppois G, Kisseleva-Romanova E, Gudipati RK, Lemoine S, et al. (2008) THO / Sub2p функционирует, чтобы координировать процессинг 3′-конца с ассоциацией ген-ядерной поры.Ячейка 135: 308–321.
- 32. Ho SN, Hunt HD, Horton RM, Pullen JK, Pease LR (1989) Сайт-направленный мутагенез путем перекрывания удлинения с использованием полимеразной цепной реакции. Gene 77: 51–59.
- 33. Strässer K, Hurt E (2001) Фактор сплайсинга Sub2p необходим для экспорта ядерной мРНК через его взаимодействие с Yra1p. Природа 413: 648–652.
- 34. Mitchell P, Petfalski E, Tollervey D (1996) 3′-конец дрожжевой 5.8S рРНК генерируется с помощью механизма процессинга экзонуклеаз.Genes Dev 10: 502–513.
- 35. Long RM, Elliott DJ, Stutz F, Rosbash M, Singer RH (1995) Пространственные последствия дефектной обработки специфических дрожжевых мРНК, выявленные флуоресцентной гибридизацией in situ. РНК 1: 1071–1078.
(PDF) Global Entrepreneurship Monitor: Отчет по Грузии за 2016-2017 гг.
4
Список цифр
Рисунок 1. Отношение населения к предпринимательству в Грузии и странах ЕС, 2016 г…….. 15
Рисунок 2. Сравнение процентного отношения населения к высокому статусу и
успешных предпринимателей и предпринимательства как желательного выбора карьеры …………… .16
Рисунок 3. Отношение к предпринимательству в Грузии и других странах, ориентированных на эффективность,
Европейские страны, 2014 и 2016 годы …………………… ………………………………………….. … 17
Рисунок 4. Предполагаемые возможности и предполагаемые возможности в Грузии и другие показатели эффективности —
европейских стран, 2016 г…………………………………………… …………………………. 20
Рисунок 5. Индивидуальное отношение к предпринимательству в Грузии и странах ЕС,
2016 ………………………………………….. ………………………………………….. ……………………… 21
Рисунок 6. Предпринимательские намерения, предполагаемые возможности, предполагаемые возможности и страх провала
в Грузии и Страны ЕС, 2016 г…………………………………………… ……………. 23
Диаграмма 7. Восприятие предпринимателей в Грузии и других ориентированных на эффективность европейских странах
, 2014 и 2016 гг ……….. ………………………………………….. ……………………………. 25
Диаграмма 8. Уровень зарождающегося предпринимательства, уровень владения новым бизнесом и TEA в Грузии и
странах ЕС, 2016 ………………………………….. ………………………………………………. ……. 27
Рисунок 9. Предпринимательская деятельность в Грузии и других странах Европы, ориентированных на повышение эффективности,
, 2014 и 2016 годы ……………….. ………………………………………….. …………………… 29
Диаграмма 10. Доля предпринимателей, мотивированных необходимостью и возможностями, в
европейских странах, ориентированных на эффективность. ……………………………………………………………………………….. 33
Рисунок 11. Доля предпринимателей, мотивированных необходимостью и возможностями, в Грузии, 2014 г.
и 2016 г …………………………. ………………………………………….. ……………………………….. 35
Рисунок 12. Прекращение деятельности (% от TEA) в европейских странах, ориентированных на эффективность,
2016 ……………………………….. ………………………………………………………………………. …… 36
Рис. 13. Основные причины прекращения деятельности в
европейских странах, ориентированных на эффективность, в 2016 году …………………. ………………………………………….. ………………………………. 37
Рисунок 14. Основные причины прекращения бизнеса в Грузия, 2014 и 2016 годы …………… 37
Рисунок 15. Предпринимательская активность сотрудников в европейских странах, ориентированных на эффективность, 2016 год….. 38
Рисунок 16. Предпринимательская деятельность сотрудников в европейских странах, ориентированных на эффективность, 2014 г. и
2016 г. …………………….. ………………………………………….. ………………………………………….. 39
Рисунок 17. Ожидаемый уровень создания рабочих мест (в процентах от TEA) в
европейских странах, ориентированных на эффективность, в 2016 году …………………. ………………………………………….. ………………… 40
Рисунок 18. Ожидаемый уровень создания рабочих мест (в процентах от TEA) в Грузии, 2014 и 2016 годы
……………… ………………………………………….. ………………………………………….. …………….. 41
Рисунок 19. Уровень инноваций для TEA в европейских странах, ориентированных на эффективность, 2016 …………… …… 43
Рисунок 20. Уровень инноваций для TEA в Грузии, 2014 и 2016 годы ………………………. …………………. 43
Рисунок 21. Международное предпринимательство в европейских странах, ориентированных на эффективность, 2016
………. ………………………………………….. ………………………………………….. ……………………. 45
Рисунок 22. Международное предпринимательство в Грузии, 2014 и 2016 годы ……… ………………. 46
Рисунок 23. Возрастная группа TEA (процент населения в возрасте 18–64 лет), 2016 г……………………….. 50
Диаграмма 24. Доля респондентов в TEA, имеющих как минимум высшее образование …. …… 51
Рисунок 25. Индикаторы рамочных условий предпринимательства в Грузии с рейтингами ……… 54
Рисунок 26. Рамочные условия предпринимательства в Грузии и среднее значение выбранной эффективности
страны …………………………………………. …………………………………….54
Эндорибонуклеаза, функционально связанная с перинуклеарными мРНП, связанная с контролем качества с комплексами ядерных пор
Abstract
Экспорт ядерной мРНК является решающим этапом в экспрессии эукариотических генов, которая в дрожжах связана с сборкой транскрипционной мессенджерской частицы рибонуклеопротеина (мРНП) и наблюдение. Было описано несколько систем наблюдения, которые отслеживают биогенез и экспорт ядерных мРНП, но механизм, с помощью которого распознаются и устраняются неподходящие мРНП, остается плохо изученным.Здесь мы сообщаем, что консервативный белок PIN-домена Swt1 представляет собой РНК-эндонуклеазу, которая участвует в контроле качества ядерных мРНП и может связываться с комплексом ядерных пор (NPC). Swt1 проявлял эндорибонуклеазную активность in vitro, которая ингибировалась точечной мутацией в предсказанном каталитическом сайте. Swt1 не обладал четкой специфичностью последовательности, но демонстрировал сильное предпочтение одноцепочечным областям. Генетические взаимодействия были обнаружены между Swt1 и комплексами THO / TREX и TREX-2, а также с компонентами перинуклеарной системы наблюдения мРНП, Mlp1, Nup60 и Esc1.Ингибирование нуклеазной активности Swt1 увеличивало уровни и цитоплазматическую утечку несплайсированной аберрантной пре-мРНК и индуцировало устойчивое ядерное накопление РНК поли (A) + в штаммах mlp1 Δ и esc1 Δ. Сверхэкспрессия Swt1 также вызывала сильное накопление РНК поли (А) + в ядре. Swt1 обычно распределяется по ядру и цитоплазме, но концентрируется в комплексах ядерных пор (NPCs) у мутанта nup133 Δ, что вызывает кластеризацию NPC и дефекты в экспорте мРНП.Данные предполагают, что эндорибонуклеаза Swt1 может временно рекрутироваться в NPCs, чтобы инициировать деградацию дефектных пре-мРНП или мРНП, захваченных на периферии ядра, чтобы избежать их цитоплазматического экспорта и трансляции.
Введение
Ядерный экспорт информационной РНК (мРНК) является фундаментальным процессом экспрессии эукариотических генов. В дрожжах вновь синтезированные пре-мРНК котранскрипционно собираются в частицы рибонуклеопротеина-мессенджера (мРНП) с помощью комплексов экспорта-транскрипции (TREX).Комплексы THO / TREX (Tho2-Hpr1-Mft1-Thp2-Tex1-Sub2-Yra1) и TREX-2 (Sac3-Thp1-Sus1-Cdc31) связывают транскрипцию мРНК с привлечением РНК-связывающих белков (например, Yra1, Npl3, и Nab2), а также консервативный рецептор экспорта мРНК Mex67-Mtr2 (TAP-p15 у человека) [1,2]. В отсутствие компонентов TREX или TREX-2 возникающие мРНК неправильно собираются в мРНП, что может привести к гибридизации с ДНК-матрицей (образование R-петель), дефектному полиаденилированию и накоплению на 3′-конце гена [ 3–5].В результате в ядре накапливаются некомпетентные к экспорту мРНП [6,7]. Во время созревания и транспорта ядерные mRNPs контролируются системами наблюдения для предотвращения экспорта и последующей трансляции аберрантных мРНК (rev. [8,9]). У мутантов TREX несовершенные мРНП могут быть нацелены и разрушены ядерной экзосомой вместе с комплексом полиаденилирования TRAMP [10].
Важной задачей контроля качества ядерной мРНК является предотвращение экспорта интрон-содержащих пре-мРНК в цитоплазму.Экспорт несплайсированных пре-мРНК редко встречается в клетках дикого типа, тогда как значительная утечка пре-мРНК наблюдается в штаммах с дефектами ранних факторов сплайсинга или для транскриптов, мутировавших в 5′-сайте сплайсинга или в последовательности точки ветвления [11 –13]. Утечка пре-мРНК, лишенных каких-либо явных дефектов сплайсинга, наблюдалась в клетках, лишенных Mlp1 (гомолог человеческого TPR) или Nup60, двух структурных компонентов нуклеоплазматической корзины комплекса ядерных пор (NPC) [14]. Это указывает на то, что Nup60-Mlp1 избирательно улавливает содержащие интроны пре-мРНК и, возможно, др. Аберрантно процессируемые мРНК, чтобы предотвратить их выход из ядра [14,15].Совсем недавно было показано, что Pml39, партнер связывания Mlp1, и Esc1, белок периферии ядра, участвующий в правильной локализации комплекса Nup60-Mlp1, вносят вклад в функцию Mlp1-Nup60 [16,17]. Однако механизм распознавания и удаления аберрантных транскриптов остается неясным.
Белок дрожжевого PIN-домена Swt1 / Yor166c (синтетический летальный с TREX 1) был идентифицирован при скрининге на синтетическую летальность с субъединицей TREX Hpr1, функционально взаимодействует с комплексом TREX и необходим для оптимальной скорости транскрипции [18].Модуль PIN (PilT N-конец) имеет длину приблизительно 130 остатков и имеет складку, аналогичную 5′-экзонуклеазам и эндонуклеазам FLAP [19,20]. Типичными чертами являются четыре инвариантных кислотных остатка, расположенные в отрицательно заряженной поверхностной щели, образующие предсказанный каталитический центр [21]. Эти остатки, вероятно, хелатируют двухвалентный ион и вместе с двумя соседними гидроксильными остатками участвуют в нуклеофильной атаке на фосфодиэфирную связь субстрата. PIN-домен человеческого SMG6 проявляет эндонуклеазную активность in vitro и необходим для его функции при нонсенс-опосредованном распаде мРНК (NMD) [22].Геном дрожжей кодирует восемь белков, содержащих домен PIN, включая Nob1 и Utp24 / Fcf1, которые помогают в эндонуклеолитическом расщеплении предшественников рРНК во время биогенеза рибосомы [23,24] и субъединицы Rrp44 / Dis3 экзосомы (C. Schneider и D. Толлервей, неопубликованные данные). Однако было продемонстрировано, что ни один из этих дрожжевых белков не функционирует как эндонуклеаза.
Здесь мы сообщаем, что эволюционно консервативный белок Swt1 проявляет РНК-эндонуклеазную активность, которая зависит от домена PIN.Swt1 необходим в клетках, нарушенных для образования котранскрипционных мРНП или контроля качества перинуклеарных мРНП, и может связываться с NPC. Эти данные показывают важную роль эндонуклеолитического расщепления в процессе наблюдения ядерной мРНК.
Результаты
SWT1 Функционально перекрывается с факторами, участвующими в сборке котранскрипционных мРНП и контроле качества перинуклеарных мРНПЧтобы идентифицировать факторы, которые взаимодействуют с TREX-2, комплексом экспорта транскрипции, который функционирует на внутренней стороне NPC, мы провели синтетическую летальность (sl) экран с штаммом sac3 Δ.Среди кандидатов мы идентифицировали мутантный аллель гена SWT1 / YOR166 . Генетическое взаимодействие было подтверждено отсутствием роста гаплоидного штамма sac3 Δ swt1 Δ, полученного в результате потери плазмиды (фигура S1). Другие компоненты TREX-2 Thp1 и Sus1 были либо sl ( thp1 Δ), либо демонстрировали сильное синергетическое взаимодействие ( sus1 Δ) с swt1 Δ, демонстрируя тесное функциональное взаимодействие между Swt1 и комплексом TREX-2 (и Рисунок S1 ).
Таблица 1
Генетические взаимодействия SWT1
Чтобы расширить эти исследования, мы проверили, перекрывается ли SWT1 функционально с другими факторами, участвующими в биогенезе и обмене мРНП. Сильные генетические взаимодействия наблюдались между swt1 Δ и компонентами THO / TREX ( tho2 Δ, hpr1 Δ, mft1 Δ, thp2 Δ и sub2–85 ) в соответствии с предыдущими выводами [18] . Напротив, не было обнаружено никаких генетических взаимодействий между swt1 Δ и рецептором экспорта мРНК Mex67 или адаптером Yra1 [18].Кроме того, мы обнаружили sl-взаимодействие между swt1Δ и NPL3 , которое кодирует мРНК-связывающий белок с множеством ролей в биогенезе мРНП [25, 26] (и рисунок S1).
Эти исследования также выявили генетические взаимодействия между SWT1 и MLP1 , NUP60 и ESC1 (и рисунок S1). Mlp1 и Nup60 являются факторами, ассоциированными с NPC, которые, как полагают, составляют ядерную корзину NPC и участвуют в контроле качества mRNP перед ядерным экспортом [14].Esc1 является белком внутренней стороны ядерной мембраны, недавно было показано, что он влияет на ассоциацию комплекса Mlp1-Nup60 с NPC [17].
Несколько факторов, участвующих в различных этапах производства, экспорта и наблюдения мРНП, не взаимодействуют генетически с swt1Δ , включая субъединицы ядерной экзосомы (Rrp6 и Rrp44 / Dis3), компоненты комплекса TRAMP (Trf4 и Mtr4), т.е. 5′-экзонуклеаза Rat1 и факторы, участвующие в обмене цитоплазматической мРНК (см. Проверенные факторы).Эти генетические анализы показывают, что Swt1 специфически взаимодействует с факторами, необходимыми для котранскрипционной сборки и перинуклеарного контроля качества ядерных мРНП.
Swt1 обладает эндорибонуклеазной активностью in vitro
PIN-домен Swt1, расположенный в центральной области (остатки 130–262), эволюционно консервативен и содержит все четыре консервативных кислотных остатка, участвующих в катализе (рис. S2). Поэтому мы проверили, обладает ли очищенный из дрожжей Swt1 нуклеазной активностью in vitro.Мы создали меченый FLAG Swt1 дикого типа и мутант Swt1, которые, как ожидается, будут каталитически неактивными (Swt1-D135N; Asp 135, как предполагается, является остатком каталитического центра). Они были экспрессированы в дрожжах и очищены с помощью аффинной хроматографии (А). Способность этих белков расщеплять одноцепочечную РНК in vitro оценивали с использованием 5′- или 3′-радиоактивно меченных олиго (A) 30 субстратов. Swt1 дикого типа эффективно расщепляет как 5′-, так и 3′-меченные олиго (A) 30 РНК. Напротив, мутантный белок Swt1-D135N не проявлял активности в отношении 3′-меченого субстрата РНК и только очень слабую активность в отношении 5′-меченного зонда (B).Остаточная активность на 5′-меченном субстрате может быть связана с загрязняющей 3′-5′-экзонуклеазой, что часто наблюдается в анализах экзонуклеаз (C. Schneider, неопубликованные данные). Отсутствие деградации 3′-меченного субстрата за счет этой активности, вероятно, отражает защиту за счет присутствия 3′-фосфата на [ 32 P] pCp-меченой РНК. Эксперименты с динамикой продолжительности также показали, что Swt1 дикого типа разлагает как 5′-, так и 3′-меченные субстраты с очень сходной кинетикой (C), что согласуется с активностью эндорибонуклеазы.
Swt1 обладает эндонуклеолитической активностью в отношении одноцепочечной РНК in vitro(A) Очищенные белки Swt1 дикого типа и мутантные белки Swt1-D135N, используемые для измерения нуклеазной активности in vitro. Указанные меченые FLAG белки Swt1 были очищены аффинно из дрожжей и проанализированы с помощью SDS-PAGE и окрашивания Кумасси. M, маркер молекулярной массы.
(B) Белок Swt1 дикого типа или мутантный белок Swt1-D135N (по 0,5 мкг каждого) инкубировали с 5 ‘- [ 32 P] -концевыми метками (левая панель) или 3′ — [ 32 P] -концевая (правая панель) олиго (A) 30 РНК в течение 0, 60 и 120 мин при 30 ° C.Продукты реакции разделяли на денатурирующем геле 12% полиакриламид / 8 М мочевина и визуализировали с помощью авторадиографии. Зонды РНК, инкубированные без белка, служили отрицательными контролями, обозначенными отрицательным знаком (-). Звездочка (*) обозначает низкую активность контаминирующей экзонуклеазы дрожжей.
(C) Динамика активности РНКазы Swt1 на 5′- [ 32 P] -конце (левая панель) или 3′- [ 32 P] -конце (правая панель) олиго (A ) 30 РНК. Анализ выполняли, как описано в (B), и аликвоты отбирали в указанные моменты времени.
(D) Swt1 дикого типа или мутантный Swt1-D135N инкубировали с 52-нуклеотидной РНК, меченной 5 ‘- [ 32 P]-концом, со стабильной концевой ножкой в течение 0, 30 и 60 минут при 30 °. C, и реакцию анализировали, как указано в (B). ОН, частичный щелочной гидролиз РНК-зонда, используемого в качестве лестницы молекулярной массы. Положения стержневых остатков обозначены синим цветом на правой стороне геля и на вторичной структуре РНК, рассчитанной с помощью MFOLD.
Чтобы оценить, функционирует ли Swt1 как эндонуклеаза, использовали структурированный субстрат РНК из 52 нуклеотидов (нт), который содержит стабильную основу (от мыши 5.8S рРНК), которая закрывает 5′- и 3′-концы РНК (D). Оставшаяся часть РНК (нуклеотиды 11–43), как предполагается, будет по крайней мере частично одноцепочечной. Swt1 дикого типа, но не мутант Swt1-D135N, эффективно эндонуклеолитически расщеплял структурированную РНК из 52 нуклеотидов с явным предпочтением одноцепочечных областей субстрата РНК (D). Взятые вместе, данные показывают, что Swt1 проявляет in vitro эндонуклеазную активность без четкой специфичности последовательности, но с предпочтением одноцепочечной РНК.
Делеция
SWT1 индуцирует накопление ядерного поли (A) + РНК и усиливает утечку цитоплазмы пре-мРНК, дефектных по сплайсингу, в mlp1 Δ и nup60 Δ мутантов дальнейшее исследование роли мутантов Swt1 в контексте экспорта связанной с транскрипцией мРНК и надзора за ядерной мРНП, мы выполнили анализы экспорта мРНК и утечки пре-мРНК с жизнеспособными двойными мутантами (рисунок S1), несущими swt1 Δ вместе с thp2 Δ, mft1 Δ (THO / TREX), sus1 Δ (TREX-2), nup60 Δ, mlp1 Δ и esc1 Δ (контроль качества перинуклеарных мРНП).Делеция SWT1 значительно усиливала накопление ядерного поли (A) + РНК по сравнению с thp2 Δ, mft1 Δ, sus1 Δ и nup60 Δ одиночными мутантами (A, S3A и неопубликованные данные) . Более того, swt1 Δ индуцировал накопление в ядре РНК poly (A) + в мутантах mlp1 Δ и esc1 Δ, что не было обнаружено у одиночных мутантов (A; см. Также [27,28]) . SWT1 участвует в контроле качества ядерных мРНП с помощью Mlp1-Nup60(A) Нарушение SWT1 индуцирует или усиливает ядерное накопление поли (A) + и SSA1 РНК в mlΔp1 esc1 Δ или nup60 Δ, соответственно.Указанные штаммы с одним и двумя мутантами выращивали при 30 ° C ( nup60 Δ) или при изменении температуры до 37 ° C в течение 90 мин ( mlp1 Δ и esc1 Δ), а локализация поли (A) + РНК и SSA1 РНК анализировали гибридизацией in situ с использованием Cy3-oligo (dT) и SSA1 -специфических зондов, соответственно. ДНК окрашивали DAPI. Процент клеток, показывающих ядерное накопление поли (A) + РНК или SSA1 мРНК, соответственно, был рассчитан для n > 150 клеток и указан под каждой панелью ( mlp1 Δ и esc1 Δ штаммов).В случае штамма nup60 Δ показан процент клеток со значительно усиленным ядерным сигналом поли (А) + (более чем в 1,5 раза выше средней интенсивности ядерного сигнала nup60 Δ).
(B) Делеция SWT1 вызывает утечку репортерных пре-мРНК в цитоплазму и синергетически усиливает утечку пре-мРНК клеток mlp1 Δ и nup60 Δ клеток. Утечка репортеров пре-мРНК lacZ , которые вызывают активность β-галактозидазы, анализировали, как описано в разделе «Материалы и методы».Отношения между активностями β-галактозидазы, полученными из репортерной пре-мРНК и безинтронной контрольной мРНК, были рассчитаны и показаны относительно отношения соответствующего штамма дикого типа. Показаны результаты четырех независимых экспериментов. Планки погрешностей указывают на стандартные отклонения.
Чтобы оценить, отражает ли накопление поли (A) + РНК накопление ядерной мРНК, мы протестировали специфический транскрипт, мРНК SSA1 (A) теплового шока. Эта мРНК явно накапливалась в ядре двойного мутанта mpl1Δ swt1 Δ, но не в соответствующих одиночных мутантах.Напротив, никакого ядерного накопления не наблюдалось при анализе ядерного экспорта тРНК (неопубликованные данные). Чтобы количественно определить, увеличились ли уровни транскрипта SSA1 в двойном мутанте mlp1 Δ swt1 Δ, мы измерили количество мРНК SSA1 до и после теплового шока с помощью полимеразной цепной реакции с обратной транскрипцией в реальном времени ( ОТ-ПЦР). Уровни SSA1 были слегка увеличены у одиночных мутантов, но синергетическое увеличение наблюдалось у двойного мутанта mlp1 Δ swt1 Δ по сравнению со штаммом дикого типа (фиг. S4).
Чтобы проверить, приводят ли двойные мутанты swt1Δ и mlp1Δ или nup60Δ к синергетическим дефектам в надзоре за пре-мРНП, мы контролировали утечку цитоплазмы несплицированных репортеров пре-мРНК [11,12,14]. Утечка репортера с интактными сигналами сплайсинга (pJCR1) не наблюдалась в штамме swt1Δ , тогда как штамм nup60Δ показал повышенную утечку несплицированной пре-мРНК, которая синергетически усиливалась в штамме swt1 Δ nup60 Δ двойной мутант (B).Затем мы проверили, проявляют ли репортеры пре-мРНК с мутацией в 5′-сайте сплайсинга (mut5’SS) или в сайте точки ветвления (mutBP) повышенную утечку цитоплазмы в клетках swt1 Δ. Примечательно, что утечка репортера mutBP была увеличена примерно в 4 раза в одинарных мутантах swt1 Δ или mlp1Δ относительно дикого типа (B; см. Также [14,16]). Утечка цитоплазмы обоих мутантных репортеров пре-мРНК снова была сильно синергетически усилена в двойном мутанте swt1Δ mlp1Δ (B).
Мы пришли к выводу, что потеря Swt1 ведет к увеличению утечки цитоплазмы дефектных по сплайсингу пре-мРНК, что может синергетически усиливаться мутациями в комплексе Mlp1-Nup60. Это указывает на то, что Swt1 и Mlp1-Nup60 играют разные, но частично совпадающие роли в надзоре ядерных мРНП. На первый взгляд кажется противоречивым, что повышенная утечка пре-мРНК сопровождается накоплением в ядре поли (A) + РНК. Однако эти два эффекта не обязательно исключают друг друга.Когда контроль качества ядер серьезно нарушен (например, в клетках mlp1 Δ swt1 Δ или nup60 Δ swt1 Δ), большее количество аберрантных мРНП может накапливаться в ядре и, следовательно, иметь более высокий шанс утечки. .
Эндонуклеазный домен необходим для функции Swt1 на физиологических уровнях экспрессии
В предыдущем исследовании [18] было показано, что Swt1, лишенный PIN (эндонуклеазного) домена, дополняет летальность swt1Δ в мутантных штаммах THO.Однако Swt1 является белком с низким содержанием (см. Ниже), и конструкции были сверхэкспрессированы с относительно сильного промотора NOP1 . Чтобы оценить, является ли эндонуклеазная активность Swt1 существенной для комплементации фенотипов swt1Δ при физиологических уровнях экспрессии, мы проанализировали каталитически неактивный мутант Swt1-D135N, экспрессируемый с эндогенного промотора (A). Swt1-D135N и Swt1 дикого типа экспрессировались на сходных уровнях (B), но Swt1-D135N не смог дополнить дефекты роста swt1Δ в любой комбинации двойных мутантов (A и неопубликованные данные).Более того, Swt1-D135N не спасает накопление ядерной мРНК, вызванное swt1Δ на фоне мутаций mlp1 Δ, nup60 Δ, sus1 Δ или thp2 Δ (C и неопубликованные данные). Наконец, утечка из ядра несплайсированного репортера пре-мРНК (mutBP) в экспрессирующих Swt1-D135N клетках (D) была неотличима от утечки, наблюдаемой в штамме swt1 Δ (B). Подобные фенотипы потери функции наблюдались для усеченного мутанта swt1 , лишенного всего PIN-домена (Swt1ΔPIN) (рисунок S5 и неопубликованные данные).
Каталитически неактивный мутант swt1-D135N Напоминает swt1 Нулевая мутация In Vivo(A) Мутант swt1-D135N не дополняет синтетически летальные / усиленные фенотипы двойного Δt1 swt1 . мутантные штаммы. Штаммы трансформировали диким типом SWT1 или мутантом swt1-D135N , и трансформанты наносили пятнами в 10-кратных серийных разведениях на указанные планшеты. Штаммы инкубировали в течение 5 дней при 30 ° C ( tho2 Δ swt1 Δ) или в течение 3 дней при 36 ° C ( thp2 Δ swt1 Δ и nup60 Δ swt1 Δ).
(B) Уровни экспрессии белков Swt1 дикого типа и мутантных белков сходны. Лизаты цельных клеток, полученные из штамма swt1 Δ или штамма swt1 Δ, экспрессирующего Swt1 или Swt1-D135N дикого типа, анализировали с помощью SDS-PAGE и вестерн-блоттинга с использованием поликлональных антител против Swt1. Обнаружение белка Arc1 поликлональным антителом против Arc1 использовали в качестве контроля загрузки.
(C) Мутант Swt1-D135N не может устранить дефект экспорта поли (A) + штамма mlp1 Δ swt1 Δ. mlp1 Δ swt1 Δ клетки, экспрессирующие Swt1 или Swt1-D135N дикого типа, анализировали на локализацию поли (A) + РНК, как в A, или окрашивали DAPI для обнаружения ДНК.
(D) Swt1-D135N-экспрессирующие клетки демонстрируют фенотип утечки ядерной пре-мРНК. Штаммы swt1 Δ и mlp1 Δ swt1 Δ трансформировали указанными плазмидами и анализировали на утечку репортера пре-мРНК lacZ (mutBP), как описано в B. показано.Планки погрешностей указывают на стандартные отклонения.
(E) Swt1-D135N клетки накапливают повышенные уровни мутированной пре-мРНК lacZ . Тотальную РНК выделяли из swt1-D135N и изогенного штамма дикого типа, экспрессирующего содержащий интрон репортерный транскрипт lacZ (норма , , для нормального) или содержащий интрон транскрипт lacZ , мутировавший в ветви — сайт точечного сплайсинга (mut . , для мутировавших). Уровни lacZ количественно определяли с помощью ОТ-ПЦР в реальном времени с использованием мРНК GAL1 в качестве контрольного транскрипта.Показаны результаты четырех независимых экспериментов. Планки погрешностей указывают стандартные ошибки среднего.
(F) Сверхэкспрессия Swt1 дикого типа, но не сверхэкспрессия каталитически неактивного мутанта Swt1-D135N, токсична для клеток. Swt1 Δ клетки, содержащие GAL1 :: SWT1 , GAL1 :: swt1-D135N или пустую плазмиду, наносили пятнами в 10-кратных серийных разведениях на планшеты, содержащие либо галактозу, либо глюкозу, и инкубировали в течение 3 дней при 30 ° C. Показаны только два последних разведения чашки с глюкозой.
(G) Сверхэкспрессия Swt1 дикого типа, но не сверхэкспрессия каталитически неактивного мутанта Swt1-D135N, вызывает сильное ядерное накопление поли (A) + РНК. Те же штаммы, что и в (F), выращивали в среде, содержащей рафинозу, до фазы ранней логарифмической активности, а затем индуцировали 2% галактозой в течение 5 часов. Локализацию РНК поли (А) + анализировали путем гибридизации in situ с использованием зонда Cy3-oligo (dT). ДНК окрашивали DAPI. Показан процент клеток, показывающих накопление ядерного сигнала поли (А) + ( n > 150 клеток).
Однако Swt1-D135N, Swt1ΔPIN и другие конструкции с частичной делецией могут дополнять swt1Δ для роста на фоне tho2Δ , когда сверхэкспрессируется под контролем сильного промотора ADh2 (фиг. S5). Напротив, другие фенотипы swt1Δ , включая утечку ядерной пре-мРНК и накопление ядерной РНК poly (A) + в штаммах mlp1Δ swt1Δ , не могли быть подавлены сверхэкспрессией Swt1-D135N или Swt1ΔPIN мутантов из ADh2. Промотор (D и неопубликованные данные).Таким образом, при физиологических уровнях экспрессии эндонуклеазная активность Swt1 важна для его функции, тогда как потребность в каталитической активности может быть частично устранена сверхэкспрессией неактивного белка Swt1.
Swt1-D135N Клетки накапливают повышенные уровни аберрантной пре-мРНКЧтобы оценить возможную роль эндонуклеазной активности Swt1 в деградации дефектных мРНП, мы определили, стабилизируется ли «аберрантный» транскрипт в swt1-D135N клетки относительно изогенного дикого типа.В качестве аномального транскрипта мы использовали интрон-содержащую конструкцию lacZ (экспрессируемую из плазмиды mutBP; см. Выше), которую использовали в анализах утечки пре-мРНК. Это таит в себе мутацию в сайте точки ветвления сплайсинга, которая эффективно блокирует сплайсинг пре-мРНК [12]. Содержащий интрон дикого типа транскрипт lacZ , кодируемый плазмидой pJCR51, использовали в качестве контрольной пре-мРНК. Накопление аберрантного транскрипта было существенно повышено в клетках swt1-D135N по сравнению со штаммом дикого типа (mut.столбцы на E), тогда как уровни нормально процессируемой мРНК lacZ не изменились между штаммами swt1-D135N и диким типом (нормальные столбцы на E). Эти результаты предполагают, что эндорибонуклеазная активность Swt1 участвует в селективном устранении дефектных транскриптов мРНК.
Сверхэкспрессия Swt1 токсична и вызывает сильное ядерное накопление пре-мРНК
Поскольку Swt1 участвует в надзоре ядерных мРНП, его экспрессия и / или нацеливание могут регулироваться.Количественный вестерн-блот-анализ с использованием антител против Swt1 показал, что in vivo уровни эндонуклеазы Swt1 очень низкие (~ 200–300 молекул на клетку) (неопубликованные данные; см. Также [18]), что может быть важно для предотвращения несоответствующей деградации РНК. В крупномасштабном анализе сообщалось, что Yor166 / Swt1 токсичен после сверхэкспрессии системой Tet-off [29]. В соответствии с этим открытием, доминантно-летальный фенотип был индуцирован сверхэкспрессией Swt1 дикого типа, но не каталитически неактивного мутанта Swt1-D135N с промотора GAL1 (F).Большинство клеток со сверхэкспрессией Swt1 показали сильное ядерное накопление как поли (A) + РНК (E), так и мРНК SSA1 , тогда как тРНК обычно экспортировалась (рисунок S3B и неопубликованные данные). Примечательно, что при анализе локализации репортерных конструкций lacZ , содержащих интрон / без интрона, накопление в ядре репортера пре-мРНК, содержащего интрон, было сильнее, чем для мРНК без интрона (рис. S3C), что согласуется с дефектом в пре-мРНК. надзор за мРНК.Эти анализы демонстрируют, что сверхэкспрессия каталитически активного Swt1 токсична и мешает экспорту мРНК и / или надзору пре-мРНК.
Swt1 ассоциирует с комплексами ядерных пор в
nup133 Δ клеткахСубклеточное фракционирование ранее показало, что меченный TAP Swt1 локализован в ядре [18]. Точное расположение Swt1 не может быть определено, однако, потому что количество зеленого флуоресцентного белка (GFP) или иммуно-меченого Swt1, экспрессируемого с эндогенного промотора, было слишком низким для визуализации (неопубликованные данные и [18]).Присоединение нескольких тегов GFP может позволить обнаруживать белки с низким содержанием (N. Daigle, A. Bancaud и J. Ellenberg, неопубликованные данные). Таким образом, мы создали функциональную конструкцию 4xGFP-Swt1, экспрессируемую с эндогенного промотора SWT1 , которая дополняла штаммы с двойной делецией swt1 Δ mlp1 Δ, swt1 Δ nup60 Δ или Δ17 swt1 Δ (Рисунок S6 и неопубликованные данные). При анализе в штамме swt1 Δ, 4xGFP-Swt1 был локализован как в ядре, так и в цитоплазме с небольшим ядерным обогащением в некоторых клетках (A).При экспрессии в мутанте nup133 Δ с кластеризацией ядерных пор [30], 4xGFP-Swt1 проявлял значительную совместную кластеризацию с NPC на периферии ядра, которая терялась при комплементации штамма nup133 Δ плазмидным производным NUP133 ( А). Когда конструкция 4xGFP-Swt1, лишенная каталитического PIN-домена (4xGFP-Swt1ΔPIN), экспрессировалась в клетках swt1 Δ, она концентрировалась в нескольких точках. Они всегда были близки к NPC, как было выявлено при совместной визуализации Nup120-mCherry (B), и 4xGFP-Swt1ΔPIN, также связанных с NPC в клетках nup133 Δ.Нацеливание Swt1ΔPIN на NPC не зависело от Mlp1, Nup60 или Esc1. Вместо этого 4xGFP-Swt1ΔPIN показал более отчетливое и пунктированное окрашивание ядерной оболочки в штаммах mlp1 Δ, nup60 Δ и esc1 Δ, чем в соответствующих изогенных штаммах дикого типа (B и неопубликованные данные). Нуклеоцитоплазматическая локализация полноразмерного 4xGFP-Swt1 в клетках mlp1 Δ, nup60 Δ и esc1 Δ не изменилась (неопубликованные данные). Вместе с генетическими данными эти наблюдения показывают, что Swt1 взаимодействует с NPC.
Swt1 ассоциирует с комплексами ядерных пор в нуклеопориновом мутанте nup133 Δ(A) 4xGFP-Swt1 (зеленый) или (B) 4xGFP-Swt1ΔPIN (зеленый) коэкспрессируются с Nup120-mCherry (красный маркер) ) в указанных штаммах ( swt1 Δ, nup133 Δ swt1 Δ, nup133 Δ swt1 Δ + NUP133 [экспрессируется из центромерной плазмиды], Δt1 mlp1 и mlp1 nup60 Δ swt1 Δ) и анализировали с помощью флуоресцентной микроскопии.Клетки также просматривались оптикой Номарского.
Обсуждение
В этом исследовании мы показываем, что Swt1 является эндонуклеазой РНК, которая может связываться с NPC и участвовать в системах, которые контролируют качество ядерных мРНП перед экспортом в цитоплазму. Эндонуклеазная активность PIN-домена Swt1 имеет решающее значение для его функции in vivo на физиологических (то есть низких) уровнях экспрессии. Сильно сверхэкспрессированные конструкции Swt1, лишенные PIN-домена, могли выполнять некоторые из функций Swt1 in vivo, тогда как сверхэкспрессия активного Swt1 была летальной.Это указывает на то, что Swt1 функционирует и как нуклеаза, и как белковый компонент комплексов наблюдения мРНП.
Чтобы избежать цитоплазматической трансляции дефектных мРНП в потенциально вредные белки, системы биогенеза мРНП и системы наблюдения взаимодействуют в ядре. Эндорибонуклеазная активность Swt1 важна, когда функция комплексов, котранскрипционно формирующих и проверяющих ядерные мРНП, нарушена, это указывает на то, что Swt1 может участвовать в деградации компонента мРНК аберрантных ядерных мРНП.Эндонуклеазная активность может быть важной, потому что транскрипционно захваченные мРНП, экспортно-некомпетентные мРНП или пре-мРНП, неспособные к сплайсингу, лишены всех доступных концов РНК, которые могли бы способствовать деградации ядерной 5′-экзонуклеазой Rat1 или 3′-экзонуклеазной активностью экзосомы. . Эндонуклеазное расщепление дефектной мРНК и мРНП с помощью Swt1 может генерировать свободные концы мРНК для дальнейшей экзонуклеолитической деградации. В соответствии с этой моделью, аберрантная репортерная конструкция пре-мРНК, которая не может быть сплайсирована, избирательно накапливалась в каталитически неактивном мутанте Swt1-D135N, и потеря Swt1 из штамма mlp1Δ стабилизировала ограниченную ядерной реакцией мРНК SSA1 .
Высокие уровни Swt1 были токсичными и индуцировали сильное ядерное накопление поли (A) + РНК. Среди нескольких возможностей, которые могут объяснить этот фенотип, сверхэкспрессия Swt1 может вызывать ошибочное расщепление РНК в правильно сформированных комплексах RNP. По нашим оценкам, количество молекул Swt1 в клетке составляет всего около 200–300 копий, и этот низкий уровень может быть важным для подавления расщепления нецелевой РНК. Накопление в ядре интрон-содержащей репортерной пре-мРНК, индуцированной сверхэкспрессией Swt1, было сильнее, чем для соответствующей безинтронной пре-мРНК, что согласуется с дефектом в наблюдении за пре-мРНК.Примечательно, что сверхэкспрессия Mlp1, но не делеция его гена, также вызывает накопление в ядре интрон-содержащих пре-мРНК [14,28], что согласуется с моделью, согласно которой Swt1 и Mlp1 функционируют вместе в этом пути.
Наши данные согласуются с ранее предполагаемой ролью Swt1 в транскрипции мРНК с помощью TREX [18]. Комплексы TREX / TREX-2 необходимы для котранскрипционной сборки мРНП, и нарушение их функции приводит к образованию аберрантных структур мРНП, которые препятствуют транскрипции RNAPII [3–5,9].Мы прогнозируем, что эти дефектные частицы мРНП представляют собой важные субстраты для деградации эндонуклеаз с помощью Swt1. В соответствии с этой моделью, дефектные и некомпетентные к экспорту мРНП, сформированные в мутантах TREX, накапливаются вместе с NPCs [9,31], где также может быть обнаружен Swt1.
В клетках дикого типа Swt1 распределялся между ядром и цитоплазмой и, следовательно, потенциально мог обнаруживать образование дефектных пре-мРНП на всем пути биогенеза мРНП. Однако Swt1 накапливался в NPC, когда они были кластеризованы в мутанте nup133 Δ [30].Эта локализация Swt1 в NPC не может быть коррелирована с нарушением экспорта мРНК ( nup133, Δ клетки также обнаруживают дефект экспорта мРНК), поскольку 4xGFP-Swt1 не накапливался в ядерной оболочке у мутантного экспортного мРНК mex67-5 (неопубликовано). данные). Более того, Swt1, лишенный своего эндонуклеазного домена, стал концентрироваться в перинуклеарных пятнах даже в клетках с нормальным набором нуклеопоринов. Мы предполагаем, что Swt1 временно рекрутируется в NPC через взаимодействия с нуклеопоринами и / или с экспортно-некомпетентными пре-мРНП, заблокированными на NPC, и функционирует там, чтобы предотвратить несоответствующий экспорт пре-мРНК.
Анализ ядерной деградации дефектных РНК у дрожжей в основном сосредоточен на активности 5′- и 3′-экзонуклеаз. Эта работа демонстрирует, что расщепление эндонуклеазой также может играть важную роль в надзоре за ядерной РНК.
Материалы и методы
Штаммы дрожжей, плазмиды и генетические манипуляции.
Штаммы дрожжей, использованные в этом исследовании, перечислены в Таблице S1; плазмидные конструкции описаны в таблице S2. Сайт-направленный мутагенез гена SWT1 для создания мутанта swt1-D135N был выполнен с помощью гибридной ПЦР [32].Скрининг синтетической летальности со штаммом sac3 Δ проводили, как описано ранее [33].
Анализ нуклеазной активности in vitro.
Для измерения активности нуклеазы Swt1 in vitro, белок FLAG-Swt1 дикого типа и мутантный белок D135N были экспрессированы из плазмид pADh211-FLAG- SWT1 или pADh211-FLAG- swt1 — D135N ( ) и очищают аффинной хроматографией против FLAG. Вкратце, собирали 4 мкл дрожжевой культуры с оптической плотностью при 600 нм (OD 600 ) приблизительно 4.0, ресуспендировали в буфере FLAG (50 мМ Трис-HCl [pH 7,5], 200 мМ NaCl, 0,5 мМ DTT, 0,05% NP-40) до конечного объема 50 мл, лизировали взбиванием шариков (Pulverissete 6; Fritsch) и инкубировали с 120 мкл агарозы anti-FLAG M2 (Sigma) в течение 120 мин при 4 ° C. Суспензию анти-FLAG промывали 750 объемами буфера FLAG, и связанный материал элюировали пептидом FLAG в соответствии с инструкциями производителя (Sigma) с последующим концентрированием и заменой буфера.
Анализы нуклеазной активности очищенных белков на 5′-конце или 3′-конце радиоактивно меченых РНК проводили при 30 ° C в 15 мМ Трис-HCl (pH 7.6), 75 мМ NaCl, 2 мМ DTT, 100 мкг / мл БСА, 0,8 Ед / мкл RNasin, 4,5% глицерина, 0,05% NP-40, 0,3 мкМ тРНК Escherichia coli и 7,5 мМ MnCl 2 . Перед добавлением 20–30 фмоль меченого субстрата РНК 20 или 30 мкл реакционной смеси, содержащей 10 пмоль белка, предварительно инкубировали в течение 15 мин при 30 ° C. Затем отбирали аликвоты по 4 мкл в моменты времени, указанные в, и реакцию останавливали добавлением буфера для загрузки РНК, содержащего формамид. Продукты реакции разделяли на денатурирующем геле 12% полиакриламид / 8 М мочевина и визуализировали с помощью авторадиографии.5 ‘- [ 32 P] — и 3′ — [ 32 P] -меченные субстраты РНК (Dharmacon) метили и очищали в геле, как описано [34].
Анализы утечки пре-мРНК.
По меньшей мере четыре независимых эксперимента были выполнены для каждого штамма, содержащего указанный репортер утечки пре-мРНК lacZ (pJCR1, pmut5’SS и pmutBP) или безинтронный контроль lacZ (pLGSD5). Трансформанты объединяли и выращивали в течение ночи в селективной среде раффинозы. Клетки разбавляли, выращивали до середины логарифмической фазы и экспрессию репортера индуцировали 2% галактозой в течение 150 мин.Анализы β-галактозидазы проводили в трех экземплярах для каждого образца в соответствии с опубликованным протоколом [11] с модификациями, описанными в [16].
Анализ уровней РНК
SSA1 и lacZ .Чтобы вызвать экспрессию SSA1 , к экспоненциально выросшим культурам применяли тепловой шок (42 ° C в течение 15 мин) путем добавления равного объема предварительно подогретой среды. Экспрессию репортеров пре-мРНК lacZ (pmutBP или pJCR51) индуцировали добавлением галактозы (конечная концентрация 2%) к культурам в ранней логарифмической фазе и их последующей инкубацией в минимальной среде галактоза-раффиноза в течение 6 часов.Клетки лизировали стеклянными шариками и встряхиванием, и общую РНК выделяли с помощью набора RNeasy Mini Kit (QIAGEN). РНК подвергали обратной транскрипции с использованием набора для обратной транскрипции QuatiTect (QIAGEN), а уровни специфического транскрипта РНК измеряли с помощью количественной ПЦР в реальном времени с использованием системы ABI Prism 7000 (Applied Biosystems) и ABsolute QPCR ROX Mix (ABgene). Каждый образец кДНК, а также контроли без обратной транскриптазы, амплифицировали в трех экземплярах с наборами праймеров и зондов, описанными в таблице S3.Уровни SSA1 и lacZ РНК были нормализованы до уровней мРНК RPB4 и GAL1 соответственно, которые, как было обнаружено, не изменились в испытанных условиях.
Разное.
Флуоресцентная гибридизация in situ (FISH) с использованием олиго (dT), SSA1, транскрипта-, тРНК- и lacZ -меченых зондов, меченных Cy3, выполнялась, как описано [33,35,14]. Локализацию слияний 4xGFP-Swt1, 4xGFP-Swt1ΔPIN и Nup120-mCherry анализировали в Zeiss Axio Imager.Микроскоп Z1 с программным обеспечением AxioVision 4.6.3. Экспрессию Swt1 анализировали иммуноблоттингом с использованием кроличьей поликлональной сыворотки против Swt1, созданной против рекомбинантного Swt1 (PickCell Laboratories), и конъюгата против кроличьей HRP в качестве вторичного антитела (BIO-RAD). Экспрессию вариантов белка Swt1, меченных FLAG, анализировали с помощью конъюгата анти-FLAG-HRP (Sigma).
Вспомогательная информация
Рисунок S1
Генетические взаимодействия между SWT1 и факторами сборки котранскрипционных мРНП и контроль качества перинуклеарных мРНП:Указанные штаммы дикого типа, одиночные и двойные мутанты были обнаружены в 10- кратные серийные разведения на планшетах, содержащих 5′-фтороротовую кислоту (FOA) в случае синтетического летального взаимодействия (левая панель) и на пластинах с синтетической полной декстрозой (SDC) -Leu после перетасовки 5′-FOA в случае синергетического дефект усиленного роста (средняя и правая панели).Планшеты инкубировали в течение 5 дней при 30 ° C (левая панель) или 3 дня при 30 ° C или 36 ° C (средняя и правая панели).
(2,23 МБ PDF)
Рисунок S2
Сравнение последовательностей PIN-доменов белков Swt1 и SMG6 / EST1A:Последовательности PIN-домена белков Swt1 из Saccharomyces cerevisiae (Swt1; (SpSwt1; Spcc24b10.15p), Drosophila melanogaster (DmSwt1; CG7206-PB) и Homo sapiens (HsSwt1; C1orf26) были выровнены с доменами PIN белков Drosophila, Drosophila и SMSAg6 человека (DmSmSAg6 и EmSmSG6). ) пользователя ClustalW.Звездочки указывают на консервативные кислотные остатки, участвующие в нуклеазной активности PIN-домена. Указаны положения соответствующих остатков Swt1.
(88 КБ PDF)
Рисунок S3
Накопление ядерной мРНК после делеции и сверхэкспрессии Swt1:(A) Удаление SWT1 усиливает ядерное накопление поли (A) + РНК TREX-2 и мутанты TREX. Указанные штаммы с одним и двумя мутантами были переведены на температуру 37 ° C на 90 мин, и локализация поли (A) + РНК была проанализирована гибридизацией in situ с использованием зонда Cy3-oligo (dT).ДНК окрашивали DAPI. Показан процент клеток, показывающих значительно усиленный ядерный сигнал поли (А) + (более 1,5-кратный средней интенсивности ядерного сигнала соответствующего одиночного мутантного штамма) ( n > 150 клеток).
(B) Сверхэкспрессия Swt1 запускает накопление в ядре мРНК SSA1 . Swt1 Δ клетки или swt1 Δ клетки, экспрессирующие GAL1 :: SWT1 , выращивали в селективной среде раффинозы до фазы раннего логарифма, индуцированной 2% галактозой в течение 5 часов, и была обнаружена локализация мРНК SSA1 гибридизацией in situ с использованием специфичного для SSA1 , Cy3-меченного зонда.ДНК окрашивали DAPI. Был рассчитан процент клеток, демонстрирующих накопление в ядре сигнала SSA1 ( n > 150 клеток), и он показан ниже.
(C) Сверхэкспрессия Swt1 вызывает накопление в ядре репортеров мРНК lacZ . Безинтронная (pLGSD5) или содержащая интрон (pJCR51) мРНК lacZ была экспрессирована в штамме swt1 Δ или GAL1 :: SWT1 дикого типа, и ее распределение анализировали, как в (B), с использованием штамма lacZ -специфический зонд.В каждом случае было исследовано не менее 300 клеток, и указан процент клеток с ядерным накоплением сигнала lacZ .
(2,30 МБ PDF)
Рисунок S4
mlp1Δ swt1Δ Двойное разрушение приводит к повышенным клеточным уровням SSA1 Транскрипт после теплового шока: Общая РНКбыла выделена из указанных культур в средней логарифмической фазе до (25 ° C) или после теплового шока до 42 ° C. на 15 мин. Уровни SSA1 количественно определяли с помощью анализа RT-PCR в реальном времени с использованием мРНК RPB4 в качестве контрольного транскрипта.Показаны репрезентативные результаты двух независимых экспериментов.
(65 КБ PDF)
Рисунок S5
Анализ делеции Swt1:(A) Схема конструкций Swt1, используемых для анализа делеции. Цифры указывают положение соответствующей аминокислоты вдоль белка Swt1.
(B) Анализ удаления Swt1. Последовательность перетасовки tho2Δ swt1Δ трансформировали указанными делеционными конструкциями SWT1 , экспрессировавшимися из эндогенного SWT1 или сильного промотора ADh2 , и способность к комплементации тестировали путем роста на пластинах 5′-FOA при 30 ° C для 5 дн.
(515 КБ PDF)
Рисунок S6
Характеристика слитых белков 4xGFP-Swt1 и 4xGFP-Swt1ΔPIN:(A) Слияние 4xGFP-Swt1 дополняет синтетически усиленные фенотипы mlp Δ1 и mlp Δ1 nup60 Δ swt1 Δ штаммы с двойными мутантами. Штаммы трансформировали плазмидой YCplac111-FLAG-4xGFP- SWT1 , YCplac111-FLAG- SWT1 или пустой плазмидой, и трансформанты наносили пятнами в 10-кратных серийных разведениях на планшеты SDC-Leu.Штаммы инкубировали 5 дней при указанных температурах.
(B) Сравнение уровней экспрессии белков Swt1, Swt1ΔPIN, 4xGFP-Swt1 и 4xGFP-Swt1ΔPIN. Лизаты цельных клеток получали из штаммов swt1 Δ или mlp Δ swt1 Δ, экспрессирующих указанные конструкции из центромерной плазмиды YCplac111, и анализировали с помощью SDS-PAGE и вестерн-блоттинга с использованием моноклональных антител против FLAG. Звездочка обозначает белок в клеточном лизате, перекрестно реагирующий с конъюгатом анти-FLAG.
(457 КБ PDF)
Таблица S1
Штаммы дрожжей, использованные в этом исследовании:(73 КБ DOC)
Таблица S2
Плазмиды, использованные в этом исследовании:(69 КБ DOC)
Таблица S3
Олигонуклеотидов ОТ-ПЦР в реальном времени, использованных в этом исследовании:(35 КБ DOC)
Escuela Politécnica Nacional | La Ciencia al Servicio del Hombre
Тодо
Todo
Acuerdo
Aniversario
capacitación
CARRERA Atlética
Casa Abierta
Celebración Ancestral
ceremonia
Ceremonia де Graduación Виртуальный
Чарла
Чарла Cientifica
Карлы де социализация
Чарла виртуальный
CHARLAS
Charlas Divulgativas
Ciclo де Conferencias
Clases Virtuales
Coloquio Internacional
Concierto
Concurso
Conferencia
Conferencia Виртуальные
CONFERENCIAS
CONGRESO
Конгресо Виртуальный
Conmemoración
Conversatorio
convocatoria
Cultural
curso
Curso Virtual
Cursos
Defensa
Diálogo
Diálogos de la Ciencia
Difusión
EJERCICIO
E NCUENTRO
Entrega de Certificados
Envío de Resúmenes
Evento artístico
Evento virtual
Eventos Graduados
Exposiciones
FERIA
FERIA
Feria virtual
9253 FiomaForo Conmemorativo
Homenaje
Inducción
Jornadas
Меса-де-Trabajo
Premiación
Presentación
Proyecto
Recolección де Алиментос
Реюньон
Руэда де Пренса
SEMINARIO
Семинарио Web
Sesion Solemne
simposio
Студенческий веб-семинар
Taller
Taller Virtual
Видеоконференция
Visita in situ
Webinario
Workshop
ʌ ʌ ʌ Arrastrar de arriba a abajo v v v
<<< Arrastrar de Lado a Lado >>>
<<< Arrastrar de Lado a Lado >>>
Тема Дестакадос
<<< Arrastrar de Lado a Lado >>>
Servicios a la Comunidad
<<< Arrastrar de Lado a Lado >>>
Редей
<<< Arrastrar de Lado a Lado >>>
.