
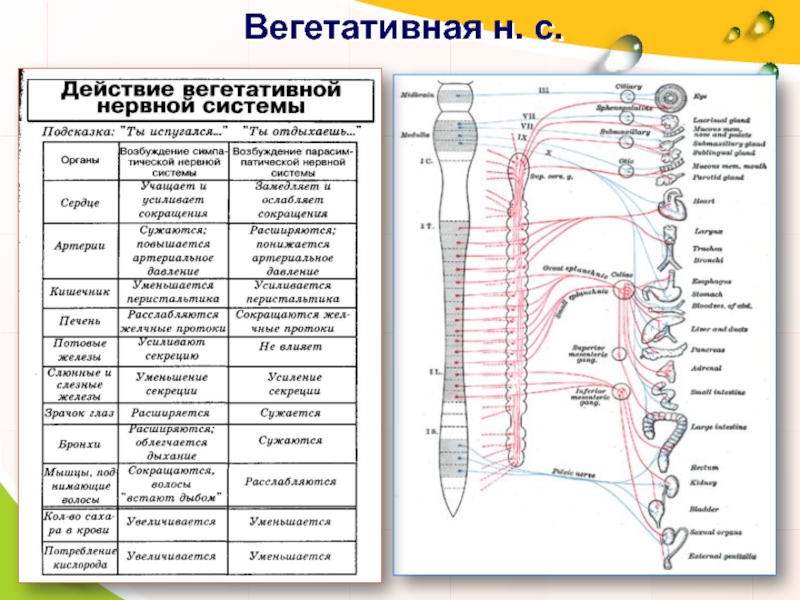
Функции вегетативной нервной системы
Вегетативная нервная система выполняет ряд функций:
Управляет деятельностью внутренних органов, кровеносных и лимфатических сосудов, осуществляя иннервацию гладкомышечных клеток и железистого эпителия.
Регулирует обмен веществ, приспосабливая его уровень к снижению или повышению функции органа. Тем самым осуществляет адаптационно-трофическую функцию, в основе которой лежит транспорт аксоплазмы — процесс непрерывного движения различных веществ от тела нейрона по отросткам в ткани. Одни из них включаются в обмен веществ, другие активируют метаболизм, улучшая трофику ткани.
Координирует работу всех внутренних органов, поддерживая постоянство внутренней среды организма.
Центры вегетативной нервной системы
Центры
вегетативной нервной системы разделяют
на сегментарные и надсегментарные
(высшие вегетативные центры).
Сегментарные центры располагаются в нескольких отделах центральной нервной системы, где выделяют 4 очага:
Мезенцефалический отдел в среднем мозге — добавочное ядро (Якубовича), nucleus accessorius, и непарное срединное ядро глазодвигательного нерва (III пара).
Бульбарный отдел в продолговатом мозге и мосту — верхнее слюноотделительное ядро, nucleus salivatorius superior, промежуточно-лицевого нерва (VII пара), нижнее слюноотделительное ядро, nucleus salivatorius inferior, языкоглоточного нерва (IX пара) и дорсальное ядроблуждающего нерва (X пара), nucleus dorsalis n. vagi.
Оба этих отдела объединяются под названием краниального и относятся к парасимпатическим центрам.
Тораколюмбальный отдел — промежуточно-боковые ядра, nuclei intermediolaterales, 16-ти сегментов спинного мозга (С8, Th1-12, L1-3).
 Они являются симпатическими центрами.
Они являются симпатическими центрами.
Сакральный отдел — промежуточно-боковые ядра, nuclei intermediolaterales, 3-х крестцовых сегментов спинного мозга (S2-4) и относятся к парасимпатическим центрам.
Высшие вегетативные центры (надсегментарные) объединяют и регулируют деятельность симпатического и парасимпатического отделов, к ним относятся:
Ретикулярная формация, ядра которой формируют центры жизненно-важных функций (дыхательный и сосудодвигательный центры, центры сердечной деятельности, регуляции обмена веществ и т.д.). Проекция дыхательного центра соответствует средней трети продолговатого мозга, сосудодвигательного центра — нижней части ромбовидной ямки. Нарушение функции ретикулярной формации проявляется вегетативно-сосудистыми расстройствами (кардио-васкулярные, вазомоторные). Кроме того страдают интегративные функции, которые необходимы для формирования целесообразного адаптивного поведения.

Мозжечок, принимая участие в регуляции двигательных актов, одновременно обеспечивает эти анимальные функции адаптационно-трофическими влияниями, которые через соответствующие центры приводят к расширению сосудов интенсивно работающих мышц, повышению уровня трофических процессов в последних. Установлено участие мозжечка в регуляции таких вегетативных функций, как зрачковый рефлекс, трофика кожи (скорость заживления ран), сокращение мышц, поднимающих волосы.
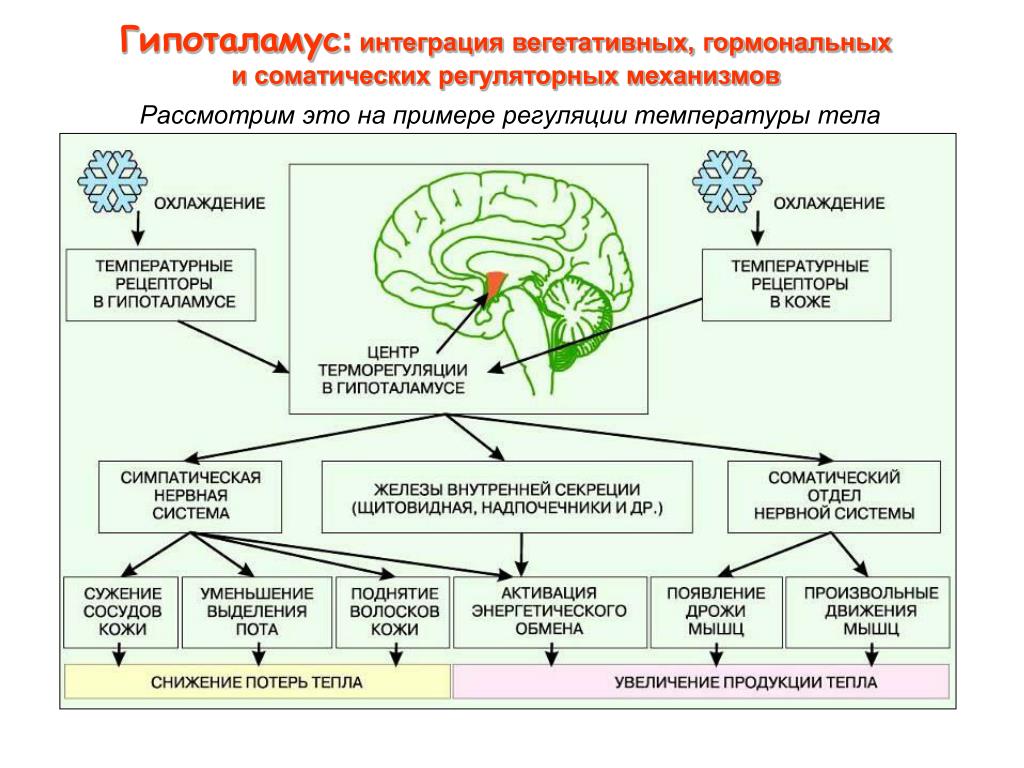
Гипоталамус — главный подкорковый центр интеграции вегетативных функций, имеет существенное значение в поддержании оптимального уровня обмена веществ (белкового, углеводного, жирового, минерального, водного) и терморегуляции. За счет связей с таламусом он получает разностороннюю информацию о состоянии органов и систем организма, а вместе с гипофизом образует функциональный комплекс — гипоталамо-гипофизарную систему.
 Гипоталамус в ней выполняет
роль своеобразного реле, включающего
гипофизарную гормональную цепь в
регуляцию различных висцеральных и
соматических функций.
Гипоталамус в ней выполняет
роль своеобразного реле, включающего
гипофизарную гормональную цепь в
регуляцию различных висцеральных и
соматических функций.Полосатое тело имеет ближайшее отношение к безусловнорефлекторной регуляции вегетативных функций. Повреждение или раздражение ядер полосатого тела вызывает изменение кровяного давления, усиление слюно- и слезоотделения, усиление потоотделения.
Высшим
центром регуляции вегетативных и
соматических функций, а также их
координации является кора
полушарий большого мозга.
Непрерывный поток импульсов от органов
чувств, сомы и внутренних органов по
афферентным путям поступает в кору
головного мозга и через эфферентную
часть вегетативной нервной системы,
главным образом через гипоталамус, кора
оказывает соответствующее влияние на
функцию внутренних органов, обеспечивая
адаптацию организма к меняющимся
условиям окружающей и внутренней среды.
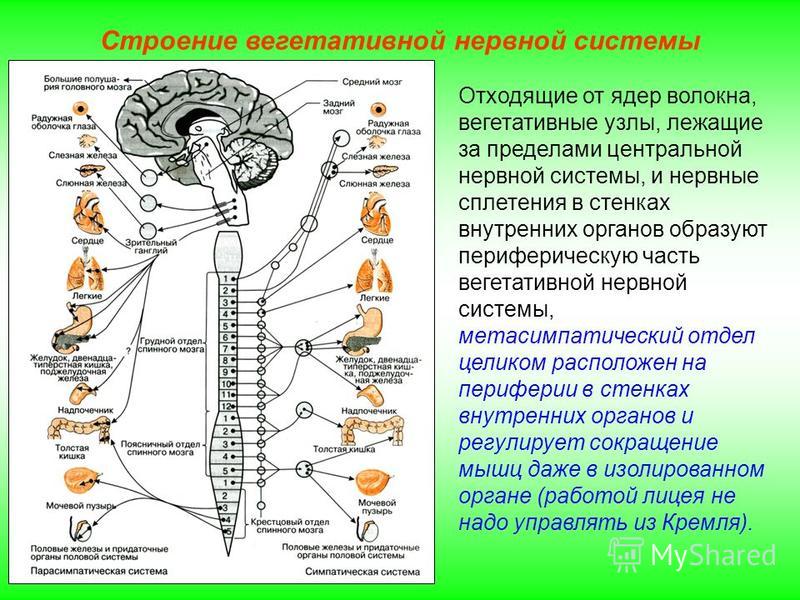
Таким образом, вегетативная нервная система, так же как и вся нервная система, построена по принципу иерархии, подчиненности. Схему организации вегетативной иннервации иллюстрирует рис.1.
Рис. 1 Принцип организации вегетативной нервной системы.
Изменения вегетативной функции у женщин с врожденными пороками развития центральной нервной системы плода
HEALTH OF WOMAN. 2016.2(108):121–124
Изменения вегетативной функции у женщин с врожденными пороками развития центральной нервной системы плода
Подольский В. В., Авраменко Т. В., Шевченко А. А.
ГУ «Институт педиатрии, акушерства и гинекологии НАМН Украины», г. Киев
Цель исследования

Материалы и методы. Проведена оценка функционального состояния вегетативной нервной системы у 45 беременных с диагностированными врожденными пороками развития ЦНС плода на основе определения вегетативного индекса Кердо (ВИ), коэффициента Хильдебрандта (КХ) и вегетативных рефлексов.
Результаты. Установлено повышение интенсивности и патологический характер вегетативных рефлексов, нарушение межсистемных соотношений с повышением КХ и ВИ на фоне действия стрессового фактора.
Заключение. Данные аспекты необходимо учитывать при ведении беременности и подготовке к родоразрешению.
Ключевые слова: вегетативная нервная система, беременность, врожденные пороки, центральная нервная система, плод.
Литература:
1. Abramchenko VV. 1996. Active management of labor. SPb., «Special literature»: 667.
2. Abramchenko VV. 2001. Psychosomatic obstetrics. SPb., «SOTIS»: 320.
3. Baevsky RM, Berseneva AP. 1997. Evaluation of adaptive capabilities of the organism and the risk of diseases. M: 236.
1997. Evaluation of adaptive capabilities of the organism and the risk of diseases. M: 236.
4. Batuev AS, Sokolova LV. 1994. The teaching of the dominant as the theoretical basis of formation of the system «Mother-child». Bulletin of St. Petersburg State University. 2:85-102.
5. Wayne AM. 2003. Autonomic disorders. M. «Medical News Agency»: 749.
6. Gusak JK, Nazarov Yu, Morozov VN. 2000. Psycho-neuro-immunological features of adaptive mechanisms of the normal reproductive cycle in women. Bulletin of new medical technologies 7(1):97-100.
7. Kustarov VN, Linde VA. 2000. Preeclampsia. St. Petersburg.: 104-105, 119.
8. Milovanov AP. 1999. Pathology system mother-placenta-fetus: A Guide for Physicians. M.
Medicine:448.
9. Pernu L. 1989. I am waiting for a child. M. Medicine:288.
10. Sidelnikova VM, Shmakov RG. 2004. Mechanisms of adaptation and disadaptation of hemostasis at pregnancy. M. Triada-X:192.
11. Stepanova SI. 1986. Biorhythmological aspects of adaptation problems. M. Science:240.
Stepanova SI. 1986. Biorhythmological aspects of adaptation problems. M. Science:240.
12. Tabelskaya TV, Tezikov Y, Lipatov IS, Frolova NA. 2015. Pathogenic orientation women dysadaptation to the gestation process. International Research Journal. 3-4 (34). URL: http://cyberleninka.ru/article/n/patogeneticheskaya-napravlennost-dezadaptatsii-zhenschin-k-protsessu-gestatsii (date of the application: 13.02.2016).
13. Talalaenko YuA, Bagrov AE, Danilova YN. 2011. Vegetative changes at pregnancy and their relation to hormonal status and development of gestational hypertensive disorders. Medical and social problems of the family 1(16):44-50.
14. Filippova GG. 2002. Psychology of motherhood. M: 240.
15. Khadartsev AA, Morozov VN, Grachev Y, Hadartseva KA, Gordeeva AY. 2012. Psycho-neuro-immunological adaptation of the program as a model dysadaptation at women with impaired reproductive cycle. Fundamental researches 5:359-365.
16. Alexa ID. 1995.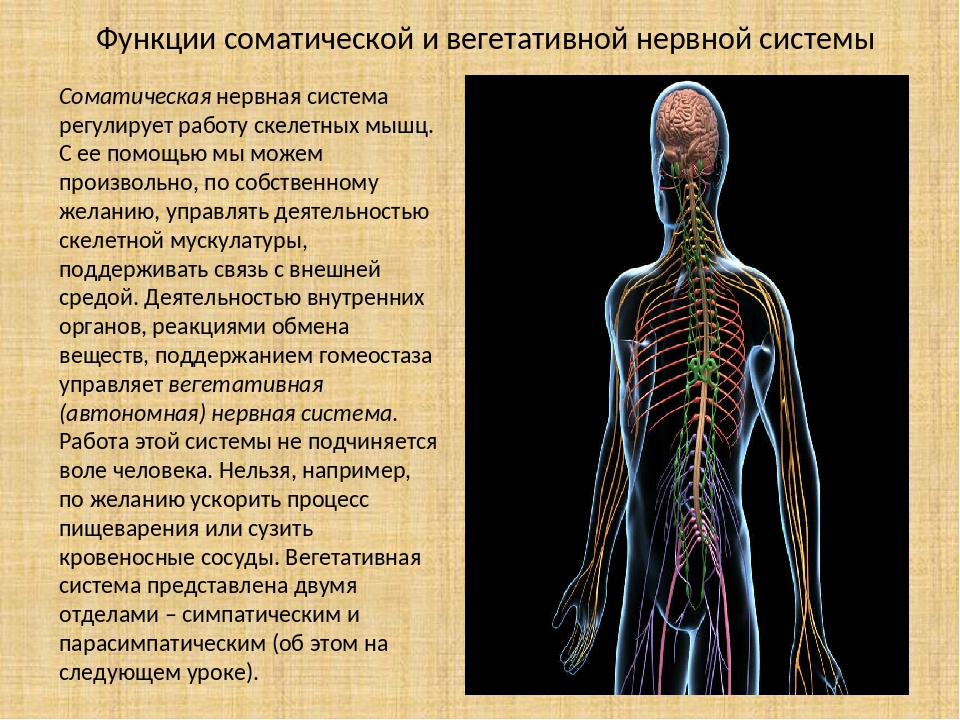 The neurohormonal mechanisms involved in the etiopathogenesis of arterial
The neurohormonal mechanisms involved in the etiopathogenesis of arterial
hypertension in pregnancy. Rev Med Chir Soc Med Nat Iasi. 99(1-2):43-51. PMid:9524655
17. Kuo CD, Chen GY, Yang MJ, Lo HM, Tsai YS. 2000. Biphasic changes in autonomic nervous activity during pregnancy. Br J Anaesth. 84(3):323-329. http://dx.doi.org/10.1093/oxfordjournals.bja.a013433; PMid:10793590
18. Matsuo H, Inoue K, Hapsari ED, Kitano K, Shiotani H. 2007. Change of autonomic nervous activity during pregnancy and its modulation of labor assessed by spectral heart rate variability analysis. Clin Exp Obstet Gynecol. 34(2):73-79. PMid:17629156
19. Ross LE, McLean LM. 2006. Anxiety disorders during pregnancy and the postpartum period: A systematic review 67(8):1285-98. http://dx.doi.org/10.4088/jcp.v67n0818
20. Soares CN, Steiner M. 2009. Perinatal depression: searching for specific tools for a closer look at this window. J Clin Psychiatry 70(9):1317-18. http://dx. doi.org/10.4088/JCP.09com05475; PMid:19818251
doi.org/10.4088/JCP.09com05475; PMid:19818251
Оценка функции сосудов и вегетативной нервной системы
Исследование рекомендуется взрослым пациентам с повышенным риском получения или развития заболеваний периферических артерий в качестве ангиологического скрининга, а также практически здоровым лицам.
Продолжительность исследования 20 мин.
Исследование позволит оценить следующие ваши индивидуальные показатели.
- Лодыжечно–плечевой индекс – маркер атеросклероза.
- Скорость распространения пульсовой волны. Позволяет оценить жесткость артериальных сосудов, выявить ранние признаки начинающегося атеросклероза, что позволяет принять своевременные меры по лечению и профилактике этого заболевания.
- Сатурация, оксигенация крови, насыщенность артериальной крови кислородом в покое и при напряжении (проба Вальсальвы) — один из основных показателей нормально функционирующего организма.

- Маркер инсулиновой резистентности организма. Инсулинорезистентность – предвестник диабета второго типа.
- Маркер эндотелиального гомеостаза. Нормальное функционирование эндотелия обеспечивает адекватную циркуляцию крови по сосудам, атеросклеротические поражения в первую очередь развиваются на поврежденном эндотелии, в эндотелии сосудов вырабатываются очень мощные факторы – местные гормоны, влияющие на тонус сосудов и соответственно на артериальное давление, выявление нарушений факторов эндотелиальной регуляции позволяет судить о функционировании организма в целом.
- Маркеры реакции на стресс. Измерение кожно-гальванического рефлекса, связанного с функцией потовых желез – может рассматриваться как показатель неспецифической нервно – психической напряженности и эмоциональности, в психофизиологии электрическую активность кожи используют как показатель ориентировочной или оборонительной реакции на внешние стимулы внутреннего эмоционального напряжения, эмоциональная активность регистрируется на ладонях и подошвах в обычных температурных условиях и усиливается в ответ на внешние возбуждающие стимулы
- Маркеры состава тела: масса сухой нежировой ткани тела (мышцы, кости), % к общей массе тела; масса жировой ткани, % к общей массе тела;-общий объем воды в теле, % к общей массе тела;-индекс массы тела.
 Опасность ожирения состоит в его ассоциации с большим количеством кардиометаболических осложнений – дислипидемией, артериальной гипертонией, ишемической болезнью сердца, сахарным диабетом 2 типа. Ожирение препятствует эффективной профилактике этих хронических заболеваний, осложняет их течение. Стандартные дозы лекарственных препаратов при нем часто менее эффективны. Причинно – следственная связь ожирения с серьезными метаболическими нарушениями и сердечно – сосудистыми заболеваниями определяют важность коррекции массы тела для снижения кардиометаболического риска.
Опасность ожирения состоит в его ассоциации с большим количеством кардиометаболических осложнений – дислипидемией, артериальной гипертонией, ишемической болезнью сердца, сахарным диабетом 2 типа. Ожирение препятствует эффективной профилактике этих хронических заболеваний, осложняет их течение. Стандартные дозы лекарственных препаратов при нем часто менее эффективны. Причинно – следственная связь ожирения с серьезными метаболическими нарушениями и сердечно – сосудистыми заболеваниями определяют важность коррекции массы тела для снижения кардиометаболического риска. - Оценка кардиометаболического риска.
Кроме того, вам будет предложена программа оздоровления с маркерами стиля жизни и рекомендации по питанию.
СТОИМОСТЬ УСЛУГ
Запись на диагностические процедуры по телефонам:
отделение функциональной диагностики +7 (499) 262-46-12
регистратура +7 (499) 262-35-99
Поликлиника ЦКБ «РЖД-Медицина»
г. Москва, ул. Новая Басманная, д. 5
Москва, ул. Новая Басманная, д. 5
Образец результатов исследования
вегетативные функции организма — Translation into English — examples Russian
These examples may contain rude words based on your search.
These examples may contain colloquial words based on your search.
Suggest an example
Other results
Он имеет полный контроль над большей частью своих вегетативных функций организма.
Он возникает в связи с разрушением нейрохимических молекл в процессе транспортации, что отражается на моторных функциях организма, состоянии его вегетативных систем и на высшей нервной деятельности.
It is caused by a breakdown of neuro-chemical molecules during transport, affecting the body’s motor functions, autonomic systems and the brain’s higher reasoning centers.
Любое нарушение структуры или функций организма…
Другими словами, инферон усиливает защитные функции организма.
Персонал отделения постоянно контролирует все функции организма пациента.
В этом случае напрямую усиливается восстановительная функция организма, не усиливая патологический сигнал.
With this technology the restorative function of the body is amplified directly, without amplifying the pathological signal. Целью Национального института физиологических наук является уточнение «механизма нормальных функций организма человека», который является основой медицинской науки.
Человек утрачивает контроль над двумя процентами функций организма каждый месяц.
Всё это уже автоматизированные функции организма.
Блокирует натриевый канал, парализует мышцы и замедляет жизненные функции организма.
Те, что отвечают за репродуктивные функции организма, Бекки.
Those that control fertility, Becky. Этот ужасный термин означает пожизненное ухудшение физических и умственных функций организма.
Говорится лишь, что смерть есть окончательное прекращение жизненных функций организма, конец жизни.
It only says that death is the final cessation of vital bodily functions; the end of life.Все передвижения, функции организма и мыслительные процессы тщательно проверяются.
Every movement, body function and thought process closely monitored.Я не понимаю, почему я должен читать книги о функциях организма.
Показатели жизненно важных функций организма замедляются настолько, что они имитируют смерть.
Vital signs so slow they mimic death.
Аннабет, ты не должна говорить шепотом о функциях организма в моем доме.
Автономная нервная система контролирует все функции организма, и отключает его, это один из способов изолировать вирус бешенства.
The autonomic nervous system controls all the functions of the body, and shutting it down is one of the ways rabies decimates its victims.Самое важное — это то, что мобильность зачастую снижается в результате ослабления функций организма.
Синдром вегетативных нарушений — диагностика и лечение в медицинском центре «Андреевские больницы
Синдром вегетативной дистонии заключается в нарушении нормального регулирования таких вегетативных функций организма, как дыхание, кровоснабжение, потоотделение, мочеиспускание и другие.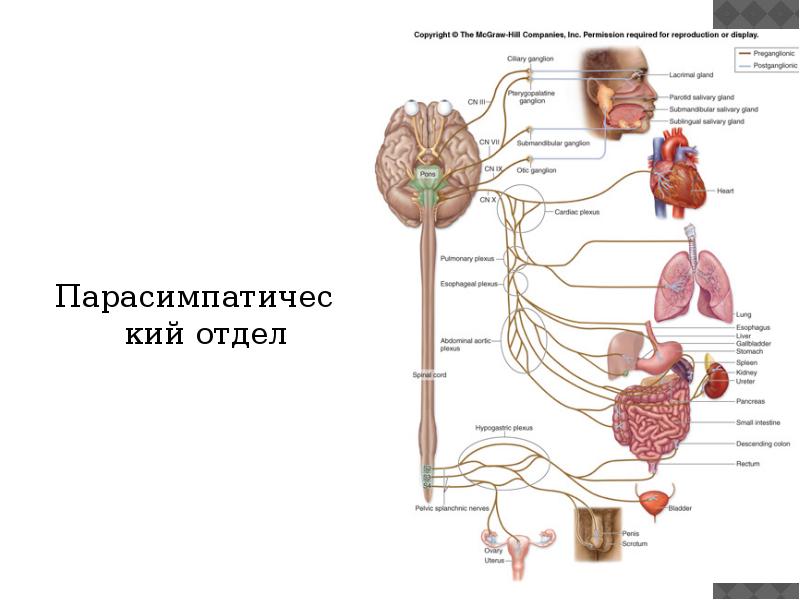 Этот синдром очень распространенный и имеет разнообразные проявления. При синдроме вегето-сосудистой дистонии могут быть задействованы почти все органы и системы, потому что в их иннервации принимает участие как центральная нервная система (то есть головной и спинной мозг), так и вегетативная нервная система, включающая парасимпатические и симпатические нервы.
Этот синдром очень распространенный и имеет разнообразные проявления. При синдроме вегето-сосудистой дистонии могут быть задействованы почти все органы и системы, потому что в их иннервации принимает участие как центральная нервная система (то есть головной и спинной мозг), так и вегетативная нервная система, включающая парасимпатические и симпатические нервы.
Причины синдрома вегетативных нарушений
Обычно этот синдром возникает в результате постоянного перенапряжения нервной системы. Может возникать из-за различных факторов, например сильного стресса, болезней, травм, злоупотребления спиртным. Перенапряжение клеток головного мозга сразу переходит в структуры спинного мозга, потом через вегетативные нервы в вегетативные узлы. Вегетативными узлами называются скопления нервных клеток с боков от позвоночника.
Вегетативные нарушения бывают распространенными, возникающими в нескольких системах органов в одно и то же время, системными, затрагивающими какую-то одну систему органов, или местными — при покраснении определенного участка кожи. Наиболее часто вегетативные проявления охватывают сердечно-сосудистую систему, так как она имеет наибольшее психологическое значение для человека.
Наиболее часто вегетативные проявления охватывают сердечно-сосудистую систему, так как она имеет наибольшее психологическое значение для человека.
Виды и симптомы синдрома вегетативных нарушений
Вегетативные нарушения обычно имеют симпатические и парасимпатические синдромы, смотря какие преобладают нарушения в той или иной области вегетативной системы организма.
К симпатическим синдромам относят симпатоадреналовые кризы, которые характеризуются неприятными ощущениями в области грудной клетки и в голове, сердцебиением, повышенным артериальным давлением и расширенными зрачками глаз. Может возникнуть чувство страха и непонятной тревоги. Заканчивается приступ обильным мочеиспусканием.
Парасимпатические кризы имеют симптомы, противоположные симпатическим проявлениям. Для них характерно головокружение, тошнота, понижение артериального давления, возникновение нарушения ритма сердца и замедленное его сокращение. Характерно чувство затрудненности дыхания и недостатка воздуха, может появиться желудочно-кишечное расстройство в виде метеоризма и позывов на дефекацию.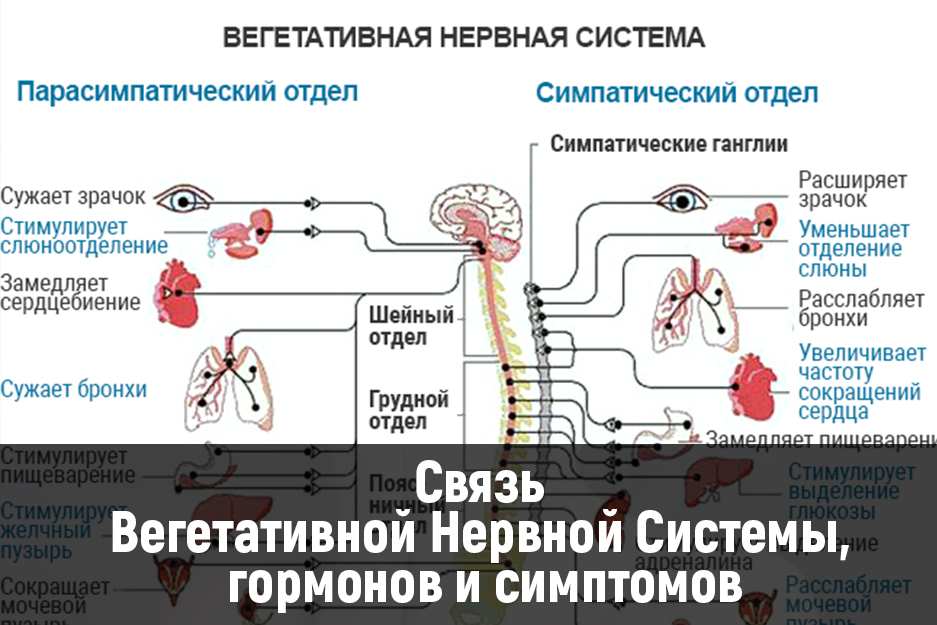
Часто синдром вегетативных нарушений носит смешанный симпато-парасимпатический характер. Вегетативные нарушения встречаются при разных заболеваниях и в качестве самостоятельного синдрома. Например, синдром вегетативной дистонии бывает первичным, конституционально обусловленным. ВСД как симптом возникает при эндокринных болезнях щитовидной железы и надпочечников, а также при эндокринных перестройках организма в норме, во время подросткового переходного возраста, беременности или климакса. Следующими по частоте являются неврозы. Вегетативные нарушения возникают при аллергиях и хронических болезнях внутренних органов.
Диагностика синдрома вегетативных нарушений
Проводится исследование вариабельности сердечного ритма, в ходе которого записывается кардиоритмограмма в течение 10 минут. Главная цель обследования – узнать, как вегетативная нервная система управляет работой сердца. Для этого проводится исследование, когда пациент находится в состоянии лежа, а затем — в состоянии стоя.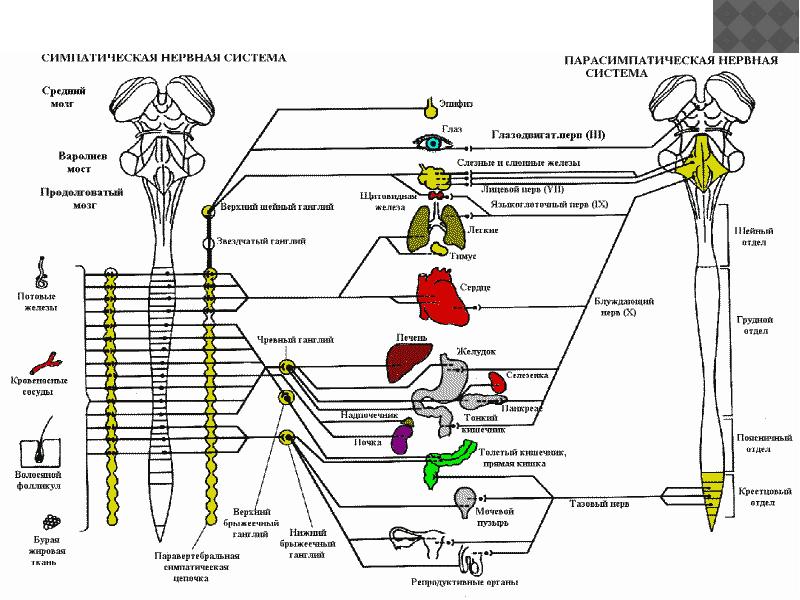 В норме при вставании его пульс должен резко участиться на 12 ударов в минуту, а потом через 20-30 ударов возвратиться к исходной норме. Когда изменения пульса, которые происходят в момент вставания, не укладываются в эти рамки, то существует нарушение деятельности вегетативной системы.
В норме при вставании его пульс должен резко участиться на 12 ударов в минуту, а потом через 20-30 ударов возвратиться к исходной норме. Когда изменения пульса, которые происходят в момент вставания, не укладываются в эти рамки, то существует нарушение деятельности вегетативной системы.
Лечение синдрома вегетативных нарушений
В лечении синдрома вегетативных нарушений ориентируются на причину появления синдрома или на основное заболевание. К главным группам препаратов для терапии синдрома вегетативных нарушений относят седативные и сосудистые лекарственные средства, иногда используют антигистаминные противоаллергические препараты.
Профилактика синдрома вегетативных нарушений
Необходимо избегать постоянного перенапряжения нервной системы, сильных стрессов, инфекционных болезней, травм, злоупотребления спиртным. Стараться больше отдыхать и следить за весом.
Расстройство вегетативной нервной системы: тревога, невроз, панические атаки
Многочисленные стрессы, постоянно сопровождающие человека, оказывают негативное влияние на нервную систему и становятся главной причиной психологического напряжения, нарушения работы органов и систем. Все это в конечном итоге приводит к истощению внутренних ресурсов, ухудшению иммунитета и развитию тревожных расстройств.
Все это в конечном итоге приводит к истощению внутренних ресурсов, ухудшению иммунитета и развитию тревожных расстройств.
Симптоматика заболевания может быть схода с мигренью, остеохондрозом, инфарктом и рядом других состояний, поэтому консультация невролога обязательна.
Вегетативная дисфункция: основные типы
Выделяют следующие типы вегетативной дисфункции (не путать с вегетососудистой дистонией (ВСД), являющейся лишь проявлением ряда заболеваний нервной системы и других органов):
- Соматофорное расстройство. Это невроз, проявляющийся в виде симптомов отсутствующих у пациента хронических заболеваний. Возможны панические атаки, приступы кашля и одышки психогенного происхождения, проблемы с пищеварением, периодические головокружения и т.д. Данный тип дисфункции в большинстве случаев обусловлен сильным или продолжительным стрессом, поэтому достаточно легко поддается лечению.
- Нарушение подкорковых структур. Повреждения возникают вследствие перенесенных травм головного мозга или резидуальной патологии ЦНС (часто диагностируется у детей).
 Для заболевания характерно пониженное артериальное давление, полуобморочное состояние, диарея и частое мочеиспускание.
Для заболевания характерно пониженное артериальное давление, полуобморочное состояние, диарея и частое мочеиспускание. - Раздражение периферических вегетативных структур, спровоцированное поражением симпатического шейного сплетения или мочекаменной болезнью.
Вегетативная дисфункция, которой страдают около 70% взрослых и 25% детей, всегда свидетельствует о наличии проблем в организме, а значит, является серьезным поводом обратиться к специалисту. Эффективность назначенного лечения во многом зависит от своевременной постановки правильного диагноза. Записаться к детскому неврологу в Уфе можно по телефону +7 (347) 216 00 22.
Причины и методы лечения
Согласно последним данным, расстройство вегетативной нервной системы может возникнуть на фоне хронического стресса, гормонального сбоя, малоподвижного образа жизни, неправильного питания, употребления табачных и алкогольных изделий, воспалительных процессов и длительного приема лекарств (в том числе самолечение). Одним из факторов является наследственность.
Одним из факторов является наследственность.
Скорректировать состояние больного можно при помощи следующих немедикаментозных методов:
- Массаж и иглорефлексотерапия. Основная задача – релаксация, снятие мышечных зажимов и улучшение кровообращения. Иглорефлексотерапия практически не имеет противопоказаний и активно применяется при лечении неврозов и других расстройств нервной системы у взрослых и детей. Благотворное влияние данных методов на состояние пациента подтверждено клинически, поэтому медицинский массаж в Уфе является одной из самых востребованных процедур.
- Психотерапия. Нередко расстройства нервной системы развиваются вследствие психических особенностей человека. Ярким примером является трудоголизм. Человек, полностью захваченный работой и не умеющий выполнять задания спустя рукава, пребывает в состоянии постоянного стресса. Не меньшим потрясением (=пусковым механизмом) может стать развод. Проработка жизненной ситуации с психотерапевтом позволяет справиться не только с первопричиной заболевания, но и с ее последствиями.

Записаться на прием кардиолога в Уфе, невролога или ортопеда можно на нашем сайте! Выберите удобное время, оставьте комментарий и дождитесь звонка нашего оператора.
Вызванные потенциалы мозга
Вызванные потенциалы мозга (ВП) – тестирование функции зрительного, акустического и соматосенсорного анализаторов. Исследуются когнитивные функции на основе эндогенного когнитивного вызванного потенциала, и исследуется центральное и периферическое звено вегетативной нервной системы с помощью вызванного кожного симпатического потенциала.
Эндогенные (когнитивные) вызванные потенциалы.
Производится оценка высших корковых функций мозга человека, таких как распознавание стимула, дифференцировка, запоминание и мыслительные процессы, связанные с принятием решения.
|
|
Показания к обследованию:
|
Соматосенсорные вызванные потенциалы.
В ходе этого исследования проводится:
- оценка сохранности афферентных путей при диагностике поражений спинного и головного мозга, периферических нервов;
- определение тяжести повреждения мозга и возможных последствий травмы и комы;
|
Показания к обследованию:
|
Акустические стволовые вызванные потенциалы.
Проводится оценка функционального состояния периферических и стволовых акустических структур.
Показания к обследованию:
|
Зрительные вызванные потенциалы.
Оценка состояния зрительного нерва, хиазмы, зрительного тракта и зрительной коры, остроты зрения и ее корригируемости, зрительных нарушений и их динамики при лечении.
Показания к обследованию:
|

Вызванные кожные симпатические потенциалы – оценка функции вегетативной нервной системы.
Показания к применению:
- дифференциальная диагностика нарушений вегетативной регуляции у больных неврологического и терапевтического профиля и вегетативных полинейропатий;
- оценка влияния неблагоприятных факторов профессиональной деятельности, индивидуальный подбор фармакотерапии вегетативных кризов, нейроциркуляторной дистонии.
Вегетативное состояние — Заболевания головного мозга, спинного мозга и нервов
Подобно людям, находящимся в коме, людям в вегетативном состоянии требуется комплексная помощь.
Обеспечение полноценного питания (нутритивной поддержки) очень важно. Людей кормят через зонд, вводимый через нос в желудок (так называемое зондовое питание). Иногда их кормят через зонд, вводимый непосредственно в желудок или тонкий кишечник через разрез в брюшной полости. Через эти трубки можно также вводить лекарства.
Через эти трубки можно также вводить лекарства.
Многие проблемы возникают из-за невозможности двигаться (иммобилизация), и меры по их предотвращению очень важны (см. Проблемы, связанные с постельным режимом). Например, может произойти следующее:
Пролежни: Лежание в одном положении может перекрыть кровоснабжение некоторых участков тела, вызывая разрушение кожи и образование пролежней.
Контрактуры: Недостаток движения также может привести к необратимому скованности и укорочению мышц (контрактурам), вызывая постоянное искривление суставов.
Сгустки крови: Отсутствие движения увеличивает вероятность образования сгустков крови в венах ног — это называется тромбозом глубоких вен.
Пролежни можно предотвратить, часто меняя положение человека и помещая защитные прокладки под части тела, которые соприкасаются с кроватью, например, пятки, чтобы защитить их.
Для предотвращения контрактур физиотерапевты осторожно перемещают суставы человека во всех направлениях (пассивные упражнения на диапазон движений) или накладывают шины на суставы в определенных положениях.
Предотвращение образования тромбов включает использование лекарств и сжатие или приподнятие ног человека. Движение конечностей, как это происходит в упражнениях с пассивным диапазоном движений, также может помочь предотвратить образование тромбов.
Поскольку люди страдают недержанием, необходимо следить за тем, чтобы кожа оставалась чистой и сухой. Если мочевой пузырь не функционирует и моча задерживается, в мочевой пузырь может быть помещена трубка (катетер) для слива мочи. Катетеры тщательно очищают и регулярно осматривают, чтобы предотвратить развитие инфекций мочевыводящих путей.
Кома и стойкое вегетативное состояние
Обзор
Что такое кома?
Кома, иногда также называемая стойким вегетативным состоянием, — это глубокое или глубокое бессознательное состояние. Стойкое вегетативное состояние — это не смерть мозга. Человек в состоянии комы жив, но не может двигаться или реагировать на свое окружение.Кома может возникнуть как осложнение основного заболевания или в результате травм, например, травмы головы. . Люди в таком состоянии потеряли свои мыслительные способности и понимание своего окружения, но сохраняют некогнитивные функции и нормальный режим сна. Даже несмотря на то, что те, кто находится в устойчивом вегетативном состоянии, теряют свои высшие функции мозга, другие ключевые функции, такие как дыхание и кровообращение, остаются относительно неизменными. Могут происходить спонтанные движения, а глаза могут открываться в ответ на внешние раздражители.Люди могут даже иногда гримасничать, плакать или смеяться. Хотя люди в устойчивом вегетативном состоянии могут казаться несколько нормальными, они не говорят и не могут реагировать на команды.
Стойкое вегетативное состояние — это не смерть мозга. Человек в состоянии комы жив, но не может двигаться или реагировать на свое окружение.Кома может возникнуть как осложнение основного заболевания или в результате травм, например, травмы головы. . Люди в таком состоянии потеряли свои мыслительные способности и понимание своего окружения, но сохраняют некогнитивные функции и нормальный режим сна. Даже несмотря на то, что те, кто находится в устойчивом вегетативном состоянии, теряют свои высшие функции мозга, другие ключевые функции, такие как дыхание и кровообращение, остаются относительно неизменными. Могут происходить спонтанные движения, а глаза могут открываться в ответ на внешние раздражители.Люди могут даже иногда гримасничать, плакать или смеяться. Хотя люди в устойчивом вегетативном состоянии могут казаться несколько нормальными, они не говорят и не могут реагировать на команды.
Есть ли лечение комы?
Когда человек оказывается вне непосредственной опасности, медицинская бригада сосредотачивается на предотвращении инфекций и поддержании здорового физического состояния. Это часто включает профилактику пневмонии и пролежней и обеспечение сбалансированного питания.Физиотерапия также может использоваться для предотвращения контрактур (постоянных мышечных сокращений) и деформаций костей, суставов и мышц, которые ограничивают восстановление для тех, кто выходит из комы.
Это часто включает профилактику пневмонии и пролежней и обеспечение сбалансированного питания.Физиотерапия также может использоваться для предотвращения контрактур (постоянных мышечных сокращений) и деформаций костей, суставов и мышц, которые ограничивают восстановление для тех, кто выходит из комы.
Каков прогноз (перспективы) для комы?
Исход комы и стойкого вегетативного состояния зависит от причины, тяжести и места неврологического повреждения. Люди могут выйти из комы с сочетанием физических, интеллектуальных и психологических проблем, требующих особого внимания.Выздоровление обычно происходит постепенно, и некоторые из них приобретают все большую способность реагировать. Некоторые люди никогда не продвигаются дальше самых простых реакций, но многие полностью восстанавливают сознание. Лица, выздоравливающие из комы, нуждаются в тщательном медицинском наблюдении. Кома редко длится более 2–4 недель. Некоторые пациенты могут восстанавливать сознание после стойкого вегетативного состояния. Другие могут оставаться в этом состоянии годами или даже десятилетиями. Наиболее частой причиной смерти людей, находящихся в устойчивом вегетативном состоянии, являются инфекции, например пневмония.
Другие могут оставаться в этом состоянии годами или даже десятилетиями. Наиболее частой причиной смерти людей, находящихся в устойчивом вегетативном состоянии, являются инфекции, например пневмония.
Какие исследования проводятся?
Национальный институт неврологических расстройств и инсульта (NINDS) и другие институты Национальных институтов здравоохранения (NIH) проводят исследования, связанные с комой, в своих лабораториях в NIH, а также поддерживают дополнительные исследования посредством грантов для крупных медицинских учреждений по всей стране. Большая часть этого исследования посвящена поиску более эффективных способов предотвращения и лечения комы.
Ресурсы
Организации
Американская ассоциация травм головного мозга, Inc.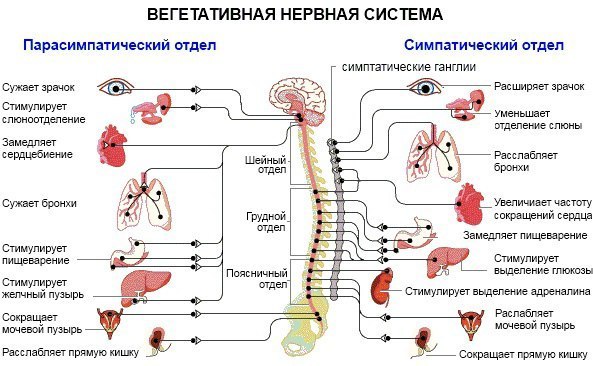
1608 Spring Hill Rd., Suite 110
Вена, VA 22182
Телефон: 703.761.0750
Бесплатный звонок: 800.444.6443
Эл. Почта: [email protected]
Веб-сайт: www.biausa.org
Фонд травмы мозга
1999 Южный Баском проспект, офис 1040
Кэмпбелл, Калифорния 95008
Телефон: 408.369.9735
Эл. Почта: [email protected]
Веб-сайт: www.braintrauma.org
Coma / Traumatic Brain Injury Recovery Association, Inc.
8300 Республика аэропорт, офис 106
Farmingdale, NY 11735
Телефон: 631-756-1826
Альянс семейных опекунов / Национальный центр опеки
Маркет 785, офис 750
Сан-Франциско, Калифорния 94103
Телефон: 415.434.3388
Бесплатный звонок: 800.445.8106
Факс: 415.434.3508
Эл. Почта: info@caregiver. org
org
Веб-сайт: www.caregiver.org
Национальный реабилитационный информационный центр (NARIC)
8400 Корпоративный Диск, Люкс 500
Landover, MD 20785
Телефон: 301.459.5900
Бесплатный звонок: 800.346.2742
Телетайп: 301.459.5984
Электронная почта: [email protected]
Веб-сайт: www.naric.com
Источник: Национальные институты здравоохранения; Национальный институт неврологических расстройств и инсульта *
Диагностика вегетативной функции для раннего выявления отравления свинцом
Abel H-H, Klüssendorf D, Koepchen H-P (1989) Связь между тонусом и ритмичностью хронотропной иннервации сердца.Арка Пфлюгера 413: R11
Google ученый
 org/Book»>
org/Book»>Abel HH, Klüssendorf D, Koralewski E, Krause R, Droh R (1992) Спектральный анализ частоты сердечных сокращений во время различных состояний активности. В: Schmidt THF, Engel BT, Blümchen G (eds) Временные вариации сердечно-сосудистой системы. Springer, Berlin Heidelberg New York, стр. 100–116
Google ученый
Bleecker, ML, Bolla KI, Agnew J, Schwartz BS, Ford DP (1991) Дозозависимые субклинические нейроповеденческие эффекты при хроническом воздействии низких уровней органических растворителей.Am J Ind Med 19: 715–728
PubMed Статья CAS Google ученый
Eckoldt K (1975) Untersuchungen über die Wirkungen der Vegetativen Herznerven mit Hilfe von unblutigen Meßverfahren. Habilitationsschrift, Humboldt Univ, Berlin, Biowiss. Фак.
Фак.
Google ученый
Eckoldt K (1984) Verfahren und Ergebnisse der Quantitativen Automatischen Analyze der Herzfrequenz und deren Spontanvariabilität.Dtsch Gesundheitswes 39: 856–863
Google ученый
Fahrenberg J (1969) Die Bedeutung Individualler Unterschiede für die Methodik der Aktivierungsforschung. Хубер, Берн, стр. 95–122
Google ученый
Gennart J, Bernard A, Lauwerys R (1992) Оценка функции щитовидной железы, яичек, почек и вегетативной нервной системы у рабочих, подвергшихся воздействию свинца. Int Arch Occup Environ Health 64: 49–57
PubMed Статья CAS Google ученый
 org/ScholarlyArticle»>
org/ScholarlyArticle»>Ирегрен А., Гамбераль Ф (1990) Поведенческая токсикология человека.Воздействие на центральную нервную систему низких доз нейротоксических веществ в рабочей среде. Scand J Work Environ Health 16 [Дополнение 1]: 17–25
PubMed Google ученый
Koepchen H-P (1977) Neurophysiologische Grundlagen der nervösen Steuerung der Herzfrequenz. Med Sport 17: 136–140
Google ученый
Koepchen H-P, Huopaniemi T (1991) Кардиореспираторная и моторная координация.Springer, Берлин Гейдельберг Нью-Йорк
Google ученый
Koepchen H-P, Abel H-H, Klüssendorf D (1989) Физиологические концепции сердечно-сосудистого и респираторного контроля: теоретическая основа и применимость к человеку. В: Droh, R, Spintge R (eds) Инновации в физиологической анестезии и мониторинге. Springer, Berlin Heidelberg New York, стр. 3–20
В: Droh, R, Spintge R (eds) Инновации в физиологической анестезии и мониторинге. Springer, Berlin Heidelberg New York, стр. 3–20
Google ученый
Lee DH, Park KS (1990) Многомерный анализ компонентов умственной и физической нагрузки в оценках синусовой аритмии.Эргономика 33: 35–47
PubMed CAS Google ученый
Малик М., Камм А.Дж. (1990) Электрофизиология, кардиостимуляция и аритмия. Clin Cardiol 13: 570–576
PubMed CAS Статья Google ученый
Мацумото Т., Фукая Ю., Йошитоми С., Арафука М., Кубо Н., Оно Ю. (1993) Взаимосвязь между воздействием свинца и периферическими нервно-мышечными функциями у рабочих, подвергшихся воздействию свинца: результаты теста на постукивание. Environ Res 61: 299–307
Environ Res 61: 299–307
PubMed Статья CAS Google ученый
Моллер Л., Кристенсен Т.С. (1992) Свинец в крови как фактор риска сердечно-сосудистых заболеваний. Am J Epidemiol 136: 1091–1100
PubMed CAS Google ученый
Moser M, Lehofer M, Sedminek A, Lux M, Zapotoczky H-G, Kenner T, Noordengraaf A (1994) Вариабельность сердечного ритма как прогностический инструмент в кардиологии.Тираж 90: 1078–1082
PubMed CAS Google ученый
Mulder G, Mulder LJM (1981) Обработка информации и сердечно-сосудистый контроль. Психофизиология 18: 392–402
PubMed Статья CAS Google ученый
 org/ScholarlyArticle»>
org/ScholarlyArticle»>Мурата К., Араки С. (1991) Дисфункция вегетативной нервной системы у рабочих, подвергшихся воздействию свинца, цинка и меди, в связи с проводимостью периферических нервов: исследование вариабельности интервала R-R.Am J Ind Med 20: 663–671
PubMed Статья CAS Google ученый
Мурата К., Араки С., Йокояма К., Учидея Е., Фудзимура Ю. (1993) Оценка функций центральной, периферической и вегетативной нервной системы у ведущих работников: нейроэлектрофизиологические исследования. Environ Res 61: 323–336
PubMed Статья CAS Google ученый
Muttray A, Jung D, Spüller M, Wildt C, Mayer-Popken O, Konietzko J (1991) Zur Wirkung von Blei auf die autonome Innervation des Herzens. В: Schäcke G, Ruppe K, Vogel-Sühring C (ред.) Arbeitsmedizin für eine gesunde Umwelt. 31. Jahrestagung in Berlin vom 11. bis 14. März 1991. Gentner, Stuttgart, pp 175–177
В: Schäcke G, Ruppe K, Vogel-Sühring C (ред.) Arbeitsmedizin für eine gesunde Umwelt. 31. Jahrestagung in Berlin vom 11. bis 14. März 1991. Gentner, Stuttgart, pp 175–177
Google ученый
Пагани М., Фурлан Р., Пиццинелли П., Кривелларо В., Черутти С., Маллиани А. (1989) Спектральный анализ вариабельности R-R и артериального давления для оценки симпато-вагусного взаимодействия во время психического стресса у людей. J Hypertens 7 [Дополнение 6]: 14–15
Google ученый
Павлоски Р. (1989) Системный подход к сердечно-сосудистой реактивности: поведенческие модели, которые ведут себя.Психофизиология 26: 468–481
PubMed Статья CAS Google ученый
 org/Book»>
org/Book»>Pfister E (1990) Die Bestimmung der Dynamischen Dauerleistungsfahigkeit Jugendlicher beim Einsatz unterschiedlicher Muskelmassen zur Begründung von Grenzwerten der physischen Belastbarkeit im 16. bis 18. Lebensjher. Habilitationsschrift, Med. Академик, Магдебург
Google ученый
Pfister E, Hartmann B (1991) Вегетативный сердечно-сосудистый контроль в диапазоне предела выносливости для динамической мышечной работы.Pflügers Arch 419 [Дополнение 1]: 86
Google ученый
Pfister E, Böckelmann I, Brosz M, Ferl T, Winter C-G (1994) Ermittlung neurotoxischer Effekte infolge langjähriger Bleiexposition in der Kupferindustrie Sachsen-Anhalts anhand psy-chometrischer Leistungsdaten. Zentralbl Arbeitsmed 44: 422–432
CAS Google ученый
 org/Book»>
org/Book»>Schmidt TFH, Engel BT, Blümchen G (1992) Временные вариации сердечно-сосудистой системы.Sprnger, Берлин Гейдельберг Нью-Йорк
Google ученый
Schubert E (1984) Welche Erkenntnisse Können aus Untersuchungen des Herzrhythmus gewonnen werden? Dtsch Gesund-heitswes 39: 845–856
Google ученый
Schwarz V (1977) Die Beeinflussung der Herzfrequenz und der Sinusarrhythmie durch Pharmakologische Vagusblockade in Ruhe und bei ergometrischer Belastung.Med Sport 17: 148–149
Google ученый
Seppäläinen AM, Hernberg S, Kock B (1979) Взаимосвязь между уровнями свинца в крови и скоростью нервной проводимости. Нейротоксикология 1: 313–332
Нейротоксикология 1: 313–332
Google ученый
Сеппяляйнен А.М., Хернберг С., Весанто Р., Кок Б. (1983) Ранние нейротоксические эффекты профессионального воздействия свинца: проспективное исследование. Neuro Toxicol 4: 181–192
Google ученый
Sroczynski J, Biskupek K, Piontrowski J, Rudzki H (1990) Влияние профессионального воздействия свинца, цинка и кадмия на показатели системы кровообращения у металлургических рабочих (на польском языке) Med Pr 41: 152–158
PubMed CAS Google ученый
Стакак А.Дж., Фабиан З., Досталек С. (1987) Спектральный анализ изменчивости интервала R-R при задержке вдоха у человека в состоянии покоя и во время эмоционального напряжения. Act Nerv Super (Прага) 29: 264–269
Act Nerv Super (Прага) 29: 264–269
Google ученый
Teruya K, Sakurai H, Omae K, Higashi T, Muto T, Kaneko Y (1991) Влияние свинца на парасимпатическую функцию сердца. Int Arch Occup Environ Health 62: 549–553
PubMed Статья CAS Google ученый
Weber P, Regel H, Leo H, Krause A (1987) COMBITEST 2, ein Gerätesystem für die apparative Psychodiagnostik.З Клин Мед 42: 1821–1825
Google ученый
Журчание, вегетативная функция и развитие языка после резекции крикотрахеальной системы у детей с афонией — Experts @ Minnesota
@article {c3b930143f4a41b74857052a7c2,
title = «Бабуля, вегетативная функция и развитие речевой функции у
детей после резекции шейки матки» abstract = «Задача: определить важность проязычного лепета путем изучения моделей развития речи и языка после резекции крикотрахеального отдела у афоничных детей. Дизайн исследования: ретроспективный обзор семи детей с афонией, перенесших крикотрахеальную резекцию нашей педиатрической бригадой грудных дыхательных путей. Анализируемые переменные включают возраст, пол, сопутствующие заболевания, степень стеноза, длину резецированной трахеи и методы коммуникации. Методы. Данные о методах общения детей до и после операции, а также об использовании ими логопедических услуг были получены путем оценки патологии речи, клинических наблюдений и стандартного телефонного опроса, дополненного документацией родителей.Послеоперационное качество голоса оценивалось с помощью исследования педиатрических исходов голоса. Результаты: Всем семи пациентам была выполнена трахеостомия до достижения 2-месячного возраста с поправкой на недоношенность. Субъекты оставались афоническими на протяжении всего периода канюляции. После резекции крикотрахеального отдела у них была начальная задержка в овладении речью. Вегетативные функции были первыми звуками гортани. Первоначально дети могли издавать эти звуки только рефлекторно, но впоследствии они приобрели произвольный контроль над этими функциями гортани.
Дизайн исследования: ретроспективный обзор семи детей с афонией, перенесших крикотрахеальную резекцию нашей педиатрической бригадой грудных дыхательных путей. Анализируемые переменные включают возраст, пол, сопутствующие заболевания, степень стеноза, длину резецированной трахеи и методы коммуникации. Методы. Данные о методах общения детей до и после операции, а также об использовании ими логопедических услуг были получены путем оценки патологии речи, клинических наблюдений и стандартного телефонного опроса, дополненного документацией родителей.Послеоперационное качество голоса оценивалось с помощью исследования педиатрических исходов голоса. Результаты: Всем семи пациентам была выполнена трахеостомия до достижения 2-месячного возраста с поправкой на недоношенность. Субъекты оставались афоническими на протяжении всего периода канюляции. После резекции крикотрахеального отдела у них была начальная задержка в овладении речью. Вегетативные функции были первыми звуками гортани. Первоначально дети могли издавать эти звуки только рефлекторно, но впоследствии они приобрели произвольный контроль над этими функциями гортани.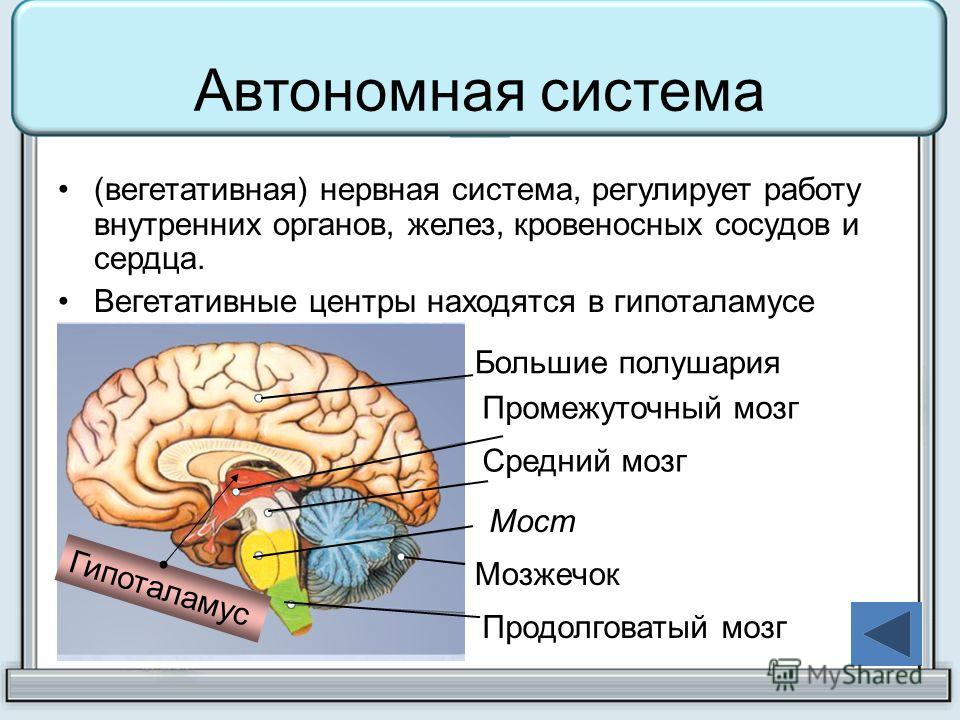 Все испытуемые прошли идентифицируемую стадию канонического лепета, которая часто происходила одновременно с вокализацией. За этим последовало появление истинной речи. Выводы: начальная задержка в овладении речью, наблюдаемая после деканюляции, наряду с наличием послеоперационной канонической стадии у всех испытуемых, подтверждает гипотезу о том, что лепет необходим для развития речи и языка. Кроме того, наличие лепета очевидно независимо от возраста, в котором развивается речь.Наконец, нет очевидной корреляции между предоперационным языком жестов и темпом развития речи. «,
Все испытуемые прошли идентифицируемую стадию канонического лепета, которая часто происходила одновременно с вокализацией. За этим последовало появление истинной речи. Выводы: начальная задержка в овладении речью, наблюдаемая после деканюляции, наряду с наличием послеоперационной канонической стадии у всех испытуемых, подтверждает гипотезу о том, что лепет необходим для развития речи и языка. Кроме того, наличие лепета очевидно независимо от возраста, в котором развивается речь.Наконец, нет очевидной корреляции между предоперационным языком жестов и темпом развития речи. «,
keywords =» Педиатрические дыхательные пути, лепет, резекция крикотрахеального канала, развитие речи, подъязычный стеноз «,
author =» Bohm, {Lauren A} и Нельсон, {Марк Э.} и Драйвер, {Линн Э.} и Грин, {Гленн Э.} «,
год =» 2010 «,
месяц = декабрь,
день =» 1 «,
doi = «10.1002 / lary.21094»,
language = «English (US)»,
volume = «120»,
pages = «2494—2497»,
journal = «Laryngoscope»,
issn = «0023-852X»,
publisher = «John Wiley and Sons Inc. «,
«,
number =» 12 «,
}
Вегетативные части растений | Служба расширения OSU
Внешние структуры растений, такие как листья, стебли, корни, цветы, плоды и семена, известны как органы растения . Орган — это организованная группа тканей, которые работают вместе для выполнения определенной функции. Эти структуры можно разделить на две группы: половые репродуктивные и вегетативные . Вегетативные части (рис. 1) включают корни, стебли, почки побегов и листья. ; они не принимают непосредственного участия в половом размножении.Вегетативные части часто используются в бесполых формах размножения, таких как черенки, бутонизация или прививка.
Корни
Часто корни упускаются из виду, вероятно, потому, что они менее заметны, чем остальная часть растения. Тем не менее, важно понимать корневые системы растений, потому что они оказывают заметное влияние на размер и жизнеспособность растений, способ размножения, адаптацию к типам почвы и реакцию на культурные обычаи и орошение.
Корни обычно происходят из нижней части растения или черенка.У них есть корневая шляпка, но отсутствуют узлы, и они никогда не имеют прямых листьев или цветов. Их основные функции — поглощать питательные вещества и влагу, закреплять растение в почве, поддерживать стебель и хранить пищу. У некоторых растений их можно использовать для размножения.
Конструкция
Внутри корень состоит из трех основных частей (рис. 2):
- Меристема находится на вершине и производит новые клетки; это область деления и роста клеток.
- За меристемой находится зона растяжения .В этой области клетки увеличиваются в размере за счет поглощения пищи и воды. По мере роста они проталкивают корень через почву.
- Зона созревания находится непосредственно под стеблем. Здесь клетки становятся специфическими тканями, такими как эпидермис, кора или сосудистая ткань.
Эпидермис корня — это самый внешний слой клеток (рис.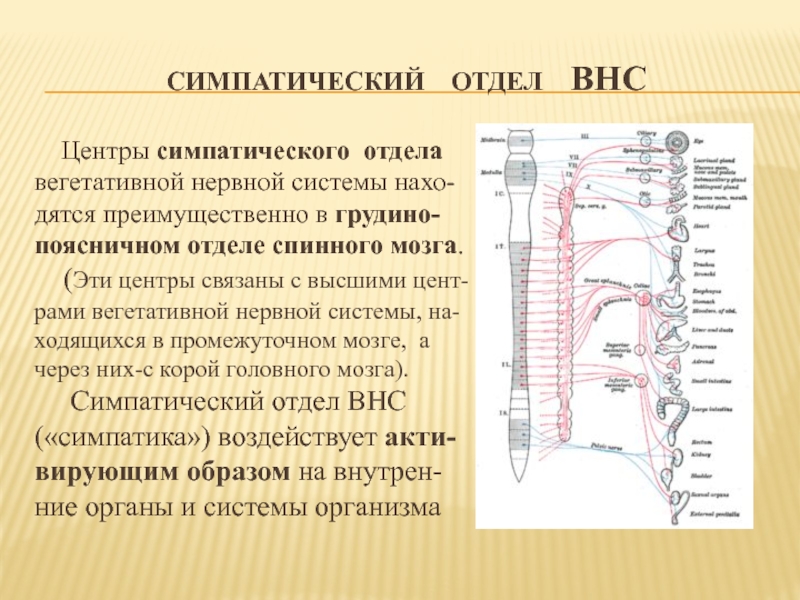 3). Эти клетки отвечают за поглощение воды и минералов, растворенных в воде. Клетки коры участвуют в перемещении воды из эпидермиса в сосудистую ткань (ксилему и флоэму) и в хранении пищи.Сосудистая ткань расположена в центре корня и проводит пищу и воду.
3). Эти клетки отвечают за поглощение воды и минералов, растворенных в воде. Клетки коры участвуют в перемещении воды из эпидермиса в сосудистую ткань (ксилему и флоэму) и в хранении пищи.Сосудистая ткань расположена в центре корня и проводит пищу и воду.
Внешне здесь важны две области: корневой покров и корневые волоски (рис. 2). Корневой колпачок — это крайний край корня. Он состоит из клеток, которые отшелушиваются по мере прорастания корня в почве. Его функция — защита корневой меристемы.
Корневые волоски — это тонкие удлиненные эпидермальные клетки, которые располагаются в небольшой зоне сразу за кончиком корня.Обычно они кажутся прекрасными невооруженным глазом. Их функция заключается в увеличении площади поверхности корня и абсорбционной способности. Корневые волоски обычно живут 1-2 дня. При пересадке растения они легко срываются или могут засохнуть на солнце.
Многие корни имеют естественные симбиотические (взаимовыгодные) отношения с определенными грибами, что улучшает способность растения поглощать воду и питательные вещества. Эта полезная ассоциация называется микориза (гриб + корень).
Эта полезная ассоциация называется микориза (гриб + корень).
Виды корней
Есть два основных типа корней: первичный, и боковой. Первичный корень берет начало в нижнем конце зародыша саженца. Если первичный корень продолжает вытягиваться вниз, становится центральным элементом корневой системы и имеет ограниченное вторичное ветвление, он называется стержневым корнем (рис. 4). У гикори и орехов пекан, а также у моркови есть стержневые корни.
Боковой корень или вторичный корень — это боковой корень или корень ветви, который возникает из другого корня.Если первичный корень перестает удлиняться и развиваются многочисленные боковые корни, образуется мочковатая корневая система. Эти боковые корни многократно разветвляются, образуя сеть питающих корней, которая есть у большинства растений.
Некоторые растения, такие как травы, в природе дают волокнистую корневую систему . В других случаях отрубание главного корня растения путем его подрезки может стимулировать образование волокнистой корневой системы. Питомники используют эту технику для деревьев, которые естественным образом дают стержневой корень, потому что деревья с компактной мочковатой корневой системой пересаживаются более успешно.
В других случаях отрубание главного корня растения путем его подрезки может стимулировать образование волокнистой корневой системы. Питомники используют эту технику для деревьев, которые естественным образом дают стержневой корень, потому что деревья с компактной мочковатой корневой системой пересаживаются более успешно.
Как растут корни
На раннем этапе развития саженец поглощает питательные вещества и влагу из почвы вокруг прорастающих семян. Полоса удобрений на несколько дюймов в каждую сторону и немного ниже посаженных семян способствует раннему росту большинства пропашных культур.
По мере того, как растение становится устойчивым, количество и распределение его корней сильно влияют на его способность поглощать влагу и питательные вещества. У большинства растений большая часть поглощающих (питающих) корней находится в верхних 12-дюймовых слоях почвы.Почвенная среда в этом регионе обычно лучше всего подходит для роста корней, с хорошим балансом плодородия, влаги и воздушного пространства.
Для роста корней важны следующие факторы:
- Корни в водонасыщенной почве плохо растут и в конечном итоге могут погибнуть из-за недостатка кислорода.
- В рыхлой, хорошо дренированной почве корни проникают намного глубже, чем в тяжелой, плохо дренированной.
- Плотный уплотненный слой почвы может ограничить или остановить рост корней.
- Контейнерные растения не только имеют ограниченную зону для роста корней, но также чувствительны к повреждению холодом, потому что ограниченное количество почвы, окружающей их корни, может не обеспечить адекватную изоляцию.
- Помимо роста вниз, корни растут в боковом направлении и часто выходят далеко за пределы капельной линии растения. Помните об этой обширной корневой системе, нарушая почву вокруг существующих деревьев и кустарников.
Корни в пищу
Увеличенный корень — съедобная часть некоторых овощных культур.Сладкий картофель — это набухший клубневидный корень; а морковь, пастернак, сальсиф и редис — удлиненные стержневые корни.
Стебли
Стебли поддерживают почки и листья и служат проводниками для переноса воды, минералов и пищи (фотосинтатов). Сосудистая система внутри стебля образует непрерывный путь от корня через стебель и, наконец, к листьям. Именно через эту систему движутся вода и пищевые продукты.
Терминология стержня
- Побег : Молодой стебель (возрастом менее 1 года) с листьями.
- Веточка : Молодой стебель (возрастом менее 1 года), находящийся в стадии покоя (без листьев).
- Ветвь : Стебель возрастом более 1 года, обычно с отходящими от него боковыми стволами.
- Ствол : Главный стебель древесного растения.
Конструкция
Сосудистая система
Эта система состоит из ксилемы , флоэмы и сосудистого камбия . Его можно рассматривать как водопровод для растений. Трубки Xylem проводят воду и растворенные минералы; Флоэма пробирок содержат пищу, например сахар.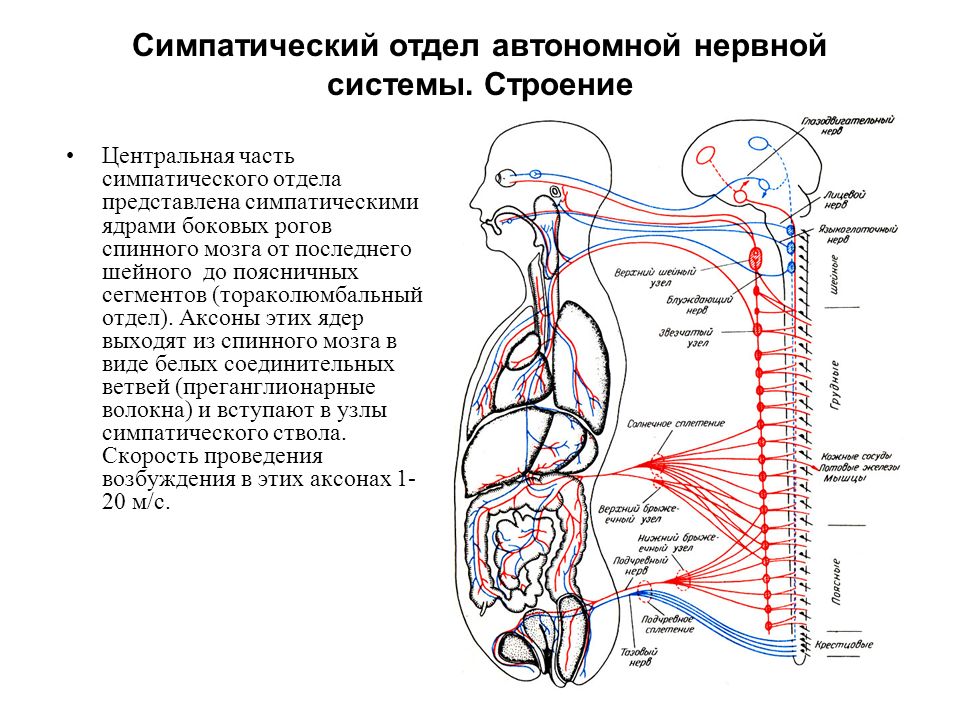 Камбий представляет собой слой меристематической ткани, которая разделяет ксилему и флоэму и непрерывно производит новые клетки ксилемы и флоэмы. Эта новая ткань отвечает за увеличение обхвата ствола.
Камбий представляет собой слой меристематической ткани, которая разделяет ксилему и флоэму и непрерывно производит новые клетки ксилемы и флоэмы. Эта новая ткань отвечает за увеличение обхвата ствола.
Сосудистый камбий важен для садоводов. Например, ткани на привитом привое и подвое должны быть выровнены. Кроме того, неосторожная обрезка сорняков может сорвать кору с дерева, повредить камбий и привести к гибели дерева.
Сосудистые системы однодольных и двудольных различаются (рис. 5). Хотя оба содержат ксилему и флоэму, эти структуры устроены в каждой по-разному. У однодольных ксилема и флоэма объединены в пучки, которые рассредоточены по стеблю. Сосудистая система двудольного растения называется непрерывной, потому что она образует кольца внутри стебля. Кольцо флоэмы находится рядом с корой и со временем становится частью коры зрелых древесных стеблей. Ксилема образует внутреннее кольцо. У древесных растений это называется заболонью и сердцевиной.
Различие сосудистой системы однодольных и двудольных представляет практический интерес для садоводов, поскольку некоторые гербициды действуют только на одну группу. Например, 2,4-Д убивает только растения с непрерывной сосудистой системой (двудольные). С другой стороны, неселективные гербициды (например, глифосат) убивают растения независимо от типа их сосудистой системы.
Узлы
Узел — это участок на стебле, где расположены почки (рис. 6). Это место высокой клеточной активности и роста, где маленькие бутоны развиваются в листья, стебли или цветы.При обрезке важно найти узлы растения. Как правило, вы хотите сделать обрезку чуть выше, но не слишком близко к узлу. Такая обрезка побуждает бутоны на этом узле начать развитие и в конечном итоге сформировать новые стебли или листья.
Область между двумя узлами называется межузлом . Его длина зависит от многих факторов, в том числе от генетики. Несколько других факторов также могут влиять на длину междоузлия:
- Пониженное плодородие почвы уменьшает длину междоузлий, а внесение удобрений с высоким содержанием азота может значительно увеличить ее.

- Недостаток света увеличивает длину междоузлий и делает стержень веретенообразным. Эта ситуация известна как растяжение, или этиоляция , и часто возникает у всходов, выращенных в помещении, и у комнатных растений, которые не получают достаточного количества солнечного света.
- Длина междоузлия также меняется в зависимости от сезона. Рост в начале сезона имеет длинные междоузлия, в то время как рост в конце сезона обычно имеет гораздо более короткие междоузлия.
- Если энергия стебля разделяется между тремя или четырьмя боковыми стеблями или направляется на рост и развитие плодов, длина междоузлий сокращается.
- Вещества, регулирующие рост растений, и гербициды также могут влиять на длину междоузлий.
Типы стержней
Стебли могут быть длинными, с большими расстояниями между листьями и почками (например, ветви деревьев, побеги клубники) или сжатыми, с небольшими расстояниями между почками или листьями (например, кроны клубники, плодовые шпоры и африканские фиалки). . Хотя стебли обычно растут над землей, иногда они растут под землей в виде корневищ, клубней, клубнелуковиц или луковиц.Все стебли должны иметь бутоны или листья, чтобы их можно было классифицировать как ткань стебля.
. Хотя стебли обычно растут над землей, иногда они растут под землей в виде корневищ, клубней, клубнелуковиц или луковиц.Все стебли должны иметь бутоны или листья, чтобы их можно было классифицировать как ткань стебля.
Надземные стебли специализированные
У некоторых растений есть специализированные надземные стебли, известные как кроны , шпоры или столоны (рис. 7). Короны (клубника, одуванчики и африканские фиалки) представляют собой сжатые стебли с листьями и цветами на коротких междоузлиях. Шпоры — короткие, толстые, боковые стержни, которые отходят от основного стержня. Это плодовые стебли груши, яблони и вишни.Если провести сильную обрезку рядом с плодоносящими шпорами, они могут вернуться к неплодоносящим стеблям, тем самым исключив потенциальный урожай плодов в этом году.
Столоны — это мясистые или полудревесные, удлиненные, горизонтальные стебли, которые часто лежат вдоль поверхности почвы. Побеги клубники — это столоны с небольшими листьями в узлах. Из этих узлов развиваются корни, и образуется дочернее растение. Этот тип вегетативного размножения — простой способ увеличить размер клубники.Растения-пауки также производят столоны, которые в конечном итоге могут стать совершенно новыми растениями.
Побеги клубники — это столоны с небольшими листьями в узлах. Из этих узлов развиваются корни, и образуется дочернее растение. Этот тип вегетативного размножения — простой способ увеличить размер клубники.Растения-пауки также производят столоны, которые в конечном итоге могут стать совершенно новыми растениями.
Специальные подземные стволы
Клубни картофеля, корневища ириса и луковицы тюльпанов — это подземные стебли, на которых хранится пища для растения (рис. 8). Иногда бывает трудно отличить корни от стеблей, но один верный способ — поискать узлы. Стебли имеют узлов ; корней нет.
В клубнях картофеля , например, «глаза» на самом деле являются узлами стебля, и каждый глазок содержит скопление почек.При выращивании картофеля из кусков семян важно, чтобы каждый кусочек имел хотя бы один глазок и был размером с мяч для гольфа, чтобы у него было достаточно энергии для раннего роста побегов и корней.
Корневища напоминают столоны, потому что они растут горизонтально от растения к растению. Некоторые корневища сжатые и мясистые (например, радужная оболочка), в то время как другие тонкие и имеют удлиненные междоузлия (например, полевица). Johnsongrass — коварный сорняк, главным образом из-за способности его корневищ к распространению.
Некоторые корневища сжатые и мясистые (например, радужная оболочка), в то время как другие тонкие и имеют удлиненные междоузлия (например, полевица). Johnsongrass — коварный сорняк, главным образом из-за способности его корневищ к распространению.
Тюльпаны, лилии, нарциссы и лук дают луковицу. представляют собой укороченные сжатые подземные стебли, окруженные мясистыми чешуйками (листьями), которые охватывают центральную почку на кончике стебля. В ноябре можно разрезать луковицу тюльпана или нарцисса пополам и увидеть все части цветка в миниатюре.
После цветения луковичного растения его флоэма переносит запасы пищи с листьев на чешуйки луковицы. Когда луковица начинает расти весной, она использует накопленную пищу.По этой причине важно не удалять листья нарциссов, тюльпанов и других растений, производящих луковицы, до тех пор, пока они не пожелтеют и не засохнут. К тому времени они закончили производство корма, который будет использоваться для цветения в следующем году.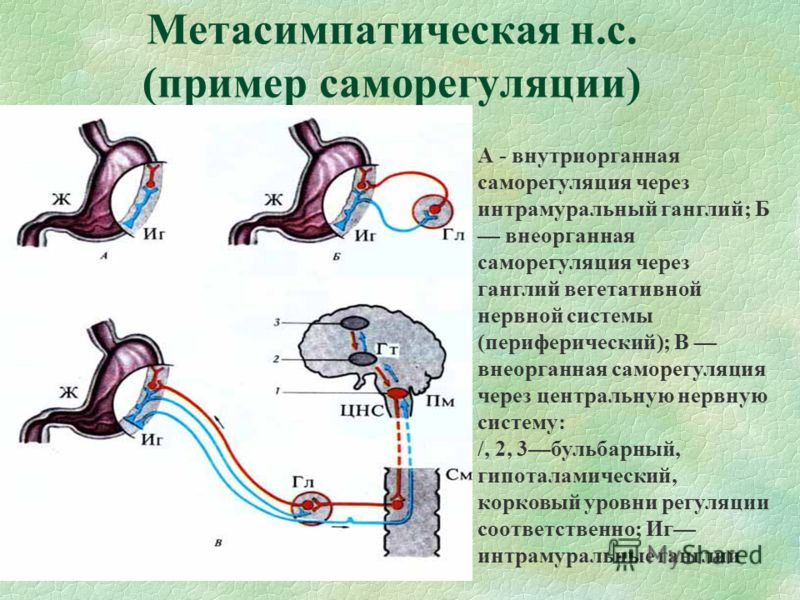
Есть два типа ламп: оболочка и несвязанная (рисунок 8). Tunicate Луковицы (например, нарциссы, тюльпаны и лук) имеют тонкое бумажное покрытие, которое на самом деле представляет собой модифицированный лист.Это помогает защитить луковицу от повреждений во время перекопки и от высыхания после выхода из почвы. Нераздельные луковицы (например, лилии) не имеют бумажного покрытия. Они очень подвержены повреждениям и высыханию, поэтому обращайтесь с ними очень осторожно.
Клубнелуковицы — еще один вид подземных стеблей. Хотя и луковицы, и клубнелуковицы состоят из стеблевой ткани, это не одно и то же. Клубнелуковицы имеют форму луковиц, но не содержат мясистых чешуек. Клубнелуковица — это твердый вздутый стебель с сухими чешуевидными листьями.Гладиолусы и крокусы производят клубнелуковицы.
Некоторые растения (например, клубневые бегонии и цикламены) производят модифицированный подземный стебель, называемый клубневым стеблем . Эти стебли короткие, плоские и увеличенные. Почки и побеги возникают сверху (макушка), а снизу растут мочковатые корни.
Эти стебли короткие, плоские и увеличенные. Почки и побеги возникают сверху (макушка), а снизу растут мочковатые корни.
Другие растения (например, георгины и сладкий картофель) производят подземные запасающие органы, называемые клубневые корни . которые часто путают с луковицами и клубнями. Однако это ткань корня, а не ткань ствола, и у нее нет ни узлов, ни междоузлий.
Стебли и размножение
Стебли часто используются для вегетативного размножения растений. Использование участков надземных стеблей, содержащих узлы и междоузлия, является эффективным способом размножения многих декоративных растений. Эти стеблевые черенки дают корни и, в конечном итоге, новые растения.
Подземные стебли также являются хорошими тканями для размножения. Можно корневища разделить на части; удалите с их родителей мелкие луковицы или кормели; и разрезать клубни на части, содержащие глазки и узлы.Все эти ткани дадут новые растения.
Виды растений и их стебли
Деревья обычно имеют один, но иногда и несколько основных стволов, которые в зрелом возрасте обычно достигают более 12 футов в высоту.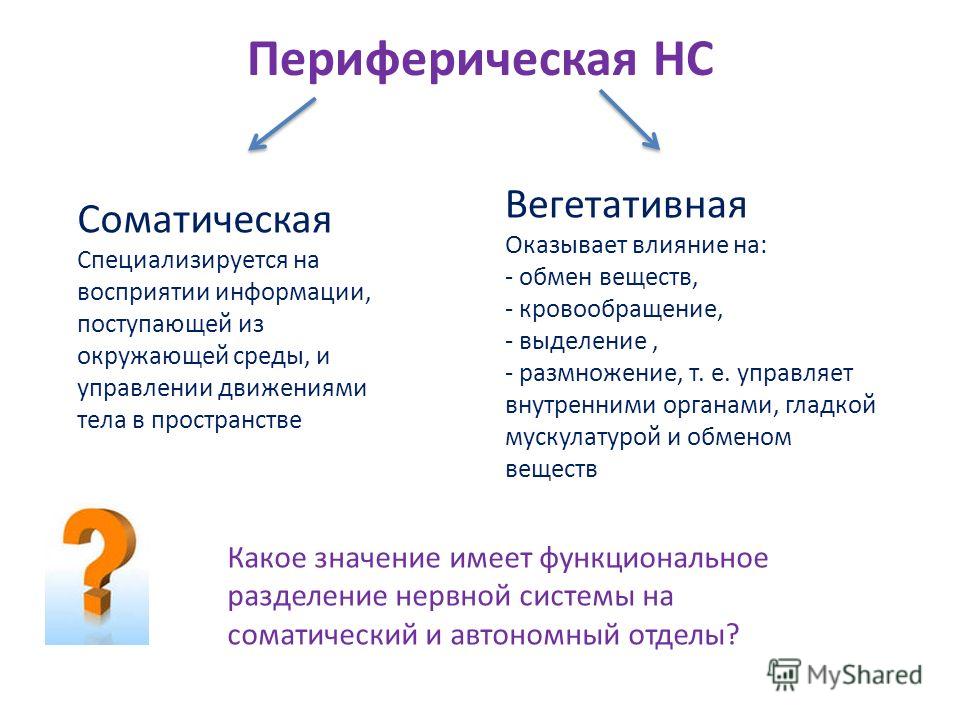 Напротив, кусты обычно имеют несколько основных стеблей, которые в зрелом возрасте обычно достигают менее 12 футов в высоту.
Напротив, кусты обычно имеют несколько основных стеблей, которые в зрелом возрасте обычно достигают менее 12 футов в высоту.
Большинство фруктовых деревьев, декоративных деревьев и кустарников имеют древесные стебли. Эти стебли содержат относительно большое количество затвердевшей ткани ксилемы в центральном ядре (сердцевине или заболони).
Травянистые или сочные стебли содержат лишь немного ткани ксилемы и обычно живут только один вегетационный период. У многолетних растений новые стебли травянистых растений развиваются из кроны (поверхности раздела корень-стебель) каждый год.
Трость (рис. 9a) — это стержень с относительно большой сердцевиной (центральная ткань, придающая силу). Обычно они живут всего 1-2 года. Примеры растений с тростником: розы, виноград, ежевика и малина. Для производства фруктов важно знать, какие стебли обрезать, как обрезать и когда обрезать.
Виноградная лоза (рис. 9b) — это растение с длинными висящими стеблями. Некоторые виноградные лозы растут вдоль земли, в то время как другие должны поддерживаться другим растением или строением. Вьющиеся лозы окружают конструкцию для поддержки. Некоторые кружатся по часовой стрелке (например, хмель и жимолость), а другие — против часовой стрелки (например, бобы и лоза голландца). Вьющиеся лозы поддерживаются либо воздушными корнями (например, английский плющ и ядовитый плющ), либо тонкими усиками, которые окружают поддерживающий объект (например, английский плющ и ядовитый плющ).g., огурцы, тыквы, виноград и пассифлоры) или усиками с липкими кончиками (например, вирджиния и японская лиана).
Некоторые виноградные лозы растут вдоль земли, в то время как другие должны поддерживаться другим растением или строением. Вьющиеся лозы окружают конструкцию для поддержки. Некоторые кружатся по часовой стрелке (например, хмель и жимолость), а другие — против часовой стрелки (например, бобы и лоза голландца). Вьющиеся лозы поддерживаются либо воздушными корнями (например, английский плющ и ядовитый плющ), либо тонкими усиками, которые окружают поддерживающий объект (например, английский плющ и ядовитый плющ).g., огурцы, тыквы, виноград и пассифлоры) или усиками с липкими кончиками (например, вирджиния и японская лиана).
Стебли в пищу
Съедобная часть некоторых культурных растений, таких как спаржа и кольраби, представляет собой увеличенный сочный стебель. Съедобные части брокколи состоят из стеблевой ткани, цветочных почек и нескольких маленьких листочков. Съедобный клубень картофеля — это мясистый подземный стебель. И хотя название говорит об обратном, съедобная часть цветной капусты на самом деле представляет собой разросшуюся стеблевую ткань.
Почки
Бутон — это неразвитый побег, из которого растут листья или части цветка. На почках деревьев и кустарников умеренной зоны обычно образуется защитный внешний слой из мелких кожистых чешуек. Однолетние растения и травянистые многолетники имеют голые бутоны с зелеными, немного сочными внешними листьями.
Почки многих растений требуют выдержки в течение определенного количества дней при температуре ниже критической, прежде чем возобновить рост весной. Этот период, часто называемый покоем, варьируется для разных растений.Например, форзиция требует относительно короткого периода отдыха и растет при первых признаках теплой погоды. С другой стороны, многим сортам персиков требуется от 700 до 1000 часов при температуре ниже 45 ° F. Во время покоя спящие бутоны могут выдерживать очень низкие температуры, но после того, как период покоя насыщен, они более подвержены повреждению холодными температурами или морозами.
Листовая почка состоит из короткого стебля с зародышевыми листьями. Листовые почки часто менее пухлые и более заостренные, чем цветочные почки (рис. 10а).Бутон цветка состоит из короткого стебля с зародышевыми частями цветка. В случае плодовых культур цветочные почки иногда называют плодовыми. Однако эта терминология неточна; Хотя цветы могут развиваться в плоды, это может не произойти из-за неблагоприятных погодных условий, отсутствия опыления или других неблагоприятных обстоятельств.
Листовые почки часто менее пухлые и более заостренные, чем цветочные почки (рис. 10а).Бутон цветка состоит из короткого стебля с зародышевыми частями цветка. В случае плодовых культур цветочные почки иногда называют плодовыми. Однако эта терминология неточна; Хотя цветы могут развиваться в плоды, это может не произойти из-за неблагоприятных погодных условий, отсутствия опыления или других неблагоприятных обстоятельств.
Расположение
Почки названы в честь их расположения на стебле (рис. 10b). Терминальные зачатки расположены на вершине (кончике) стебля.Боковые ( пазушные, ) зачатки расположены по бокам стебля и обычно возникают там, где лист встречается со стеблем (пазуха , ). В некоторых случаях пазуха содержит более одной почки.
Придаточные зачатки возникают не в терминальном или подмышечном положении, а на других участках. Они могут развиваться из корней, стебля , междоузлия , края листовой пластинки или костной ткани на отрезанном конце стебля или корня. Придаточные почки позволяют стеблям, листьям и корневым черенкам развиваться в совершенно новые растения.
Придаточные почки позволяют стеблям, листьям и корневым черенкам развиваться в совершенно новые растения.
Почки в пищу
Увеличенные почки или их части являются съедобной частью некоторых садовых культур. Кочанная капуста и кочанный салат — примеры необычайно больших верхушечных бутонов. Сочные пазушные почки — съедобная часть брюссельской капусты. В случае шарообразного артишока съедается мясистая базальная часть прицветников бутона вместе с его твердым стеблем. Брокколи — важнейшее садовое растение со съедобными цветочными бутонами. В этом случае поедаются части стебля, а также небольшие листья, связанные с цветочными бутонами.
Листья
Функционирование и устройство
Основная функция листьев — поглощать солнечный свет для производства растительных сахаров посредством процесса, называемого фотосинтез . Поверхности листьев плоские, чтобы обеспечить большую площадь для эффективного поглощения света. Лезвие представляет собой расширенную тонкую структуру по обе стороны от средней жилки и обычно является самой большой и заметной частью листа (рис. 11).
11).
Лист удерживается на расстоянии от стебля стеблевидным отростком, называемым черешком , а основание черешка прикреплено к стеблю в узле .Черешки различаются по длине или могут отсутствовать полностью, в этом случае листовая пластинка описывается как сидячая, или бесстебельная.
Узел, где черешок встречается со стеблем, называется пазухой листа . В пазухе находятся отдельные почки или скопления почек, называемые пазушными почками. Они могут быть активными или бездействующими; в правильных условиях они разовьются в стебли или листья.
Листовая пластинка состоит из нескольких слоев (Рисунок 12a и Рисунок 12b). Сверху и внизу находится слой толстых твердых клеток, называемый эпидермисом .Его основная функция — защита других слоев ткани листа. Расположение клеток эпидермиса определяет текстуру поверхности листа. У некоторых листьев, например у африканской фиалки, есть волоски (опушение ), которые являются продолжением эпидермальных клеток, благодаря которым листья кажутся бархатными.
Кутикула является частью эпидермиса. Он образует восковой слой под названием cutin , который защищает лист от обезвоживания и болезней. Количество кутина на листе увеличивается с увеличением интенсивности света.По этой причине, перемещая растения из тени на солнечный свет, делайте это постепенно в течение нескольких недель. Постепенное воздействие солнечного света позволяет нарастать кутиновому слою и защищать листья от быстрой потери воды или солнечного ожога.
Восковый кутин также отталкивает воду. По этой причине многие пестициды содержат аэрозольные добавки, которые помогают продукту прилипать или проникать через слой кутина.
Специальные эпидермальные клетки, называемые замыкающими клетками , открываются и закрываются в ответ на внешние раздражители, такие как изменения погоды и света.Они регулируют проникновение воды, кислорода и углекислого газа в лист и из него через крошечные отверстия, называемые устьицами . У большинства видов большинство устьиц расположено на нижней стороне листьев.
У большинства видов большинство устьиц расположено на нижней стороне листьев.
Условия, при которых растения теряют много воды (высокая температура, низкая влажность), стимулируют закрытие замыкающих клеток. В мягкую погоду они остаются открытыми. Сторожевые клетки также закрываются при отсутствии света.
Между верхним и нижним эпидермисом находится мезофилл .Он разделен на плотный верхний слой (палисадный мезофилл) и нижний слой, содержащий много воздушного пространства (губчатый мезофилл). Внутри клеток мезофилла находятся хлоропластов , в которых происходит фотосинтез.
Виды листьев
Есть много видов листьев растений. Наиболее распространенные и заметные листья называются листвой и являются основным местом фотосинтеза. Однако есть много других видов модифицированных листьев:
- Чешуйчатые листья (катафиллы) находятся на корневищах и почках, которые они закрывают и защищают.

- Семенные листья ( семядоли ) встречаются на зародышевых растениях. В них хранится корм для развивающегося саженца.
- Колючки и усики , такие как те, что встречаются на растениях барбариса и гороха, защищают растение или помогают поддерживать его стебли.
- Хранение листьев , например, на луковичных и суккулентных растениях, для хранения продуктов.
- Прицветники часто ярко окрашены. Например, эффектные структуры на кизилах и пуансеттиях представляют собой прицветники, а не лепестки.
жилкование
Сосудистые пучки ксилемы и флоэмы проходят от стебля через черешок и в виде жилок в пластинку листа. Термин жилкование относится к тому, как расположены жилки в лезвии. Существует два основных типа жилкования: с параллельными жилками и с сетчатыми жилками (рис. 13).
В листе с параллельными прожилками многочисленные прожилки идут по существу параллельно друг другу и соединяются сбоку мельчайшими прямыми прожилками. Листья с параллельными жилками чаще всего встречаются у однодольных растений. Наиболее распространенный тип параллельных жилок встречается у растений семейства злаковых, жилки которых проходят от основания листа к его вершине. Другой тип параллельного жилкования встречается у таких растений, как банан, калла и пикерник, жилки которых идут сбоку от средней жилки.
Листья с параллельными жилками чаще всего встречаются у однодольных растений. Наиболее распространенный тип параллельных жилок встречается у растений семейства злаковых, жилки которых проходят от основания листа к его вершине. Другой тип параллельного жилкования встречается у таких растений, как банан, калла и пикерник, жилки которых идут сбоку от средней жилки.
В листе с сетчатыми прожилками (также называемым с сетчатыми прожилками ) жилки ответвляются от главного ребра или ребер и разделяются на более тонкие прожилки.Эти прожилки затем объединяются в сложную сеть. Эта система переплетенных жилок делает лист более устойчивым к разрыву, чем структура с параллельными жилками. Листья с сетчатыми прожилками встречаются у двудольных растений.
Сетчатое жилкование может быть перистым или пальчатым. У перистого жилкования жилки проходят латерально от средней жилки до края (например, у яблок, вишен и персиков). В жилковании пальчато- основные жилки отходят наружу, как ребра веера, от основания листовой пластинки (e. г., виноград и клены).
г., виноград и клены).
Листья как идентификаторы растений
Листья используются для идентификации растений. Жилкование листа (Рис. 13), форма лезвия и края (Рис. 14 и 15), а также форма вершины и основания (Рис. 16) могут быть важными отличительными характеристиками.
Тип листа (рис. 17) также важен для идентификации. Есть два типа листов: простой и составной . В простых листах листовая пластина представляет собой единый сплошной блок. Сложные листья состоят из нескольких отдельных листочков, растущих на одном черешке. Некоторые листья сложны вдвойне. Тип листа может сбивать с толку, потому что простой лист с глубокими лопастями может выглядеть как сложный.
Расположение листьев вдоль стебля также используется при идентификации растений (рис. 18). Существует четыре типа расположения створок:
- Противоположные листья расположены поперек стебля друг от друга, по два листа на каждом узле.
- Чередующиеся (спиральные) листья расположены попеременно вдоль стебля, только по одному листу в каждом узле.
- Мутовчатые листья расположены по кругу вдоль стебля.
- Рошеобразные листья расположены в виде розетки вокруг стебля с очень короткими узлами.
Листья в пищу
Листовая пластинка является основной съедобной частью некоторых садовых культур, в том числе чеснока, капусты, одуванчиков, эндивий, капусты, листового салата, горчицы, петрушки, шпината, мангольда и другой зелени. Съедобная часть лука-порея, лука и флорентийского фенхеля представляет собой скопление оснований мясистых листьев.Черешок — это съедобный продукт сельдерея и ревеня.
Устойчивое вегетативное состояние — AMBOSS
Последнее обновление: 7 марта 2019 г.
Резюме
Устойчивое вегетативное состояние (ПВС) — это клиническое состояние, при котором функция коры головного мозга нарушается, а функция ствола мозга сохраняется в течение более длительного периода. чем один месяц. Черепно-мозговая травма и диффузная церебральная гипоксия — наиболее частая этиология. Пациенты в ПВС могут казаться бодрствующими, но не осознают и не осознают, и они не могут общаться с другими или целенаправленно взаимодействовать со своим окружением.Однако они по-прежнему могут дышать самостоятельно, циклы сна и бодрствования сохраняются, а вегетативная функция сохраняется, по крайней мере, частично. ПВС — это клинический диагноз, и его важно отличать от комы (отсутствие циклов сна-бодрствования) и смерти мозга (отсутствие циклов сна-бодрствования или функции ствола мозга). Окончательного лечения не существует. Хотя большинство пациентов остаются в вегетативном состоянии в течение многих лет, некоторые могут выздоравливать самопроизвольно. Вегетативное состояние объявляется постоянным, когда восстановление маловероятно (т.например, ПВС вследствие травмы продолжительностью более 1 года или ПВС вследствие нетравматической причины продолжительностью более 3 месяцев).
чем один месяц. Черепно-мозговая травма и диффузная церебральная гипоксия — наиболее частая этиология. Пациенты в ПВС могут казаться бодрствующими, но не осознают и не осознают, и они не могут общаться с другими или целенаправленно взаимодействовать со своим окружением.Однако они по-прежнему могут дышать самостоятельно, циклы сна и бодрствования сохраняются, а вегетативная функция сохраняется, по крайней мере, частично. ПВС — это клинический диагноз, и его важно отличать от комы (отсутствие циклов сна-бодрствования) и смерти мозга (отсутствие циклов сна-бодрствования или функции ствола мозга). Окончательного лечения не существует. Хотя большинство пациентов остаются в вегетативном состоянии в течение многих лет, некоторые могут выздоравливать самопроизвольно. Вегетативное состояние объявляется постоянным, когда восстановление маловероятно (т.например, ПВС вследствие травмы продолжительностью более 1 года или ПВС вследствие нетравматической причины продолжительностью более 3 месяцев).
Этиология
Ссылки: [1]
Патофизиология
Широко распространенное поражение белого вещества обоих полушарий головного мозга и / или таламуса с двух сторон без повреждения ствола мозга. Это приводит к снижению корковой функции с сохранением контроля ствола мозга над дыханием, сердечной функцией, циклами сна и бодрствования и рефлексами.
Это приводит к снижению корковой функции с сохранением контроля ствола мозга над дыханием, сердечной функцией, циклами сна и бодрствования и рефлексами.
Каталожные номера: [2] [3] [4]
Клинические особенности
- Обычно предшествует коматозное состояние, требующее интенсивной терапии и постепенного восстановления вегетативной функции.
- Но отсутствие восстановления высших психических функций через 1 месяц и следующие особенности:
- Неповрежденные циклы сна-бодрствования с периодами бодрствования
- Потеря восприятия и самосознания
- Отсутствие целенаправленной реакции на внешние раздражители (напр.д., тактильные, слуховые, визуальные и т. д.)
- Неспособность общаться и очевидное понимание языка
- Расстройство движений глаз с потерей фиксации
- Случайные нецелевые движения конечностями и спонтанная улыбка, плач, хрюканье или крик
- Вегетативная функция и рефлексы сохранены в различной степени
Литература: [5]
Диагностика
- Клинический диагноз, основанный на сохранении циклов сна-бодрствования ≥ 1 месяца, потере восприятия и самосознания, а также сохранении вегетативных функций в различной степени
- Нейровизуализация
- КТ / МРТ: неспецифические диффузные или мультифокальные аномалии серого и белого вещества; снижение мозгового кровотока
- ПЭТ: диффузное снижение скорости метаболизма глюкозы в головном мозге
- ЭЭГ: диффузная генерализованная полиморфная дельта- и / или тета-активность
Дифференциальный диагноз
| Расстройства сознания | ||||
|---|---|---|---|---|
| Стойкое вегетативное состояние | Кома | Синдром запертого состояния | Смерть мозга | |
| Восприятие 909 Отсутствует | Присутствует | Отсутствует | ||
| Циклы сна-бодрствования | Присутствует | Отсутствует | Присутствует | Отсутствует |
| Моторная функция | Нет целенаправленного движения | 920 Нет целенаправленного движения и движения движение глаз сохранено | Отсутствуют или только рефлекторные движения позвоночника | |
| Рефлексы ствола головного мозга | Переменные | Переменные | Переменные | Отсутствуют |
| Функция дыхания | 909 909 | |||
Каталожные номера: [5] [6] [7]
Перечисленные здесь дифференциальные диагнозы не являются исчерпывающими.
Прогноз
- Вегетативное состояние определяется как постоянное, если восстановление маловероятно.
- Вероятность выздоровления зависит от этиологии
- ПВС из-за травмы → восстановление маловероятно через ≥ 12 месяцев
- ПВС нетравматической этиологии → восстановление маловероятно через ≥ 3 месяцев
- Низкая продолжительность жизни
- Прекращение жизнеобеспечения может быть рассмотрено, если шансы на выздоровление очень низкие (например, ≥ 12 месяцев при травматической ПВС).Однако для пациентов без дополнительных указаний это сложный юридический и этический вопрос, который требует обсуждения с членами семьи и экспертами.
Каталожные номера: [1] [2]
Лечение
- Лечение поддерживающее; окончательного лечения не существует.
Ссылки
- Caplan LR.
 Синдром запертости. В: Сообщение TW, под ред. Новости . Уолтем, Массачусетс: UpToDate. https://www.uptodate.com/contents/locked-in-syndrome .Последнее обновление: 25 августа 2014 г. Дата обращения: 3 апреля 2017 г.
Синдром запертости. В: Сообщение TW, под ред. Новости . Уолтем, Массачусетс: UpToDate. https://www.uptodate.com/contents/locked-in-syndrome .Последнее обновление: 25 августа 2014 г. Дата обращения: 3 апреля 2017 г. - Ашвал С., Крэнфорд Р., Бернат Дж. Л. и др. Медицинские аспекты стойкого вегетативного состояния. N Engl J Med . 1994; 330 (21): с.1499-1508. DOI: 10,1056 / nejm199405263302107. | Открыть в режиме чтения QxMD
- Земан А. Стойкое вегетативное состояние. Ланцет . 1998; 351 (9096): с.144. DOI: 10,1016 / s0140-6736 (05) 78109-5. | Открыть в режиме чтения QxMD
- Вегетативное состояние и состояние минимального сознания. https://www.msdmanuals.com/professional/neurologic-disorders/coma-and-impaired-consciousness/vegetative-state-and-minimally-conscious-state .
Обновлено: 1 декабря 2017 г.
Доступ: 28 ноября 2018 г.

- Адамс Дж. Х. Невропатология вегетативного состояния после острого инсульта головного мозга. Мозг . 2000; 123 (7): с.1327-1338. DOI: 10.1093 / мозг / 123.7.1327. | Открыть в режиме чтения QxMD
- Патофизиология вегетативного состояния. https://www.jwatch.org/jn201003300000003/2010/03/30/pathophysiology-vegetative-state . Обновлено: 30 марта 2010 г. Доступ: 28 ноября 2018 г.
- Wijdicks EFM, Cranford RE. Клиническая диагностика затяжных состояний нарушения сознания у взрослых. Mayo Clin Proc . 2005; 80 (8): с.1037-1046. DOI: 10.4065 / 80.8.1037. | Открыть в режиме чтения QxMD
Бесплодное соцветие1 Функции в органогенезе во время вегетативного развития и развития соцветий кукурузы
Abstract
Кукуруза ( Zea mays ) имеет сильно разветвленное соцветие из-за образования различных типов пазушных меристем. Характеристика бесплодного соцветия класса мутантов привела к открытию генов, необходимых для инициации пазушной меристемы в соцветии. Предыдущие исследования показали, что бесплодное соцветие2 ( bif2) кодирует серин / треониновую протеинкиназу, которая регулирует транспорт ауксина, а бесплодный стебель1 ( ba1 ) кодирует основной транскрипционный фактор спираль-петля-спираль, который действует ниже ауксина. транспорт. Здесь мы охарактеризуем Бесплодное соцветие1 ( Bif1 ), классическую полудоминантную мутацию кукурузы.Онтогенетический, гистологический и генетический анализы показывают, что мутанты Bif1 дефектны в инициации всех пазушных меристем в соцветии. Эксперименты с ОТ-ПЦР в реальном времени показывают, что и bif2 , и ba1 экспрессируются на более низких уровнях у мутантов Bif1 . Анализ двойных мутантов показывает, что Bif1 проявляет эпистатическое взаимодействие с ba1 и синергетическое взаимодействие с bif2 .
Характеристика бесплодного соцветия класса мутантов привела к открытию генов, необходимых для инициации пазушной меристемы в соцветии. Предыдущие исследования показали, что бесплодное соцветие2 ( bif2) кодирует серин / треониновую протеинкиназу, которая регулирует транспорт ауксина, а бесплодный стебель1 ( ba1 ) кодирует основной транскрипционный фактор спираль-петля-спираль, который действует ниже ауксина. транспорт. Здесь мы охарактеризуем Бесплодное соцветие1 ( Bif1 ), классическую полудоминантную мутацию кукурузы.Онтогенетический, гистологический и генетический анализы показывают, что мутанты Bif1 дефектны в инициации всех пазушных меристем в соцветии. Эксперименты с ОТ-ПЦР в реальном времени показывают, что и bif2 , и ba1 экспрессируются на более низких уровнях у мутантов Bif1 . Анализ двойных мутантов показывает, что Bif1 проявляет эпистатическое взаимодействие с ba1 и синергетическое взаимодействие с bif2 . Резкое фенотипическое усиление наблюдается у Bif1; bif2 двойные мутанты подразумевают, что bif1 играет перекрывающуюся роль с bif2 в инициации боковых органов во время вегетативного развития.Фенотипическое сходство мутантов Bif1 с bif2 и снижение транспорта ауксина у мутантов Bif1 предполагает, что bif1 функционирует как регулятор транспорта ауксина в кукурузе.
Резкое фенотипическое усиление наблюдается у Bif1; bif2 двойные мутанты подразумевают, что bif1 играет перекрывающуюся роль с bif2 в инициации боковых органов во время вегетативного развития.Фенотипическое сходство мутантов Bif1 с bif2 и снижение транспорта ауксина у мутантов Bif1 предполагает, что bif1 функционирует как регулятор транспорта ауксина в кукурузе.
ОРГАНОГЕНЕЗ в растениях контролируется меристемами (Steeves and Sussex 1989). Периферическая зона меристемы инициирует зачатки органов, в то время как центральная зона остается недифференцированной, чтобы позволить органогенезу продолжаться бесконечно (McSteen and Hake 1998; Veit 2006).Ауксин играет фундаментальную роль в органогенезе периферической зоны меристемы (Reinhardt et al. 2000, 2003; Vernoux et al. 2000). Растения с мутациями в генах, необходимых для биосинтеза, транспорта или ответа ауксина, имеют дефекты органогенеза (Okada et al. 1991; Bennett et al. 1995; Przemeck et al. 1996; Vernoux et al. 2000 ; Cheng et al. 2006, 2007). Эти и другие исследования показали, что ауксин необходим для зарождения листьев во время вегетативного развития и зарождения цветков во время репродуктивного развития (Okada et al. 1991; Рейнхардт и др. 2000, 2003; Бенкова и др. 2003; Scanlon 2003; Heisler et al. 2005; Ченг и Чжао 2007; Ву и Макстин 2007).
1991; Bennett et al. 1995; Przemeck et al. 1996; Vernoux et al. 2000 ; Cheng et al. 2006, 2007). Эти и другие исследования показали, что ауксин необходим для зарождения листьев во время вегетативного развития и зарождения цветков во время репродуктивного развития (Okada et al. 1991; Рейнхардт и др. 2000, 2003; Бенкова и др. 2003; Scanlon 2003; Heisler et al. 2005; Ченг и Чжао 2007; Ву и Макстин 2007).
Во время вегетативной фазы роста апикальная меристема побега (SAM) на кончике развивающегося побега многократно продуцирует фитомеры, состоящие из узла, междоузлия, листа и пазушной меристемы, расположенной в пазухе листа (Steeves and Sussex 1989 ; Макстин и Лейзер 2005). Подмышечные меристемы могут вырасти и превратиться в боковую ветвь, известную у кукурузы как побеги, которая повторяет рост главного побега.Во время репродуктивной фазы роста апикальная меристема побега превращается в меристему соцветия, производящую модифицированные фитомеры. У многих растений листья редуцируются, образуя прицветники, а пазушные меристемы увеличиваются для образования цветков (Steeves and Sussex 1989). У кукурузы образуются сильно разветвленные соцветия (McSteen et al. 2000; Bommert et al. 2005; Bortiri and Hake 2007). Мужское соцветие, кисточка, растет на верхушке растения и состоит из главного колоса с несколькими длинными ветвями у основания (рис. 1А).Главный колос и ветви дают короткие ветви, называемые парами колосков. Колоск — строительный блок всех соцветий травы (Clifford 1987; Kellogg 2000). У кукурузы колоск состоит из двух листовидных чешуек, охватывающих два соцветия. Женское соцветие — колос — образуется из пазушной меристемы, расположенной на несколько узлов ниже кисточки. И в кисточке, и в початке соцветия окружены листовыми структурами, называемыми lemma и palea, окружающими пару лепестковидных структур, называемых lodicules, и репродуктивные органы, тычинки и плодолистики.Плодолистики обрываются кисточкой, а тычинки обрываются в ухе, образуя отдельные мужские и женские соцветия (Irish 1996).
У многих растений листья редуцируются, образуя прицветники, а пазушные меристемы увеличиваются для образования цветков (Steeves and Sussex 1989). У кукурузы образуются сильно разветвленные соцветия (McSteen et al. 2000; Bommert et al. 2005; Bortiri and Hake 2007). Мужское соцветие, кисточка, растет на верхушке растения и состоит из главного колоса с несколькими длинными ветвями у основания (рис. 1А).Главный колос и ветви дают короткие ветви, называемые парами колосков. Колоск — строительный блок всех соцветий травы (Clifford 1987; Kellogg 2000). У кукурузы колоск состоит из двух листовидных чешуек, охватывающих два соцветия. Женское соцветие — колос — образуется из пазушной меристемы, расположенной на несколько узлов ниже кисточки. И в кисточке, и в початке соцветия окружены листовыми структурами, называемыми lemma и palea, окружающими пару лепестковидных структур, называемых lodicules, и репродуктивные органы, тычинки и плодолистики.Плодолистики обрываются кисточкой, а тычинки обрываются в ухе, образуя отдельные мужские и женские соцветия (Irish 1996).
Для образования этого сильно разветвленного соцветия меристема соцветия производит четыре типа пазушных меристем (таблица 1), которые дают начало различным структурам зрелого соцветия (Cheng et al. 1983; Irish 1997; McSteen et al. 2000; Bommert et al. 2005). Первичные подмышечные меристемы имеют две альтернативные судьбы.Первые первичные подмышечные меристемы, которые возникают, — это меристемы ветвей (BM). КМ индетерминантные и разрастаются, превращаясь в длинные ветви у основания кисточки, что повторяет рост главного колоса (Таблица 1). Следующие первичные подмышечные меристемы, которые возникают, — это меристемы пары колосков (SPM). ВП детерминантны и дают короткие ветви с парой колосков (табл. 1). SPM продуцируют вторичные подмышечные меристемы, называемые меристемами колосков (SM), которые затем производят третичные подмышечные меристемы, называемые цветочными меристемами (FM), которые в конечном итоге производят цветочные органы.Судьба первичных подмышечных меристем как неопределенных (ветвь) по сравнению с определенными (пара колосков) регулируется путем ramosa ( ra ) (Vollbrecht et al.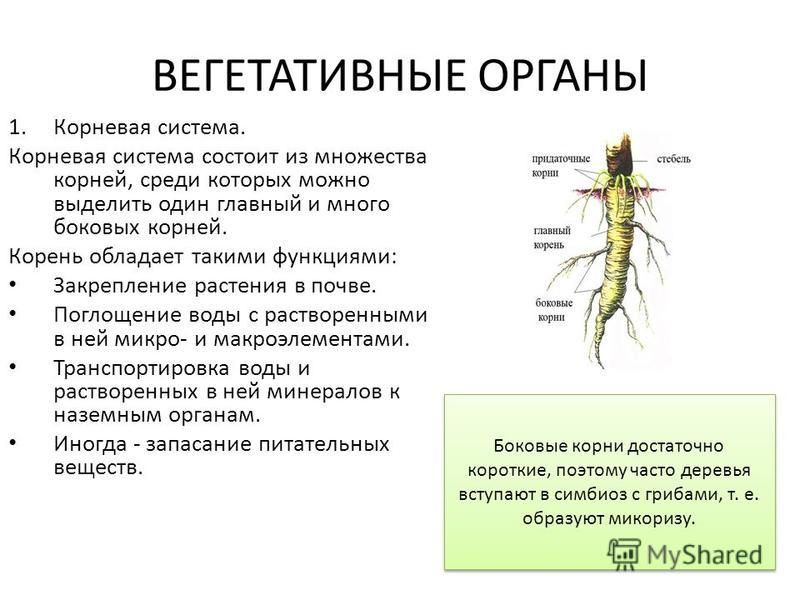 2005; Bortiri et al. 2006a) ; McSteen 2006; Satoh-Nagasawa et al. 2006; Kellogg 2007). Гены ra1 и ra2 кодируют факторы транскрипции, которые необходимы для определения детерминированности SPM (Vollbrecht et al., 2005; Bortiri et al. 2006a). У мутанта ra1 имеются дополнительные длинные ответвления на кисточке и ухе (Gernart 1912; Vollbrecht et al. 2005).
2005; Bortiri et al. 2006a) ; McSteen 2006; Satoh-Nagasawa et al. 2006; Kellogg 2007). Гены ra1 и ra2 кодируют факторы транскрипции, которые необходимы для определения детерминированности SPM (Vollbrecht et al., 2005; Bortiri et al. 2006a). У мутанта ra1 имеются дополнительные длинные ответвления на кисточке и ухе (Gernart 1912; Vollbrecht et al. 2005).
Пазушные меристемы в соцветии кукурузы
В бесплодных соцветиях Локусы кукурузы идентифицируют гены, необходимые для инициации пазушных меристем. бесплодный стебель1 ( ba1 ) кодирует основной транскрипционный фактор спираль-петля-спираль, необходимый для инициации подмышечной меристемы как во время вегетативного, так и во время репродуктивного развития (Gallavotti et al. 2004). ba1 Мутанты не дают побегов, колосьев, ветвей, колосков и цветков (Ritter et al. 2002). бесплодное соцветие2 Мутанты ( bif2 ) также имеют меньше колосьев и меньше ветвей, колосков, цветков и органов цветка из-за дефектов инициации пазушных меристем в соцветии (McSteen and Hake 2001). Мутанты bif2 также имеют дефекты инициации подмышечной меристемы во время вегетативного развития (McSteen et al. 2007). Ген bif2 кодирует серин / треониновую протеинкиназу, коортологичную PINOID , которая регулирует транспорт ауксина у Arabidopsis (Christensen et al. 2000; Benjamins et al. 2001; Lee and Cho 2006; McSteen и др. 2007).
2002). бесплодное соцветие2 Мутанты ( bif2 ) также имеют меньше колосьев и меньше ветвей, колосков, цветков и органов цветка из-за дефектов инициации пазушных меристем в соцветии (McSteen and Hake 2001). Мутанты bif2 также имеют дефекты инициации подмышечной меристемы во время вегетативного развития (McSteen et al. 2007). Ген bif2 кодирует серин / треониновую протеинкиназу, коортологичную PINOID , которая регулирует транспорт ауксина у Arabidopsis (Christensen et al. 2000; Benjamins et al. 2001; Lee and Cho 2006; McSteen и др. 2007).
Здесь мы охарактеризуем еще одно бесплодное соцветие , мутация , бесплодное соцветие1 ( Bif1 ). Bif1 — полудоминантная мутация, которая придает фенотип меньшему количеству ветвей, колосков, цветков и органов цветка в соцветии.Хотя Bif1 является классической мутацией кукурузы, впервые выделенной> 30 лет назад (Neuffer et al.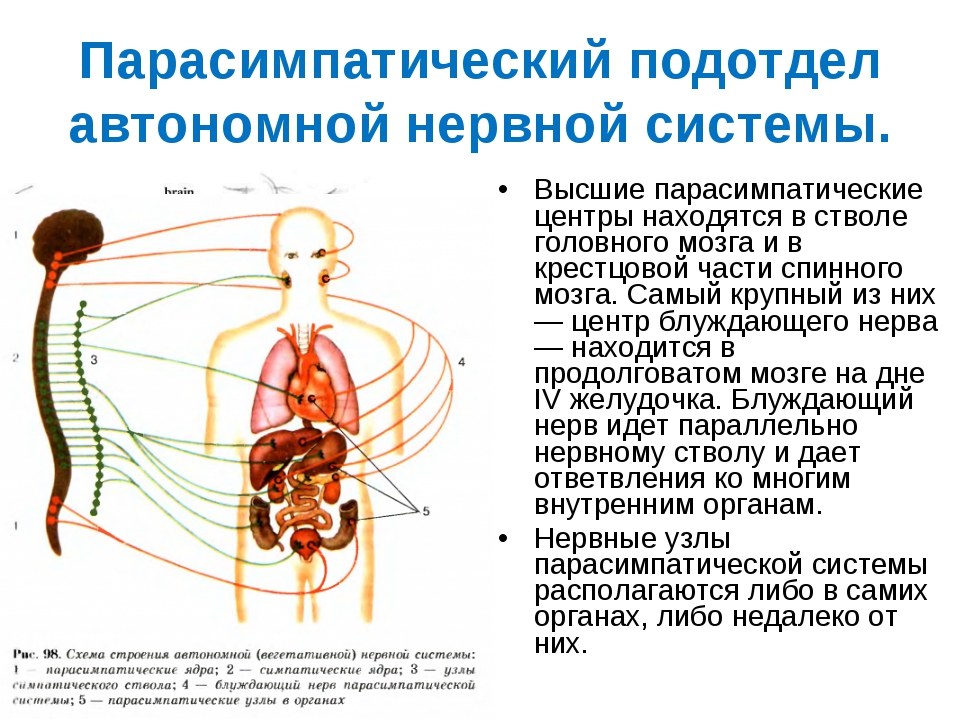 1997), фенотип ранее подробно не анализировался. Здесь мы сообщаем, что дефекты у мутантов Bif1 обусловлены дефектами инициации пазушных меристем в соцветии. Мы протестировали взаимодействие между Bif1 и bif2 или ba1 , используя экспрессию и анализ двойных мутантов. Мы показываем, что Bif1 эпистатичен по отношению к ba1 и что экспрессия ba1 значительно снижена у мутантов Bif1 .Мы показываем, что мутанты Bif1 имеют много фенотипических сходств с мутантами bif2 и что экспрессия bif2 также снижена у мутантов Bif1 . Резкое усиление фенотипа, наблюдаемое в Bif1; Растения с двойным мутантом bif2 указывают на то, что bif1 играет избыточную роль с bif2 в зарождении листьев во время вегетативного развития. Мутанты Bif1 имеют пониженные уровни транспорта ауксина, что означает, что функция bif1 заключается в регуляции транспорта ауксина.
1997), фенотип ранее подробно не анализировался. Здесь мы сообщаем, что дефекты у мутантов Bif1 обусловлены дефектами инициации пазушных меристем в соцветии. Мы протестировали взаимодействие между Bif1 и bif2 или ba1 , используя экспрессию и анализ двойных мутантов. Мы показываем, что Bif1 эпистатичен по отношению к ba1 и что экспрессия ba1 значительно снижена у мутантов Bif1 .Мы показываем, что мутанты Bif1 имеют много фенотипических сходств с мутантами bif2 и что экспрессия bif2 также снижена у мутантов Bif1 . Резкое усиление фенотипа, наблюдаемое в Bif1; Растения с двойным мутантом bif2 указывают на то, что bif1 играет избыточную роль с bif2 в зарождении листьев во время вегетативного развития. Мутанты Bif1 имеют пониженные уровни транспорта ауксина, что означает, что функция bif1 заключается в регуляции транспорта ауксина.
МАТЕРИАЛЫ И МЕТОДЫ
Анализ фенотипа зрелого соцветия
Bif1 :Аллель Bif1-1440 был получен из Центра запасов кукурузы Genetics Coop (инвентарный номер 827C) и восемь раз скрещен с генетическим материалом B73 фон. Количественный анализ был проведен на растениях, выращенных до зрелости (9–10 недель) летом в поле в Рок-Спрингс, штат Пенсильвания. Приведены репрезентативные данные за один полевой сезон. Для анализа количества ветвей и колосков анализировали по 8–10 растений каждого генетического класса.Для определения количества цветков и органов цветка было проанализировано 100 колосков каждого генетического класса.
Анализы двойных мутантов:
Все исходные мутанты были подвергнуты обратному скрещиванию минимум пять раз на B73 перед использованием для создания двойных мутантов с Bif1 . Все семьи с двойными мутантами выращивались летом в поле в Рок-Спрингс, штат Пенсильвания. Чтобы уменьшить воздействие на окружающую среду, все семьи были посажены дважды в разных местах поля и в течение двух отдельных полевых сезонов.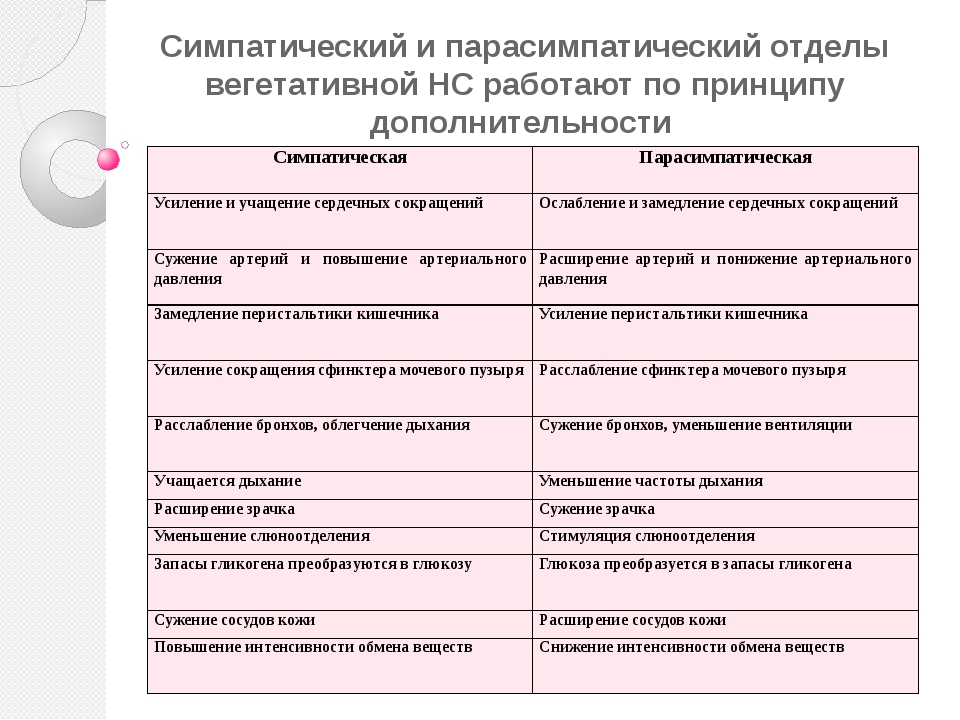 В каждом месте было высажено от двух до трех семейств F 2 по 120 ядер. Представленные здесь данные представляют собой репрезентативный набор данных, собранных в течение полевого сезона 2007 года. Анализ хи-квадрат не смог отклонить нулевую гипотезу об ожидаемом количестве растений в каждом генотипическом классе (дополнительная таблица S1).
В каждом месте было высажено от двух до трех семейств F 2 по 120 ядер. Представленные здесь данные представляют собой репрезентативный набор данных, собранных в течение полевого сезона 2007 года. Анализ хи-квадрат не смог отклонить нулевую гипотезу об ожидаемом количестве растений в каждом генотипическом классе (дополнительная таблица S1).
Bif1; ra1:
Аллель ra1-R был использован для генерации Bif1; ra1 семьи, разделяющие двойные мутанты (Vollbrecht et al. 2005).При погашении Bif1; Двойные мутанты ra1 оценивали по фенотипу кисточки и уха. Была проанализирована архитектура соцветий не менее 10 растений каждого генетического класса. Подсчитывали количество первичных и вторичных ветвей кисточки и общее количество колосков. Также подсчитывали количество колосков на ветке у основания кисточки, а также количество колосков в верхних двух сантиметрах главного колоса.
Bif1; bif2:
Семьи, разделяющие Bif1; bif2 двойные мутанты были созданы с использованием аллеля bif2-77 (McSteen et al. 2007). Для генотипирования ткань листа собирали с двухнедельных растений в 96-луночные планшеты и измельчали с использованием Tissue Lyzer (QIAGEN, Valencia, CA). ДНК экстрагировали в соответствии с протоколом, модифицированным для формата 96-луночного планшета от Chen and Dellaporta (1994), без стадии экстракции фенолом хлороформом. ПЦР проводили для генотипирования растений на мутацию bif2-77 с использованием праймеров bif2-57 (5 ‘CAG CCT GCC GCG CTG CTC CAGC 3’) и bif2-250 (5 ‘CGG CGC AGC AGC CTG AAG TCC 3 ‘), Которые предназначены для пересечения сайта встраивания в этот аллель bif2 (McSteen et al. 2007). Второй набор реакций ПЦР с использованием праймера bif2-57 с праймером, расположенным во вставке (bif2-77, 5 ‘CAG TGG CGG GCC TAG AAA TTT G 3’), использовали для подтверждения этого результата. Bif1 / Bif1; bif2 / bif2 растения были легко идентифицированы как растения, генотипированные как гомозиготные по мутации bif2 , но которые также имели чрезвычайно низкий рост и тяжелый фенотип метелки.
2007). Для генотипирования ткань листа собирали с двухнедельных растений в 96-луночные планшеты и измельчали с использованием Tissue Lyzer (QIAGEN, Valencia, CA). ДНК экстрагировали в соответствии с протоколом, модифицированным для формата 96-луночного планшета от Chen and Dellaporta (1994), без стадии экстракции фенолом хлороформом. ПЦР проводили для генотипирования растений на мутацию bif2-77 с использованием праймеров bif2-57 (5 ‘CAG CCT GCC GCG CTG CTC CAGC 3’) и bif2-250 (5 ‘CGG CGC AGC AGC CTG AAG TCC 3 ‘), Которые предназначены для пересечения сайта встраивания в этот аллель bif2 (McSteen et al. 2007). Второй набор реакций ПЦР с использованием праймера bif2-57 с праймером, расположенным во вставке (bif2-77, 5 ‘CAG TGG CGG GCC TAG AAA TTT G 3’), использовали для подтверждения этого результата. Bif1 / Bif1; bif2 / bif2 растения были легко идентифицированы как растения, генотипированные как гомозиготные по мутации bif2 , но которые также имели чрезвычайно низкий рост и тяжелый фенотип метелки. Первоначальный фенотипический анализ выявил избыток растений с гомозиготным фенотипом Bif1 / Bif1 .Это превышение было отнесено к Bif1 / + ; bif2 / Двойные мутанты bif2 , которые напоминали тяжелые гомозиготы Bif1 по фенотипу, но были генотипированы как гомозиготные по мутации bif2 . Дальнейшее подтверждение этого результата было получено при скрещивании Bif1 / +; bif2 / + растений до + / +; bif2 / + и определение того, что одна восьмая часть потомства напоминала гомозигот Bif1 .
Первоначальный фенотипический анализ выявил избыток растений с гомозиготным фенотипом Bif1 / Bif1 .Это превышение было отнесено к Bif1 / + ; bif2 / Двойные мутанты bif2 , которые напоминали тяжелые гомозиготы Bif1 по фенотипу, но были генотипированы как гомозиготные по мутации bif2 . Дальнейшее подтверждение этого результата было получено при скрещивании Bif1 / +; bif2 / + растений до + / +; bif2 / + и определение того, что одна восьмая часть потомства напоминала гомозигот Bif1 .
В зрелом возрасте измеряли высоту каждого растения от земли до кончика кисточки.Для подсчета количества листьев каждый пятый лист каждого растения обрезали ножницами для розжига, начиная через 3 недели после появления всходов и через равные промежутки времени в течение всего полевого сезона. Это позволило нам получить точную оценку общего количества листьев в конце полевого сезона, потому что, если бы мы считали только в конце полевого сезона, мы бы пропустили опущенные листья. Для анализа количества ветвей метелки и колосков использовали по десять растений каждого генетического класса.
Для анализа количества ветвей метелки и колосков использовали по десять растений каждого генетического класса.
Bif1; ba1:
Аллель ba1-ref был использован для генерации Bif1; ba1 семьи, разделяющие двойные мутанты (Gallavotti et al. 2004). Ткань собирали и ДНК экстрагировали, как описано для Bif1; bif2 растений. Для генотипирования растений использовали праймер ba04 (5 ‘TGG CAT TGC ATG GAA GCG TGT ATG AGC 3’), расположенный в промоторе ba1 , и праймер ba05 (5 ‘TCC TAG ACA TGC ATA TCT GAA CCA GAG CT 3’), расположенный в helitron в аллеле ba1-ref , который амплифицировал продукт в гетерозиготных и гомозиготных растениях ba1 . Вторая реакция ПЦР с праймерами ba04 и ba07 (5 ‘GCT AAG CTA CTG TAA GCG GGA TGG ACA 3’) амплифицировала продукт в растениях дикого типа и гетерозиготных растениях. Bif1 / Bif1; ba1 / ba1 двойные мутанты были классифицированы как растения, генотипированные как гомозиготные по ba1 , но с гладкой тонкой кисточкой, сходной с гомозиготами Bif1 . Bif1 / +; ba1 / ba1 двойные мутанты были классифицированы как растения, генотипированные как гомозиготные по ba1 , которые выглядели как ba1 , но с немного более гладкой кисточкой.
Bif1 / +; ba1 / ba1 двойные мутанты были классифицированы как растения, генотипированные как гомозиготные по ba1 , которые выглядели как ba1 , но с немного более гладкой кисточкой.
Статистический анализ:
Компьютерная программа Minitab v.15 (Minitab, State College, PA) использовался для выполнения всего статистического анализа. Наборы данных сравнивались с двухвыборочными двусторонними тестами t . Данные, представленные в виде гистограмм, представляют собой среднее значение данных, а все полосы ошибок показывают стандартную ошибку среднего.
Сканирующая электронная микроскопия, РНК
in situ, гибридизация и гистология: Кисточки были получены из семей, разделяющих Bif1 , выращенных в теплице в течение 5 недель. Кисти вскрывали и фиксировали на льду в течение ночи в 3 раза.7% формалина, 50% этанола, 10% уксусной кислоты (FAA), а затем дегидратация с помощью серии этанола. Уши были получены от растений Bif1 , выращенных в поле в течение 8 недель.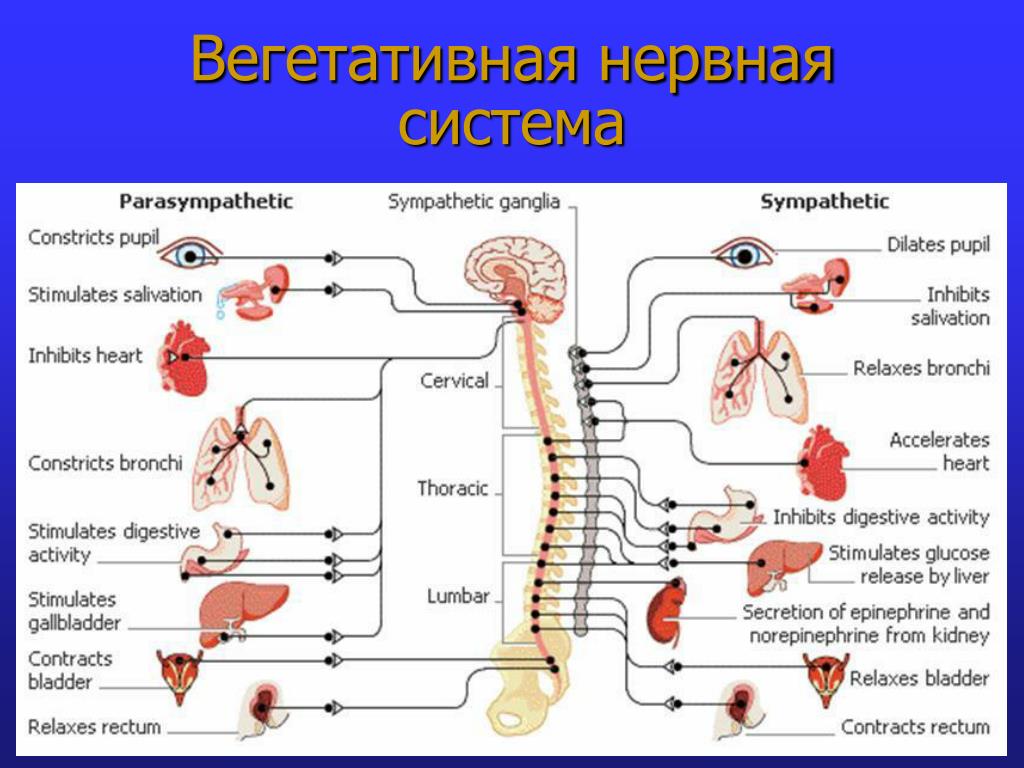 Уши препарировали и фиксировали на льду в течение ночи в 4% формальдегиде в фосфатно-солевом буфере. Для сканирующей электронной микроскопии (SEM) меристемы сушили до критической точки (BAL-TEC CPD 030; Techno Trade, Manchester, NH), а затем устанавливали на углеродные штыри. Образцы покрывали напылением слоем золотого палладия 0,7 Å (BAL-TEC SCD 050, Techno Trade) и просматривали с помощью SEM (JSM 5400; JEOL, Peabody, MA) с использованием ускоряющего напряжения 10 кВ.Для разделения образцы заливали парафином (Paraplast Plus; McCormick Scientific, Сент-Луис). Срезы толщиной 8 мкм вырезали с использованием парафинового микротома Finnesse (Thermo Fisher, Waltham, MA) и помещали на предметные стекла с покрытием (Probe-On Plus; Fisher Scientific, Waltham, MA). Для гибридизации RNA in situ с предметные стекла зондировали DIG-меченным РНК-антисмысловым зондом kn1 согласно Jackson et al. (1994). Для гистологии слайды депарафинизировали с использованием histoclear (National Diagnostics, Atlanta), гидратировали серией этанола, окрашивали 0.
Уши препарировали и фиксировали на льду в течение ночи в 4% формальдегиде в фосфатно-солевом буфере. Для сканирующей электронной микроскопии (SEM) меристемы сушили до критической точки (BAL-TEC CPD 030; Techno Trade, Manchester, NH), а затем устанавливали на углеродные штыри. Образцы покрывали напылением слоем золотого палладия 0,7 Å (BAL-TEC SCD 050, Techno Trade) и просматривали с помощью SEM (JSM 5400; JEOL, Peabody, MA) с использованием ускоряющего напряжения 10 кВ.Для разделения образцы заливали парафином (Paraplast Plus; McCormick Scientific, Сент-Луис). Срезы толщиной 8 мкм вырезали с использованием парафинового микротома Finnesse (Thermo Fisher, Waltham, MA) и помещали на предметные стекла с покрытием (Probe-On Plus; Fisher Scientific, Waltham, MA). Для гибридизации RNA in situ с предметные стекла зондировали DIG-меченным РНК-антисмысловым зондом kn1 согласно Jackson et al. (1994). Для гистологии слайды депарафинизировали с использованием histoclear (National Diagnostics, Atlanta), гидратировали серией этанола, окрашивали 0.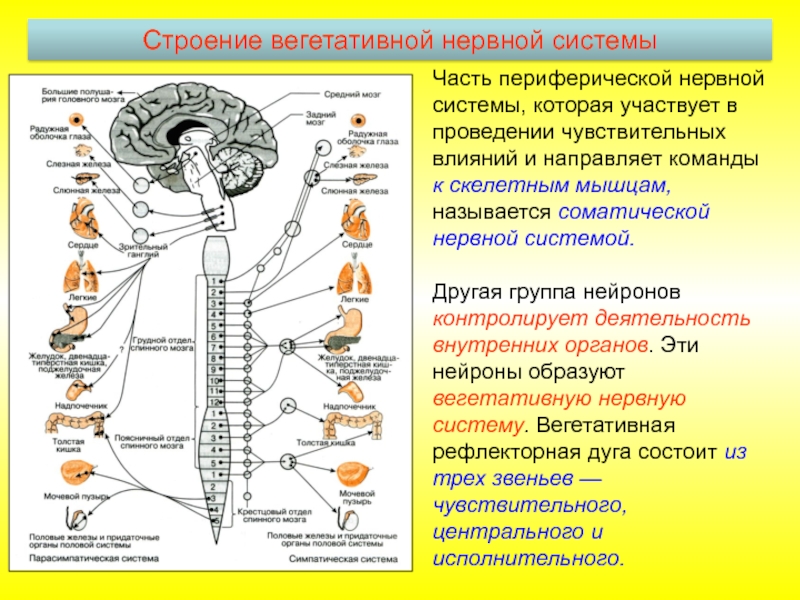 05% толуидиновый синий O (ТВО) в течение 30 секунд, обезвоженный и закрытый покровным стеклом с помощью Histomount (Thermo-Shandon, Pittsburgh). Все слайды просматривали в ярком поле с помощью вертикального микроскопа Eclipse 80i (Nikon, Мелвилл, Нью-Йорк) и фотографировали цифровой камерой DXM1200F (Nikon).
05% толуидиновый синий O (ТВО) в течение 30 секунд, обезвоженный и закрытый покровным стеклом с помощью Histomount (Thermo-Shandon, Pittsburgh). Все слайды просматривали в ярком поле с помощью вертикального микроскопа Eclipse 80i (Nikon, Мелвилл, Нью-Йорк) и фотографировали цифровой камерой DXM1200F (Nikon).
Анализ экспрессии:
Суммарная РНК была выделена из кисточек (5-7 мм) и 8-недельных ушей (20-22 мм) от гомозигот Bif1 и нормальных братьев и сестер в возрасте 5–6 недель с использованием набор для растений Nucleospin RNA в соответствии с протоколом производителя (Macherey-Nagel, Durel, Германия).Одна кисточка или початок (~ 8–12 мг сырой массы) использовалась для экстракции РНК, с тремя биологическими повторами каждого типа образца. Всего 200 нг РНК из каждого образца обрабатывали ДНКазой с использованием набора ДНКазы I (Ambion, Остин, Техас) для удаления загрязнения геномной ДНК. Обратную транскрипцию проводили с использованием набора ABI High-Capacity RT (Applied Biosystems, Foster City, CA) с инкубацией при 25 ° в течение 10 минут и затем при 37 ° в течение 2 часов. Праймеры для ОТ-ПЦР в реальном времени и зонды Taqman, меченные 5′-FAM- (карбоксифлуоресцеин) и 3′-BHQ1- (гаситель черных дыр) (Biosearch Technologies, Novato, CA), были сконструированы с использованием Primer Express версии 2.0 (Applied Biosystems). Пять микролитров кДНК использовали в качестве матрицы для реакций ОТ-ПЦР в реальном времени с использованием универсальной смеси TaqMan 2X (Applied Biosystems), за исключением того, что бетаин (Sigma, St Louis) добавляли до конечной концентрации 0,5 мкМ в реакциях bif2 . Реакции RT – PCR проводили в 96-луночных планшетах с использованием машины для ПЦР в реальном времени ABI 7300 (Applied Biosytems). Для обнаружения экспрессии bif2 зонд Такмана был (FAM-5 ‘CTC CGC CAC CGC ATG CCC 3′-BHQ), а праймеры RT – PCR были bif2F (5’ CTG CGT CGT CAC GGA GTT C 3 ‘) и bif2R (5 ‘TGC CCA TCA TGT GCA GGT ACT 3’).Для обнаружения экспрессии ba1 зонд Такмана был (FAM-5 ‘ACG CGG CTT CCC CAT CAT CCA 3′-BHQ), а праймеры RT – PCR были ba1F (5’ TGG ATC CAT ATC ACT ACC AAA CCA 3 ‘ ) и ba1R (5 ′ ACC GGG TGC TGG AGG TAA G 3 ′).
Праймеры для ОТ-ПЦР в реальном времени и зонды Taqman, меченные 5′-FAM- (карбоксифлуоресцеин) и 3′-BHQ1- (гаситель черных дыр) (Biosearch Technologies, Novato, CA), были сконструированы с использованием Primer Express версии 2.0 (Applied Biosystems). Пять микролитров кДНК использовали в качестве матрицы для реакций ОТ-ПЦР в реальном времени с использованием универсальной смеси TaqMan 2X (Applied Biosystems), за исключением того, что бетаин (Sigma, St Louis) добавляли до конечной концентрации 0,5 мкМ в реакциях bif2 . Реакции RT – PCR проводили в 96-луночных планшетах с использованием машины для ПЦР в реальном времени ABI 7300 (Applied Biosytems). Для обнаружения экспрессии bif2 зонд Такмана был (FAM-5 ‘CTC CGC CAC CGC ATG CCC 3′-BHQ), а праймеры RT – PCR были bif2F (5’ CTG CGT CGT CAC GGA GTT C 3 ‘) и bif2R (5 ‘TGC CCA TCA TGT GCA GGT ACT 3’).Для обнаружения экспрессии ba1 зонд Такмана был (FAM-5 ‘ACG CGG CTT CCC CAT CAT CCA 3′-BHQ), а праймеры RT – PCR были ba1F (5’ TGG ATC CAT ATC ACT ACC AAA CCA 3 ‘ ) и ba1R (5 ′ ACC GGG TGC TGG AGG TAA G 3 ′). Контролем для нормализации был убиквитин : зонд Такмана был (5’FAM-AAA TCC ACC CGT CGG CAC CTC C 3′-BHQ), а праймеры RT – PCR были ubqF (5’CTC TTT CCC CAA CCT CGT GTT 3 ‘ ) и ubqR (5 ‘ACG AGC GGC GTA CCT TGA 3’). Три технических повтора каждой реакции ПЦР в реальном времени были выполнены в трех биологических повторах для каждого эксперимента, и весь эксперимент был повторен дважды.Нормализованные уровни относительной экспрессии определяли с использованием метода сравнительных пороговых значений (Livak and Schmittgen 2001).
Контролем для нормализации был убиквитин : зонд Такмана был (5’FAM-AAA TCC ACC CGT CGG CAC CTC C 3′-BHQ), а праймеры RT – PCR были ubqF (5’CTC TTT CCC CAA CCT CGT GTT 3 ‘ ) и ubqR (5 ‘ACG AGC GGC GTA CCT TGA 3’). Три технических повтора каждой реакции ПЦР в реальном времени были выполнены в трех биологических повторах для каждого эксперимента, и весь эксперимент был повторен дважды.Нормализованные уровни относительной экспрессии определяли с использованием метода сравнительных пороговых значений (Livak and Schmittgen 2001).
Анализы транспорта ауксина:
Анализы транспорта ауксина проводили с использованием метода, модифицированного из Okada et al. (1991) и McSteen et al. (2007). Незрелые соцветия колосов отделяли от растений, выращенных в поле в течение 8 недель. Незрелые уши имели размер от 2 до 3 см. На этой стадии развития меристема соцветия все еще инициировала SPM на кончике, а цветочные органы производились у основания.Два сантиметра кончика уха помещали в любой ориентации в пробирки объемом 2 мл, содержащие 100 мкл 1,5 мкм 3- [5 (n) — 3 H] индолуксусной кислоты (удельная активность 25 Ки / ммоль; GE Healthcare, Пискатауэй, штат Нью-Джерси) в 0,5 × среды Мурашиге и Скуга (Сигма, Сент-Луис). Некоторые пробирки также содержали 20 мкм N -1-нафтилфталаминовую кислоту (Chemservice, West Chester, PA). После 24-часовой инкубации в темноте незрелые ушные вкладыши промокали и 5 мм от конца, который не был погружен в раствор, помещали в сцинтилляционную жидкость (Ready safe; Beckman Coulter, Fullerton, CA) и считали на жидкостном сцинтилляционном счетчике ( LSC6000, Бекман Коултер).Для первоначального эксперимента с нормальными ушами для каждого лечения использовали по три уха, и эксперимент повторяли три раза. Для эксперимента с мутантами Bif1 использовали по три уха из каждого генотипического класса, и эксперимент повторяли четыре раза. Представлены данные, которые являются репрезентативными для одного эксперимента.
Некоторые пробирки также содержали 20 мкм N -1-нафтилфталаминовую кислоту (Chemservice, West Chester, PA). После 24-часовой инкубации в темноте незрелые ушные вкладыши промокали и 5 мм от конца, который не был погружен в раствор, помещали в сцинтилляционную жидкость (Ready safe; Beckman Coulter, Fullerton, CA) и считали на жидкостном сцинтилляционном счетчике ( LSC6000, Бекман Коултер).Для первоначального эксперимента с нормальными ушами для каждого лечения использовали по три уха, и эксперимент повторяли три раза. Для эксперимента с мутантами Bif1 использовали по три уха из каждого генотипического класса, и эксперимент повторяли четыре раза. Представлены данные, которые являются репрезентативными для одного эксперимента.
РЕЗУЛЬТАТЫ
Мутация Bif1 была восстановлена в эксперименте по мутагенезу EMS (Neuffer and Sheridan, 1977) и была картирована на хромосоме 8 с использованием генетических и цитогенетических инструментов (http: // www.maizegdb.org). Используя маркеры SSR, мы точно сопоставили Bif1 между idp98 и umc1360 в ячейке 8.02. Bif1 представляет собой полудоминантную мутацию, при которой гомозигота имеет более тяжелый фенотип, чем гетерозигота. Мутация Bif1 придает фенотип меньшего количества ветвей и колосков в кисточке и меньшего количества ядер в колосе, но фенотип ранее подробно не анализировался (Coe et al., 1988; Sheridan 1988; Veit et al. 1993; Neuffer et al. 1997; McSteen et al. 2000).
Мутанты Bif1 дают меньше ветвей и колосков:Мутантные кисточки Bif1 имели редкий вид с меньшим количеством ветвей и колосков по сравнению с нормальными братьями и сестрами (рис. 1A). Кисточки растений, гомозиготных по Bif1 , были затронуты сильнее, чем гетерозиготы (рис. 1А). Растения, гетерозиготные по Bif1 , давали колосья с неравномерной греблей из-за уменьшения количества зерен и бесплодия на кончике (рис. 1В).Растения, гомозиготные по Bif1 , давали уши с очень небольшим количеством ядер, так что был виден голый рахис (фигура 1B).
Рисунок 1.—Фенотип зрелого соцветия мутанта Bif1 . (A) Зрелые кисточки нормальных, Bif1 / + и Bif1 / Bif1 растений. У обычной кисточки длинные ветки обозначены у основания главного колоса. Пары колосков покрывают ветви и главный колос. У мутантов Bif1 в кисточке снижено количество ветвей и колосков.(B) Зрелые колосья нормальных растений, растений Bif1 / + и Bif1 / Bif1 , демонстрирующих меньшее количество ядер и неорганизованные ряды у мутантов Bif1 . (C) Количественная оценка количества веток кисточки. (D) Количественная оценка количества колосков кисточки. (E) Процент колосков, которые встречаются поодиночке против парных. Столбцы представляют собой среднее значение, а столбцы ошибок представляют стандартную ошибку среднего.
Количественный анализ зрелой метелки Bif1 показал, что мутанты Bif1 не способны продуцировать полный набор ветвей метелки (рис. 1С).Анализ количества колосков показал статистически значимое снижение количества колосков у растений, гетерозиготных и гомозиготных по Bif1 (рис. 1D). У растений, гомозиготных по Bif1 , образующиеся колоски иногда производились по отдельности, а не парами (рис. 1E). Уменьшение количества продуцируемых ветвей и колосков указывает на то, что инициация или поддержание первичных подмышечных меристем, BM и SPM, является дефектным в соцветиях Bif1 .
Мутанты Bif1 не могут инициировать SPM:Фенотип зрелого соцветия Bif1 предполагает наличие дефектов на ранних стадиях развития соцветия.Чтобы проверить это, СЭМ использовали для визуализации развивающегося соцветия. К 5 неделям роста нормальные соцветия образовали несколько боковых ветвей, причем SPM были видны как правильные бугорки на боках как главного колоса, так и ветвей (рис. 2А). На той же стадии развития растения, гетерозиготные по Bif1 , имели пониженное количество SPM (фиг. 2B). Растения, гомозиготные по Bif1 , имели более тяжелый фенотип с очень небольшим количеством или, в некоторых случаях, без SPM (Рисунок 2C). Бесплодная поверхность рахиса была очень немного ребристой (рис. 2С).Гомозиготное ухо Bif1 имело такой же фенотип, что и кисточка, с несколькими инициированными SPM (рис. 2E). В отличие от кисточки, меристема соцветия колоса была фасцирована (рис. 2Е).
Рис. 2.—Изображения развивающихся соцветий Bif1 , полученные с помощью сканирующей электронной микроскопии (СЭМ). (A) Нормальная кисточка, показывающая файлы развивающихся меристем пары колосков (SPM) на боках меристемы соцветия (IM). (B) Bif1 / + кисточка с уменьшенным количеством SPM. (C) Bif1 / Bif1 кисточка с несколькими SPM.(D) Нормальное ухо, демонстрирующее организованные ряды SPM. (E) Колос Bif1 / Bif1 с фасциированной меристемой соцветия и небольшим количеством SPM. Пруток, 100 мкм.
Чтобы определить, были ли какие-либо гистологические доказательства образования SPM, мы использовали TBO для окрашивания срезов развивающихся соцветий Bif1 . Поскольку меристематические клетки имеют меньшие вакуоли, чем дифференцированные клетки, SPM окрашиваются TBO более интенсивно, чем окружающие ткани. В нормальных соцветиях развивающиеся SPM были видны как регулярные группы плотно окрашенных клеток на боках соцветия (рис. 3A).У растений Bif / + SPM, видимые на боках соцветия, были похожи на нормальные, за исключением того, что их было меньше (рис. 3B). Гомозиготы Bif1 в основном не продуцировали SPM (рис. 3C). Вместо этого поверхность бесплодных областей соцветия иногда имела очень слегка приподнятые выступы, которые были менее интенсивно окрашены, чем SPM, что указывает на то, что небольшие гребни, видимые с помощью SEM, состоят из дифференцированной ткани (рис. 3C).
Рисунок 3.—Гистология и РНК in situ гибридизация с kn1 в развивающихся кистях Bif1 .(A – C) Продольные срезы кисточек 5-недельного возраста, окрашенные TBO, с SPM, видимыми как области интенсивного окрашивания. (A) Три развивающихся SPM на боках меристемы соцветия в нормальной кисточке. (B) Bif1 / + , показывающий один SPM в той же области, что и три SPM в нормальном состоянии. (C) Bif1 / Bif1 с небольшим выступом на поверхности позвоночника, но без признаков развития SPM. (D – F) РНК in situ гибридизация с kn1 . (D) Меристематические клетки и сосудистая сеть обозначены экспрессией kn1 в нормальных кистях.Отсутствие kn1 на боках меристемы соцветия (IM) указывает на образование подавленных зачатков прицветников (BR), которые прилегают к SPM. (E) Соцветия Bif1 / + имеют меньшее количество областей экспрессии kn1 на боках соцветия. (F) Соцветие Bif1 / Bif1 с экспрессией kn1 только в меристеме соцветия и в сосудистой сети. Пруток, 100 мкм.
Для определения наличия каких-либо молекулярных свидетельств образования SPM, гибридизацию РНК in situ с kn1 использовали в качестве маркера для идентификации меристематических тканей. kn1 экспрессируется в меристемах, где он необходим для поддержания меристемы, и не экспрессируется как дифференцированные зачатки органа (Jackson et al. 1994; Kerstetter et al. 1997; Vollbrecht et al. 2000). У нормальных растений экспрессия kn1 была четко видна в меристеме соцветия, а также в сосудистой сети и стебле (фигура 3D). kn1 не экспрессировался на боках меристемы соцветия, поскольку зародыши прицветника (последующий рост которых подавляется) инициируются (обозначены BR на фигуре 3D).Однако kn1 сильно экспрессируется в SPMs, которые формируются в пазухах зачатков прицветников (Figure 3D). У мутантов Bif1 , kn1 экспрессировался как нормальный в меристеме, сосудистой сети и стебле соцветия (фиг. 3, E и F). В растениях, гетерозиготных по Bif1 , области без экспрессии kn1 чередовались с областями экспрессии kn1 , как ожидалось для нескольких SPM, которые инициируют (фиг. 3E). У растений, гомозиготных по Bif1 , обычно не было свидетельств образования SPM на боках меристемы соцветия (фиг. 3F).Поскольку подавление kn1 было видно на боках меристемы соцветия Bif1 (рис. 3, E и F), это указывает на то, что зачатки прицветников отделены у мутантов Bif1 и что случайные небольшие выступы видны у Bif1. Мутанты могут быть подавленными зачатками прицветника. Однако in situs с kn1 ясно показывают, что SPM не образуются в пазухах зачатков прицветников у растений, гомозиготных по Bif1 .
Мутанты Bif1 имеют дефекты в инициации SM, а не в детерминации SPM:В нормальных растениях на главном колосе и боковых ветвях SPM продуцируют пару SM. Мутанты Bif1 имеют уменьшенное количество колосков, в значительной степени из-за уменьшенного количества SPM. Когда SPMs инициируются у мутантов Bif1 , они часто производят одиночные, а не парные колоски (Рис. 1E). Это указывает на то, что мутанты Bif1 имеют дефекты либо в инициации SM, либо в детерминированности SPM. Чтобы различать эти две возможности, мы сконструировали двойные мутанты между Bif1 и ramosa1 ( ra1 ). ra1 кодирует фактор транскрипции «цинковые пальцы» EPF, который придает детерминированность SPM (Vollbrecht et al. 2005). У мутантов ra1 SPMs лишены детерминированности и вырастают, чтобы стать длинными неопределенными ветвями, вместо того, чтобы давать детерминированные пары колосков (Gernart 1912; Vollbrecht et al. 2005). В результате мутанты ra1 и образуют дополнительные длинные ответвления как в кисточке, так и в початке (рис. 4, A и B).
Рисунок 4.—Анализ Bif1; ra1 двойные мутанты. (A) Фенотип зрелой метелки, показывающий все генетические классы из сегрегации Bif1 ; ra1 семейство.(B) фенотип уха сегрегирующего Bif1 ; ra1 семейство. (C – E) Изображения с большим увеличением, показывающие отдельные ветви от ушей. (C) Ответвление от уха ra1 / ra1 . (D) Ответвление от Bif1 / +; ra1 / ra1 ухо. (E) Bif1 / Bif1; ra1 / ra1 ухо. (F – I) Количественный анализ Bif1; ra1 двойные мутанты. Для всех диаграмм столбцы представляют собой среднее значение набора данных, а столбцы ошибок представляют стандартную ошибку среднего. (F) Среднее количество ветвей на кисть.(G) Среднее количество колосков на кисточку. (H) Среднее количество колосков на ветке, измеренное на ветке у основания кисточки. (I) Среднее количество колосков на верхних 2 см главного колоса кисточки.
Мы обнаружили, что даже на фоне мутанта ra1 гомозиготы Bif1 были неспособны образовывать дополнительные длинные ответвления в кисточке (рис. 4A). Модель Bif1 / Bif1; Двойной мутант ra1 / ra1 имел фенотип бесплодной кисточки, подобный Bif1 (фиг. 4A).Количественный анализ показал, что общее количество ветвей и количество колосков статистически не различались между Bif1 / Bif1; ra1 / ra1 и Bif1 / Bif1 (номер ответвления, -значение P = 0,054; номер колоска, -значение P = 0,192; рис. 4, F и G). Следовательно, когда у гомозигот Bif1 не были продуцированы SPM, ra1 не мог действовать на них. С другой стороны, в Bif1 / Bif1; ra1 / ra1 , несколько ветвей выросли из рахиса (Рисунок 4B, крупный план показан на рисунке 4E).Оказалось, что когда SPM инициировались в Bif1 / Bif1; ra1 / ra1 уха, они превратились в ответвления из-за отсутствия ra1 (рис. 4, B и E). Следовательно, мутант Bif1 не имеет дефектов в детерминированности SPM после того, как SPMs инициированы.
Дальнейшее понимание было получено путем характеристики Bif1 / +; Двойные мутанты ra1 / ra1 . Кисточка Биф1 / +; Двойные мутанты ra1 / ra1 имели более длинные ветки и колоски, чем Bif1 / + (фиг. 4A).Количественный анализ показал значительное увеличение количества филиалов у Bif1 / +; ra1 / ra1 по сравнению с Bif1 / + (значение P <0,001, рис. 4F). Это говорит о том, что когда возникли ВОП, они выросли и стали боковыми ответвлениями. Однако ветки на Bif1 / +; ra1 / ra1 были более бесплодными, чем типичные ветви ra1 , и не имели статистически различающегося количества колосков по сравнению с растениями Bif1 / + ( P -значение = 0.94; Рис. 4, G – I). Аналогично Bif1 / +; ra1 / ra1 был более разветвленным, чем Bif1 / Bif1; ra1 / ra1 , но на ветвях образовалось мало колосков (рис. 4В, крупный план отдельной ветви, показанной на рис. 4D). Эти результаты указывают на то, что мутанты Bif1 имеют дефекты инициации вторичных подмышечных меристем, SM, скорее, чем дефекты детерминированности первичных подмышечных меристем, SPM.
Колоски и меристемы цветков являются дефектными у мутантов
Bif1 :Рассечение нескольких колосков, образовавшихся у мутантных растений Bif1 , показало, что колоски Bif1 имели меньше цветков, чем обычно, и у цветков было меньше цветковых органов.Для количественной оценки этих дефектов было вырезано 100 колосков как с гетерозиготных, так и гомозиготных растений по Bif1 и определено количество цветков и органов цветка относительно нормальных сибсов.
У нормальных растений каждый колоск несет пару цветков (рис. 5А). У гетерозиготных растений Bif1 только 64% колосков давали два цветочка (рис. 5A), тогда как у гомозигот Bif1 только 3% колосков давали два цветочка (рис. 5A). Эти результаты показывают, что у мутантов Bif1 SM дефектны, поскольку они неспособны инициировать нормальное количество третичных подмышечных меристем, FM.
Рисунок 5.—Количественная оценка количества цветков и органов цветка у мутантов Bif1 . (A) Процент колосков, содержащих два, один или ноль цветков на один колоск. (B) Количественная оценка леммы и числа палеи на цветочек. (C) Процент цветков, содержащих указанное количество тычинок на цветочек.
Нормальные кисточки дают соцветия, содержащие лемму, палею, две лодикулы и три тычинки. Количественный анализ показал, что у мутантов Bif1 соцветия имели меньше цветочных органов, чем обычно, причем гомозигота была поражена сильнее, чем гетерозигота (рис. 5, B и C).Количество лемм и палеев было уменьшено у мутантов Bif1 с 87% колосков от гетерозигот Bif1 и 56% колосков от гомозигот Bif1 , продуцирующих оба органа (рис. 5B). Лодикулы не учитывались, так как их небольшой размер и прозрачность затрудняли точный подсчет их под препарирующим микроскопом. Число тычинок было снижено у мутантов Bif1 только с 29% цветков Bif1 / + и 12% соцветий Bif1 / Bif1 , содержащих три нормальные тычинки (фигура 5C).Интересно, что небольшой процент цветков мутанта Bif1 содержал четыре тычинки, что указывает на то, что количество цветочных органов может как увеличиваться, так и уменьшаться. Неспособность инициировать нормальное количество органов цветков указывает на то, что FMs также являются дефектными у мутантов Bif1 .
Исследования экспрессии показывают, что
bif2 и ba1 экспрессируются на более низком уровне у мутантов Bif1 :Подобно Bif1 , bif2 и ba1 мутанты также являются дефектными в инициации всех типов мутантов. пазушные меристемы соцветия (McSteen, Hake, 2001; Ritter et al. 2002). Чтобы определить, влияет ли мутация Bif1 на экспрессию bif2 или ba1 , были проведены эксперименты с ОТ-ПЦР в реальном времени. bif2 и ba1 оба экспрессируются в кистях и ушах у нормальных растений (Gallavotti et al. 2004; McSteen et al. 2007). bif2 экспрессируется в подмышечных меристемах, латеральных органах и сосудистой сети, тогда как ba1 имеет более ограниченный паттерн экспрессии во время инициации подмышечной меристемы (Gallavotti et al. 2004; McSteen et al. 2007). РНК выделяли из незрелых кисточек и колосьев растений, гомозиготных по Bif1 , и от нормальных братьев и сестер. Эксперименты с ОТ-ПЦР в реальном времени показали, что уровни РНК bif2 и ba1 были снижены в кистях и початках растений, гомозиготных по Bif1 (фиг. 6, A – D). Уровни bif2 были снижены до 36–62% от нормального уровня в кисточке и ушах, соответственно (рис. 6, А и В), что частично можно объяснить сокращением количества BM, SPM, SM, FM, и органы цветков у мутантов Bif1 .С другой стороны, уровни ba1 резко снизились до 4–13% от нормального уровня (рис. 6, C и D). Принимая во внимание, что ba1 экспрессируется в очень ограниченном паттерне при инициировании подмышечной меристемы, эти результаты подтверждают, что мутация Bif1 влияет на ранние стадии инициации подмышечной меристемы.
Рисунок 6.—Анализ ОТ-ПЦР в реальном времени экспрессии bif2 и ba1 в мутантах Bif1 . (A) Уровень экспрессии bif2 в незрелой кисточке мутантов Bif1 по сравнению с нормальными братьями и сестрами.(B) Уровень экспрессии bif2 в незрелых ушах мутантов Bif1 относительно нормальных братьев и сестер. (C) Уровень экспрессии ba1 в незрелой кисточке мутантов Bif1 относительно нормальных братьев и сестер. (D) Уровень экспрессии ba1 в незрелых ушах мутантов Bif1 относительно нормальных братьев и сестер. Среднее значение плюс или минус SE показано для одного репрезентативного эксперимента с использованием трех биологических и трех технических повторов для каждого образца.
Анализ двойных мутантов показывает, что
bif1 и bif2 играют роль в вегетативном развитии: мутантыbif2 имеют фенотип, очень похожий на фенотип гомозигот Bif1 , с очень небольшим количеством ветвей кистей и колосков (McSteen и Hake 2001; McSteen et al. 2007). Для определения генетического взаимодействия между Bif1 и bif2 были сконструированы линии двойных мутантов. Bif1; Растения с двойным мутантом bif2 оказывали очень сильное влияние как на вегетативное развитие, так и на развитие соцветий.
Фенотип соцветия:
Фенотип соцветия Bif1; Двойные мутанты bif2 были более серьезными, чем у одинарных мутантов без ветвей или колосков (Рисунок 7, A – C). Количественный анализ показал, что отсутствие колосков у Bif1 / Bif1; Двойные мутанты bif2 / bif2 показали статистически значимое снижение количества колосков по сравнению с Bif1 / Bif1 ( P -значение = 0,004) или bif2 одиночными мутантами ( P -значение = 0.001, фигура 7C). Кроме того, генетический и молекулярный анализ показал, что растения, которые были гетерозиготными по Bif1 и гомозиготными по bif2 , напоминали гомозиготы Bif1 (фиг. 7, A – C). Эти результаты предполагают, что bif1 и bif2 играют избыточные роли в инициации ветвей и колосков в соцветии.
Рисунок 7.— АнализBif1; bif2 двойные мутанты. (A) Фенотип зрелой кисточки Bif1; bif2 сегрегация семьи.(B и C) Количественная оценка характеристик кисточки в Bif1 ; bif2 сегрегационная семья. (B) Среднее количество веток с кисточкой. (C) Среднее количество колосков на кисточку. (D) Вегетативный фенотип Bif1; семейство bif2 , показывающее уменьшенную высоту растений в Bif1; bif2 двойные мутанты. (E и F) Количественная оценка вегетативных фенотипов. (E) Средняя высота растения в сантиметрах. (F) Среднее количество листьев.
Вегетативный фенотип:
Bif1; Растения с двойным мутантом bif2 были меньше половины высоты нормальных растений (фиг. 7, D и E).Чтобы определить, вызвано ли уменьшение высоты растений разницей в количестве продуцируемых фитомеров, подсчитывали количество листьев (рис. 7F). И Bif1 , и bif2 (McSteen et al. 2007) сами по себе оказывают незначительное влияние на количество листьев, с небольшим, но статистически значимым сокращением количества листьев по сравнению с обычными братьями и сестрами ( P -значение = 0,001, рисунок 7F). Однако Bif1 / Bif1; Двойной мутант bif2 / bif2 имел неаддитивный эффект с большим и значительным уменьшением количества листьев по сравнению с Bif1 / Bif1 (значение P <0.001) или bif2 / bif2 ( P -значение = 0,001) одиночных мутантов. Резкое влияние на количество листьев в Bif1; Двойной мутант bif2 подразумевает, что bif1 и bif2 также играют избыточные роли в продукции листьев вегетативной апикальной меристемой побега.
Анализ двойных мутантов показывает, что
Bif1 эпистатичен по отношению к ba1 в кисточке :Мутант бесплодный стебель1 ( ba1 ) испытывает дефицит как в вегетативной, так и в соцветийной части пазушной меристемы. отсутствуют побеги и колосья, а также веточки и колоски на кисточке (Ritter et al. 2002; Gallavotti et al. 2004). Хотя эпистаз сложно определить, когда мутанты имеют сходный фенотип, мутанты Bif1 можно отличить от мутантов ba1 по внешнему виду стебля соцветия (рахиса). Мутанты Bif1 имеют гладкий тонкий стержень, в то время как у мутантов ba1 толстый стержень с очень регулярными выраженными выступами из-за образования более крупных, чем обычно, подавленных зачатков прицветника (Ritter et al. 2002). Bif1 / +; двойные мутанты ba1 / ba1 напоминали одиночные мутанты ba1 ; однако поверхность рахиса была немного более гладкой, чем обычно наблюдается у кистей ba1 (рис. 8A). Bif1 / Bif1; Двойные мутанты ba1 / ba1 напоминали гомозиготы Bif1 с гладкой тонкой кисточкой оси. Как Bif1 / Bif1; Двойной мутант ba1 / ba1 устраняет регулярные выступы, обычно наблюдаемые у мутантов ba1 , это указывает на то, что Bif1 эпистатичен по отношению к ba1 в кисточке.Фенотип соцветия двойного мутанта не был усилен в отношении количества колосков, что не было неожиданным, поскольку мутанты ba1 обычно не производят никаких колосков (фигура 8B). Точно так же двойной мутант не дал ушей (рис. 8C). Более того, этот анализ также показал, что одни только мутанты Bif1 не имели каких-либо дефектов в производстве ушей (фигура 8C). В отличие от взаимодействия между Bif1 и bif2 , не наблюдалось усиления вегетативных дефектов Bif1 на ba1 (данные не показаны), что указывает на то, что ba1 не играет избыточной роли в инициации листьев во время вегетативного периода. разработка.
Рисунок 8.—Анализ Bif1; ba1 двойные мутанты. (A) Фенотип зрелой кисточки Bif1; ba1 семья. (B) Среднее количество колосков на кисточку. (C) Среднее количество колосьев на растение.
мутанты Bif1 имеют пониженный уровень транспорта ауксина:Поскольку мутанты Bif1 имели столь сильное взаимодействие с bif2 , который играет роль в транспорте ауксина, мы проверили, имеют ли мутанты Bif1 дефекты в транспорт ауксина. bif2 мутанты обладают пониженным уровнем транспорта ауксина в зрелом стебле соцветия кисточки (McSteen et al. 2007). Предварительные эксперименты показали, что мутанты Bif1 аналогичным образом обладали сниженным транспортом в зрелом стебле соцветия кисточки (данные не показаны). Однако мутанты Bif1 и bif2 имеют пониженную сосудистую сеть в зрелом стебле соцветия (McSteen et al. 2007 и данные не показаны). Мутанты Bif1 также имеют сокращение сосудистой сети в незрелых соцветиях кисточки на ранней стадии развития (дополнительная фигура S1, A – C).Однако мутанты Bif1 не обладают значительным сокращением сосудистой сети в развивающемся соцветии колоса (дополнительная фигура S1, D – F). Поэтому, чтобы определить, имели ли мутанты Bif1 дефекты транспорта ауксина на ранней стадии развития, мы разработали протокол для измерения транспорта ауксина в соцветии уха.
Незрелые колосовые соцветия длиной до 2 см инкубировали в течение ночи в любой ориентации в растворе 1,5 мкм 3 H-меченой ИУК.Уши дикого типа показали заметный уровень базипетального транспорта, который подавлялся совместной инкубацией с 20 мкМ N -1-нафтилфталаминовой кислоты (NPA), часто используемым ингибитором транспорта ауксина (рис. 9A, дорожки 1 и 2). Однако акропетальный транспорт на этом этапе развития был очень низким (рис. 9А, дорожки 3 и 4).
Рисунок 9.—Измерение транспорта ауксина в нормальных соцветиях и соцветиях Bif1 . Темное затенение, без NPA; светлая штриховка, с NPA. (A) Измерение базипетального (дорожки 1 и 2) и акропетального (дорожки 3 и 4) транспорта в нормальных ушах.(B) Измерение базипетального транспорта в незрелых ушах семьи сегрегации для Bif1 . Дорожки 1 и 2, нормальные братья и сестры; дорожки 3 и 4, Bif1 / +; полосы 5 и 6, Bif1 / Bif1 .
Чтобы проверить, имеют ли соцветия колоса Bif1 сниженный уровень транспорта ауксина, базипетальный транспорт измеряли у растений, которые были гетерозиготными или гомозиготными по Bif1 по сравнению с нормальными братьями и сестрами (фигура 9B). Растения, которые были гетерозиготными по Bif1 , имели примерно одну треть уровня транспорта, как у нормальных братьев и сестер (фигура 9B, дорожка 3), и эти уровни дополнительно снижались путем совместной инкубации с 20 мкМ NPA (фигура 9B, дорожка 4).У гомозиготных ушей Bif1 наблюдалось еще большее снижение активного транспорта ауксина (фигура 9B, дорожка 5), которое не отличалось значительно от такового у нормальных братьев и сестер, получавших NPA (значение P = 0,75). Следовательно, мутанты Bif1 имеют пониженный уровень транспорта ауксина, что указывает на то, что первичный дефект у мутантов Bif1 может заключаться в регуляции транспорта ауксина.
ОБСУЖДЕНИЕ
Мы определили нового участника на пути инициации подмышечной меристемы во время развития соцветий кукурузы.Мутанты Bif1 имеют очень сходный фенотип с мутантами bif2 с дефектами инициации всех пазушных меристем в соцветии. Синергетическое взаимодействие Bif1 с bif2 указывает на то, что bif1 действует избыточно с bif2 как во время вегетативного развития, так и во время развития соцветий. Мы предполагаем, что дефекты в мутантах Bif1 вызваны снижением транспорта ауксина и что функция bif1 заключается в регулировании транспорта ауксина.
bif1 играет роль в инициации подмышечной меристемы:Растения, гомозиготные по Bif1 , имеют фенотип, очень похожий на мутанты bif2 (McSteen and Hake 2001). Сходство с мутантами bif2 включает уменьшение количества ветвей, колосков, цветков и органов цветка в кисточке и уменьшение количества ядер в колосе. Более того, мутанты Bif1 продуцируют одиночные, а не парные колоски, что также характерно для мутантов bif2 .Кисточка и рахис уха гладкие, с редкими неровными выступами, как у bif2 . Кроме того, меристема верхушечного соцветия колоса может быть фасциирована, как и у bif2 .
Характеристика развивающегося соцветия с помощью анализа SEM, гистологии и экспрессии kn1 показывает, что существует специфический дефект в инициации подмышечной меристемы у мутантов Bif1 . Мы предполагаем, что bif1 играет роль в инициации пазушной меристемы в соцветии.У мутантов поражены все пазушные меристемы соцветия — BM, SPM, SM и FM. Однако, в отличие от bif2 , мутанты Bif1 не имеют дефектов инициации подмышечной меристемы, дающей начало отростку уха. Уши образуются у мутантов Bif1 и как нормальные, и не наблюдается усиления дефектов числа ушей в Bif1; bif2 двойные мутанты (данные не представлены). Более того, в отличие от bif2 , анализ двойных мутантов с teosinte разветвленный1 ( tb1 ) (Doebley et al. 1997; Хаббард и др. 2002; McSteen et al. 2007) показывает, что мутация Bif1 не оказывает существенного влияния на продукцию вегетативной подмышечной меристемы (побеги) (данные не показаны). Таким образом, одно из немногих различий между мутациями Bif1 и bif2 заключается в степени их влияния на продукцию побегов и колосьев.
Во время вегетативного развития мутанты Bif1 имеют небольшое, но значительное уменьшение количества листьев, что приводит к сопутствующему уменьшению высоты растения.Мутанты bif2 также оказывают незначительное влияние на зарождение листьев во время вегетативного развития (McSteen et al. 2007). Драматический эффект Bif1; Двойной мутант bif2 по вегетативному развитию указывает на то, что bif1 и bif2 играют избыточные роли в продукции листьев вегетативной апикальной меристемой. Таким образом, наш анализ показывает, что помимо роли bif1 и bif2 в инициации пазушных меристем во время развития соцветий, bif1 и bif2 также играют перекрывающиеся роли в продукции боковых органов во время вегетативного развития.
Роль
bif1 в транспорте ауксина:Градиенты ауксина необходимы для полярного роста растений (Benkova et al. 2003; Heisler et al. 2005). У мутантов pinformed1 ( pin1 ) и pinoid ( pid ) у Arabidopsis снижение транспорта ауксина отменяет инициацию подмышечных меристем, что приводит к фенотипу соцветия pin , аналогичному соцветию . Фенотип кукурузы (Okada et al. 1991; Bennett et al. 1995; Galweiler et al. 1998; Рейнхардт и др. 2003). Двойные мутанты у членов семейства гена YUCCA , необходимые для биосинтеза ауксина, также вызывают фенотип соцветия pin (Cheng et al. 2006). Однако либо потеря, либо усиление функции фактора транскрипции MONOPTEROS приводит к фенотипу соцветия pin , иллюстрируя, что потеря или усиление передачи сигналов ауксина отменяет инициацию подмышечной меристемы (Przemeck et al. 1996; Hardtke et al. 2004). Следовательно, дефекты биосинтеза, транспорта или реакции ауксина приводят к неспособности инициировать аксиллярные меристемы в соцветии у Arabidopsis (Cheng and Zhao 2007).
Мы предполагаем, что bif1 действует вместе с bif2 в контроле транспорта ауксина в соцветиях кукурузы. Многие из фенотипов, наблюдаемых у мутантов Bif1 и bif2 , также наблюдаются у растений, обработанных ингибиторами транспорта ауксина (Wu and McSteen 2007).Например, неспособность инициировать пазушные меристемы в соцветии, одиночные колоски, снижение сосудистой сети и меньшее количество листьев также наблюдается у растений, обработанных полярными ингибиторами транспорта ауксина (Scanlon 2003; Wu and McSteen 2007). Таким образом, мы проверили уровни транспорта ауксина в соцветиях мутантов Bif1 и обнаружили, что транспорт ауксина снижается, что означает, что bif1 играет роль в транспорте ауксина.
Мутация Bif1 является полудоминантной, поэтому она может быть либо доминантной потерей функции ( e.g ., антиморф или гипоморф) или доминантная мутация увеличения функции (, например, ., гиперморф или неоморф). Невозможно было использовать анализ дозировки, чтобы определить, является ли Bif1 мутацией потери или усиления функции, поскольку Bif1 не обнаруживается известными линиями транслокации на хромосоме 8. Однако, поскольку мутация Bif1 вызывает снижение транспорта ауксина, мы можем сделать вывод, что ген bif1 является либо положительным, либо отрицательным регулятором транспорта ауксина.
Генетическое взаимодействие между
Bif1 и другими бесплодными соцветиями мутаций:Чтобы определить генетическое взаимодействие между Bif1 и ранее известными мутациями бесплодного соцветия , мы провели анализ двойных мутантов и экспрессии. Мы заключаем, что bif1 действует перед ba1 как Bif1; Двойной мутант ba1 напоминал Bif1 в кисточке. Кроме того, уровни экспрессии ba1 были резко снижены у мутантов Bif1 .Дальнейшее подтверждение этой гипотезы обеспечивается предположением, что ba1 действует ниже транспорта ауксина (Wu and McSteen 2007). Мутант ba1 продуцирует прицветники очень регулярным образом, указывая на то, что филлотаксис у мутанта не нарушен и что транспорт ауксина является нормальным (Ritter et al. 2002). Более того, ba1 не экспрессируется после обработки ингибиторами транспорта ауксина, что указывает на то, что экспрессия ba1 зависит от транспорта ауксина (Wu and McSteen 2007).Мы предполагаем, что ba1 , являясь фактором транскрипции, необходим для ответа на сигнал ауксина для инициации подмышечной меристемы. Мы предполагаем, что bif1 действует выше транспорта ауксина и, следовательно, выше ba1.
Анализ экспрессии показывает, что уровни bif2 несколько снижены у мутантов Bif1 . Некоторое снижение экспрессии bif2 можно объяснить отсутствием структур, которые экспрессируют bif2 , или этот результат может означать, что Bif1 действует выше bif2 .Однако синергетический эффект наблюдается у Bif1; Двойные мутанты bif2 подразумевают, что bif1 и bif2 имеют перекрывающиеся функции. Оба мутанта имеют очень похожий фенотип, но двойной мутант намного более серьезен, чем любой одиночный мутант, что указывает на то, что bif1 и bif2 могут играть избыточную роль в вегетативном развитии и развитии соцветий. Эффект дозировки Bif1; Взаимодействие bif2 также подтверждает, что они влияют на один и тот же процесс.Из результатов анализа двойных мутантов и экспрессии вместе с предыдущими результатами мы предполагаем, что bif1 и bif2 оба действуют выше транспорта ауксина.
Чтобы определить молекулярный механизм, с помощью которого bif1 регулирует транспорт ауксина, будущая работа будет идентифицировать ген bif1 путем клонирования на основе карты. Благодаря секвенированию генома кукурузы и доступности последовательности генома родственных трав, ходьба по хромосомам стала обычным явлением для кукурузы (Salvi et al. 2002; Wang et al. 2005; Alleman et al. 2006; Bortiri et al. 2006а, б; Satoh-Nagasawa et al. 2006; Тарамино и др. 2007). Многие регуляторы транспорта ауксина были идентифицированы у других видов; однако только мутанты pin и pid имеют фенотип соцветий pin (Brown et al. 2001; Gil et al. 2001; Noh et al. 2001; Geisler et al. 2003 ; Geldner et al. 2003; Multani et al. 2003; Bennett et al. 2006; Sieburth et al. 2006). Ближайшие гомологи кукурузы pin и pid не картируются с Bif1 , что указывает на то, что ген bif1 , возможно, может быть новым регулятором транспорта ауксина в соцветии.
Благодарности
P.M. благодарит Сару Хейк за ее наставничество в начале разработки проекта бесплодных соцветий .Мы благодарим Тони Омейса, У. Скотта Харккома и Боба Оберхейма за уход за растениями в теплицах Департамента биологии и на фермах Департамента садоводства и растениеводства и почвоведения в Рок-Спрингс, штат Пенсильвания. Мы благодарим Кэрри Барриос за выполнение некоторых генетических скрещиваний, Молли Савейкис за первоначальный анализ SEM, Мисси Хазен за обучение работе с SEM, Деб Гроув за помощь в проведении экспериментов RT – PCR в реальном времени и студентов бакалавриата Джейсона Хоара, Джеффри Бутербо, Кима Филлипсу, Мэтту Дэвису и Крису Куку за помощь в анализе двойных мутантов в полевых условиях.Мы благодарим сотрудников лабораторий Брауна и Макстина за обсуждение и критическое прочтение рукописи. Это исследование было профинансировано стипендией Huck Institutes of Life Science для выпускников S.B. и грантом Национального научного фонда № IOS-0416616 к P.M.
- Получено 5 ноября 2007 г.
- Принято 15 февраля 2008 г.
- Авторское право © 2008 Американского генетического общества
Ссылки
- ↵
Alleman, M., Л. Сидоренко, К. МакГиннис, В. Сешадри, Дж. Э. Дорвейлер, и др. , 2006 РНК-зависимая РНК-полимераза необходима для парамутации кукурузы. Nature 442 : 295–298.
- ↵
Benjamins, R., A. Quint, D. Weijers, P. Hooykaas и R. Offringa, 2001 Протеинкиназа PINOID регулирует развитие органов у Arabidopsis путем усиления полярного транспорта ауксина. Разработка 128 : 4057–4067.
- ↵
Бенкова, Е., М. Мичневич, М. Зауэр, Т. Тейхманн, Д. Сейфертова и др. , 2003 Местные, зависимые от оттока градиенты ауксина как общий модуль для формирования органов растений. Ячейка 115 : 591–602.
- ↵
Беннет, С. Р. М., Дж. Альварес, Г. Боссингер и Д. Р. Смит, 1995 Морфогенез у мутантов пиноидов Arabidopsis thaliana. Плант Дж. 8 : 505–520.
- ↵
Беннет, Т., Т. Сиберер, Б. Виллет, Дж.Букер, C. Luschnig et al. , 2006 Путь Arabidopsis MAX контролирует ветвление побегов путем регулирования транспорта ауксина. Curr. Биол. 16 : 553–563.
- ↵
Боммерт П., Н. Сато-Нагасава, Д. Джексон и Х. Ю. Хирано, 2005 г. Генетика и эволюция соцветий и развитие цветков у трав. Physiol растительной клетки. 46 : 69–78.
- ↵
Bortiri, E., and S. Hake, 2007 Цветение и детерминированность кукурузы.J. Exp. Бот. 58 : 909–916.
- ↵
Bortiri, E., G. Chuck, E. Vollbrecht, T. Rocheford, R. Martienssen et al. , 2006a ramosa2 кодирует белок LATERAL ORGAN BOUNDARY domain, который определяет судьбу стволовых клеток в меристемах ветвей кукурузы. Растительная клетка 18 : 574–585.
- ↵
Бортири, Э., Д. Джексон и С. Хейк, 2006b Достижения в геномике кукурузы: появление позиционного клонирования.Curr. Opin. Plant Biol. 9 : 164–171.
- ↵
Браун Д. Э., А. М. Рашотт, А. С. Мерфи, Дж. Норманли, Б. В. Таг и др. , 2001 Флавоноиды действуют как негативные регуляторы транспорта ауксина in vivo у Arabidopsis. Plant Physiol. 126 : 524–535.
- ↵
Чен, Дж. И С. Л. Деллапорта, Минипреп по растительной ДНК на основе мочевины, 1994, стр. 526–527 в издании The Maize Handbook , под редакцией М. Фрилинга и В.Walbot. Спрингер-Верлаг, Нью-Йорк.
- ↵
Cheng, P. C., R. I. Greyson и D. B. Walden, 1983 г. Инициирование органов и развитие однополых цветков в кисточке и колосе Zea mays. Am. J. Bot. 70 : 450–462.
- ↵
Cheng, Y., X. Dai and Y. Zhao, 2007 Ауксин, синтезируемый флавинмонооксигеназами YUCCA, необходим для эмбриогенеза и формирования листьев у Arabidopsis. Растительная клетка 19 : 2430–2439.
- ↵
Cheng, Y. F., and Y. D. Zhao, 2007 Роль ауксина в развитии цветков. J. Integr. Plant Biol. 49 : 99–104.
- ↵
Cheng, Y. F., X. H. Dai и Y. D. Zhao, 2006 Биосинтез ауксина флавинмонооксигеназами YUCCA контролирует образование органов цветков и сосудистых тканей у Arabidopsis. Genes Dev. 20 : 1790–1799.
- ↵
Christensen, S. K., N. Dagenais, J.Чори и Д. Вейгель, 2000 Регулирование ауксинового ответа протеинкиназой PINOID. Ячейка 100 : 469–478.
- ↵
Клиффорд, Х. Т., 1987 Морфология колосков и цветков, стр. 21–30 в издании Grass Systematics and Evolution , под редакцией Т. Р. Содерстрома, К. В. Хилу, К. С. Кэмпбелла и М. Э. Баркуорта. Smithsonion Institution Press, Вашингтон, округ Колумбия.
- ↵
Коу, Э. Х., М. Г. Нойффер и Д. А. Хойзингтон, 1988 г. Генетика кукурузы, стр.81–258 в Corn and Corn Improvement , под редакцией Дж. Ф. Спрэга и Дж. У. Дадли. ASA-CSSA-SSSA, Мэдисон, Висконсин.
- ↵
Doebley, J., A. Stec и L. Hubbard, 1997 Эволюция апикального доминирования у кукурузы. Природа 386 : 485–488.
- ↵
Gallavotti, A., Q. Zhao, J. Kyozuka, R. B. Meeley, M. Ritter et al. , 2004 Роль бесплодного стебля1 в архитектуре кукурузы. Природа 432 : 630–635.
- ↵
Galweiler, L., C.H. Guan, A. Muller, E. Wisman, K. Mendgen et al. , 1998 Регулирование полярного транспорта ауксина с помощью AtPIN1 в сосудистой ткани Arabidopsis. Наука 282 : 2226–2230.
- ↵
Geisler, M., H.U. Kolukisaoglu, R. Bouchard, K. Billion, J. Berger et al. , 2003 TWISTED DWARF1, уникальный иммунофилин-подобный белок, закрепленный на плазматической мембране, взаимодействует с переносчиками AtPGP1 и AtPGP19, подобными множественной лекарственной устойчивости Arabidopsis.Мол. Биол. Ячейка 14, : 4238–4249.
- ↵
Geldner, N., N. Anders, H. Wolters, J. Keicher, W. Kornberger et al. , 2003 Arabidopsis GNOM ARF-GEF обеспечивает рециклинг эндосом, транспорт ауксина и зависимый от ауксина рост растений. Мобильный 112 : 219–230.
- ↵
Gernart, W., 1912 Новый подвид Zea mays L. Am. Nat. 46 : 616–622.
- ↵
Гил, П., E. Dewey, J. Friml, Y. Zhao, K. C. Snowden et al. , 2001 BIG: калоссиноподобный белок, необходимый для полярного транспорта ауксина у Arabidopsis. Genes Dev. 15 : 1985–1997.
- ↵
Hardtke, C. S., W. Ckurshumova, D. P. Vidaurre, S. A. Singh, G. Stamatiou et al. , 2004 Перекрывающиеся и не повторяющиеся функции факторов ответа на ауксин Arabidopsis МОНОПТЕРОС и НЕФОТОТРОПНЫЙ ГИПОКОТИЛ 4. Развитие 131 : 1089–1100.
- ↵
Heisler, M. G., C. Ohno, P. Das, P. Sieber, G. V. Reddy et al. , 2005 Паттерны транспорта ауксина и экспрессии генов во время развития зачатков, выявленные с помощью живых изображений меристемы соцветий Arabidopsis. Curr. Биол. 15 : 1899–1911.
- ↵
Hubbard, L., P. McSteen, J. Doebley и S. Hake, 2002 Паттерны экспрессии и мутантный фенотип teosinte разветвленный1 коррелируют с подавлением роста кукурузы и теозинте.Генетика 162 : 1927–1935.
- ↵
Irish, E. E., 1996 Регулирование определения пола у кукурузы. BioEssays 18 : 363–369.
- ↵
Irish, E. E., 1997 Класс II семян метелки мутации свидетельствуют о множестве типов меристем соцветий у кукурузы (Poaceae). Являюсь. J. Bot. 84 : 1502–1515.
- ↵
Джексон Д., Б. Фейт и С. Хейк, 1994 Экспрессия генов гомеобокса кукурузы узловых1 в апикальной меристеме побега предсказывает паттерны морфогенеза вегетативного побега.Разработка 120 : 405–413.
- ↵
Келлог, Э. А., 2000 Травы: пример макроэволюции. Анну. Rev. Ecol. Syst. 31 : 217–238.
- ↵
Келлог, Э. А., 2007 Цветочные проявления: генетический контроль соцветий травы. Curr. Opin. Plant Biol. 10 : 26–31.
- ↵
Керстеттер, Р. А., Д. Лауденсия-Чингкуанко, Л. Г. Смит и С. Хейк, 1997 Мутации с потерей функции в гене гомеобокса кукурузы, узелковый1 , нарушают поддержание меристемы побегов.Разработка 124 : 3045–3054.
- ↵
Lee, S. H., and H. T. Cho, 2006 PINOID положительно регулирует отток ауксина в волосковых клетках корня арабидопсиса и клетках табака. Растительная клетка 18 : 1604–1616.
- ↵
- ↵
McSteen, P., 2006 Ветвление: путь ramosa и эволюция морфологии соцветий травы. Растительная клетка 18 : 518–522.
- ↵
Макстин, П., and S. Hake, 1998 Генетический контроль развития растений. Curr. Opin. Biotechnol. 9 : 189–195.
- ↵
McSteen, P., and S. Hake, 2001 бесплодное соцветие2 регулирует развитие пазушных меристем в соцветиях кукурузы. Разработка 128 : 2881–2891.
- ↵
Макстин П. и О. Лейзер, 2005 г. Ветвление побегов. Анну. Rev. Plant Biol. 56 : 353–374.
- ↵
Макстин, П., Д. Лауденсия-Чингкуанко и Дж. Коласанти, 2000 Цветочек под любым другим названием: контроль идентичности меристемы у кукурузы. Trends Plant Sci. 5 : 61–66.
- ↵
McSteen, P., S. Malcomber, A. Skirpan, C. Lunde, X. Wu et al. , 2007 бесплодное соцветие2 кодирует коортолог серин / треонинкиназы PINOID и необходим для органогенеза во время соцветия и вегетативного развития кукурузы. Plant Physiol. 144 : 1000–1011.
- ↵
Мултани, Д. С., С. П. Бриггс, М. А. Чемберлин, Дж. Дж. Блейксли, А. С. Мерфи и др. , 2003 Утрата переносчика МЛУ в компактных стеблях мутантов кукурузы br2 и сорго dw3 . Наука 302 : 81–84.
- ↵
Neuffer, M. G., and K. A. Sheridan, 1977 Доминантные мутанты от пыльцы, обработанной EMS. Maize Newsl. 51 : 59–60.
- ↵
Нойффер, М.Г., Э. Х. Коу и С. Р. Весслер, 1997 Мутанты кукурузы. Лабораторный пресс Колд-Спринг-Харбор, Плейнвью, Нью-Йорк.
- ↵
Но, Б., А. С. Мерфи и Э. П. Сполдинг, 2001 Гены Arabidopsis, подобные множественной лекарственной устойчивости, необходимые для транспорта ауксина и опосредованного ауксином развития. Растительная клетка 13 : 2441–2454.
- ↵
Окада, К., Дж. Уэда, М. К. Комаки, К. Дж. Белл и Ю. Шимура, 1991 Потребность в системе полярного транспорта ауксина на ранних стадиях формирования цветочных бутонов Arabidopsis.Растительная клетка 3 : 677–684.
- ↵
Przemeck, G. K. H., J. Mattsson, C. S. Hardtke, Z. R. Sung и T. Berleth, 1996 г. Исследования роли гена Arabidopsis MONOPTEROS в развитии сосудов и аксиализации растительных клеток. Planta 200 : 229–237.
- ↵
Reinhardt, D., T. Mandel и C. Kuhlemeier, 2000 Ауксин регулирует инициацию и радиальное положение боковых органов растения. Растительная клетка 12 : 507–518.
- ↵
Reinhardt, D., E. R. Pesce, P. Stieger, T. Mandel, K. Baltensperger et al. , 2003 Регулирование филлотаксиса с помощью полярного транспорта ауксина. Природа 426 : 255–260.
- ↵
Риттер, М. К., К. М. Падилла и Р. Дж. Шмидт, 2002 г. Мутант кукурузы бесплодный стебель1 является дефектным в развитии подмышечной меристемы. Являюсь. J. Bot. 89 : 203–210.
- ↵
Salvi, S., R. Tuberosa, E. Chiapparino, M. Maccaferri, S. Veillet et al. , 2002 К позиционному клонированию Vgt1 , QTL, контролирующего переход от вегетативной фазы к репродуктивной у кукурузы. Завод Мол. Биол. 48 : 601–613.
- ↵
Сато-Нагасава, Н., Н. Нагасава, С. Малкомбер, Х. Сакаи и Д. Джексон, 2006 г. Фермент метаболизма трегалозы контролирует структуру соцветий кукурузы. Природа 441 : 227–230.
- ↵
Скэнлон, М.J., 2003 Полярный ингибитор транспорта ауксина N-1-нафтилфталаминовая кислота нарушает зарождение листьев, регуляцию белка KNOX и образование краев листьев у кукурузы. Plant Physiol. 133 : 597–605.
- ↵
Шеридан, В. Ф., 1988 Генетика развития кукурузы: гены морфогенеза. Анну. Преподобный Жене. 22 : 353–385.
- ↵
Зибурт, Л. Э., Г. К. Мудай, Э. Дж. Кинг, Г. Бентон, С. Ким и др. , 2006 SCARFACE кодирует ARF-GAP, который необходим для нормального оттока ауксина и формирования рисунка вены у Arabidopsis.Растительная клетка 18 : 1396–1411.
- ↵
Стивз Т. и И. Сассекс, 1989 Паттерны в развитии растений. Cambridge University Press, Кембридж, Великобритания.
- ↵
Taramino, G., M. Sauer, J. L. Stauffer, D. Multani, X. Niu et al. , 2007 Кукуруза ( Zea mays L.) Ген RTCS кодирует белок LOB-домена, который является ключевым регулятором зародышевого семенного и постэмбрионального зародышевых корней.Плант Дж. 50 : 649–659.
- ↵
Вейт Б., 2006 Сети передачи сигналов стволовых клеток в растениях. Завод Мол. Биол. 60 : 793–810.
- ↵
Вейт Б., Р. Дж. Шмидт, С. Хейк и М. Ф. Янофски, 1993 Развитие цветков кукурузы — новые гены и старые мутанты. Растительная клетка 5 : 1205–1215.
- ↵
Vernoux, T., J. Kronenberger, O. Grandjean, P. Laufs and J. Traas, 2000 PIN-FORMED 1 регулирует судьбу клеток на периферии апикальной меристемы побега.Разработка 127 : 5157–5165.
- ↵
Vollbrecht, E., L. Reiser and S. Hake, 2000 Размер меристемы побега зависит от инбредного фона и присутствия гена гомеобокса кукурузы, узловатый1. Разработка 127 : 3161–3172.
- ↵
Фолльбрехт, Э., П. С. Спрингер, Л. Гох, Э. С. Баклер, И. В. и Р. Мартиенсен, 2005 Архитектура систем цветочных ветвей кукурузы и родственных трав. Природа 436 : 1119–1126.
- ↵
Wang, H., T. Nussbaum-Wagler, B. Li, Q. Zhao, Y. Vigouroux et al.
