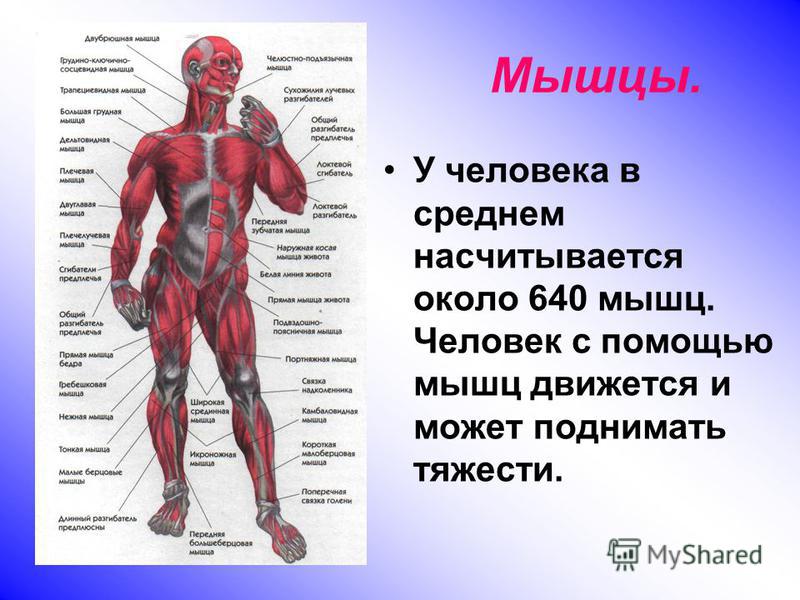
Скелетная мышца как орган
Дано определение органа. Выделены характерные особенности органа: целостность, своеобразная форма, размеры и положение, совокупность различных клеток и тканей, специфическая функция. На основе этих признаков скелетная мышца рассмотрена как орган. Описаны основная и вспомогательные функции скелетных мышц.
Скелетная мышца как орган
Что такое орган?
Прежде чем разбираться, что представляет собой скелетная мышца как орган, давайте поймем, что такое орган.
Слово орган происходит от древнегреческого слова «ὄργανον», что означает орудие или инструмент.
Википедия определяет орган как обособленную совокупность различных типов клеток и тканей, выполняющую определённую функцию в пределах живого организма.
М.Ф. Иваницкий (1985) указывает на следующие характерные признаки: «…орган, как компонент системы, анатомически и функционально обособлен от соседних образований. Органом называют часть тела, которая в процессе развития вида и особи приобрела своеобразие положения, формы, размеров, внутреннего строения, функций и взаимодействует с другими органами. Орган – это целостная конструкция, состоящая из различных тканей и подразделяющаяся на более мелкие части. Эти анатомические образования включают в себя структурно-функциональные единицы органа».
Орган – это целостная конструкция, состоящая из различных тканей и подразделяющаяся на более мелкие части. Эти анатомические образования включают в себя структурно-функциональные единицы органа».
Есть еще одно определение скелетной мышцы как органа, которое мне нравится.
«Скелетная мышца – орган, имеющий определенный источник развития, характерную форму и строение, расположение, источники кровоснабжения и иннервации, пути лимфооттока, выполняющий определенную функцию» (https://www.bsmu.by/downloads/kafedri/k_anatomia/stud/2017-2/mr9.pdf обращение 4.06.2020).
Из этих определений можно выделить следующие характерные признаки органа:
- Орган обособлен от соседних образований. Его характеризует целостность, своеобразная форма, размеры и положение.
- Орган может состоять из различных клеток и тканей.
- Чаще всего орган состоит из структурно-функциональных единиц.
- У органа всегда имеется специфическая, только ему присущая функция.
Следует отметить, что в организме человека много органов.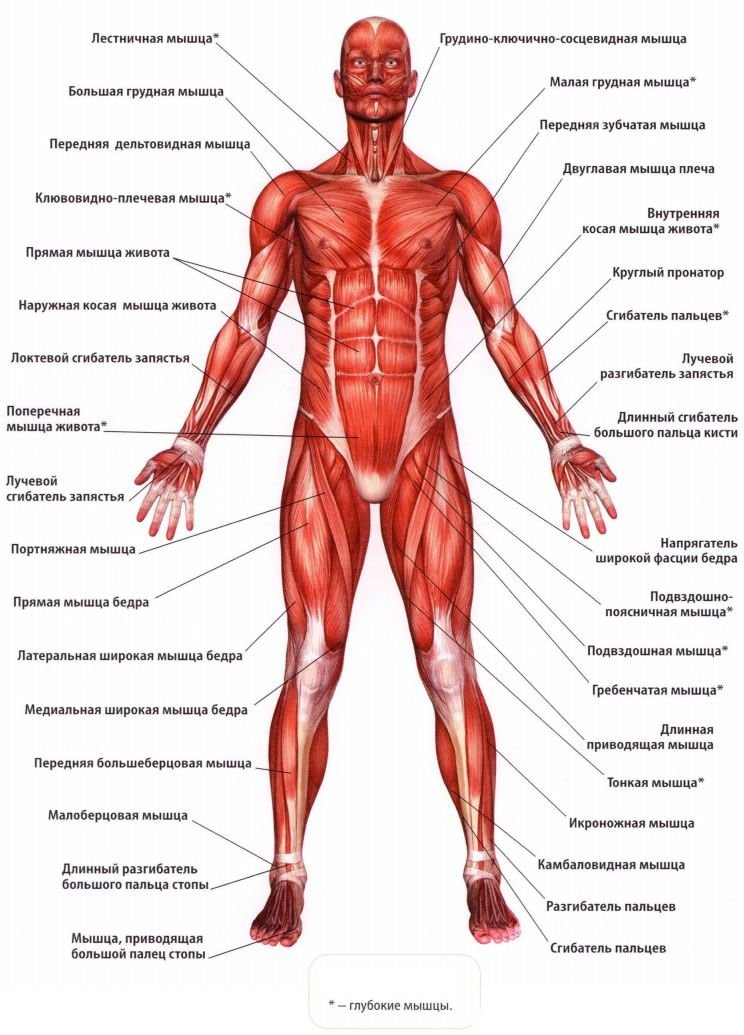 Например, различают внутренние органы: сердце, печень, почки и т.д. И везде мы видим обособленную совокупность различных типов клеток и тканей, которая выполняет определенную функцию в пределах живого организма.
Например, различают внутренние органы: сердце, печень, почки и т.д. И везде мы видим обособленную совокупность различных типов клеток и тканей, которая выполняет определенную функцию в пределах живого организма.
Теперь рассмотрим скелетную мышцу как орган.
Обособленность и целостность мышцы
Скелетная мышца – это орган, который обособлен от других мышц и элементов опорно-двигательного аппарата человека за счет того, что снаружи каждая мышца окружена оболочками, которые отделяют одну мышцу от других мышц.
Форма мышцы
Скелетные мышцы имеют брюшко и сухожильные концы, за счет которых мышца прикрепляется к костям или другим образованиям. Бывают и другие способы прикрепления мышц. Проксимальное сухожилие или проксимальная часть мышцы, связанная с костью, называется головкой и является началом мышцы. Дистальное сухожилие или дистальный конец мышцы, прикрепляющийся к другой кости, называется хвостом; это место принято называть прикреплением мышцы. Форма скелетных мышц разнообразна. Различают веретенообразные, прямые, круглые, квадратные, дельтовидные, трапециевидные и.т.д.
Различают веретенообразные, прямые, круглые, квадратные, дельтовидные, трапециевидные и.т.д.
Размеры мышц
Скелетные мышцы имеют различные размеры. Они могут быть очень маленькими, как например, мышцы, обеспечивающие перемещение глазного яблока и изменение толщины хрусталика. А бывают очень большие мышцы, например четырехглавая мышца бедра или ягодичные мышцы. Основными показателями, характеризующими размеры мышцы являются: объем, площадь поперечного сечения и длина мышцы. Увеличение объема скелетных мышц называется гипертрофией.
Положение мышц
Особенностью прикрепления скелетных мышц является то, что они начинаются на одной кости, а прикрепляются к другой. Благодаря этому скелетные мышцы обеспечивают движения и локомоцию человека, а также сохранение положения тела.
Совокупность различных клеток и тканей
Скелетная мышца представляет собой совокупность различных клеток и тканей. Составляющими скелетной мышцы являются: поперечно-полосатая мышечная ткань, рыхлая и плотная соединительные ткани, а также нервная ткань. Лимфатические и кровеносные сосуды состоят из соединительной ткани, гладкой мышечной ткани и эпителия. Мышечная ткань формирует основную часть мышцы – её брюшко, рыхлая соединительная ткань образует мягкий скелет мышцы, а плотная – сухожилия.
Структурно-функциональная единица мышцы
Структурно-функциональной единицей скелетной мышцы является мышечное волокно. В скелетных мышцах человека насчитываются сотни тысяч мышечных волокон. В некоторых мышцах (икроножной) количество мышечных волокон достигает одного миллиона.
Состав мышцы
Мышечное волокно, группы мышечных волокон и вся мышца в целом окружены соединительно-тканными оболочками различной плотности. Плотная соединительная ткань, покрывающая всю мышцу или группы мышц, называется фасцией.
Мышечные волокна соединяются с сухожилием, которое прикрепляется к кости. Мышечные волокна могут также напрямую прикрепляться к кости. Сухожилия у различных мышц неодинаковы. У мышц конечностей в основном наблюдаются узкие и длинные сухожилия.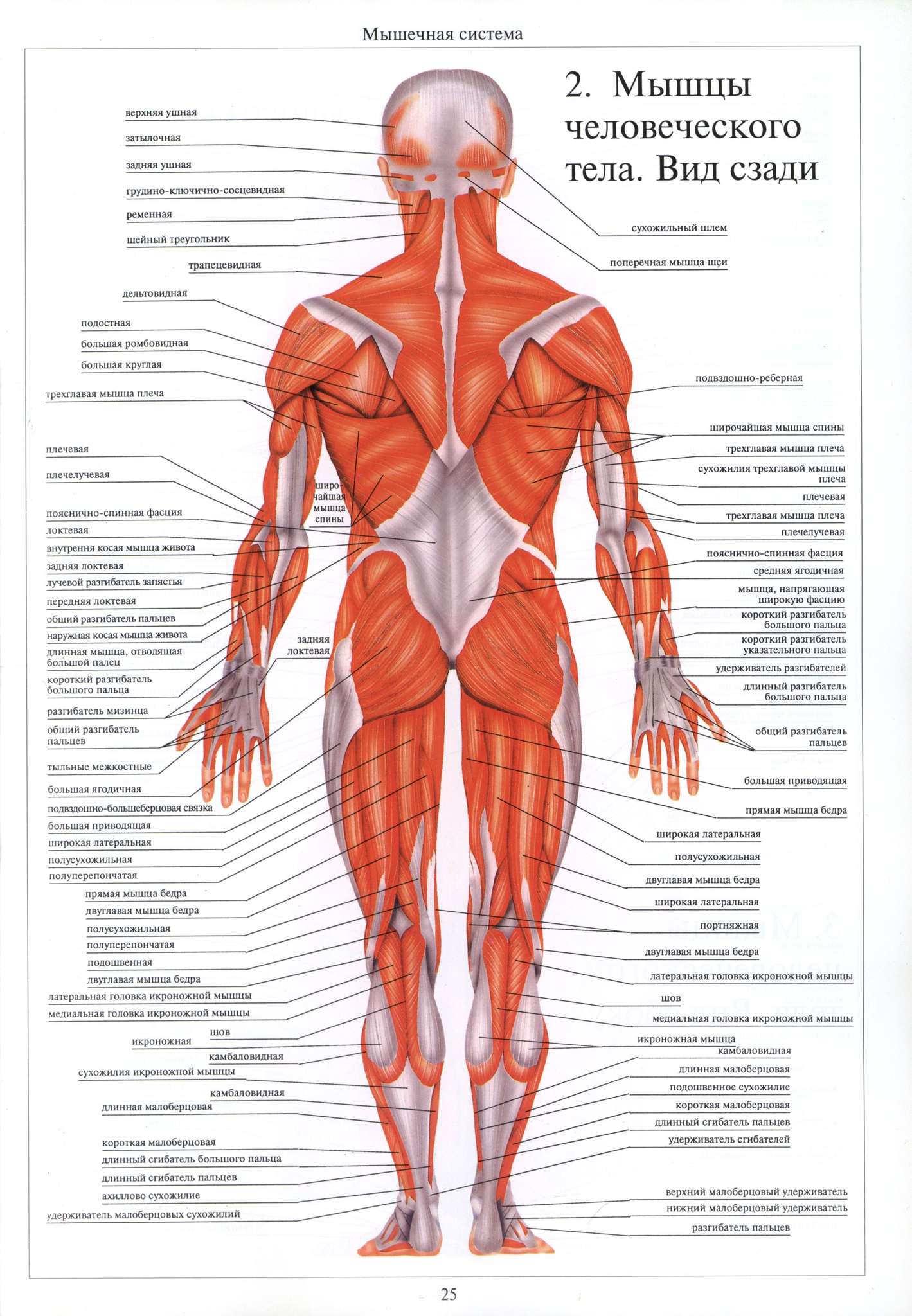 У мышц, участвующих в формировании стенок брюшной полости имеется широкое плоское сухожилие, которое называется апоневрозом.
У мышц, участвующих в формировании стенок брюшной полости имеется широкое плоское сухожилие, которое называется апоневрозом.
Иннервация мышцы осуществляется двигательными, чувствительными и вегетативными нервами. Также мышца снабжена кровеносными и лимфатическими сосудами. В мышце имеются рецепторы, реагирующие на изменение длины, скорости и напряжения мышцы. Внутренней средой мышцы является тканевая жидкость, которая по составу похожа на лимфу.
Более подробно строение и функции мышц описаны в моих книгах
Функции скелетных мышц
Основная (специфическая) функция скелетных мышц
Под воздействием нервных импульсов скелетные мышцы сокращаются (развивают напряжение). Благодаря этому скелетные мышцы приводят в движение кости (части тела) друг относительно друга или наоборот, обеспечивают их неподвижность. Это обеспечивает передвижение тела в пространстве (ходьба, бег, прыжки и т. д.), выполнение разнообразных манипуляций (работа), сохранение равновесия тела.
Вспомогательные функции
Кроме основной, скелетные мышцы выполняют ряд вспомогательных функций:
- Скелетные мышцы участвуют в выполнении жизненно важных функций организма человека, таких как дыхание, глотание, зрительная функция.
- Скелетные мышцы обеспечивают различные физиологические отправления (роды, мочеиспускание, дефекацию).
- Скелетные мышцы стабилизируют суставы, а также своды стопы.
- При сокращении скелетных мышц облегчается ток крови по венам и лимфы по лимфатическим сосудам. В этом случае скелетные мышцы действуют в качестве «насоса».
- Скелетные мышцы обладают вязкостью. Вязкость мышцы возникает из-за трения мышечных волокон друг о друга, а также мышечных волокон о соединительно-тканные оболочки. Поэтому при сокращении скелетные мышцы нагреваются, что способствует увеличению теплопродукции организмом человека.
- Скелетные мышцы участвуют в образовании стенок полостей тела, например, брюшной полости.
Литература
Иваницкий М. Ф. Анатомия человека: Учебник для ин-тов физ. культ.– М. Физкультура и спорт, 1985.- 544 с.
Ф. Анатомия человека: Учебник для ин-тов физ. культ.– М. Физкультура и спорт, 1985.- 544 с.
Самсонова А.В. Гипертрофия скелетных мышц человека: Учебное пособие.- 5-е изд. — СПб.: Кинетика, 2018.- 159 с.
С уважением, А.В. Самсонова
Урок 13. Строение мышц человека
Строение мышц человека
Урок биологии человека –
8 класс
Разработан преподавателем биологии: Колесниковой Л.А.
Мышечная ткань
- Мышечная ткань составляет основную массу мышц и осуществляет их сократительную функцию. Сократимые элементы развиваются из нескольких источников и имеют различное строение.
- Выделяют гладкую и поперечно-полосатую мышечную ткань, которая, в свою очередь, подразделяется на скелетную и сердечную .
Функция мышечной ткани:
сократительная
Мышечная ткань
Поперечно-полосатая
скелетная мышца
Поперечнополосатая
сердечная мышца
Гладкая мышечная ткань
Мышечная ткань
Скелетные мышцы образованы поперечнополосатой мышечной тканью, волокна которой собраны в пучки.
Поперечно-полосатая
скелетная мышца
Мышечная ткань
Сердечная мышца, как и скелетная, состоит из поперечнополосатых мышечных волокон. Эти волокна в определённых участках как бы сливаются (переплетаются). Поперечнополосатая сердечная мышца, не подчиняется нашей воле и работает не переставая от первого в жизни сокращения до последнего.
Поперечнополосатая
сердечная мышца
Мышечная ткань
Стенки внутренних органов (сосудов, кишечника, мочевого пузыря) образованы гладкой мышечной тканью.
Сокращения гладкой (неисчерченной)
мышечной ткани происходят произвольно, медленно.
Гладкая мышечная ткань
Мышечный корсет
Мышцы – активная часть опорно-двигательной системы, обеспечивающая все многообразие движений, совершаемых в организме человека. Благодаря мышцам тело сохраняет равновесие, перемещается в пространстве, осуществляются дыхательные движения грудной клеткой и диафрагмой, глотание, образуется голос, осуществляются движения глаз, работа внутренних органов, в том числе и сердца.
Благодаря мышцам тело сохраняет равновесие, перемещается в пространстве, осуществляются дыхательные движения грудной клеткой и диафрагмой, глотание, образуется голос, осуществляются движения глаз, работа внутренних органов, в том числе и сердца.
Мышца – musculus – мышь!
Строение мышц
Мышца – орган, состоящий из мышечной ткани, плотной соединительной ткани, кровеносных сосудов и нервов, и выполняющий функцию сокращения.
- Скелетные мышцы состоят из пучков поперечнополосатых мышечных волокон.
- Каждый мышечный пучок покрывает соединительнотканная пленка, а всю мышцу – фасция.
Строение мышц
- Брюшко состоит из множества пучков поперечнополосатых мышечных волокон, покрытых соединительнотканными оболочками.
- Сухожилия обеспечивают соединение с костями, состоят из плотной соединительной ткани.
Мышцы – органы тела, состоящие из мышечной ткани, способной сокращаться под влиянием нервных импульсов.
В организме человека около 600 мышц.
- В мышцах расположены:
- — кровеносные сосуды – кровь обеспечивает поступление О2 и питательных веществ, уносит продукты распада
- — лимфатические сосуды – способствуют дополнительному оттоку жидкости от мышц
- — нервные окончания:
- рецепторы – воспринимают степень растяжения и сокращения мышц
- эффекторы – получают команды из ЦНС
Классификация мышц
- Сгибатели и разгибатели
- Приводящие и отводящие
- Супинаторы и пронаторы
- Вдоха и выдоха
- Жевательные и мимические
Мышцы, совершающие одинаковые движения – синергисты .
Мышцы, совершающие противоположные движения – антагонисты .
Классификация мышц
2. По расположению
По расположению
- Мышцы головы
- Мышцы шеи
- Мышцы туловища
- Мышцы конечностей
- Мышечная координация – согласованная работа мышц.
- Синергисты – мышцы, выполняющие одно и то же движение.
- Антагонисты – мышцы, выполняющие противоположные действия.
- У плачущего ребенка работает 43 мышцы, а у смеющегося – 17 = смеяться энергетически выгодней.
- Свойства мышц:
- возбудимость – способность реагировать на нервные импульсы – раздражители
- сократимость – способность уменьшать длину при увеличении толщины
- растяжимость – способность увеличивать длину при уменьшении толщины
- эластичность – способность принимать прежнее положение после растяжения
- Наиболее эффективно мышцы работают в 1330.

- КПД мышц 20%. 80% — тепловые потери.
Практическая работа
- Используя рисунки учебника и анатомическое описание, определите местоположение мышечных групп и выполняемые ими движения. Заполните таблицу:
Расположение мышц
Название мышцы или мышечной группы
Мышцы головы
Функции мышцы или мышечной группы
Мышцы шеи
Мышцы туловища спереди
Мышцы туловища сзади
Мышцы руки
Мышцы ноги
Мышцы или группы мышц и их функции
- Мимические мышцы — мышцы, одним концом прикрепляющиеся к костям черепа, а другим – к коже или только к коже.
- Жевательные — мышцы, приводящие в движение нижнюю челюсть.

- Трапециевидная – мышца шеи и спины, откидывающая назад голову.
- Кивательные (грудино-ключично-сосцевидные) – мышцы, наклоняющие голову вперед, когда сокращаются вместе, и наклоняющие голову в стороны, когда сокращаются поодиночке.
- Широчайшая — мышца спины, отводящая руки за спину.
Мышцы или группы мышц и их функции
- Грудные (большие и малые) – мышцы груди, сводящие плечи и руки, согнутые в локте, вперед.
- Межреберные – мышцы, обеспечивающие расширение и спадение грудной клетки
- Пресс – мышцы, поднимающие туловище из положения «лежа» в положение «сидя» при фиксированных ногах.
- Ягодичные, глубокие мышцы спины — мышцы, обеспечивающие прямохождение.
- Дельтовидная – мышца руки, отводящая плечо в сторону.
- Предплечье – мышцы, сгибающие пальцы и кисть в кулак.

Мышцы или группы мышц и их функции
- Скелетные мышцы приводят в движение кости относительно друг друга в составах, кроме того, они участвуют в образовании стенок брюшной и грудной полостей, таза. Входят в состав стеной верхней части пищевода и гортани. Осуществляют движение глазного яблока, дыхательные и глотательные движения.
- Мимические мышцы – мышцы лица, не связаны с суставами, выражают эмоции человека.
Закрепление изученного материала
- По предложенной схеме дайте объяснения классификации мышц человека.
Мышцы
Поперечно — полосатые
Гладкие
(мышцы внутренних органов)
Сердечная
Скелетные
(автоматизм)
Медленно сокращаются, мало интенсивен обмен в-в, большая растяжимость, могут находиться в состоянии длительного сокращения.
Быстро сокращаются, интенсивный обмен веществ
Дайте обоснованный ответ!
- Жевательные мышцы – самые сильные (усилие 70 кг).
- Самая крепкая связка – бертиниева, укрепляющая тазобедренный сустав (выдерживает 350 кг.)
- 3) Нужны ли эти свойства мышц и связок человеку?
- 1) Почему мышцы к костям прикрепляются под некоторым углом?
- 2) Почему в местах прикрепления мышц образуются шероховатости и бугристость?
Дайте обоснованный ответ!
4) Почему у тренированных людей рельеф костей более выражен?
Домашнее задание:
- §13 прочитать.
- Повторить § 8, статью «Нервная ткань», и § 9 «Нервная регуляция».
- По желанию: подготовить материал о работах И.М. Сеченова в области гигиены труда.
Иван Михайлович Сеченов
(1829 – 1905)
Всего доброго!
Основы рисунка — А.
 Ф. Шембель
Ф. ШембельАнатомическая характеристика мышц головы человека
Форма всей головы, ее внешний вид обусловливаются не только костным скелетом, т.е. черепом, но и залегающими под кожным покровом так называемыми мышцами.
Мышцы (мускулы) находятся поверх костей, прикрепляясь к ним, кроме тех, которые начинаются от кожи и соединяются с ней. Мышцы с помощью сухожилий перекидываются через суставы, соединяя кости, например, как это наблюдается у лобной и затылочной костей.
В отличие от черепа, где много неподвижных костей, мышцы относятся к активным органам движения. Являясь органом движения, мышца имеет остов, именуемый стромой и функционирующие элементы под названием мышечные волокна. Строма — это рыхлая соединительная ткань, связывающая мышечные волокна в отдельные пучки, покрытые оболочкой, которую называют фасцией. Роль фасции — не давать мышце смещаться в сторону, отделить ее от других мускулов. Следовательно, каждая мышца — целостный орган, наделенный способностью сокращаться и в связи с этим приводить в движение ту или иную часть головы: открывать и закрывать рот, говорить, расширять или сужать ноздри, моргать глазом и т.д.
Естественно, что движительная способность мышц регулируется нервной системой организма.
Любая мышца состоит из двух частей. Одна часть — брюшко — наделена природным свойством сокращаться и расслабляться, в то время как другая — сухожилие — просто передает действие мышцы. Внутренняя структура мускула схематически выглядит так: мышечные волокна крепятся под определенным углом к сухожильным пластинам, ввиду чего образуется перистое строение мышцы (рис. 53).
53).
Рис. 53
Разнообразная форма принадлежит всем мышцам человека. Она бывает веретенообразной, перистой и двуперистой, широкой, длинной с параллельными волокнами, короткой, веерообразной, двуглавой и двубрюшной. Форма мышцы связана с функцией.
Мышечный покров головы человека принято рассматривать с двух позиций: как жевательную систему и как мимическую.
Жевательные мышцы имеют объемную мускульную часть, образующую при сокращении хорошо различаемый рельеф. Свое название они получили благодаря естественной потребности живого организма в пище, но не только этим ограничивается их функция. Разговорная речь невозможна без этих мышц, некоторое участие жевательные мускулы принимают в мимических движениях человеческого лица.
К жевательным мышцам относятся: собственно жевательная мышца, височная мышца (рис. 54). Они расположены в области головы, лежат поверхностно, их сокращения прослеживаются под кожным покровом.
Рис. 54
54
По анатомическому строению собственно жевательная мышца (парная) начинается от скуловой дуги и заканчивается у угла нижнечелюстной кости. Рельефность этих двух мускулов заметна при приеме пищи. Участие мышц в мимике лица проявляется, когда человек находится в состоянии сильнейшего напряжения, связанного с физической болью, эмоциональным переживанием — горем, несчастьем, неприятностью, гневом, ссорой, угрозой, отвращением и т.п.
Свое начало височная мышца (парная) берет от височной впадины, широко пролегает по ней и внизу переходит в узкое сухожилие, продолженное между стенкой черепа и скуловой дугой до венечного отростка нижнечелюстной кости, где и прикрепляется. По форме височная мышца веерообразная. Она не так рельефна, как жевательная мышца, но признаки ее участия в жевании и мимических проявлениях можно обнаружить. Малорельефность височной мышцы влияет на резкость выступов височной линии и скуловой дуги у худых людей.
Есть и другие жевательные мышцы, но так как они лежат в глубине и не влияют на пластическую анатомию лица, то нами не рассматриваются. Эти мышцы, например крыловидные, участвуют в движениях нижней челюсти и сочетаются с действиями собственно жевательной и височной.
Эти мышцы, например крыловидные, участвуют в движениях нижней челюсти и сочетаются с действиями собственно жевательной и височной.
Мимические мышцы подразделяются на три группы. Подразделение их связано с областью расположения на лицевой части головы и естественными связями и взаимовлияниями.
К верхней группе мимических мышц головы человека относятся: лобная мышца (лобное брюшко), сухожильный шлем (черепной апоневроз, т.е. расширенная пластина плоского сухожилия, несущего функцию перенесения силы мышц на место воздействия), затылочная мышца (затылочное брюшко), мышца гордецов (пирамидальный мускул), мышца — сморщиватель брови (мускул боли), круговая мышца глаза, мышца — подниматель верхнего века.
Лобная мышца (парная) имеет вертикальность мышечных волокон, прикрепляется вверху к сухожильному шлему, внизу — к коже у бровей (рис. 55). Сокращаясь, данная мышца собирает кожу лба, поднимая брови, что ведет к образованию поперечных кожных складок, исчезающих при спокойном состоянии людей.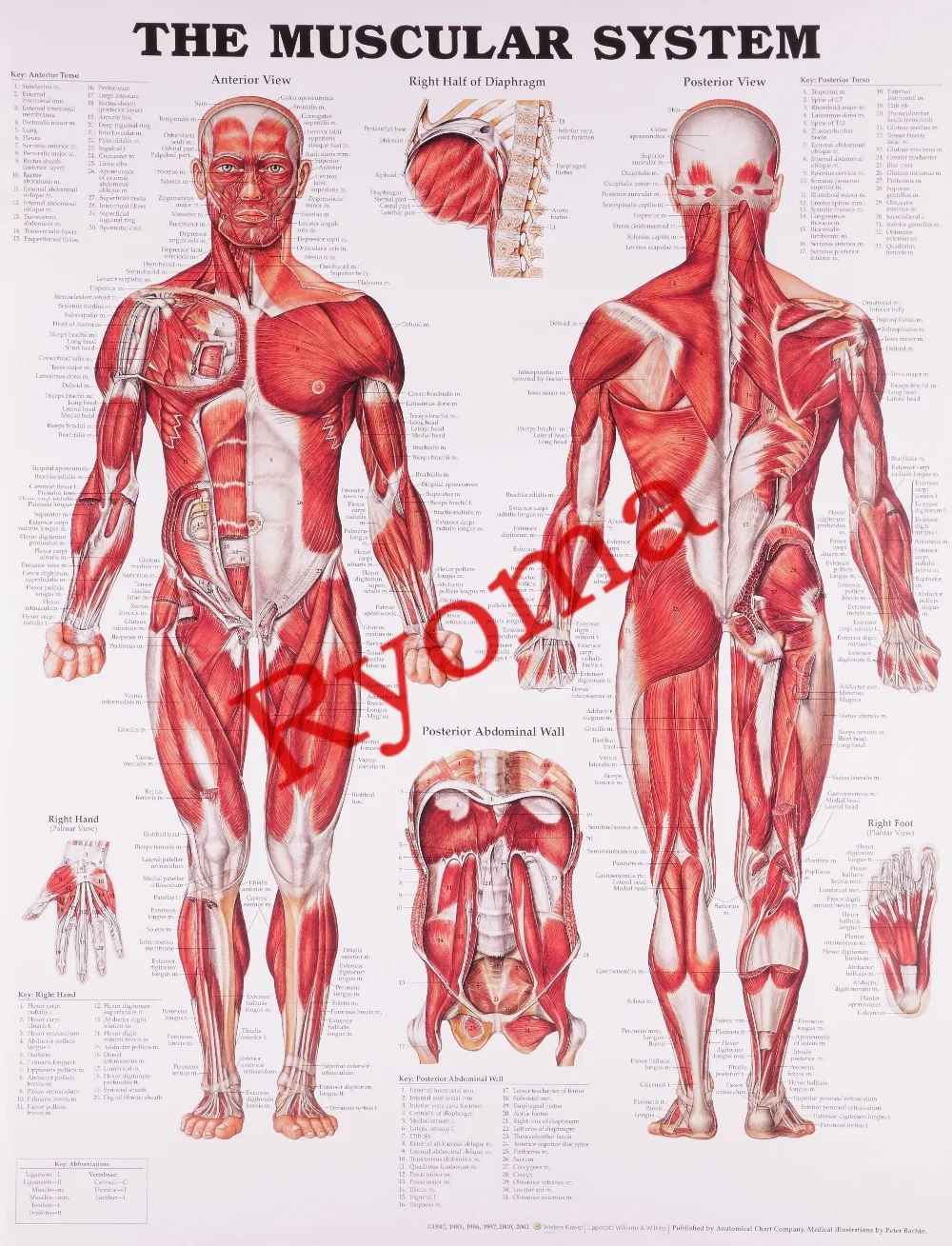 Сокращение лобной мышцы наблюдается у человека при выражении внимания или удивления. Живописцы, скульпторы досконально знают мимические мышцы головы, иначе им бы не удавалось передавать в своих произведениях всевозможные состояния души человека.
Сокращение лобной мышцы наблюдается у человека при выражении внимания или удивления. Живописцы, скульпторы досконально знают мимические мышцы головы, иначе им бы не удавалось передавать в своих произведениях всевозможные состояния души человека.
Соединяющее лобное брюшко с затылочным сухожилием покрывает почти всю верхнюю часть черепа и названо поэтому сухожильным шлемом. Он соединен с теменной костью. Затылочная мышца «вынуждена» прикрепляться к затылочной кости, ибо больше негде. Лобное и затылочное брюшки не могут, кроме того, сокращаться одновременно, хотя есть люди, сумевшие при надлежащей тренировке добиться изолированного сокращения каждой из мышц.
Рис. 55
Лобная мышца продолжается с каждой стороны до спинки носа и переходит в отдельную небольшую парную мышцу — мышцу гордецов, получившую свое название, вероятно, из-за того, что является антагонистом (соперницей) лобной, имеющей свойство поднимать кожу лба, в то время как пирамидальная (так еще называют мышцу гордецов) — опускать. При сокращении пирамидальной мышцы образуется поперечная складка у корня носа. Мышца прикрепляется внизу к носовой косточке, а вверху к коже на внутреннем конце брови.
При сокращении пирамидальной мышцы образуется поперечная складка у корня носа. Мышца прикрепляется внизу к носовой косточке, а вверху к коже на внутреннем конце брови.
Надпереносье является местом, от которого начинаются и тянутся вправо и влево под кожей бровей до их середин, где крепятся к коже, так называемые мускулы боли (их два), иначе именуемые мышцы-сморщиватели бровей. При сокращении мышц сближаются брови, перегибаются под углом, образуются крупные складки, идущие вертикально. Мышцы активно функционируют при мучительных воспоминаниях, печали, обиде, боли и т.д.
Природа создала глазницу, в которой располагается чудеснейший орган — глазное яблоко. Снаружи глаза располагается его круговая мышца. Она состоит из двух основных частей, одна из которых носит название глазничной, а другая — вековой. Первая занимает всю окружность глазницы, заходя за се края, вторая находится на верхнем и нижнем веках, откуда и произошло название. Мышцы глаза способствуют защите органа зрения от внешних влияний, самостоятельно сокращаясь, играют большую роль в мимике лица человека.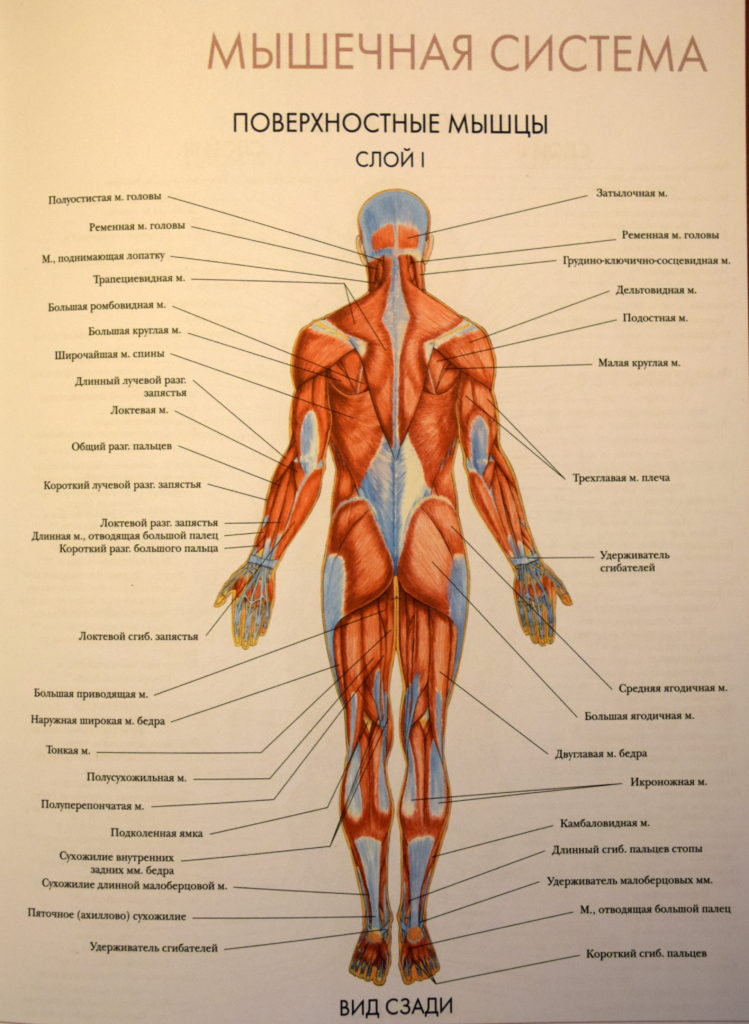 Недаром существует выражение «глаза — зеркало души».
Недаром существует выражение «глаза — зеркало души».
Выражение всего лица человека зависит от того, какое эмоциональное состояние захватило его.
У глазного яблока есть своя мышца, специально предназначенная для большой подвижности верхнего века. Она называется мышцей — поднимателем верхнего века (рис. 56).
Рис. 56
Следующей группой мимических мышц занята область ротового отверстия и ноздрей: круговая мышца рта, большая скуловая мышца (мышца смеха), малая скуловая мышцы (мышца плача), мышца подниматель верхней губы и крыла носа, носовая мышца, мышца опускатсль перегородки носа, мышца подниматель угла рта, мышца опускатель нижней губы, мышца опускатель угла рта, подбородочная мышца и щечная мышца (рис. 57).
Непарная мышца, залегающая в коже в виде концентрических волокон вокруг отверстия рта, носит название круговой мышцы рта. Подразделяясь на две части — внутреннюю и наружную, эта мышца предназначена при сокращении закрывать ротовое отверстие.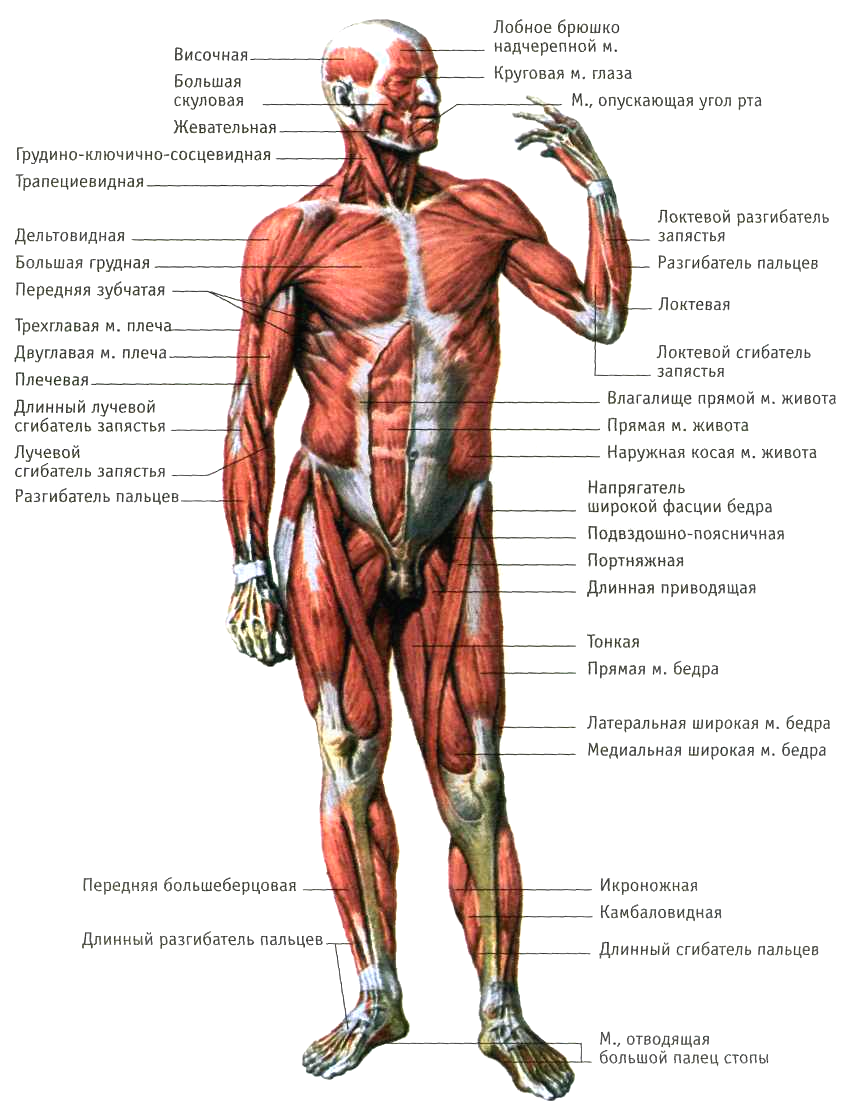 Наружная часть мышцы — губная. Внутренняя залегает под розовой каймой губ. Короче говоря, роль круговой мышцы рта такова, что не будь ее, рот постоянно был бы открыт.
Наружная часть мышцы — губная. Внутренняя залегает под розовой каймой губ. Короче говоря, роль круговой мышцы рта такова, что не будь ее, рот постоянно был бы открыт.
Кожа угла рта заканчивает протяжение начинающейся от скуловой кости большой скуловой мышцы, идущей вниз и вперед и немного вплетающейся в круговую мышцу рта. Ее еще называют мышцей смеха, так как наиболее заметно сокращение этой мышцы при смехе. Эта мышца парная, поэтому сокращаются они одновременно (улыбка появляется при легком сокращении; при этом углы рта поднимаются, а нижние концы носогубных складок слегка изгибаются), во время смеха особенно сильно. Рот при улыбке приоткрыт, при смехе вовсе растягивается.
Рис. 57
Рис. 58
От скуловой кости берет свое начало еще одна мышца, тоже парная, которая идет вниз и внутрь. Она названа малой скуловой и расположена почти параллельно большой скуловой мышце. Сильное сокращение малой скуловой мышцы проявляется при плаче, отчего ее называют мышцей плача (рис. 58).
58).
Верхнюю губу поднимает мышца, прикрепляющаяся вверху к краю глазной впадины, а внизу — к крылу носа и к поднимаемой им губе.
Перегородку носа опускает мышца, находящаяся рядом со своей парой. Ее начало — от альвеолярного отростка верхнечелюстной кости (над средним зубом-резцом), завершение — в перегородке носа.
Носовая мышца имеет две части — поперечную и крыльную, начинается от верхней челюсти в месте, находящемся над наружным зубом-резцом и над зубом-клыком. Поперечная часть носовой мышцы устремляется вверх, огибает крыло носа и прикрепляется к сухожилию, покрывающему хрящевую часть. С внутренней стороны поперечной части носовой мышцы поднимается кверху крыльная, которая затем прикрепляется к крылу носа. Сокращение обеих носовых мышц приводит к тому, что на коже носа образуются продольные складки, растягиваются ноздри.
Передняя поверхность нижнечелюстной кости у основания зубов служит началом спускающейся вниз и вплетающейся в кожу подбородка так называемой подбородочной мышцы (парная).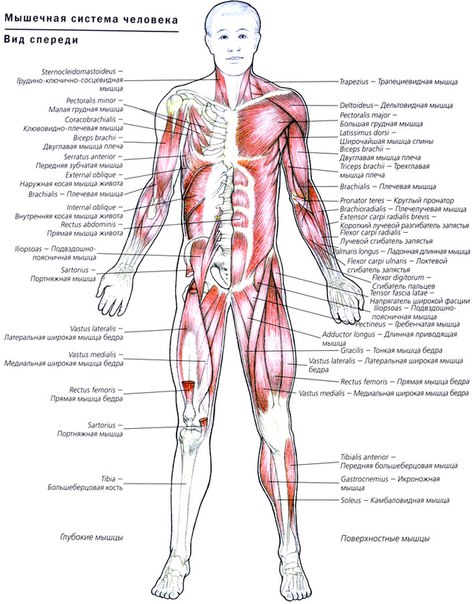
Все остальные мимические мышцы функционируют в зависимости от проявлений настроения и поведения человека.
Последнюю группу составляют мышцы ушной раковины (рис. 59).
Рис. 59
Группа состоит из трех слабо развитых небольших мышц — передней, верхней и задней. При сокращении мышцы почти не дают смещаться ушной раковине вперед, вверх и назад. Лишь немногие люди умеют «шевелить» ушами.
Ознакомившись с мышцами головы человека, вы обязаны твердо усвоить, что объемная форма ее образуется благодаря не только выступающим частям черепа, но и мышечному покрову.
Скелетных мышц | Анатомия и физиология
Цели обучения
- Описать слои соединительной ткани, упаковывающие скелетную мышцу
- Объясните, как мышцы работают с сухожилиями для движения тела
- Определить участки волокон скелетных мышц
- Описание муфты возбуждения-сжатия
Самая известная особенность скелетных мышц — это их способность сокращаться и вызывать движение.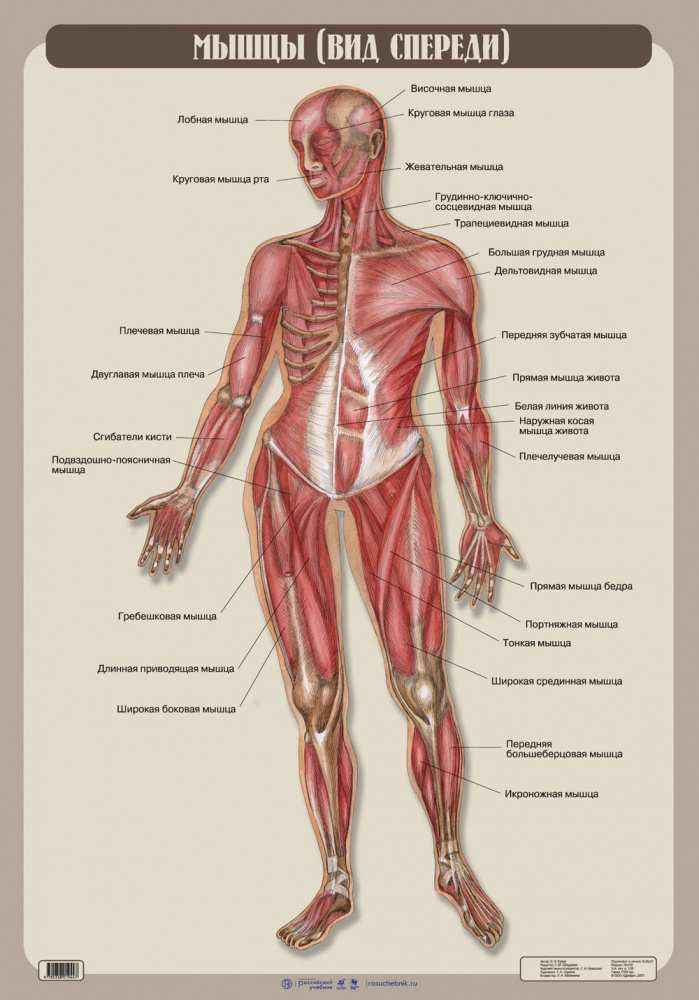 Скелетные мышцы действуют не только для создания движения, но и для остановки движения, например, противостояния силе тяжести для сохранения осанки.Необходимы небольшие, постоянные корректировки скелетных мышц, чтобы удерживать тело в вертикальном или сбалансированном положении в любом положении. Мышцы также предотвращают чрезмерное движение костей и суставов, поддерживая стабильность скелета и предотвращая повреждение или деформацию скелетных структур. Суставы могут полностью смещаться или смещаться, если потянуть за связанные кости; мышцы работают, чтобы суставы оставались стабильными. Скелетные мышцы расположены по всему телу в отверстиях внутренних путей, чтобы контролировать движение различных веществ.Эти мышцы позволяют произвольно контролировать такие функции, как глотание, мочеиспускание и дефекация. Скелетные мышцы также защищают внутренние органы (особенно органы брюшной полости и таза), выступая в качестве внешнего барьера или щита от внешних травм и поддерживая вес органов.
Скелетные мышцы действуют не только для создания движения, но и для остановки движения, например, противостояния силе тяжести для сохранения осанки.Необходимы небольшие, постоянные корректировки скелетных мышц, чтобы удерживать тело в вертикальном или сбалансированном положении в любом положении. Мышцы также предотвращают чрезмерное движение костей и суставов, поддерживая стабильность скелета и предотвращая повреждение или деформацию скелетных структур. Суставы могут полностью смещаться или смещаться, если потянуть за связанные кости; мышцы работают, чтобы суставы оставались стабильными. Скелетные мышцы расположены по всему телу в отверстиях внутренних путей, чтобы контролировать движение различных веществ.Эти мышцы позволяют произвольно контролировать такие функции, как глотание, мочеиспускание и дефекация. Скелетные мышцы также защищают внутренние органы (особенно органы брюшной полости и таза), выступая в качестве внешнего барьера или щита от внешних травм и поддерживая вес органов.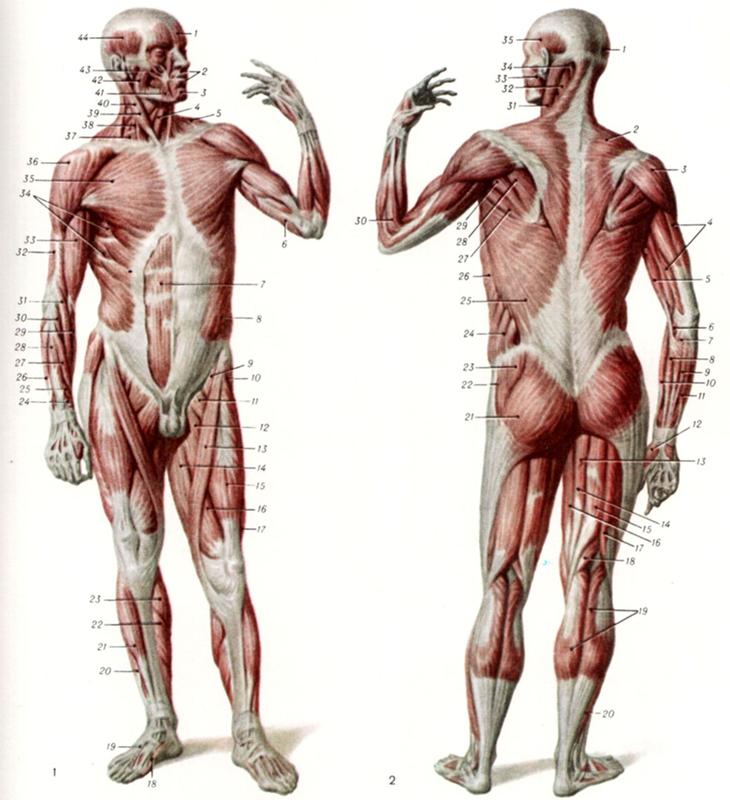
Скелетные мышцы способствуют поддержанию гомеостаза в организме, выделяя тепло. Для сокращения мышц требуется энергия, а при расщеплении АТФ выделяется тепло. Это тепло очень заметно во время упражнений, когда продолжительное движение мышц вызывает повышение температуры тела, а в случаях сильного холода, когда дрожь вызывает случайные сокращения скелетных мышц, чтобы генерировать тепло.
Каждая скелетная мышца — это орган, состоящий из различных интегрированных тканей. Эти ткани включают волокна скелетных мышц, кровеносные сосуды, нервные волокна и соединительную ткань. Каждая скелетная мышца состоит из трех слоев соединительной ткани (называемых «мизия»), которые окружают ее и обеспечивают структуру мышцы в целом, а также разделяют мышечные волокна внутри мышцы (рис. 7.5). Каждая мышца обернута оболочкой из плотной соединительной ткани неправильной формы, называемой эпимизием , которая позволяет мышце сокращаться и мощно двигаться, сохраняя при этом ее структурную целостность. Эпимизий также отделяет мышцу от других тканей и органов в этой области, позволяя мышце двигаться независимо.
Эпимизий также отделяет мышцу от других тканей и органов в этой области, позволяя мышце двигаться независимо.
Внутри каждой скелетной мышцы мышечные волокна организованы в отдельные пучки, каждый из которых называется пучком , за счет среднего слоя соединительной ткани, называемого перимизием .Эта фасцикулярная организация часто встречается в мышцах конечностей; он позволяет нервной системе запускать определенное движение мышцы, активируя подмножество мышечных волокон в пучке или пучке мышцы. Внутри каждого пучка каждое мышечное волокно заключено в тонкий слой соединительной ткани из коллагена и ретикулярных волокон, называемый эндомизиумом . Эндомизий содержит внеклеточную жидкость и питательные вещества, поддерживающие мышечные волокна. Эти питательные вещества поступают в мышечную ткань через кровь.
Эти питательные вещества поступают в мышечную ткань через кровь.
В скелетных мышцах, которые работают с сухожилиями, натягивая кости, коллаген в трех тканевых слоях (мизия) переплетается с коллагеном сухожилия. На другом конце сухожилия оно срастается с надкостницей, покрывающей кость. Напряжение, создаваемое сокращением мышечных волокон, затем передается через мизию на сухожилие, а затем на надкостницу, чтобы тянуть кость для движения скелета. В других местах мезия может срастаться с широким, похожим на сухожилие листом, называемым апоневрозом , или с фасцией, соединительной тканью между кожей и костями.Широкий слой соединительной ткани в нижней части спины, в который срастаются широчайшие мышцы спины («широчайшие»), является примером апоневроза.
Каждая скелетная мышца также богато снабжена кровеносными сосудами для питания, доставки кислорода и удаления отходов. Кроме того, каждое мышечное волокно в скелетной мышце снабжается аксональной ветвью соматического мотонейрона, которая сигнализирует волокну о сокращении. В отличие от сердечных и гладких мышц, единственный способ функционального сокращения скелетных мышц — это передача сигналов от нервной системы.
В отличие от сердечных и гладких мышц, единственный способ функционального сокращения скелетных мышц — это передача сигналов от нервной системы.
Волокна скелетных мышц
Поскольку клетки скелетных мышц длинные и цилиндрические, их обычно называют мышечными волокнами. Волокна скелетных мышц могут быть довольно большими для клеток человека: диаметром до 100 мкм м и длиной до 30 см (7,6 дюйма) в Sartorius верхней части ноги. На раннем этапе развития эмбриональные миобласты, каждый со своим собственным ядром, сливаются с сотнями других миобластов, образуя многоядерные волокна скелетных мышц.Множественные ядра означают множественные копии генов, позволяющие производить большое количество белков и ферментов, необходимых для сокращения мышц.
Другая терминология, связанная с мышечными волокнами, восходит к греческому слову sarco , что означает «плоть». Плазматическая мембрана мышечных волокон называется сарколеммой , цитоплазма называется саркоплазмой , а специальный гладкий эндоплазматический ретикулум, который хранит, высвобождает и извлекает ионы кальция (Ca ++ ) называется саркоплазматической сетью (SR) (Рисунок 7. 6). Как будет вскоре описано, функциональной единицей волокна скелетных мышц является саркомер, высокоорганизованная структура сократительных миофиламентов актина (тонкая нить) и миозина (толстая нить), а также другой опоры. белки.
6). Как будет вскоре описано, функциональной единицей волокна скелетных мышц является саркомер, высокоорганизованная структура сократительных миофиламентов актина (тонкая нить) и миозина (толстая нить), а также другой опоры. белки.
Саркомер
Решетчатый вид волокон скелетных мышц обусловлен расположением миофиламентов актина и миозина в последовательном порядке от одного конца мышечного волокна до другого. Каждая группа этих микрофиламентов называется саркомером и образует функциональную единицу мышечного волокна.
Интерактивная ссылка
Посмотрите это видео, чтобы узнать больше о макро- и микроструктуре скелетных мышц. а) Как называются «точки соединения» между саркомерами? (б) Как называются «субъединицы» в миофибриллах, которые проходят по длине волокон скелетных мышц? в) Что такое «двойная нить жемчуга», описанная в видео? (d) Что придает волокну скелетных мышц поперечно-полосатый вид?
а) Как называются «точки соединения» между саркомерами? (б) Как называются «субъединицы» в миофибриллах, которые проходят по длине волокон скелетных мышц? в) Что такое «двойная нить жемчуга», описанная в видео? (d) Что придает волокну скелетных мышц поперечно-полосатый вид?
Сам саркомер связан с миофибриллами, которые проходят по всей длине мышечного волокна и прикрепляются к сарколемме на своем конце. По мере сокращения миофибрилл сокращается вся мышечная клетка.Поскольку миофибриллы имеют диаметр примерно 1,2 мкм м, от сотен до тысяч (каждая с тысячами саркомеров) можно найти внутри одного мышечного волокна. Каждый саркомер имеет длину примерно 2 мкм, м, имеет трехмерное цилиндрическое расположение и граничит с структурами, называемыми Z-дисками (также называемыми Z-линиями, поскольку изображения двумерные), к которым прикреплены миофиламенты актина. закреплен (рисунок 7.7). Поскольку актин и его комплекс тропонин-тропомиозин (выступающий от Z-дисков к центру саркомера) образуют нити, которые тоньше миозина, его называют тонкой нитью саркомера.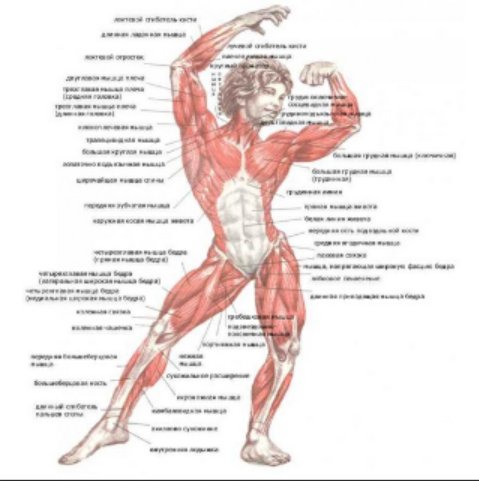 Комплекс тропонин-тропомиозин регулирует процесс сокращения. Точно так же, поскольку нити миозина и их многочисленные головки (выступающие от центра саркомера к Z-дискам, но не полностью к ним) имеют большую массу и толще, их называют толстой нитью . саркомер.
Комплекс тропонин-тропомиозин регулирует процесс сокращения. Точно так же, поскольку нити миозина и их многочисленные головки (выступающие от центра саркомера к Z-дискам, но не полностью к ним) имеют большую массу и толще, их называют толстой нитью . саркомер.
Нервно-мышечное соединение
Другая специализация скелетных мышц — это место, где терминал моторного нейрона встречается с мышечным волокном, называемое нервно-мышечным соединением (НМС) . Здесь мышечное волокно в первую очередь реагирует на сигналы двигательного нейрона. Каждое волокно скелетной мышцы в каждой скелетной мышце иннервируется двигательным нейроном в НМС. Сигналы возбуждения от нейрона — единственный способ функционально активировать сокращение волокна.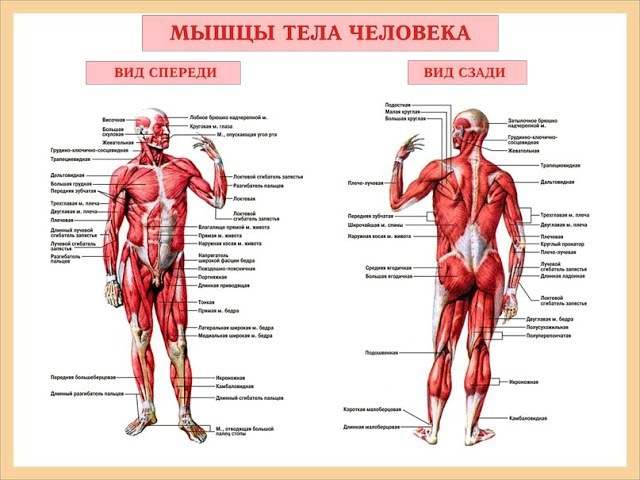
Интерактивная ссылка
Каждое волокно скелетных мышц снабжается двигательным нейроном в СМС. Посмотрите это видео, чтобы узнать больше о том, что происходит в СМП. а) Каково определение моторной единицы? б) Каковы структурные и функциональные различия между большой моторной единицей и малой моторной единицей? (c) Вы можете привести примеры каждого из них? (d) Почему нейромедиатор ацетилхолин разлагается после связывания с его рецептором?
Муфта возбуждения-сжатия
У всех живых клеток есть мембранные потенциалы или электрические градиенты на мембранах.Внутренняя часть мембраны обычно составляет от -60 до -90 мВ относительно внешней стороны. Это называется мембранным потенциалом клетки. Нейроны и мышечные клетки могут использовать свои мембранные потенциалы для генерации электрических сигналов. Они делают это, контролируя движение заряженных частиц, называемых ионами, через свои мембраны для создания электрических токов. Это достигается за счет открытия и закрытия специализированных белков в мембране, называемых ионными каналами. Хотя токи, создаваемые ионами, движущимися через эти канальные белки, очень малы, они формируют основу как нейронной передачи сигналов, так и сокращения мышц.
Хотя токи, создаваемые ионами, движущимися через эти канальные белки, очень малы, они формируют основу как нейронной передачи сигналов, так и сокращения мышц.
И нейроны, и клетки скелетных мышц электрически возбудимы, что означает, что они способны генерировать потенциалы действия. Потенциал действия — это особый тип электрического сигнала, который может перемещаться по клеточной мембране в виде волны. Это позволяет быстро и точно передавать сигнал на большие расстояния.
Хотя термин связь возбуждения-сокращения сбивает с толку или пугает некоторых студентов, он сводится к следующему: для сокращения скелетного мышечного волокна его мембрана должна сначала быть «возбуждена» — другими словами, она должна быть стимулирована к сокращению. запускать потенциал действия.Потенциал действия мышечных волокон, который движется по сарколемме в виде волны, «связан» с фактическим сокращением через высвобождение ионов кальция (Ca ++ ) из SR. После высвобождения Ca ++ взаимодействует с защитными белками, заставляя их отодвигаться, так что сайты связывания актина становятся доступными для прикрепления миозиновыми головками. Затем миозин тянет актиновые нити к центру, укорачивая мышечные волокна.
После высвобождения Ca ++ взаимодействует с защитными белками, заставляя их отодвигаться, так что сайты связывания актина становятся доступными для прикрепления миозиновыми головками. Затем миозин тянет актиновые нити к центру, укорачивая мышечные волокна.
В скелетных мышцах эта последовательность начинается с сигналов соматического моторного отдела нервной системы.Другими словами, этап «возбуждения» в скелетных мышцах всегда запускается сигналом нервной системы (рис. 7.8).
Рисунок 7.8. Концевая пластина двигателя и иннервация В СМП терминал аксона высвобождает ACh. Моторная пластинка — это место расположения ACh-рецепторов в сарколемме мышечного волокна. Когда молекулы ACh высвобождаются, они диффундируют через небольшое пространство, называемое синаптической щелью, и связываются с рецепторами. Моторные нейроны, которые заставляют скелетные мышечные волокна сокращаться, берут начало в спинном мозге, а меньшее их количество находится в стволе мозга для активации скелетных мышц лица, головы и шеи. Эти нейроны имеют длинные отростки, называемые аксонами, которые специализируются на передаче потенциалов действия на большие расстояния — в данном случае от спинного мозга до самой мышцы (которая может находиться на расстоянии до трех футов). Аксоны нескольких нейронов связываются вместе, образуя нервы, как провода, связанные вместе в кабель.
Эти нейроны имеют длинные отростки, называемые аксонами, которые специализируются на передаче потенциалов действия на большие расстояния — в данном случае от спинного мозга до самой мышцы (которая может находиться на расстоянии до трех футов). Аксоны нескольких нейронов связываются вместе, образуя нервы, как провода, связанные вместе в кабель.
Передача сигналов начинается, когда нейрон с потенциалом действия перемещается по аксону двигательного нейрона, а затем по отдельным ветвям и заканчивается в НМС.В NMJ окончание аксона выпускает химический мессенджер, или нейротрансмиттер , называемый ацетилхолин (ACh) . Молекулы ACh диффундируют через крошечное пространство, называемое синаптической щелью , и связываются с рецепторами ACh, расположенными в моторной концевой пластине сарколеммы на другой стороне синапса. Как только ACh связывается, канал в рецепторе ACh открывается, и положительно заряженные ионы могут проходить в мышечное волокно, вызывая деполяризацию , что означает, что мембранный потенциал мышечного волокна становится менее отрицательным (ближе к нулю.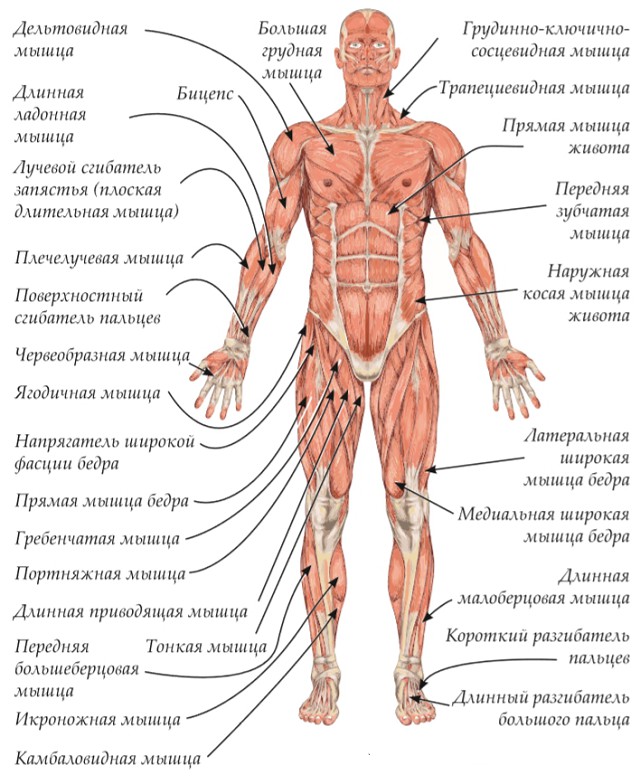 )
)
По мере того, как мембрана деполяризуется, другой набор ионных каналов, называемый потенциалозависимые натриевые каналы , запускается для открытия. Ионы натрия попадают в мышечные волокна, и потенциал действия быстро распространяется (или «вспыхивает») по всей мембране, инициируя связь возбуждения и сокращения.
В мире возбудимых мембран все происходит очень быстро (только подумайте, как быстро вы можете щелкнуть пальцами, как только решите это сделать). Сразу после деполяризации мембраны она реполяризуется, восстанавливая отрицательный мембранный потенциал.Между тем, ACh в синаптической щели расщепляется ферментом ацетилхолинэстеразой (AChE), так что ACh не может повторно связываться с рецептором и повторно открывать свой канал, что может вызвать нежелательное расширенное возбуждение и сокращение мышц.
Распространение потенциала действия по сарколемме является возбуждающей частью взаимодействия возбуждения-сокращения. Напомним, что это возбуждение фактически запускает высвобождение ионов кальция (Ca ++ ) из их хранилища в SR клетки. Чтобы потенциал действия достигал мембраны SR, в сарколемме есть периодические инвагинации, называемые Т-канальцами («Т» означает «поперечный»).Вы помните, что диаметр мышечного волокна может достигать 100 мкм м, поэтому эти Т-канальцы гарантируют, что мембрана может приблизиться к SR в саркоплазме. Расположение Т-канальца с мембранами SR с обеих сторон называется триадой (рис. 7.9). Триада окружает цилиндрическую структуру, называемую миофибриллой , которая содержит актин и миозин.
Т-канальцы несут потенциал действия внутрь клетки, что запускает открытие кальциевых каналов в мембране соседнего SR, заставляя Ca ++ диффундировать из SR в саркоплазму.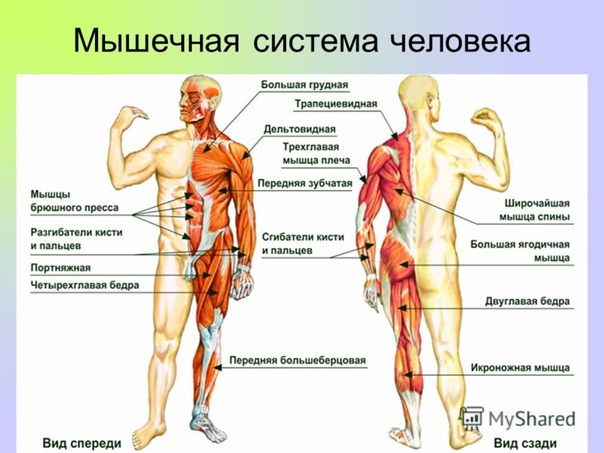 Именно поступление Ca ++ в саркоплазму инициирует сокращение мышечного волокна его сократительными единицами, или саркомерами.
Именно поступление Ca ++ в саркоплазму инициирует сокращение мышечного волокна его сократительными единицами, или саркомерами.
Сетевая структура опорно-двигательного аппарата человека формирует нейронных взаимодействий на нескольких временных масштабах
Абстрактный управления двигателем человеческого требует координации мышечной активности при анатомических ограничений, накладываемых опорно-двигательного аппарата. Взаимодействия внутри центральной нервной системы имеют фундаментальное значение для координации движений, но принципы функциональной интеграции остаются плохо изученными.Мы использовали сетевой анализ, чтобы исследовать взаимосвязь между анатомической и функциональной связностью между 36 мышцами. Анатомические сети определялись физическими связями между мышцами, а функциональные сети основывались на межмышечной когерентности, оцениваемой во время постуральных задач. Мы нашли модульную структуру функциональных сетей, которые сильно сформированным анатомических ограничений костно-мышечной системы. Изменения постуральных задач были связаны с частотно-зависимой реконфигурацией связи между функциональными модулями.Эти данные раскрывают различные паттерны функционального взаимодействия между мышцами, участвующими в гибкой организации мышечной активности во время постурального контроля. Наш сетевой подход к системе двигателя предлагает уникальное окно в нейронную схему вождения костно-мышечную системы.
Изменения постуральных задач были связаны с частотно-зависимой реконфигурацией связи между функциональными модулями.Эти данные раскрывают различные паттерны функционального взаимодействия между мышцами, участвующими в гибкой организации мышечной активности во время постурального контроля. Наш сетевой подход к системе двигателя предлагает уникальное окно в нейронную схему вождения костно-мышечную системы.
ВВЕДЕНИЕ
Человеческое тело представляет собой сложную систему, состоящую из множества подсистем и регуляторных путей. Костно-мышечная система дает структуру тела и создает способность двигаться. Он состоит из более чем 200 костей скелета, соединительной ткани и более 300 скелетных мышц.Мышцы прикрепляются к костям через сухожилие и могут вызывать движение вокруг сустава при сокращении. Центральная нервная система контролирует эти движения через двигательные нейроны спинного мозга, которые служат последним общим путем к мышцам ( 1 ). В то время как анатомо-физиологические компоненты опорно-двигательного аппарата хорошо охарактеризованы ( 2 , 3 ), организационные принципы нервного контроля остаются мало изучены. Здесь мы выяснить взаимосвязь между анатомическим строением костно-мышечной системы и функциональной организации распределенной нейронной схемы, из которых двигательные поведения мола.
Здесь мы выяснить взаимосвязь между анатомическим строением костно-мышечной системы и функциональной организации распределенной нейронной схемы, из которых двигательные поведения мола.
Традиционная идея о том, что кора головного мозга контролирует мышцы взаимно однозначно, была опровергнута несколькими линиями доказательств ( 4 ). Например, широко признано, что много степеней свободы (степени свободы) опорно-двигательного аппарата запретить простое взаимно-однозначное соответствие между двигательной задачей и конкретным решением двигателя; скорее, мышцы связаны и контролируются вместе ( 5 ). Связь между мышцами — механическими или нервными — уменьшает количество эффективных степеней свободы и, следовательно, количество потенциальных моделей движения.Таким образом, эта муфта снижает сложность управления двигателем ( 6 ).
Продолжаются споры о природе сцепления между мышцами. Механическое соединение в опорно-двигательной системе ограничивает паттерны движения, которые могут быть сгенерированы ( 7 , 8 ). Например, биомеханика конечности ограничивает относительные изменения в длине мускульно-сухожильных мышц субпространством с малой размерностью, что приводит к коррелированным афферентным входам в двигательные нейроны спинного мозга ( 9 ).Связь между мышцами также может быть результатом дублирования нейронных цепей, управляющих двигательными нейронами спинного мозга ( 10 ). Электрофизиологические исследования показывают, что комбинация всего лишь нескольких последовательных паттернов мышечной активации — или мышечной синергии — может генерировать широкий спектр естественных движений ( 11 ). Некоторые из этих паттернов уже присутствуют с рождения и не меняются в процессе развития, тогда как другие паттерны усваиваются ( 12 ). Такое расположение поддерживает представление о том, что нервно-мышечная система имеет модульную организацию, которая упрощает задачу управления ( 13 ).Спинальная схема состоит из сети премоторных интернейронов и двигательных нейронов, которые могут генерировать основные двигательные паттерны, опосредуя синергетический драйв множеству мышц ( 14 ).
Например, биомеханика конечности ограничивает относительные изменения в длине мускульно-сухожильных мышц субпространством с малой размерностью, что приводит к коррелированным афферентным входам в двигательные нейроны спинного мозга ( 9 ).Связь между мышцами также может быть результатом дублирования нейронных цепей, управляющих двигательными нейронами спинного мозга ( 10 ). Электрофизиологические исследования показывают, что комбинация всего лишь нескольких последовательных паттернов мышечной активации — или мышечной синергии — может генерировать широкий спектр естественных движений ( 11 ). Некоторые из этих паттернов уже присутствуют с рождения и не меняются в процессе развития, тогда как другие паттерны усваиваются ( 12 ). Такое расположение поддерживает представление о том, что нервно-мышечная система имеет модульную организацию, которая упрощает задачу управления ( 13 ).Спинальная схема состоит из сети премоторных интернейронов и двигательных нейронов, которые могут генерировать основные двигательные паттерны, опосредуя синергетический драйв множеству мышц ( 14 ). Эти спинномозговые сети могут кодировать программы скоординированных моторных выходов ( 15 ), которые могут использоваться для преобразования нисходящих команд для многосуставных движений в соответствующие скоординированные мышечные синергии, которые поддерживают эти движения ( 3 ).
Эти спинномозговые сети могут кодировать программы скоординированных моторных выходов ( 15 ), которые могут использоваться для преобразования нисходящих команд для многосуставных движений в соответствующие скоординированные мышечные синергии, которые поддерживают эти движения ( 3 ).
Сети может представить альтернативную точку зрения на модульной организации костно-мышечной системы.Сообщества или модульные структуры, которые относятся к плотно связанным группам узлов с только редкими связями между этими группами, являются одной из наиболее важных особенностей сложных сетей ( 16 ). Исследование структур сообщества широко используется в различных областях, таких как сети мозга ( 17 ). Этот подход был недавно применен к исследованию структуры и функции опорно-двигательного аппарата: Анатомическое сеть может быть построена путем сопоставления происхождение и вставку мышц ( 18 , 19 ).Ранее мы показали, как можно построить функциональные мышечные сети, оценивая межмышечную когерентность с помощью поверхностной электромиографии (ЭМГ), записанной с различных мышц ( 20 ).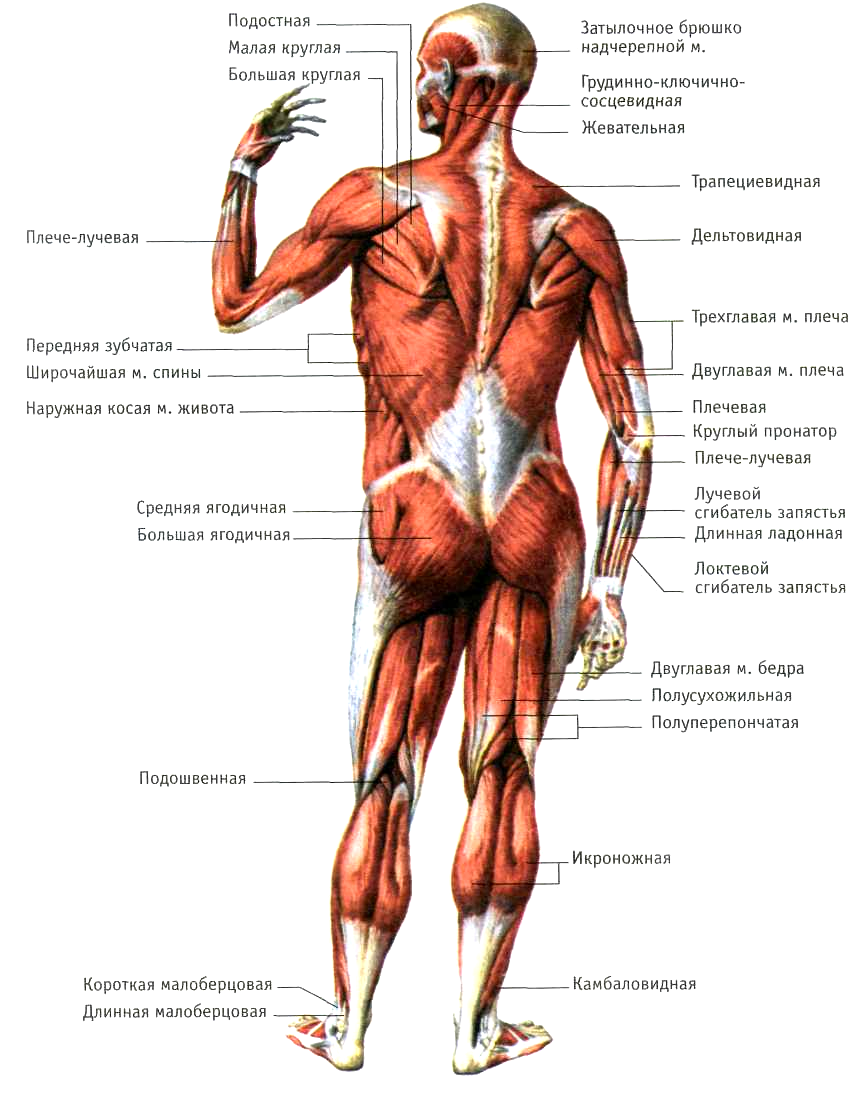 Эти функциональные сети обнаруживают функциональную связь между группами мышц во многих частотных диапазонах. Согласованность между ЭМГ указывает на коррелированные или общие входы в двигательные нейроны спинного мозга, которые генерируются общими структурными связями или синхронизацией в двигательной системе ( 10 , 21 , 22 ).Таким образом, паттерны функциональной связности позволяют оценивать структурные пути в двигательной системе с использованием неинвазивных записей ( 23 ).
Эти функциональные сети обнаруживают функциональную связь между группами мышц во многих частотных диапазонах. Согласованность между ЭМГ указывает на коррелированные или общие входы в двигательные нейроны спинного мозга, которые генерируются общими структурными связями или синхронизацией в двигательной системе ( 10 , 21 , 22 ).Таким образом, паттерны функциональной связности позволяют оценивать структурные пути в двигательной системе с использованием неинвазивных записей ( 23 ).
Здесь мы исследуем организационные принципы, управляющие человеческим моторным контролем, сравнивая структуру сообщества анатомических и функциональных сетей. Мы используем мультиплексный анализ модульности ( 24 ) для оценки структуры сообщества функциональных мышечных сетей по частотам и постуральным задачам. В биомеханических свойств костно-мышечной системы сдерживало характера движения, которые могут быть сгенерированы, мы ожидаем, что подобную структуру сообщества анатомических и функциональных сетей мышц.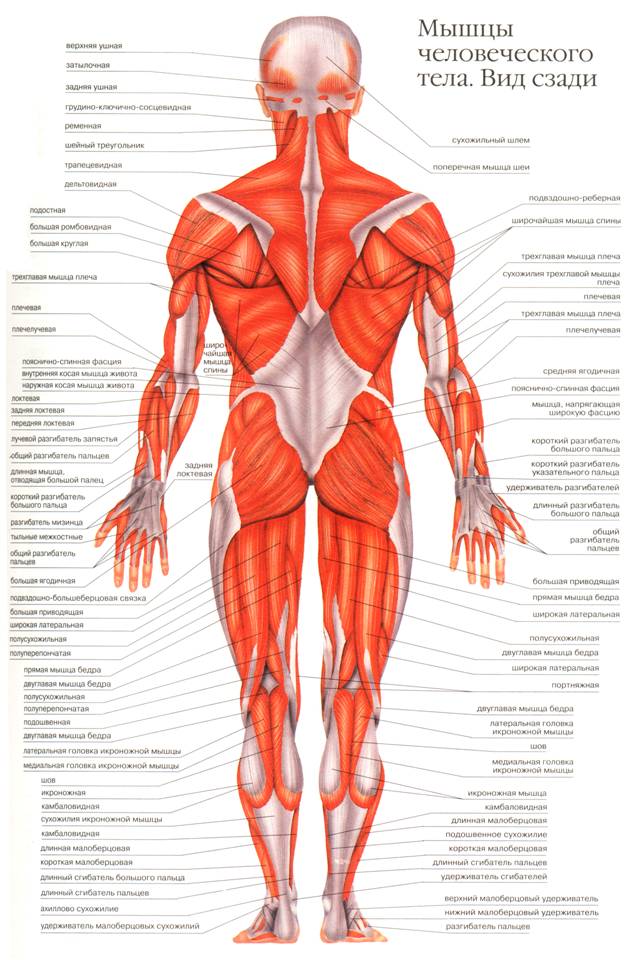 Отклонения в структуре сообщества указывают на дополнительные ограничения, налагаемые центральной нервной системой. Мы также сравниваем функциональную взаимосвязь между модулями во время выполнения различных задач, чтобы исследовать изменения в функциональной организации во время поведения. В то время как средняя функциональная связность ограничена анатомическими ограничениями, мы ожидаем, что функциональные мышечные сети переконфигурируются для обеспечения зависимых от задач моделей координации между мышцами. Эти задачи модуляции будут означать, что функциональные взаимодействия между мышцами не зашитые, но вместо этого определяются динамической связью в центральной нервной системе, которая имеет форму анатомической топологией костно-мышечную системы.
Отклонения в структуре сообщества указывают на дополнительные ограничения, налагаемые центральной нервной системой. Мы также сравниваем функциональную взаимосвязь между модулями во время выполнения различных задач, чтобы исследовать изменения в функциональной организации во время поведения. В то время как средняя функциональная связность ограничена анатомическими ограничениями, мы ожидаем, что функциональные мышечные сети переконфигурируются для обеспечения зависимых от задач моделей координации между мышцами. Эти задачи модуляции будут означать, что функциональные взаимодействия между мышцами не зашитые, но вместо этого определяются динамической связью в центральной нервной системе, которая имеет форму анатомической топологией костно-мышечную системы.
РЕЗУЛЬТАТЫ
Мы оценили взаимосвязь между анатомической и функциональной связностью ключевых мышц, участвующих в задачах контроля позы (36 мышц, распределенных по всему телу). Мы исследовали мышечно-ориентированную сеть, в которой узлы представляют мышцы, а края сети представляют собой анатомические связи или функциональные отношения между мышцами.
Анатомическая мышечная сеть
Анатомическая мышечная сеть была определена путем картирования физических связей между мышцами ( 19 , 25 ) на основе общей анатомии человека ( 2 ).Анатомическая сеть представляет собой плотно связанную симметричную сеть (плотность сети 0,27; рис. 1). Анализ модульности выявил пять модулей, которые разделили анатомическую мышечную сеть на основные части тела (правая рука, левая рука, туловище, правая нога и левая нога) с модульностью 0,39.
Рис. 1 Сообщество анатомической мышечной сети. ( A ) Топологическое представление анатомической сети. Узлы сети представляют мышцы, а края представляют собой анатомические связи между мышцами, которые прикреплены к одной и той же кости или соединительной ткани.Пять модулей имеют цветовую кодировку. ( B ) Пространственное представление анатомической мышечной сети, отображаемой на теле человека ( 53 ). Размер каждого узла представляет собой количество других узлов, к которым он подключен.
Функциональная мышечная сеть
Функциональная мышечная сеть была определена путем сопоставления коррелированных входных сигналов с различными мышцами. Чтобы составить карту функциональных сетей, мы измерили поверхностную ЭМГ тех же 36 мышц, в то время как здоровые участники выполняли разные постуральные задачи.Был использован полнофакторный план, в котором мы варьировали контроль позы (нормальное положение и нестабильность в передне-заднем или медиально-латеральном направлении) и поведение при указании (не указывать и указывать доминирующей рукой или обеими руками; см. Материалы и методы подробнее). Мы использовали эти задачи, чтобы экспериментально манипулировать необходимой координацией между мышцами и вызвать изменения в функциональной мышечной сети. Мы оценили функциональную связность посредством межмышечной когерентности между всеми мышечными комбинациями и использовали неотрицательную матричную факторизацию (NNMF), чтобы разложить эти спектры когерентности на частотные компоненты и соответствующие веса границ.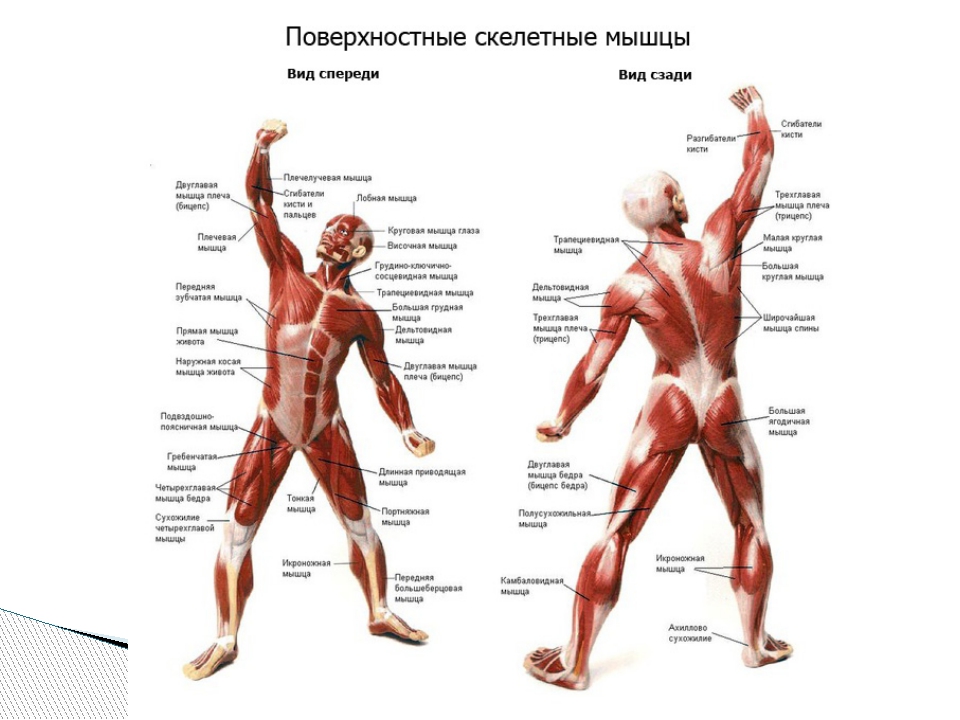 Это дало набор взвешенных сетей с соответствующими спектральными отпечатками (частотными компонентами).
Это дало набор взвешенных сетей с соответствующими спектральными отпечатками (частотными компонентами).
Мы наблюдали четыре отдельных частотных компонента (компонент 1, от 0 до 3 Гц; компонент 2, от 3 до 11 Гц; компонент 3, от 11 до 21 Гц; компонент 4, от 21 до 60 Гц; рис. 2A), которые служат отдельными слоев мультиплексной сети и объяснил большую часть дисперсии спектров когерентности ( R 2 = 0,90). Для получения минимально связной двоичной сети между уровнями и для сохранения постоянного количества ребер между слоями (относительный порог, 0.035). Используя мультиплексный анализ модульности, мы получили фиксированную структуру сообщества по всем четырем частотам и девяти состояниям, которые выявили шесть модулей: правое плечо, двусторонние предплечья, туловище, правое бедро, левое верхнее положение и двустороннее голени (рис. 2B). . На рис. 2С показано, как эти модули распределяются по телу. Отчетливые топологии сети наблюдались на разных уровнях с более широко подключенной сетью на более низких частотах и более разделенной сетью на более высоких частотах: плотность сети равнялась 0.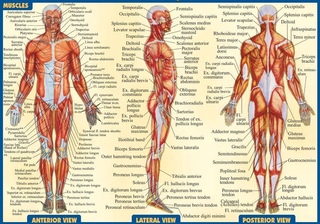 10, 0,09, 0,08 и 0,06, а модульность составляла 0,46, 0,60, 0,64 и 0,75 для компонентов 1, 2, 3 и 4 соответственно (рис. 2D).
10, 0,09, 0,08 и 0,06, а модульность составляла 0,46, 0,60, 0,64 и 0,75 для компонентов 1, 2, 3 и 4 соответственно (рис. 2D).
( A ) Частотные спектры четырех компонентов, полученные с использованием NNMF. ( B ) Мультиплексная структура сообщества функциональной мышечной сети в зависимости от частот и условий. Доминирующая рука всех участников отображается на правой стороне человеческого тела.( C ) Пространственное представление средней мышечной сети, отображаемой на теле человека ( 53 ). Размер узлов представляет собой количество других узлов, к которым они подключены, а ширина ребер — количество ребер по слоям. ( D ) Бинарные мышечные сети для каждого слоя.
Сравнение анатомических и функциональных сетей
Сообщества анатомических и функциональных мышечных сетей были очень похожи (индекс Рэнда, 0. 80; скорректированный индекс Rand — 0,36; P <0,001). Связи между двусторонними мышцами предплечья и двусторонних мышц голени в функциональных сетях, которые отсутствовали в анатомической сети, представляют собой заметное различие между анатомической и функциональной сетями. Это отражено в структуре сообщества функциональных сетей, где двусторонние мышцы голени и двусторонние мышцы предплечья были сгруппированы в отдельные модули (рис. 2С).
Сравнение анатомического расстояния (длины пути) и функциональной связности показало, что анатомически близлежащие узлы с большей вероятностью получат общий ввод (рис.3). Сначала мы исследовали процент всех возможных краев, то есть количество краев выше порога, которое уменьшалось в зависимости от анатомического расстояния: 11,3, 0,9, 0,6 и 0,0% для анатомических расстояний 1, 2, 3 и 4, соответственно. Это снижение с расстоянием было еще более заметным для более высокочастотных компонентов (рис. 3B). Затем мы исследовали распределение функциональных весов в зависимости от анатомического расстояния. Наибольший вес наблюдался для ребер, соединяющих мышцы в одном модуле.Края в большинстве модулей имели анатомическое расстояние 1. Лишь несколько ребер имели анатомическое расстояние 2 или 3, и все эти края находились в модулях предплечья и голени. В частности, края, соединяющие двусторонние мышцы голени, имели относительно большой вес на анатомическом расстоянии 3 (рис. 3C).
Наибольший вес наблюдался для ребер, соединяющих мышцы в одном модуле.Края в большинстве модулей имели анатомическое расстояние 1. Лишь несколько ребер имели анатомическое расстояние 2 или 3, и все эти края находились в модулях предплечья и голени. В частности, края, соединяющие двусторонние мышцы голени, имели относительно большой вес на анатомическом расстоянии 3 (рис. 3C).
( A ) Матрица смежности и расстояния анатомической мышечной сети.Максимальное анатомическое расстояние (длина пути) составляет 4. ( B ) Процент функциональных границ пороговых сетей в экспериментальных условиях как функция анатомического расстояния. ( C ) Распределение веса краев функциональных сетей в зависимости от анатомического расстояния для каждого слоя. Веса были усреднены по условиям эксперимента. Края, соединяющие мышцы в одном модуле, имеют цветовую кодировку (rUA, правое плечо; FA, двусторонние предплечья; T, торс; rUL, правая верхняя нога; lUL, левая верхняя нога; и LL, двусторонние нижние ноги) и серые точки.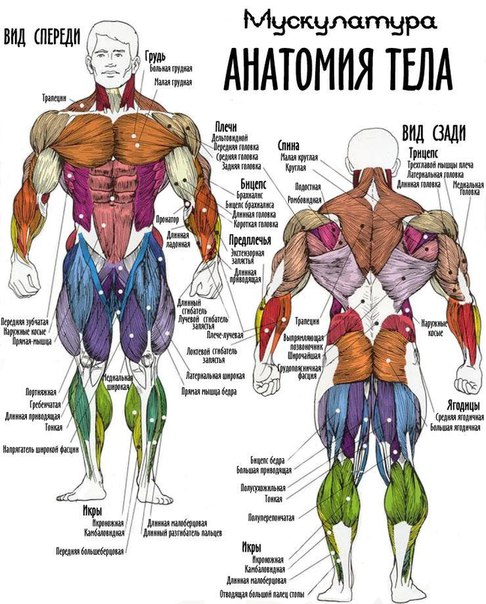 представляют ребра между модулями.
представляют ребра между модулями.
Модуляции, зависящие от задачи
Далее мы попытались изучить влияние задачи на эту взаимосвязь между структурой и функцией. Это было достигнуто за счет использования кластерных графиков для сравнения функциональных мышечных сетей в разных условиях выполнения задачи. Функциональные модули, идентифицированные с помощью предыдущего анализа модульности мультиплексирования, образуют узлы этих кластерных графов. На рис. 4А показаны сгруппированные графики для девяти экспериментальных условий и для четырех частотных составляющих. Кластеризованные графы были очень разреженными, так как модули имеют плотные внутримодульные связи, но разреженные связи между узлами в других модулях.Большинство краев наблюдались между модулями мышц ног (голень, правая верхняя часть ноги и левая верхняя часть ноги) при самых низких частотных компонентах (от 0 до 3 и от 3 до 11 Гц), что соответствует нижним показателям модульности, в частности, когда постуральная стабильность вызвано нестабильностью в передне-заднем или медиально-латеральном направлении.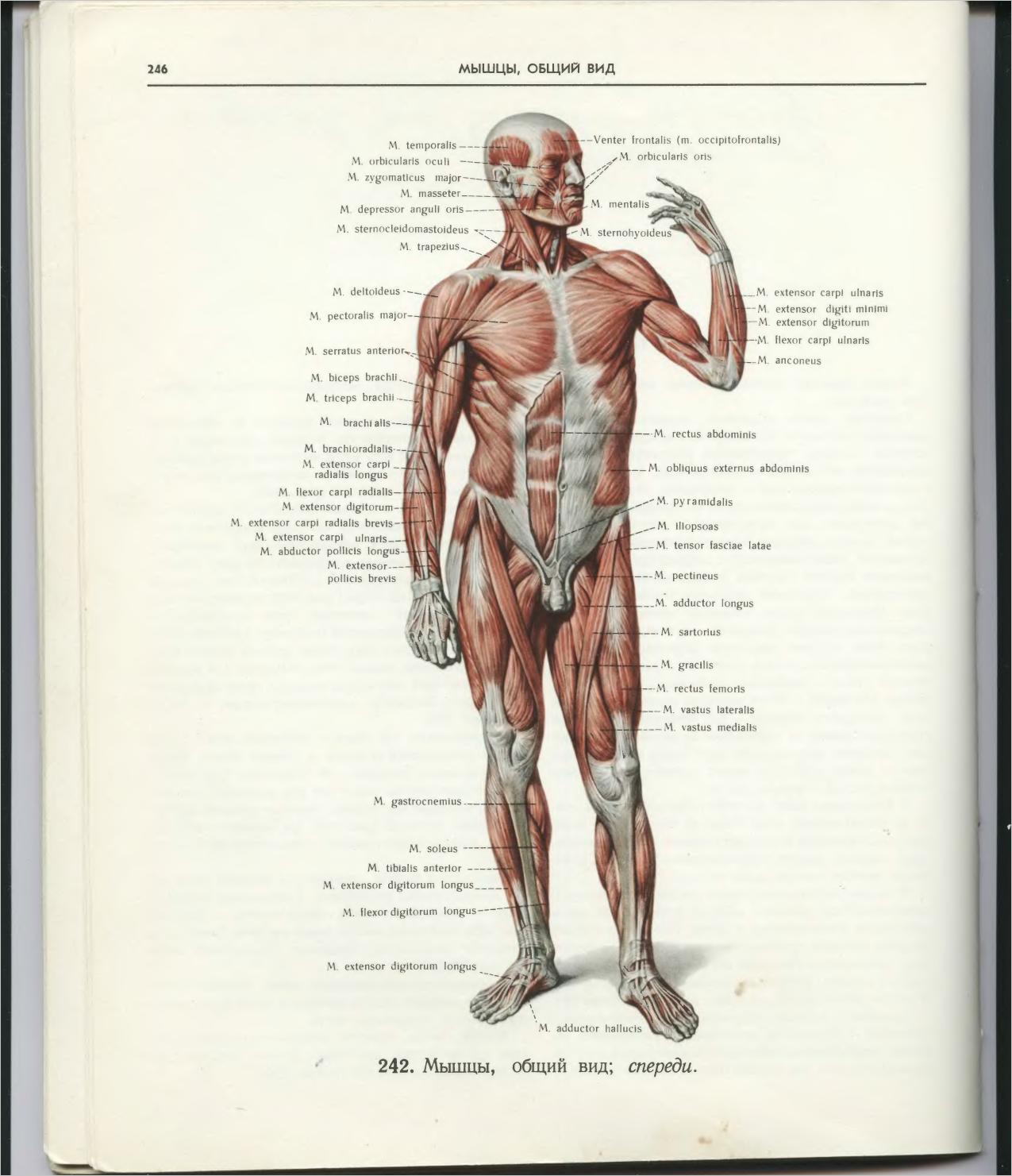 Края между мышечными модулями руки (правое плечо и предплечье) и туловищем в основном наблюдались на более высокочастотных компонентах (от 11 до 21 и от 21 до 60 Гц) во время наведения (одно- и бимануальное).
Края между мышечными модулями руки (правое плечо и предплечье) и туловищем в основном наблюдались на более высокочастотных компонентах (от 11 до 21 и от 21 до 60 Гц) во время наведения (одно- и бимануальное).
( A ) Кластерные графики для девяти экспериментальных условий (столбцы) и четырех частотных компонентов (строки). Узлы — это модули, идентифицированные с помощью анализа модульности мультиплексирования. Размер узла представляет собой плотность сети внутри, а ширина краев представляет собой плотность соединения между модулями. ( B ) Пространственное представление функциональных модулей на теле человека: правое плечо (rUA), двусторонние предплечья (FA), туловище (T), правое плечо (rUL), левое плечо (lUL) и двустороннее голени (LL).Мы использовали наборы инструментов для обработки геометрии, чтобы создать цветные сетки ( 54 ) и отобразить их на теле человека ( 53 ). ( C ) Значительные различия в связности кластерных графов между условиями стабильности. Оценивали два контраста: нормальная стабильность — передне-задняя нестабильность и нормальная стабильность — медиально-латеральная нестабильность. Использовался тест перестановки, и семейный контроль ошибок поддерживался с помощью поправки Бонферрони (84 сравнения).Значимые различия ( P, , , скорректированный, <0,05) имеют цветовую кодировку: красный означает увеличение, а синий - уменьшение среднего веса. Цветные края и узлы показывают значительные изменения в связях между модулями и внутри них соответственно. ( D ) Значительные различия в связности сгруппированных графов между условиями наведения. Были оценены два контраста: отсутствие указания — однозначное наведение и отсутствие указания — бимануальное наведение.
( C ) Значительные различия в связности кластерных графов между условиями стабильности. Оценивали два контраста: нормальная стабильность — передне-задняя нестабильность и нормальная стабильность — медиально-латеральная нестабильность. Использовался тест перестановки, и семейный контроль ошибок поддерживался с помощью поправки Бонферрони (84 сравнения).Значимые различия ( P, , , скорректированный, <0,05) имеют цветовую кодировку: красный означает увеличение, а синий - уменьшение среднего веса. Цветные края и узлы показывают значительные изменения в связях между модулями и внутри них соответственно. ( D ) Значительные различия в связности сгруппированных графов между условиями наведения. Были оценены два контраста: отсутствие указания — однозначное наведение и отсутствие указания — бимануальное наведение.
Влияние задач на устойчивость в основном ограничивалось модулями ног (рис.4С). Повышенная связность наблюдалась во время постуральной нестабильности (переднезадней и медиально-латеральной) по сравнению с нормальным стоянием в большинстве частотных компонентов. На самом низком частотном компоненте (от 0 до 3 Гц) возможность подключения увеличилась внутри и между большинством модулей ног ( P, , , скорректированное, <0,01). Только небольшие различия наблюдались при частоте от 3 до 11 Гц: повышенная связь между модулями туловища и голени во время передне-задней нестабильности [+ 25% (диапазон от -9 до 46%), P скорректировано = 0.01] и снижение связности в модуле туловища во время медиально-латеральной нестабильности [-21% (диапазон от -50 до 0,3%), P скорректировано = 0,01]. Связность снова увеличилась на самых высоких частотных компонентах (от 11 до 21 и от 21 до 60 Гц) внутри и между модулями туловища и ног (правая верхняя нога, левая верхняя нога и нижняя нога; P с поправкой <0,02).
На самом низком частотном компоненте (от 0 до 3 Гц) возможность подключения увеличилась внутри и между большинством модулей ног ( P, , , скорректированное, <0,01). Только небольшие различия наблюдались при частоте от 3 до 11 Гц: повышенная связь между модулями туловища и голени во время передне-задней нестабильности [+ 25% (диапазон от -9 до 46%), P скорректировано = 0.01] и снижение связности в модуле туловища во время медиально-латеральной нестабильности [-21% (диапазон от -50 до 0,3%), P скорректировано = 0,01]. Связность снова увеличилась на самых высоких частотных компонентах (от 11 до 21 и от 21 до 60 Гц) внутри и между модулями туловища и ног (правая верхняя нога, левая верхняя нога и нижняя нога; P с поправкой <0,02).
Задания по наведению показали другую схему по сравнению с заданиями позы, но эффекты одно- и бимануального наведения были очень похожими (рис.4D). Во время наведения связь в модуле туловища снизилась на самых низких частотных составляющих [от 0 до 3 Гц, −61% (диапазон от −90 до −1%), P с поправкой <0,005; От 3 до 11 Гц, -59% (диапазон от -86 до 2%), P с поправкой <0,02] и между туловищем и правым модулем плеча только при самой низкой частотной составляющей [от 0 до 3 Гц, - 67% (диапазон от -93 до -9%), P скорректированный <0,005]. Напротив, во время одноручного наведения наблюдалось значительное увеличение возможности соединения в правом верхнем модуле плеча по сравнению с отсутствием наведения на высокочастотные компоненты [от 11 до 21 Гц, + 64% (диапазон, от -4 до 95%), P исправлено = 0.005; От 21 до 60 Гц, + 66% (диапазон от –12 до 93%), P скорректированный = 0,015]. Кроме того, увеличилось соединение между модулями туловища и предплечья [+ 41% (диапазон от -8 до 82%), P исправлено, <0,01] и между правым плечом и модулями предплечья [+44 % (диапазон от 0 до 82%), P скорректировано <0,005] во время наведения (одноручного и двухручного) по сравнению с отсутствием наведения на частотную составляющую 3 (от 11 до 21 Гц).
Напротив, во время одноручного наведения наблюдалось значительное увеличение возможности соединения в правом верхнем модуле плеча по сравнению с отсутствием наведения на высокочастотные компоненты [от 11 до 21 Гц, + 64% (диапазон, от -4 до 95%), P исправлено = 0.005; От 21 до 60 Гц, + 66% (диапазон от –12 до 93%), P скорректированный = 0,015]. Кроме того, увеличилось соединение между модулями туловища и предплечья [+ 41% (диапазон от -8 до 82%), P исправлено, <0,01] и между правым плечом и модулями предплечья [+44 % (диапазон от 0 до 82%), P скорректировано <0,005] во время наведения (одноручного и двухручного) по сравнению с отсутствием наведения на частотную составляющую 3 (от 11 до 21 Гц).
ОБСУЖДЕНИЯ Мы использовали сетевой подход для изучения структурно-функциональной взаимосвязи костно-мышечной системы человека.были выявлены несколько принципов, регулирующих функциональную зависимость между мышцами: (я) Функциональные модели с подключением между мышцами сильно формируется анатомических ограничений опорно-двигательного аппарата, с функциональной связи сильнейших в анатомических модулей и уменьшается в зависимости от анатомической расстояния; (ii) двусторонняя связь между гомологичными верхними и между гомологичными нижними конечностями является ключевой характеристикой функциональных мышечных сетей; (iii) функциональные отношения зависят от задачи, при этом постуральные задачи по-разному влияют на функциональную связность в разных частотных диапазонах. Использование мультиплексного подхода позволяет интегрировать функциональные мышечные сети на разных частотах и обеспечивает объединяющее окно в распределенные схемы центральной нервной системы человека, которые контролируют движения, иннервируя двигательные нейроны спинного мозга.
Использование мультиплексного подхода позволяет интегрировать функциональные мышечные сети на разных частотах и обеспечивает объединяющее окно в распределенные схемы центральной нервной системы человека, которые контролируют движения, иннервируя двигательные нейроны спинного мозга.
Определение отношений между анатомическими и функциональными мышечными сетями имеет решающее значение для понимания того, как координируются движения. Предыдущие исследования исследовались либо как биомеханические свойства опорно-двигательного аппарат ограничивает модели движения, которые могут быть сформированы ( 8 , 9 ) или как шаблоны активации мышц можно объяснить сочетание всего лишь нескольких моделей активации когерентных мышц ( 11 ).Наши совместные анализы анатомических и функциональных сетей мышц обнаруживают сильную взаимосвязь между анатомическими связями в костно-мышечной системы и коррелированных входных данных для спинальных двигательных нейронов. Этот вывод основан на предыдущих исследованиях, показывающих, что общий вход наиболее силен для спинномозговых мотонейронов, которые иннервируют пары мышц, которые анатомически и функционально тесно связаны ( 10 , 21 ).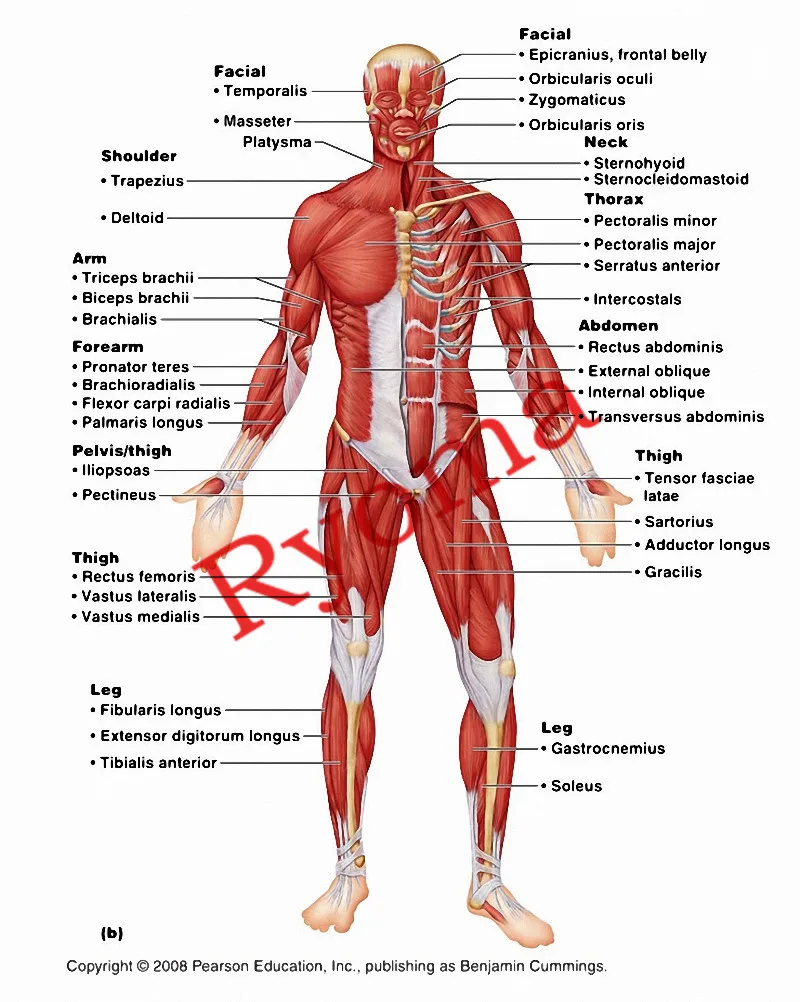 Сходство между структурными и функциональными сетями было признаком исследования сетей мозга ( 26 ), а топология сетей мозга зависит от пространственного встраивания мозга ( 27 ).Настоящие результаты показывают, что принципы, управляющие воплощенными структурными и функциональными сетями, также применимы к нейронным цепям, которые контролируют движения, и, следовательно, могут отражать общий принцип центральной нервной системы.
Сходство между структурными и функциональными сетями было признаком исследования сетей мозга ( 26 ), а топология сетей мозга зависит от пространственного встраивания мозга ( 27 ).Настоящие результаты показывают, что принципы, управляющие воплощенными структурными и функциональными сетями, также применимы к нейронным цепям, которые контролируют движения, и, следовательно, могут отражать общий принцип центральной нервной системы.
Сходство между анатомической и функциональной связностью может указывать на то, что анатомическая структура ограничивает функциональные взаимодействия между мышцами. Анатомические связи между мышцами остаются в основном неизменными на протяжении всей жизни ( 28 ), и более вероятно, что быстро меняющиеся функциональные сети ограничены гораздо более медленными изменениями анатомических сетей, чем наоборот.Эти ограничения могут быть наложены посредством афферентной активности. Скелетно-мышечные свойства человеческого тела ограничивают постуральную динамику ( 9 ), и эти механические связи приводят к коррелированной проприоцептивной обратной связи с двигательными нейронами спинного мозга. Влияние биомеханики на функциональных сетей мышц, как ожидается, будет наиболее выражена на компоненты нижних частот, так как мышцы действуют как фильтр нижних частот на нейрональных входов и кинематика опорно-двигательного аппарата разворачиваться на медленном масштабе времени.Это генерирует коррелированную активность на низких частотах, которая возвращается к моторным нейронам спинного мозга через сенсорные афференты. Пространственное распределение общего входа будет возможно отражать топологию опорно-двигательного аппарата.
Влияние биомеханики на функциональных сетей мышц, как ожидается, будет наиболее выражена на компоненты нижних частот, так как мышцы действуют как фильтр нижних частот на нейрональных входов и кинематика опорно-двигательного аппарата разворачиваться на медленном масштабе времени.Это генерирует коррелированную активность на низких частотах, которая возвращается к моторным нейронам спинного мозга через сенсорные афференты. Пространственное распределение общего входа будет возможно отражать топологию опорно-двигательного аппарата.
Анатомические ограничения также могут быть наложены во время развития нервной системы. Во время раннего развития изменения в топографическом распределении окончаний аксонов нисходящих проектов зависят от паттернов двигательной активности и анатомической связи между мышцами ( 29 ).Аналогичным образом, большие изменения функциональной связи наблюдаются у младенцев в возрасте от 9 до 25 недель, что отражает чувствительный период, когда функциональные связи между волокнами кортикоспинального тракта и спинномозговыми мотонейронами подвергаются зависимой от активности реорганизации ( 30 ).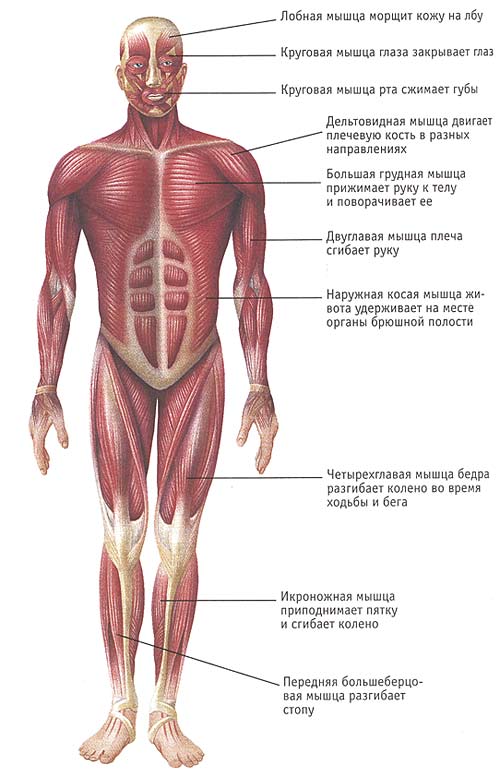 Анатомия опорно-двигательного аппарата будет ограничивать модели двигательной активности, которые могут быть выполнены.
Анатомия опорно-двигательного аппарата будет ограничивать модели двигательной активности, которые могут быть выполнены.
На анатомическую и функциональную связь между мышцами также могут влиять внешние факторы. Например, паттерны связности нисходящих путей частично определены генетически ( 31 ).Соматотопическая организация наблюдается во всей нервно-моторной системе, а структура сообщества анатомической мышечной сети отражает организацию модулей управления первичной моторной корой ( 19 ). Точно так же пространственная организация мотонейронов спинного мозга также связана с анатомической организацией мышц ( 32 ), и мышцы, которые анатомически расположены близко друг к другу, также иннервируются теми же спинными нервами (рис. S2). ( 2 ).Топографическая организация спинномозговых мотонейронов сходна для разных видов ( 33 ) и, следовательно, может быть результатом эволюционной консервации ( 34 ). Костно-мышечная анатомия и нейронные пути являются, следовательно, и с учетом своего родом генетического контроля.
Функциональная связь не полностью определялась анатомией; мы наблюдали несколько ключевых различий между анатомическими и функциональными мышечными сетями. Двусторонние модули, состоящие из мышц верхних или нижних конечностей, были ключевой характеристикой функциональной мышечной сети, которая отсутствовала в анатомической сети.Две двусторонние мышцы предплечья (поверхностный сгибатель пальцев и разгибатель пальцев) показали когерентную активность в диапазоне от 3 до 11 Гц, что согласуется с предыдущими исследованиями, показывающими бимануальное сцепление на частоте ~ 10 Гц между гомологичными мышцами кисти и предплечья ( 35 , 36 ). Наблюдаемая бимануальная связь на частотах от 3 до 11 Гц может быть вызвана оливо-церебеллярной системой, которая, как известно, производит колебания в этом частотном диапазоне и участвует в формировании функциональных мышечных коллективов ( 35 ).Двусторонние мышцы предплечья были слабо связаны с другими мышцами (рис. 2), что может отражать относительно высокую долю прямых кортикоспинальных проекций — и, таким образом, относительно низкую долю расходящихся выступов — к мотонейронам, иннервирующим мышцы рук и предплечий ( 37 ).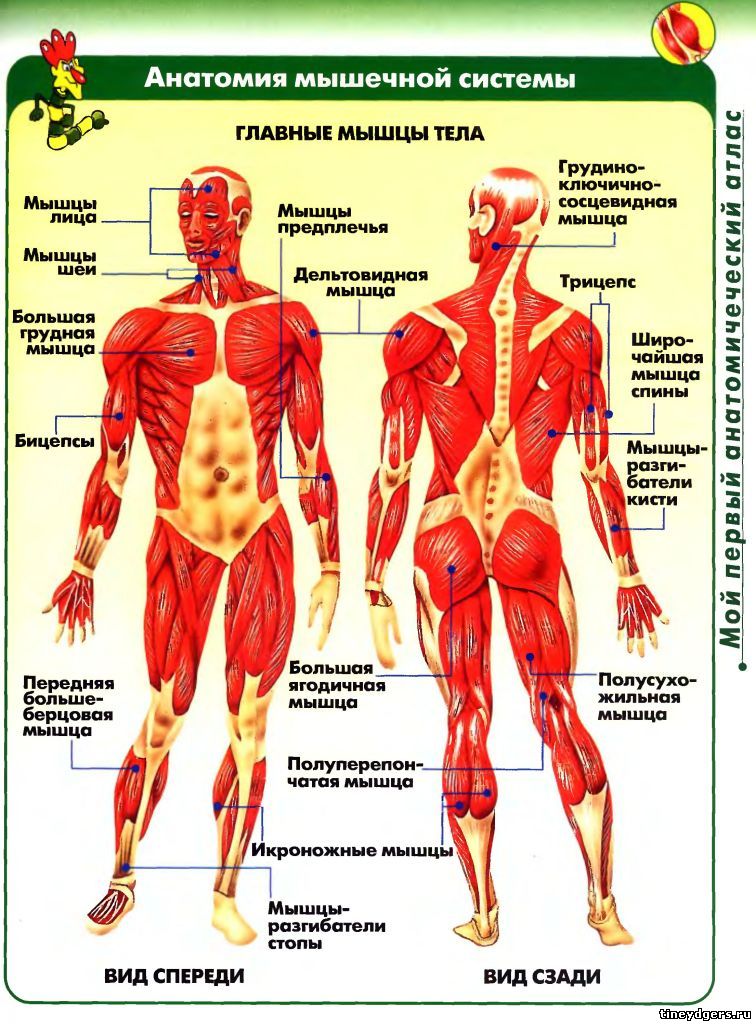
Напротив, двусторонний модуль мышц голени показал сильную связь во многих частотных диапазонах, что согласуется с предыдущими анализами функциональных мышечных сетей ( 20 ), и показал самые сильные дальнодействующие связи, наблюдаемые в настоящем исследовании (рис.3С). Двусторонняя связь между мышцами руки и ноги во время балансировки может быть создана вестибулоспинальным трактом, который, как известно, участвует в постуральной стабильности и иннервирует серое вещество спинного мозга с обеих сторон ( 21 ). Двусторонняя связь наблюдалась на всех уровнях кортикоспинальной оси ( 38 ) и имеет первостепенное значение для функциональных сетей мозга, особенно между гомологичными лево-правыми областями коры ( 39 ). Настоящие результаты предполагают, что двустороннее сцепление также является определяющим признаком функциональных мышечных сетей.Различия в функциональной связности между двусторонними мышцами руки и двусторонними мышцами ног указывают на то, что функциональная мышечная сеть, такая как анатомическая мышечная сеть ( 25 ), не демонстрирует последовательной гомологии.
Функциональная связность отображала различные зависящие от задачи модуляции, которые были связаны с задачей, которую выполняли субъекты: функциональная связность была увеличена внутри и между модулями ног во время постуральной нестабильности и увеличилась внутри и между модулями руки и верхней части тела в условиях наведения.Таким образом, функциональная связь между мышцами зависит от задачи ( 21 , 36 ), что может свидетельствовать о наличии многофункциональных цепей, в которых данный паттерн анатомической связи может генерировать различные паттерны функциональной активности в различных условиях ( 40 ). Такая распределенная схема создает основу для поддержки множества вариантов поведения, которые управляются согласованными действиями большой распределенной сети, а не простыми выделенными путями. Таким образом, лежащая в основе сетевая связность ограничивает возможные паттерны популяционной активности низкоразмерным многообразием, охватываемым несколькими независимыми паттернами — нейронными режимами — которые обеспечивают основные строительные блоки нейронной динамики и моторного контроля ( 41 ).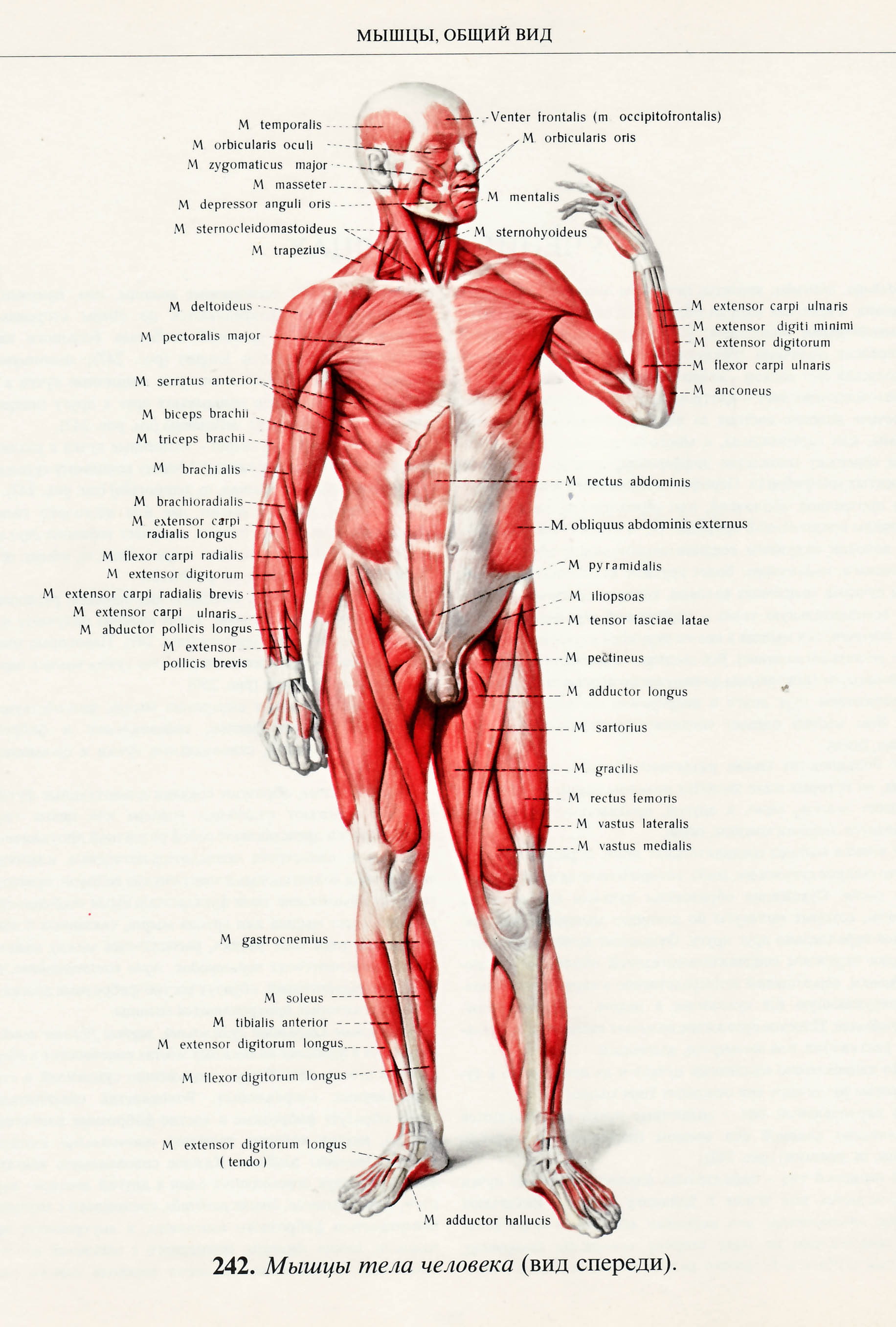 Опять же, это обнаруживает сходство с недавними исследованиями функциональных принципов когнитивных сетей в мозге ( 42 ).
Опять же, это обнаруживает сходство с недавними исследованиями функциональных принципов когнитивных сетей в мозге ( 42 ).
Изменения, зависящие от задачи, происходили с разной частотой, что указывает на функционирование организации мультиплексной сети, при этом четыре частотных компонента отражают разные типы взаимодействий между мышцами. Четыре различных частотных компонента (от 0 до 3, от 3 до 11, от 11 до 21 и от 21 до 60 Гц) были извлечены с помощью NNMF. Эти полосы частот полностью соответствуют найденным ранее ( 20 ), демонстрируя надежность этого открытия.Интересная возможность состоит в том, что эти частотные компоненты отражают спектральные отпечатки различных путей, которые проецируются на двигательные нейроны спинного мозга. Было высказано предположение, что эти разные частоты могут играть определенную роль в кодировании моторных сигналов ( 43 ). Функциональная связь на низкочастотных компонентах может быть результатом афферентных путей, в то время как функциональная связность на более высоких частотах может отражать коррелированный вход от нисходящих путей.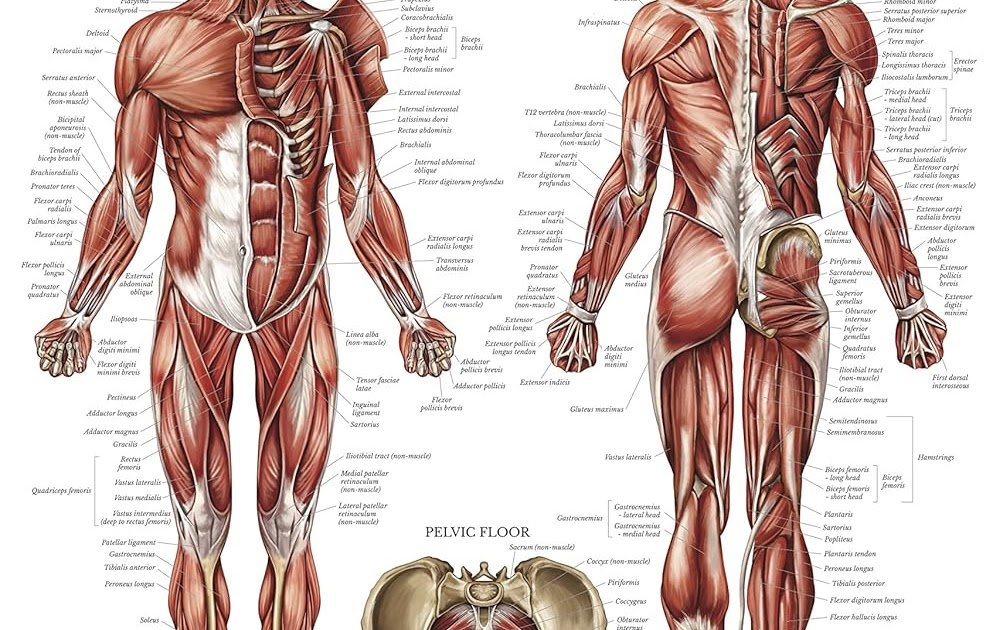
Таким образом, наш сетевой анализ выявил широкую функциональную взаимосвязь между мышцами, что свидетельствует о коррелированных входах в спинномозговые двигательные нейроны на разных частотах.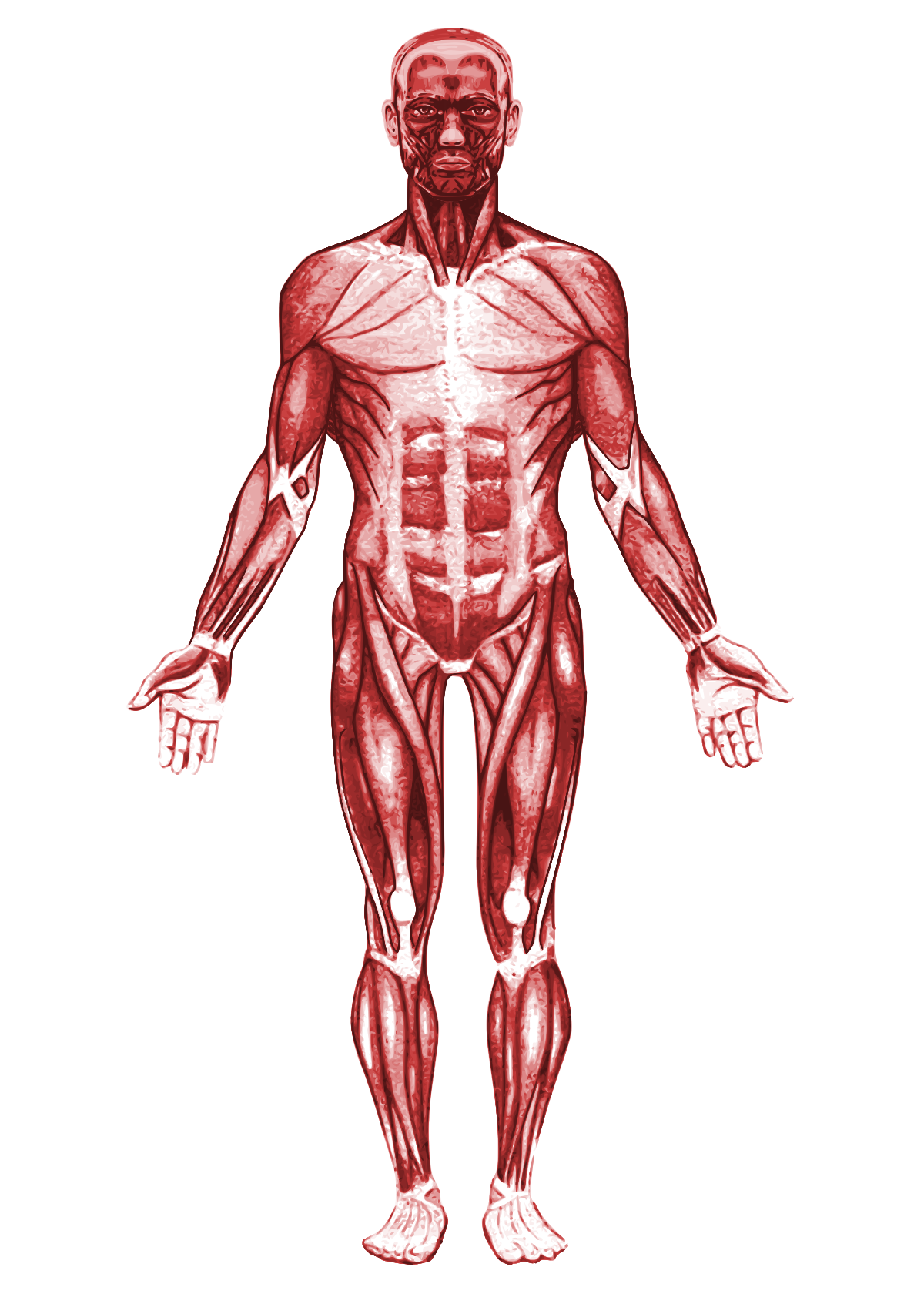 Коррелированные входные данные указывают на расходящиеся проекции или латеральные связи в нервных путях, которые иннервируют двигательные нейроны спинного мозга и, следовательно, могут использоваться для оценки спинномозговых сетей ( 23 ).Эти результаты согласуются с картированием «многие ко многим», а не с сопоставлением «один к одному» между мозгом и мышцами ( 4 ), в котором сложные движения возникают за счет относительно тонких изменений в коактивации различных распределенных функциональных режимов. Мы представляем новый подход, который совмещается движение неврологии с текущими исследованиями в области сетей мозга, показывая, как центральную нервную систему взаимодействует с костно-мышечной системой человеческого организма. Этот подход вписывается в более широкие рамки сетевой физиологии, исследуя взаимодействия мозга и тела ( 45 ).Подобно текущим результатам, исследования сетевой физиологии показали, что динамические взаимодействия между системами органов опосредуются через определенные диапазоны частот ( 46 ).
Коррелированные входные данные указывают на расходящиеся проекции или латеральные связи в нервных путях, которые иннервируют двигательные нейроны спинного мозга и, следовательно, могут использоваться для оценки спинномозговых сетей ( 23 ).Эти результаты согласуются с картированием «многие ко многим», а не с сопоставлением «один к одному» между мозгом и мышцами ( 4 ), в котором сложные движения возникают за счет относительно тонких изменений в коактивации различных распределенных функциональных режимов. Мы представляем новый подход, который совмещается движение неврологии с текущими исследованиями в области сетей мозга, показывая, как центральную нервную систему взаимодействует с костно-мышечной системой человеческого организма. Этот подход вписывается в более широкие рамки сетевой физиологии, исследуя взаимодействия мозга и тела ( 45 ).Подобно текущим результатам, исследования сетевой физиологии показали, что динамические взаимодействия между системами органов опосредуются через определенные диапазоны частот ( 46 ). Мы расширили этот подход, исследуя сетевую топологию функциональных взаимодействий между мышцами, которые опосредуются нервными путями в спинном мозге. В будущих исследованиях может быть увеличено количество исследуемых мышц, включая электроэнцефалографию для картирования сетей мозг-тело и изучение кортикального контроля мышечных сетей, а также рассмотрение индивидуальных различий в анатомии.
Мы расширили этот подход, исследуя сетевую топологию функциональных взаимодействий между мышцами, которые опосредуются нервными путями в спинном мозге. В будущих исследованиях может быть увеличено количество исследуемых мышц, включая электроэнцефалографию для картирования сетей мозг-тело и изучение кортикального контроля мышечных сетей, а также рассмотрение индивидуальных различий в анатомии.
С точки зрения системной биологии, головной и спинной мозг переплетаются с телом — они «воплощены» ( 7 ), и, таким образом, сетевой анализ мозга может быть расширен для исследования внутренней организации функциональных сетей в позвоночнике человека. шнур ( 47 ). Функциональные взаимодействия между надспинальными, спинными и периферическими регионами могут быть интегрированы с использованием сетевого анализа в качестве общей основы. Такая интегрированная структура хорошо подходит для обеспечения новых взглядов и методов лечения неврологических расстройств ( 48 ).
МАТЕРИАЛЫ И МЕТОДЫ
Сбор данных
Четырнадцать здоровых участников (7 мужчин и 7 женщин; средний возраст 25 ± 8 лет; 10 правшей и 4 левши) без каких-либо неврологических или моторных расстройств или сахарного диабета и с индекс массы тела ниже 25 были включены в это исследование. Эксперименты были одобрены Комитетом по этике Human Movement Sciences Vrije Universiteit Amsterdam (ссылка ECB 2014-78) и проводились в полном соответствии с Хельсинкской декларацией.Все участники были письменно и устно проинформированы о процедуре и подписали информированное согласие перед участием.
Участникам было предложено выполнить девять различных заданий позы. Был использован полнофакторный план, в котором устойчивость позы (нормальное положение, нестабильность в передне-заднем направлении и нестабильность в медиально-латеральном направлении) и поведение при указании (отсутствие указания, указание доминирующей рукой и указание обеими руками) варьировались. . Постуральной стабильностью манипулировали с помощью балансира с одной степенью свободы, которая позволяла двигаться в передне-заднем или медиально-латеральном направлении. В задаче наведения участники держали лазерную указку доминирующей рукой (одноручная) или двумя руками (двуручная) и наводили ее на белую мишень (25 см 2 ), расположенную на расстоянии 2,25 м, параллельно поперечной оси. ось тела на высоте акромиона участника. Таким образом, эксперимент состоял из девяти (3 × 3) экспериментальных условий. Продолжительность испытания составляла 30 с, каждое условие повторялось шесть раз.
В задаче наведения участники держали лазерную указку доминирующей рукой (одноручная) или двумя руками (двуручная) и наводили ее на белую мишень (25 см 2 ), расположенную на расстоянии 2,25 м, параллельно поперечной оси. ось тела на высоте акромиона участника. Таким образом, эксперимент состоял из девяти (3 × 3) экспериментальных условий. Продолжительность испытания составляла 30 с, каждое условие повторялось шесть раз.
Биполярная поверхностная ЭМГ была записана для 36 мышц, распределенных по всему телу (18 двусторонних мышц; Таблица 1).Мы выбрали репрезентативную группу пар антагонистических мышц, участвующих в постуральном контроле, которые можно правильно измерить с помощью поверхностной ЭМГ из-за их расположения и размера. ЭМГ регистрировали с использованием трех 16-канальных систем Porti (TMSi), фильтровали верхними частотами онлайн на 5 Гц и дискретизировали на 2 кГц.
Анатомическая мышечная сеть
Анатомическая мышечная сеть была определена путем картирования физических связей между мышцами. Узлы представляют 36 мышц (18 слева и 18 справа), а края сети представляют собой сухожильные прикрепления мышц к костям и соединительной ткани.Структурные связи были определены на основе происхождения и прикрепления мышц ( 2 ). Кости, которые не показывают или почти не двигаются в суставе между ними, считались одной жесткой костной структурой, то есть тазом, грудным скелетом или ossa cranii ( 25 ). Связи между мышцами и костями, перечисленные в таблице 2, обозначают двудольную сеть C , с мышцами как одной группой и костями как второй группой. Затем мы создали сеть, ориентированную на мышцы, как одномодовые проекции C : B = CC T ( 19 ).Это дало взвешенную матрицу смежности, где веса отражают количество прикреплений, которыми связаны две мышцы. Мы преобразовали это в бинарную сеть, установив для всех ненулевых весов значение 1.
Узлы представляют 36 мышц (18 слева и 18 справа), а края сети представляют собой сухожильные прикрепления мышц к костям и соединительной ткани.Структурные связи были определены на основе происхождения и прикрепления мышц ( 2 ). Кости, которые не показывают или почти не двигаются в суставе между ними, считались одной жесткой костной структурой, то есть тазом, грудным скелетом или ossa cranii ( 25 ). Связи между мышцами и костями, перечисленные в таблице 2, обозначают двудольную сеть C , с мышцами как одной группой и костями как второй группой. Затем мы создали сеть, ориентированную на мышцы, как одномодовые проекции C : B = CC T ( 19 ).Это дало взвешенную матрицу смежности, где веса отражают количество прикреплений, которыми связаны две мышцы. Мы преобразовали это в бинарную сеть, установив для всех ненулевых весов значение 1.
Происхождение и введение основаны на общей анатомии человека, как описано Martini et al . ( 2 ), таким образом игнорируя возможные индивидуальные различия между участниками.
( 2 ), таким образом игнорируя возможные индивидуальные различия между участниками.
Функциональная мышечная сеть
Мы отразили данные левшей, чтобы создать доминирующую и недоминантную стороны.Данные ЭМГ были предварительно обработаны для удаления артефактов движения и электрокардиографии (ЭКГ). ЭМГ подвергалась полосовой фильтрации (от 1 до 400 Гц), а анализ независимых компонентов использовался для удаления загрязнения ЭКГ ( 49 ). Для каждого участника были удалены один или два независимых компонента. Затем данные ЭМГ подвергались высокочастотной фильтрации (20 Гц) для удаления низкочастотных артефактов движения. После предварительной обработки огибающие ЭМГ были извлечены с помощью амплитуды Гильберта ( 22 ).
Мы следовали процедуре, описанной Boonstra et al .( 20 ) для извлечения функциональных мышечных сетей из поверхностной ЭМГ. Сначала была оценена комплексная когерентность и усреднена по испытаниям в каждом условии для каждого участника. Абсолютное значение когерентности было возведено в квадрат, чтобы получить согласованность в квадрате величины. Межмышечная связность оценивалась между всеми 630 парами мышц. Затем NNMF использовался для разложения этих спектров когерентности по всем комбинациям мышц, состояниям и участникам на четыре отдельных частотных компонента и соответствующие веса.Это дало набор весов для каждого частотного компонента, который определял неориентированную взвешенную сеть для каждого условия и участника.
Абсолютное значение когерентности было возведено в квадрат, чтобы получить согласованность в квадрате величины. Межмышечная связность оценивалась между всеми 630 парами мышц. Затем NNMF использовался для разложения этих спектров когерентности по всем комбинациям мышц, состояниям и участникам на четыре отдельных частотных компонента и соответствующие веса.Это дало набор весов для каждого частотного компонента, который определял неориентированную взвешенную сеть для каждого условия и участника.
Эти функциональные сети были преобразованы в бинарные сети для облегчения сравнения с анатомической сетью. Для получения минимально связной сети по условиям и частотным компонентам были выбраны веса. Эта процедура определения порога дает единственное уникальное пороговое значение, которое соответствует порогу перколяции ( 50 ).Это приводило к разреженным сетям, в которых каждый узел был подключен по крайней мере к одному другому узлу ребром на одном из уровней мультиплексной сети.
Структура сообщества
Для извлечения модулей из анатомических сетей использовался алгоритм Лувена. Поскольку алгоритм Лувена является стохастическим, мы использовали консенсусную кластеризацию, чтобы получить стабильное разбиение на 1000 итераций ( 51 ). Мультиплексный анализ модульности ( 24 ) использовался для идентификации модулей функциональной мышечной сети в зависимости от условий и частотных компонентов.Мы использовали MolTi, автономное графическое программное обеспечение, для обнаружения сообществ в мультиплексных сетях путем оптимизации модульности мультиплекса с помощью адаптированного алгоритма Лувена (https://github.com/gilles-didier/MolTi). Модули были извлечены по 36 двоичным сетям (9 × 4). Мы использовали индекс Rand и скорректированный индекс Rand для сравнения модулей анатомических и функциональных мышечных сетей ( 16 ).
Сравнение функциональных сетей в разных условиях
Чтобы упростить сравнение функциональных сетей в разных условиях задачи, мы грубо обработали сети ( 52 ).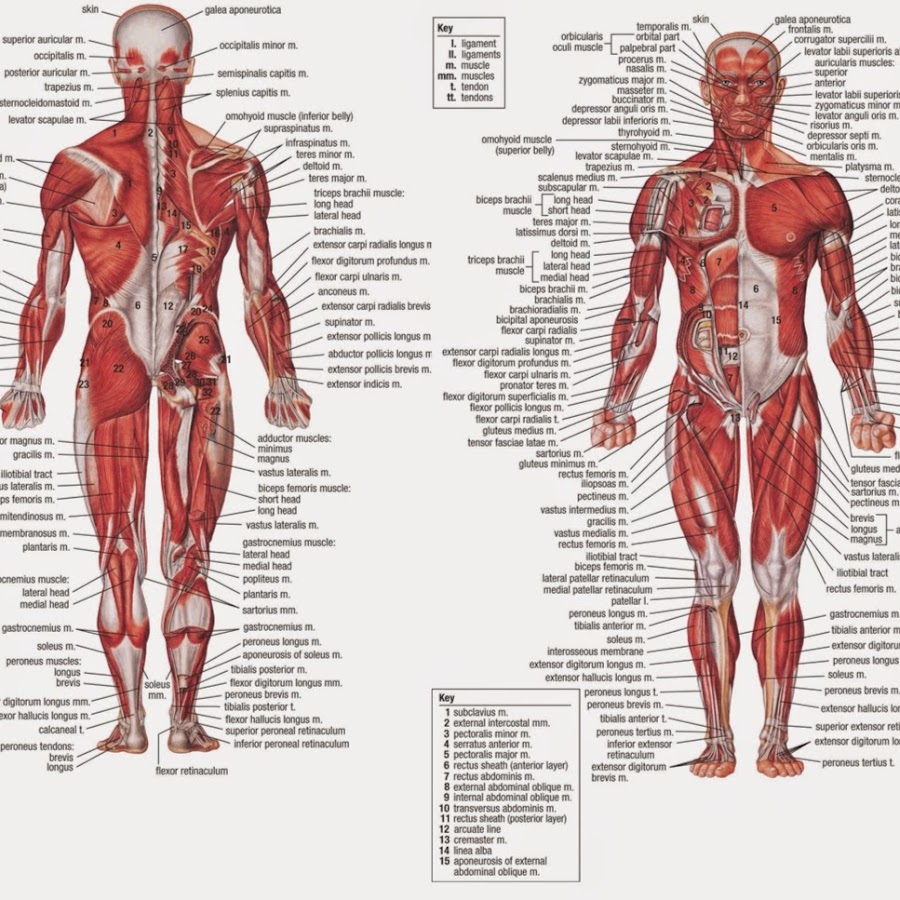
Для сравнения сгруппированных сетей по условиям мы использовали простые контрасты между условиями задач и количественные различия в количестве соединений между модулями и внутри них. Мы проверили четыре контраста: (i) одноручное и (ii) бимануальное наведение по сравнению с отсутствием наведения и (iii) передне-заднее и (iv) медиально-латеральную нестабильность по сравнению с нормальным стоянием. Чтобы проверить статистическую значимость этих контрастов, мы выполнили парные перестановочные тесты отдельно для каждого из элементов матрицы ( 52 ). Кластерные сети имели гораздо меньшую размерность по сравнению с исходными функциональными мышечными сетями (21 ребро вместо 630). Контроль ошибок на уровне семьи поддерживался с помощью поправки Бонферрони для исправления множественных сравнений (4 × 21 = 84 сравнения).
Кластерные сети имели гораздо меньшую размерность по сравнению с исходными функциональными мышечными сетями (21 ребро вместо 630). Контроль ошибок на уровне семьи поддерживался с помощью поправки Бонферрони для исправления множественных сравнений (4 × 21 = 84 сравнения).
ДОПОЛНИТЕЛЬНЫЕ МАТЕРИАЛЫ
Дополнительные материалы к этой статье доступны по адресу http://advances.sciencemag.org/cgi/content/full/4/6/eaat0497/DC1
раздел S1. Альтернативная анатомическая мышечная сеть
сечение S2.Сеть спинномозговых нервов
участок S3. Весовые функциональные сети
рис. S1. Матрица смежности анатомических мышечных сетей.
рис. S2. Структура сообщества спинномозговой нервной сети.
рис. S3. Структура сообщества взвешенной функциональной сети.
стол S1. Иннервация спинномозгового нерва.
Это статья в открытом доступе, распространяемая в соответствии с условиями некоммерческой лицензии Creative Commons Attribution, которая разрешает использование, распространение и воспроизведение на любом носителе, при условии, что в результате будет использовано , а не для коммерческих целей и при условии, что оригинальная работа правильно процитирована.
ССЫЛКИ И ПРИМЕЧАНИЯ
- ↵
К. Шеррингтон, Интегративное действие нервной системы (архив CUP, 1910).
- ↵
Ф. Х. Мартини, М. Дж. Тиммонс, М. П. МакКинли, Анатомия человека (3-е издание) (Прентис Холл, 2000).
- ↵
E. Pierrot-Deseilligny, D. Burke, Схема человеческого спинного мозга: его роль в двигательном контроле и двигательных расстройствах (Cambridge Univ. Press, 2005).
- ↵
- ↵
N.Бернштейн, Координация и регулирование движений (Пергамон, 1967).
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- 000
- 000
- 000
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵ 9000 9000
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- 000
- 000
- 000Н.
 Макаров, Г. М. Ноэтчер, А. Назарян, Низкочастотное электромагнитное моделирование электрических и биологических систем с использованием MATLAB (Wiley, 2015), стр. 89–130.
Макаров, Г. М. Ноэтчер, А. Назарян, Низкочастотное электромагнитное моделирование электрических и биологических систем с использованием MATLAB (Wiley, 2015), стр. 89–130. - ↵
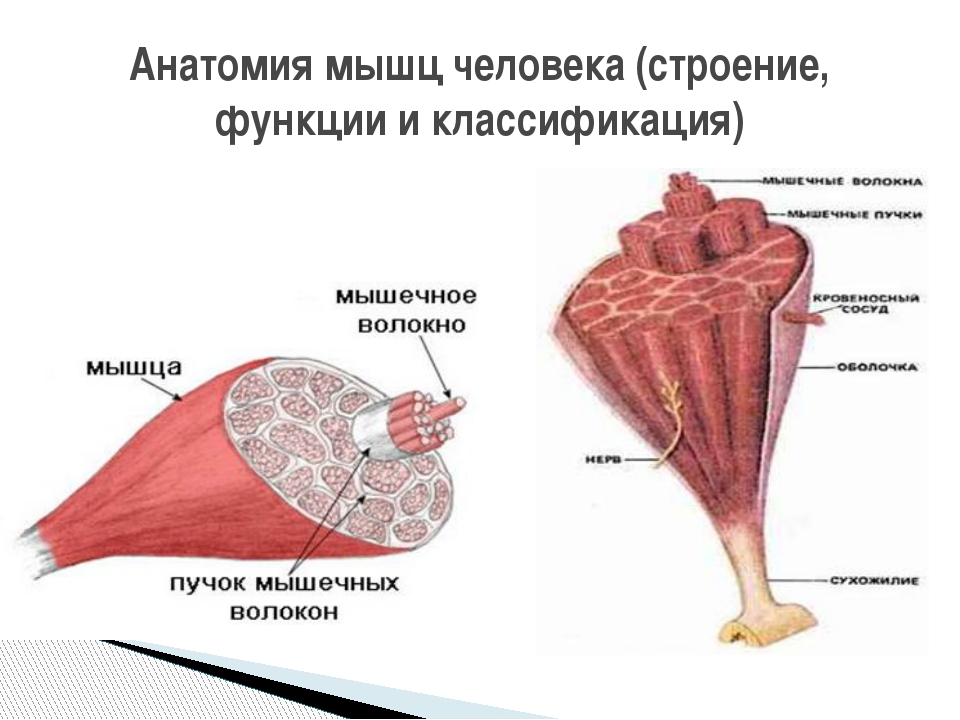 J.N.K. получил данные ЭМГ. J.N.K. и T.W.B. провели оценку анатомической сети. J.N.K., L.L.G. и T.W.B. провел оценку функциональной сети. T.W.B. провели статистический анализ. J.N.K. и T.W.B. написал рукопись.Все авторы редактировали рукопись. Конкурирующие интересы: Авторы заявляют, что у них нет конкурирующих интересов. Доступность данных и материалов: Все данные, необходимые для оценки выводов в статье, присутствуют в документе и / или дополнительных материалах. Дополнительные данные, относящиеся к этой статье, могут быть запрошены у авторов. Данные ЭМГ, использованные в этом исследовании, доступны на сайте Zenodo (http://doi.org/10.5281/zenodo.1185196).
J.N.K. получил данные ЭМГ. J.N.K. и T.W.B. провели оценку анатомической сети. J.N.K., L.L.G. и T.W.B. провел оценку функциональной сети. T.W.B. провели статистический анализ. J.N.K. и T.W.B. написал рукопись.Все авторы редактировали рукопись. Конкурирующие интересы: Авторы заявляют, что у них нет конкурирующих интересов. Доступность данных и материалов: Все данные, необходимые для оценки выводов в статье, присутствуют в документе и / или дополнительных материалах. Дополнительные данные, относящиеся к этой статье, могут быть запрошены у авторов. Данные ЭМГ, использованные в этом исследовании, доступны на сайте Zenodo (http://doi.org/10.5281/zenodo.1185196).- Copyright © 2018 Авторы, некоторые права защищены; обладатель исключительной лицензии Американской ассоциации содействия развитию науки.Нет претензий к оригинальным работам правительства США. Распространяется по некоммерческой лицензии Creative Commons Attribution 4.0 (CC BY-NC).
Анатомия и структура ткани скелетных мышц
В человеческом теле есть 3 типа мышечной ткани: скелетная, гладкая и сердечная. В этом видео и статье я собираюсь обсудить скелетной мышечной ткани .
В этом видео и статье я собираюсь обсудить скелетной мышечной ткани .
Ткань скелетных мышц
Скелетные мышцы чаще всего прикрепляются к костям и помогают двигать телом.В отличие от двух других типов мышечной ткани, скелетные мышцы сокращаются на произвольной основе, через соматическую нервную систему , , что позволяет вам двигать телом по желанию.
Скелетные мышцы также выполняют важные функции, такие как поддержка осанки, защита нежных органов, и они даже выделяют тепло во время сокращения, что помогает телу поддерживать надлежащую температуру.
Структура скелетных мышц
Каждая скелетная мышца считается органом и состоит из слоев соединительной ткани, мышечных волокон, кровеносных сосудов и нервов.Скелетные мышцы прикрепляются к костям через сухожилия или посредством прямого прикрепления.
Если вы посмотрите на эту схему мышц, вы заметите внешний слой соединительной ткани, который называется эпимизий .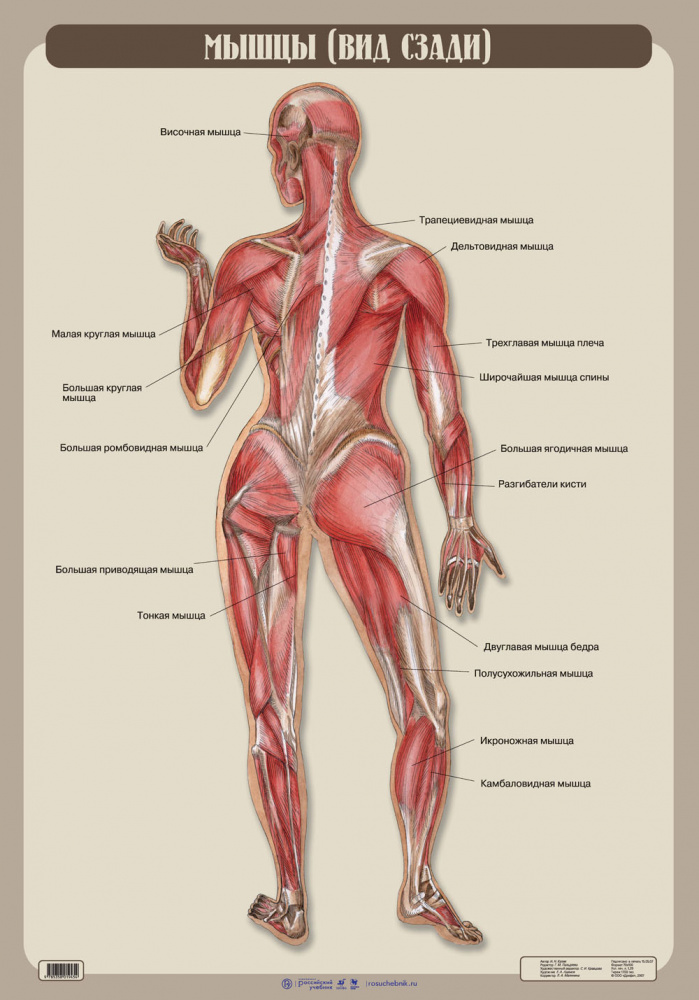 Приставка «эпи» означает на или над (эпидермис — это слой на вашей коже), а «mysium» происходит от греческого слова, которое означает «мышца». Следовательно, эпимизий представляет собой слой соединительной ткани, который покрывает или покрывает весь мышечный орган.
Приставка «эпи» означает на или над (эпидермис — это слой на вашей коже), а «mysium» происходит от греческого слова, которое означает «мышца». Следовательно, эпимизий представляет собой слой соединительной ткани, который покрывает или покрывает весь мышечный орган.
Затем вы заметите, что мышечные волокна сгруппированы в нечто, называемое пучками , что означает «пучки».Эти пучки окружены соединительной тканью, которая называется перимизий и . «Пери» означает «вокруг», и снова «mysium» относится к мышцам. Итак, перимизий находится вокруг пучков, которые связывают эти мышечные волокна.
Внутри пучков другой слой соединительной ткани, называемый эндомизием , окружает отдельные мышечные клетки. «Эндо» означает «внутри», это поможет вам вспомнить, что он окружает отдельные мышечные волокна внутри пучка.
Мышечные волокна
Теперь давайте взглянем на отдельные мышечные клетки, которые называются мышечными волокнами . Эти волокна длинные, цилиндрические и содержат несколько ядер. Эти мышечные волокна обернуты клеточной мембраной, называемой сарколемма .
Эти волокна длинные, цилиндрические и содержат несколько ядер. Эти мышечные волокна обернуты клеточной мембраной, называемой сарколемма .
Внутри каждого мышечного волокна есть крошечные стержни, называемые миофибриллами, которые окружены саркоплазмой. Эти миофибриллы, также называемые фибриллами, состоят из повторяющихся сегментов, называемых саркомерами, которые представляют собой крошечные единицы, ответственные за сокращение скелетных мышц.
Структура саркомера
Когда мы более подробно рассмотрим структуру саркомера, вы заметите эти зигзагообразные секции, которые отмечают конечную точку каждого саркомера.Они называются дисками Z или линиями Z , и они позволяют прикреплять тонкие (актиновые) филаменты, а также эластичный белок, называемый тайтином.
Каждый саркомер содержит тонких (актиновых) волокон и толстых (миозиновых) волокон . Тонкие (актиновые) филаменты, представленные ниже синим цветом, прикрепляются к Z-диску.
Эти толстые (миозиновые) филаменты, представленные ниже красным цветом, прикрепляются к эластичному, упругому белку, называемому тайтин , который затем прикрепляется к Z-диску.Актиновые и миозиновые нити взаимодействуют во время сокращения мышц, о чем я расскажу чуть позже. «М-линии» или «М-полосы» закрепляют центр миозиновых волокон, удерживая их вместе, одновременно действуя как амортизатор.
Повязки и зоны саркомера
Чтобы помочь нам понять части саркомера, анатомы делят срезы на полосы или зоны. Расположение нитей внутри этих лент объясняет вид волокон скелетных мышц полосатым, (полосатым).Это важная характеристика скелетных и сердечных мышц , которую вы должны запомнить: они обе содержат полосок .
- Полоса : Во-первых, на каждом саркомере имеется «полоса A », которая представляет собой участок, который содержит всю длину толстой миозиновой нити вместе с перекрывающимися частями тонких актиновых нитей.
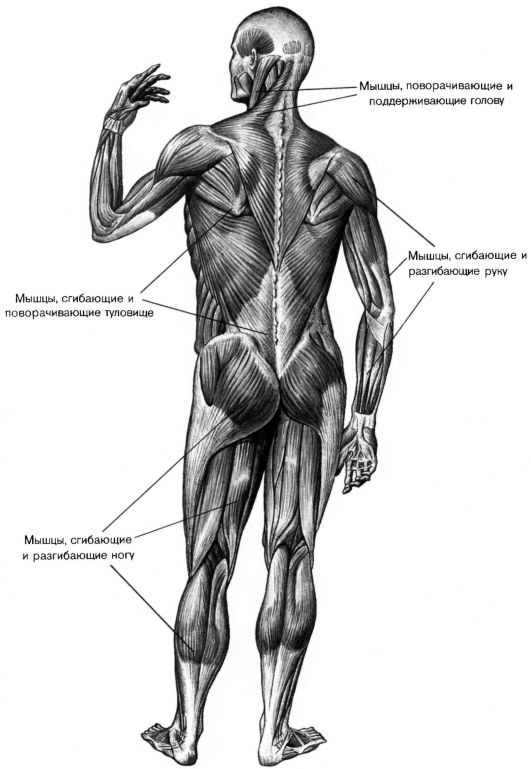 Этот участок составляет темную часть рисунка из полосок.
Этот участок составляет темную часть рисунка из полосок. - I band : « I band » — это часть саркомера, которая окружает Z-диск и содержит только тонкие (актиновые) нити.Этот участок составляет более светлую полосу рисунка.
- Зона H : Зона H — это участок внутри полосы A, который состоит из толстых миозиновых нитей и встроенных в них M-линий. В расслабленном состоянии в этой части саркомера нет тонких нитей.
- Z-диск : И снова Z-диск — это зигзагообразная часть, которая отмечает конец каждого саркомера и позволяет прикреплять актиновые нити и тайтин.
Сокращение скелетных мышц
Во время сокращения мышцы толстые (миозиновые) нити, расположенные внутри саркомера, изгибаются, и узловатая головная часть прикрепляется к тонким актиновым нитям, сдвигая их к средней линии саркомера.Это скольжение нитей заставляет саркомер укорачиваться или сокращаться. Поскольку это происходит вдоль всех саркомеров внутри миофибрилл, сокращается все мышечное волокно, что в конечном итоге приводит к сокращению или сокращению всего мышечного органа.
Это известно как теория скользящих волокон мышечного сокращения.
Бесплатные викторины и другие видео по анатомии
Пройдите бесплатный тест по анатомии скелетных мышц, чтобы проверить свои знания, или просмотрите наше видео о скелетных мышцах.Кроме того, вы можете посмотреть наши лекции по анатомии и физиологии на YouTube или проверить наши заметки по анатомии и физиологии.
Ультраструктура мышцы — скелет — скользящая нить
Мышечная ткань имеет уникальный гистологический вид, который позволяет ей выполнять свою функцию. Существует три основных типа мышц:
- Скелет — поперечно-полосатая мышца, которая находится под произвольным контролем соматической нервной системы. Отличительные признаки — цилиндрические клетки и множественные периферические ядра.
- Сердечная — поперечно-полосатая мышца, которая находится только в сердце. Отличительными признаками являются отдельные ядра и наличие интеркалированных дисков между клетками.

- Гладкая — мышца без поперечно-полосатой полосы, которая непроизвольно контролируется вегетативной нервной системой. Отличительным признаком является наличие одного веретенообразного центрального ядра на клетку.
В этой статье мы рассмотрим гистологию скелетной мышцы — ее состав, гистологический вид и клинические корреляции.
Состав скелетных мышц
Мышечная клетка очень специализирована для своего назначения. Одиночная клетка образует одно мышечное волокно, а ее клеточная мембрана известна как сарколемма .
Т-канальцы уникальны для мышечных клеток. Это инвагинации сарколеммы, которые проводят заряд, когда клетка деполяризована.
Мышечные клетки также имеют специализированный эндоплазматический ретикулум — он известен как саркоплазматический ретикулум и содержит большой запас ионов кальция.
Мышцы также имеют сложную опорную структуру из соединительной ткани. Каждое мышечное волокно окружено тонким слоем соединительной ткани, известной как endomysium . Затем эти волокна группируются в пучки, известные как пучки , которые окружены слоем соединительной ткани, известным как перимизий . Многие пучки составляют мышцу, которая, в свою очередь, окружена толстым слоем соединительной ткани, известной как эпимизий .
Каждое мышечное волокно окружено тонким слоем соединительной ткани, известной как endomysium . Затем эти волокна группируются в пучки, известные как пучки , которые окружены слоем соединительной ткани, известным как перимизий . Многие пучки составляют мышцу, которая, в свою очередь, окружена толстым слоем соединительной ткани, известной как эпимизий .
Ультраструктурный вид скелетных мышц
Поперечно-полосатый вид волокон скелетных мышц обусловлен организацией двух сократительных белков: актина (тонкая нить) и миозина (толстая нить).
Функциональная единица сокращения в волокне скелетных мышц — это саркомер , который проходит от линии Z до линии Z. Саркомер разбит на несколько частей:
- Z-линия — где актиновые филаменты закреплены.
- Линия M — где закреплены миозиновые нити.

- I полоса — содержит только актиновые филаменты.
- Полоса — длина миозиновой нити, может содержать перекрывающиеся актиновые нити.
- H зона — содержит только миозиновые нити.
Полезное сокращение — MHAZI — линия M находится внутри зоны H, которая находится внутри диапазона A, тогда как линия Z находится внутри диапазона I.
Рис. 2. Саркомер измеряется от линии Z до линии Z [/ caption]Скользящая нить, модель
Модель скользящей нити описывает механизм сокращения скелетных мышц
Актин и миозин
Мышечные волокна образуются из двух сократительных белков — актина и миозина.
Миозиновые филаменты имеют множество головок, которые могут связываться с участками актинового филамента. Актиновые филаменты связаны с двумя другими регуляторными белками, тропонином и тропомиозином. Тропомиозин — это длинный белок, который проходит вдоль актинового филамента и блокирует сайты связывания миозиновой головки.
Тропомиозин — это длинный белок, который проходит вдоль актинового филамента и блокирует сайты связывания миозиновой головки.
Тропонин — это небольшой белок, который связывает тропомиозин с актином. Он состоит из трех частей:
- Тропонин I — связывается с актиновым филаментом.
- Тропонин Т — связывается с тропомиозином.
- Тропонин C — может связывать ионы кальция.
Муфта возбуждения-сжатия
Уникальная структура тропонина является основой связи возбуждения-сжатия :
- Когда деполяризация происходит в нервно-мышечном соединении , она проводится вниз по t-канальцам, вызывая огромный приток ионов кальция в саркоплазму из саркоплазматического ретикулума.
- Этот кальций связывается с тропонином C , вызывая изменение конформации, которое перемещает тропомиозин от участков связывания миозиновой головки актиновых филаментов.
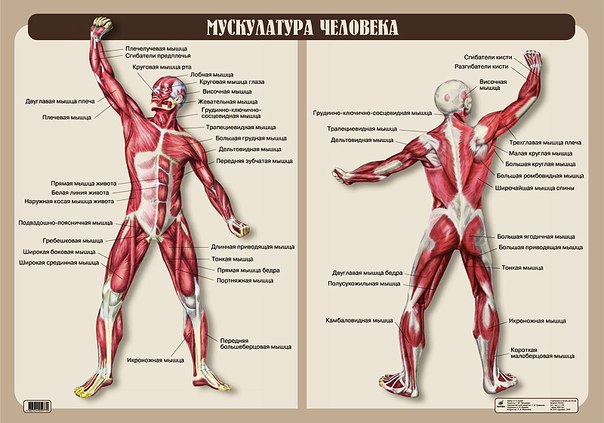
- Это позволяет головке миозина связываться с актином, образуя поперечную связь. Затем происходит силовой ход , когда миозиновые головки поворачиваются в «гребном движении», перемещая актин мимо миозина к линии M .
- Затем АТФ связывается с головкой миозина, заставляя ее отсоединяться от актина и позволяя процессу повторяться.
Следовательно, при сокращении длина волокон не меняется. Однако длина саркомера уменьшается из-за скольжения актиновых филаментов по миозину.Зона H и полоса I укорачиваются, в то время как полоса A остается той же длины. Это сближает линии Z и приводит к уменьшению общей длины саркомера.
Рис. 3. Модель мышечного сокращения со скользящей нитью. [/ caption][старт-клиника]
Клиническая значимость — мышечная дистрофия Дюшенна
Мышечная дистрофия Дюшенна — это рецессивное Х-сцепленное генетическое заболевание, при котором не продуцируется дистрофин , белок, который прикрепляет сарколемму к миофиламентам.
Это приводит к разрыву мышечных волокон при сокращении, вызывая прогрессирующую мышечную слабость и истощение.
Заболевание имеет раннее начало, пациенты часто становятся инвалидами к 12 годам.
[окончание клинической]
Структура мышечной системы
Мышечная система представляет собой сложную совокупность тканей, каждая из которых имеет свое назначение. Мышечную систему часто рассматривают вместе со скелетной системой и называют мышечно-скелетной системой.Понимание компонентов мышечной системы, включая различные типы соединительных тканей, — хороший способ понять, как работают тела и физические движения.
Скелетные мышцы
Скелетные мышцы и ткань скелетных мышц — это структуры, которые приходят в голову большинству людей, когда они думают о «мышцах». Ткань скелетных мышц — это грубая мышечная ткань, оптимизированная для сокращения и движения частей тела. Ткани скелетных мышц часто связаны с частями мышечной системы, которые находятся под нашим сознательным контролем. Плотное снабжение кровеносных сосудов и нервов в скелетной ткани способствует сокращению, которое натягивает сухожилия и кости, вызывая движение скелета.
Плотное снабжение кровеносных сосудов и нервов в скелетной ткани способствует сокращению, которое натягивает сухожилия и кости, вызывая движение скелета.
Гладкие мышцы
Гладкие мышечные ткани часто связаны с подсознательным контролем над различными системами в организме. Гладкая мышечная ткань связана с желудком, кишечником и мочевыводящей системой. Гладкие мышечные ткани контролируют кровоток в основных органах и играют ключевую роль в регулировании артериального давления.
Жесткие сухожилия
Сухожилия соединяют скелетные мышцы с костями, которые будут перемещать мышцы.Сухожилия прочные и гибкие, они очень устойчивы к разрывам и поломкам. Однако при чрезмерном растяжении связки могут быть растянуты или повреждены. Поскольку сухожилия не заживают легко, обратитесь к врачу за травмированным сухожилием. Спортсмены склонны к разрыву ахиллова сухожилия, которое соединяет икроножную мышцу с костью в пятке. Слезы могут быть довольно болезненными и могут потребовать хирургического вмешательства.
Слезы могут быть довольно болезненными и могут потребовать хирургического вмешательства.
Гибкие связки
Связка — это волокнистый материал, соединяющий две кости, которые перемещаются через скелетные мышцы.Они обеспечивают устойчивость костей как при движении скелетными мышцами, так и во время отдыха. Связки тесно связаны с мышечной системой. Совместное функционирование и диапазон движений контролируются связками. В колене находится множество связок. Часто бывает, что колено травмирует переднюю крестообразную связку (ПКС) во время резких остановок или поворотов.
Жировая ткань
Жировая ткань — еще один ключевой компонент скелетно-мышечной системы. Жировая ткань — это соединительная ткань, которая хранит энергию в виде жировых клеток и смягчает суставы.Это важно для защиты суставов во время напряженных сокращений скелетных мышц, например, при занятиях спортом и других тяжелых физических упражнениях. Сердце и другие основные органы защищены жировой тканью. Слишком много жировой ткани увеличивает риск таких проблем, как диабет.
Сердце и другие основные органы защищены жировой тканью. Слишком много жировой ткани увеличивает риск таких проблем, как диабет.
Анатомия мышц — объяснение структуры скелетных мышц
Хотя клетки скелетных мышц бывают разных форм и размеров, основная структура клетки скелетных мышц остается неизменной. Если бы вы взяли одну целую мышцу и разрезали ее, вы бы обнаружили, что мышца покрыта слоем соединительной мышечной ткани, известной как эпимизий.
Видео по анатомии мышц
Скопируйте и вставьте этот HTML-код на свою веб-страницу, чтобы встроить его.
Эпимизий защищает мышцу от трения о другие мышцы и кости. Он также продолжается в конце мышцы, образуя (вместе с другими соединительными тканями) сухожилие мышцы. Глядя на поперечное сечение мышцы, вы можете увидеть пучки волокон, известные как фасцикулы, которые окружены другой соединительной тканью, называемой перимизием.
Каждый Fasciculi содержит от 10 до 100 мышечных волокон, в зависимости от рассматриваемой мышцы. Большая сильная мышца, такая как мышцы, образующие четырехглавую мышцу, будет иметь большое количество волокон в каждом пучке. Мышцы меньшего размера, используемые для точных движений, например, в руке, будут содержать гораздо меньше волокон на фасцикулы.
Большая сильная мышца, такая как мышцы, образующие четырехглавую мышцу, будет иметь большое количество волокон в каждом пучке. Мышцы меньшего размера, используемые для точных движений, например, в руке, будут содержать гораздо меньше волокон на фасцикулы.
Мышечные волокна
Если внимательно посмотреть на каждое мышечное волокно, можно увидеть, что они тоже покрыты волокнистой соединительной тканью, известной как эндомизий, которая изолирует каждое мышечное волокно.Мышечные волокна могут иметь диаметр от 10 до 80 микрометров и длину до 35 см.
Под эндомизием и вокруг мышечного волокна находится сарколемма, которая представляет собой клеточную мембрану волокон, а под ней находится саркоплазма, цитоплазма клетки, студенистая жидкость, заполняющая большинство клеток.
Он содержит гликоген и жиры для получения энергии, а также митохондрии, которые являются электростанциями клетки, внутри которых вырабатывается энергия клетки.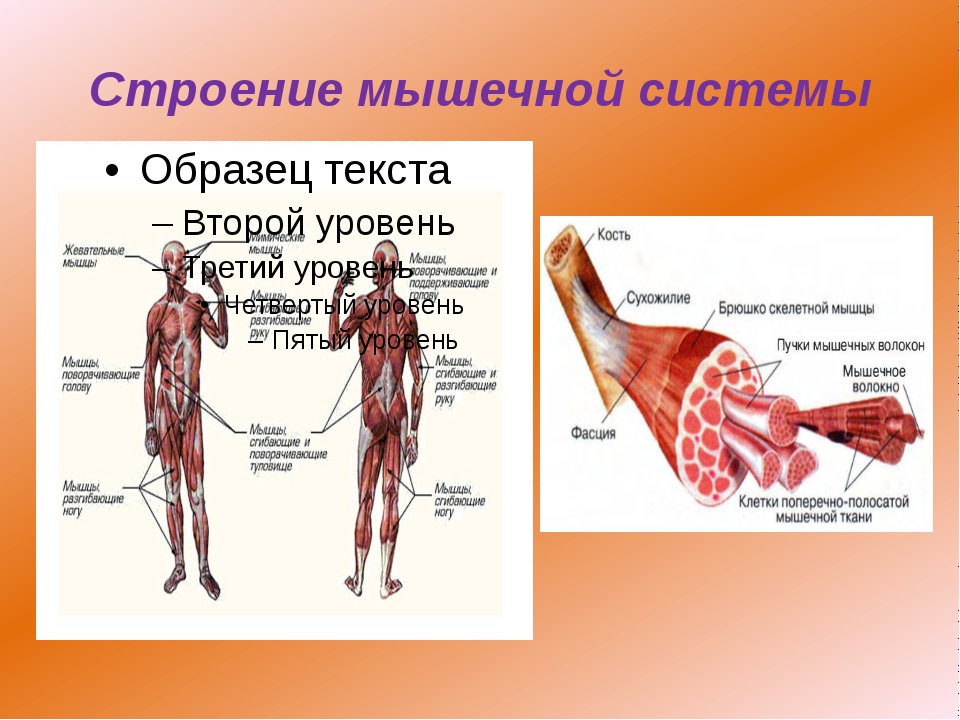
Миофибриллы
Каждое мышечное волокно само содержит цилиндрические органеллы, известные как миофибриллы.Каждое мышечное волокно содержит от сотен до тысяч миофибрилл. Это связки белков актина и миозина, которые проходят по всей длине мышечного волокна и играют важную роль в сокращении мышц.
Миофибриллы окружены сетью канальцев и каналов, называемой саркоплазматической сеткой, в которой хранится кальций, который важен для сокращения мышц. Поперечные канальцы проходят внутрь от сарколеммы через миофибриллы, по которым проходят нервные импульсы.
Каждая миофибрилла затем может быть разбита на функциональные повторяющиеся сегменты, называемые саркомерами.Для получения дополнительной информации о саркомерах и сокращении мышц ознакомьтесь с теорией скользящих волокон.
Тесты по анатомии мышц
медицинских иллюстраций мышечной системы | Готово к созданию и лицензированию
Медицинские и анатомические иллюстрации мышечной системы
В этой студии медицинского искусства работают художники, прошедшие подготовку в области анатомии, которая позволяет нам точно иллюстрировать анатомию мышц человека. Создание иллюстраций, которые не только реалистичны, но и соответствуют научным стандартам и анатомической точности.
Создание иллюстраций, которые не только реалистичны, но и соответствуют научным стандартам и анатомической точности.
Мы показываем, что по сути является невидимой анатомией под кожей, создавая медицинскую иллюстрацию, которая может быть ключевым элементом, чтобы помочь тем, кто преподает анатомию и естественнонаучное образование. Медицинские иллюстрации, медицинские визуальные эффекты, подобные этим, также могут сосуществовать с письменными медицинскими данными или выступать в качестве самостоятельных изображений, чтобы помочь передать идею анатомического образования.
Наши медицинские иллюстрации красочные и привлекательные и, следовательно, визуально привлекательные, они помогают продвигать любой тип письменного медицинского текста, публикации анатомии и используются во многих медицинских и научных областях образования и маркетинга.
Команда medical artist.com работает полный рабочий день в качестве поставщика коммуникаций в области медицины и науки, и вы можете связаться с нами в любое время, чтобы обсудить любые требования к проекту.