
Как устроены мышцы? И за счет чего они растут / Хабр
Пандемия заставила нас вести менее подвижный образ жизни. Мы закрылись дома, перестали бегать по утрам (я не бегал, но вдруг, в отличие от меня у вас были на это силы). Это поспособствовало накоплению запасов к зиме (или к лету, если вы живете в Австралии), и особенно ударило по тем, кто пытается держать себя в форме. В эти липофильные (буквально — сродство к жирам) времена мы начинаем чаще задумываться о том, что пора бы заняться какой-нибудь двигательной активностью даже не выходя из дома: покачать пресс, поотжиматься, скачать наконец фитнесс приложение (о них подробнее тут), или пойти в зал — это для совсем бесстрашных. В связи с этим мне хотелось бы поговорить о нескольких вещах, которые важно знать, чтобы лучше понимать, как тренировки воздействуют на наше тело и почему к одним нагрузкам оно хорошо приспособлено, а к другим — нет.В этой статье мы поговорим о мышцах, о том какие они бывают и за счет чего растут
Строение мышечной ткани
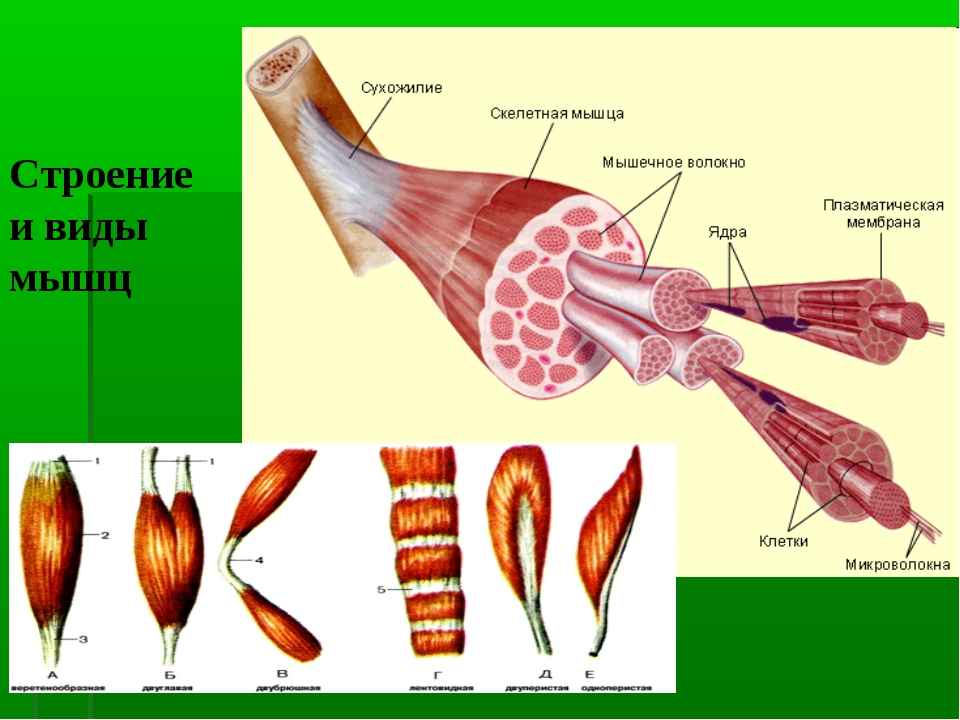
Мышцы относительно сложно устроены. Они представляют из себя совокупность мышечных волокон, объединённых в пучки, покрытые соединительной тканью (перимизием). Все вместе пучки окружены плотной оболочкой из соединительной ткани (эпимизием). При этом перимизий не только отделяет один пучок от другого, но и соединяет их с эпимизием. Обе эти оболочки достаточно плотные. В каждом пучке находятся обособленные мышечные волокна, каждое из которых покрыто рыхлой, куда менее плотной соединительной тканью (эндомизием). Эндомизий как бы связывает мышечные волокна внутри пучка. Артерии, проходя через эпимизий начинают ветвится в перимизии, распадаясь на отдельные капилляры в эндомизии.
На рисунке хорошо видно, что большую часть мышечной клетки занимают сократительные структуры, однако базовые органеллы, такие как ядра, эндоплазматический ретикулум тоже присутствуют. Митохондрии, увы не нарисованы, но они там тоже есть.
Какие бывают мышцы?
Существует несколько классификаций мышц: по форме, числу головок, положению, месту прикрепления и направлению мышечных пучков.
Остановимся на классификации мышц по направлению мышечных пучков, так как именно она обьясняет достаточно сильное отличие в силовых возможностях мышц (а это нас и интересует).
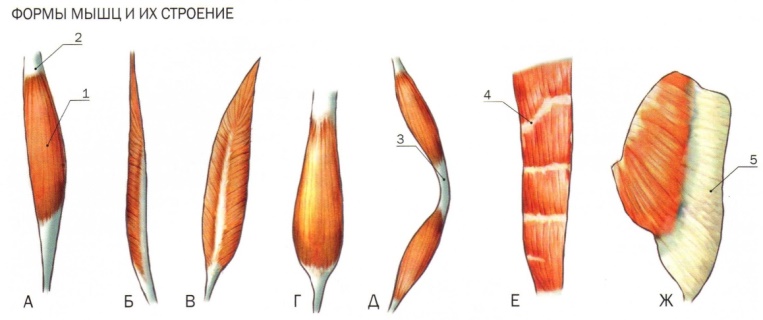
В веретенообразных мышечных пучках волокна расположены параллельно длинной оси мышцы (например, бицепс). При перистом расположении мышечные волокна расположены под углом к длинной оси (идеальные примеры — икроножная и камбаловидная мышцы). Давайте посмотрим как это выглядит.
Слева — веретенообразная мышца, справа — двуперистая
За счет перистого строения в одной мышце удается упаковать куда больше мышечных волокон одинакового объема, чем в веретенообразных мышцах того же диаметра. Соответственно, мышцы с перистым расположением волокон обладают куда большей «силой тяги».
Тут замечательный пример — икроножная и камбаловидная мышцы. За счет своего перистого строения они в 6 и, соответственно, 12 раз сильнее веретеновидных мышц аналогичного диаметра. Это и логично, ведь им необходимо поднимать вес всего тела при каждом новом шаге.
Однако, у перистых мышц есть и существенный недостаток. За счет того, что волокна расположены под углом к длинной оси мышцы, сама мышца сокращается меньше чем отдельное волокно. По сути, изменение длины всей мышцы при сокращении равняется изменению длины волокна, умноженному на косинус угла перистости. Чаще всего угол перистости находится в диапазоне от 2 до 27 градусов. Камбаловидная мышца, расположенная прямо под икроножной, имеет угол перистости в 27 градусов (cos = 0.89). Соответственно, при сокращении мышечных волокон внутри камбаловидной мышцы на x см, реально длина мышцы сократится на 0.
Иначе говоря, перистые мышцы нужны там, где речь идет о преодолении большой силы на малом пути. Например, при подъеме на носочки амплитуда движения небольшая (если сравнивать ее с разгибанием/сгибанием руки). У нас нет прямой необходимости вставать на носочки с очень большой скоростью, если, конечно, вы не увлекаетесь балетом. Однако, в целом вставать на носочки нам приходится довольно часто. Соответственно, мышцы, которые отвечают за подъем, должны поднимать вес всего тела, пусть даже и в ущерб скорости. Сгибателям и разгибателям рук тоже нужно быть сильными, но им точно нельзя жертвовать скоростью, чтобы первым дотянуться до яблока на дереве или оттолкнуть хищника (ну, эволюционно так сложилось). Поэтому, там, где нужно действовать оперативно, тело чаще использует веретенообразные мышцы.
Быстрые и медленные мышечные волокна
В одной мышце сосуществует несколько типов волокон, которые отличится по таким параметрам, как скорость, сила сокращения и утомляемость. Причина этого лежит в различиях метаболических процессов и в отличиях сократительных элементов. Давайте посмотрим на это явление подробнее:
1. Медленные окислительные (I тип) — красные
Это волокна сравнительно тонкого диаметра, которые имеют низкий порог активации мотонейрона. А значит именно они выполняют обыденные сокращения — ведь мозгу достаточно послать слабую команду для сокращения таких волокон. Также, красные волокна сокращаются относительно медленно (порядка 100-110 мс).
Кровоснабжаются эти волокна хорошо и имеют высокое содержание миоглобина (используется как депо кислорода). Крупные митохондрии позволяют им работать на протяжении более длительного времени.
Название — окислительные, очень логично, поскольку получение энергии ими осуществляется за счет аэробного дыхания (процесс длительный и требует наличие кислорода). Обычно это подразумевает окисление глюкозы до пирувата в процессе гликолиза, с последующим окислением до углекислого газа в цикле Кребса.
Красные волокна выполняют основную работу когда вы печатаете на клавиатуре, идете на работу или даже бегаете по утрам (только если не очень быстро).
2. Быстрые гликолитические волокна (II тип) — белые
Волокна данного типа в целом более толстые и сильные и куда больше подвержены гипертрофии (увеличению в размере). Для них характерна большая скорость сокращения (порядка 50 мс), но и большая утомляемость.
Название гликолитический происходит от основного способа получения ими энергии (в результате гликолиза). Данный способ позволяет получить АТФ быстро и не требует кислорода, то есть, является анаэробным. Однако, у него низкая эффективность — всего 2 молекулы АТФ из 1 молекулы глюкозы.
Для белых волокон характерен высокий порог активации мотонейрона. Это значит, чтобы задействовать данный тип волокон, мозг должен послать сильную команду на сокращение. Получается, что в обычной жизни, такие волокна слабо задействованы.
В разных мышцах доля белых волокон различается. Так, например, в уже упомянутых икроножных — быстрых волокон довольно мало, поскольку икры чаще всего выполняют монотонную работу и должны быть довольно выносливыми. А вот у разгибателей плеча (трицепса) большинство волокон — белые, ведь сокращаться ему нужно быстро. Будь мы в дикой природе, я бы сказал, что такие волокна в основном отвечают за реализацию стратегии бей, или беги.
Среди быстрых волокон выделяют два подтипа.
IIа тип: быстрые окислительно-гликолитические, или просто быстрые окислительные волокна. По сути это почти те же быстрые волокна, но чуть меньшей толщины. Они более выносливы, чем волокна IIb типа, но утомляются быстрее, чем волокна I типа. При сокращении данный тип волокон развивает среднюю силу, используя в качестве источников энергии как окислительные (используются медленными), так анаэробные механизмы (используются быстрыми волокнами).
IIb тип: быстрые гликолитические волокна — толстые, быстрые, сильные волокна.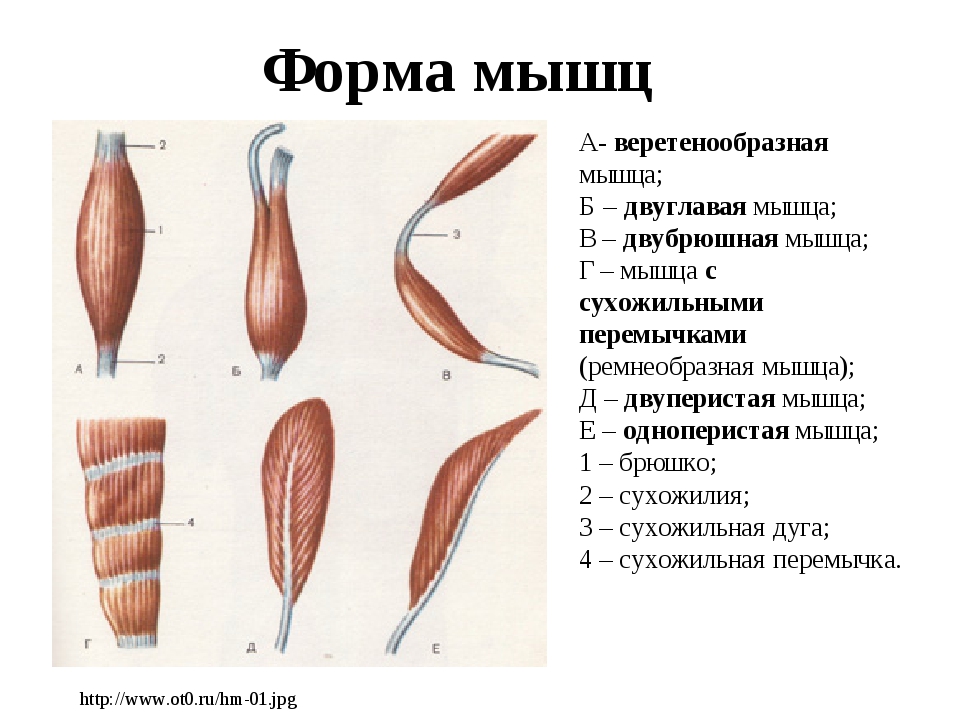 Для них характерна быстрая утомляемость и высокий порог активации мотонейрона. Для получения энергии используют те же механизмы, что и быстрые волокна.
Для них характерна быстрая утомляемость и высокий порог активации мотонейрона. Для получения энергии используют те же механизмы, что и быстрые волокна.
На рисунке сверху показано условное распределение быстрых и медленных волокон, а так же указаны типичные примеры мышц с преобладанием конкретного типа волокон.
Увеличение мышечной массы: гипертрофия или гиперплазия?
Количество волокон в одной и той же мышце у разных людей может существенно отличаться. Изначально считалось, что число мышечных волокон генетически детерминировано и не меняется в течение жизни. Соответственно и мышечный рост обусловлен не увеличением числа мышечных волокон, а увеличением их диаметра (гипертрофия).
Однако в последнее время появляется все больше работ, показывающих возможность увеличения числа волокон (гиперплазия) у животных, например, у птиц. Обычно, причиной гиперплазии у животных служит экстремальное растяжение мышц на протяжении длительного времени (от пары часов, до нескольких суток). Если кто-то подумал, что есть птицы, приверженцы экстремальной йоги — спешу вас разочаровать. Эти экстремальные растяжения являются частью экспериментов и достигаются не самым приятным образом.
Так за счет какого процесса происходит развитие и рост мышц у нас с вами?
Существующие работы по исследованию мышечного роста у человека показывают, что именно увеличение толщины волокон является причиной увеличения объема его мышц. И именно силовые нагрузки приводят к гипертрофии мышечных волокон человека
Почему разные мышцы растут по разному?
Наиболее привычный и понятный для нас способ тренироваться — это обычные силовые тренировки. Под воздействием таких тренировок происходит гипертрофия быстрых и части промежуточных волокон (IIa), в то время, как медленные волокна чаще остаются за бортом.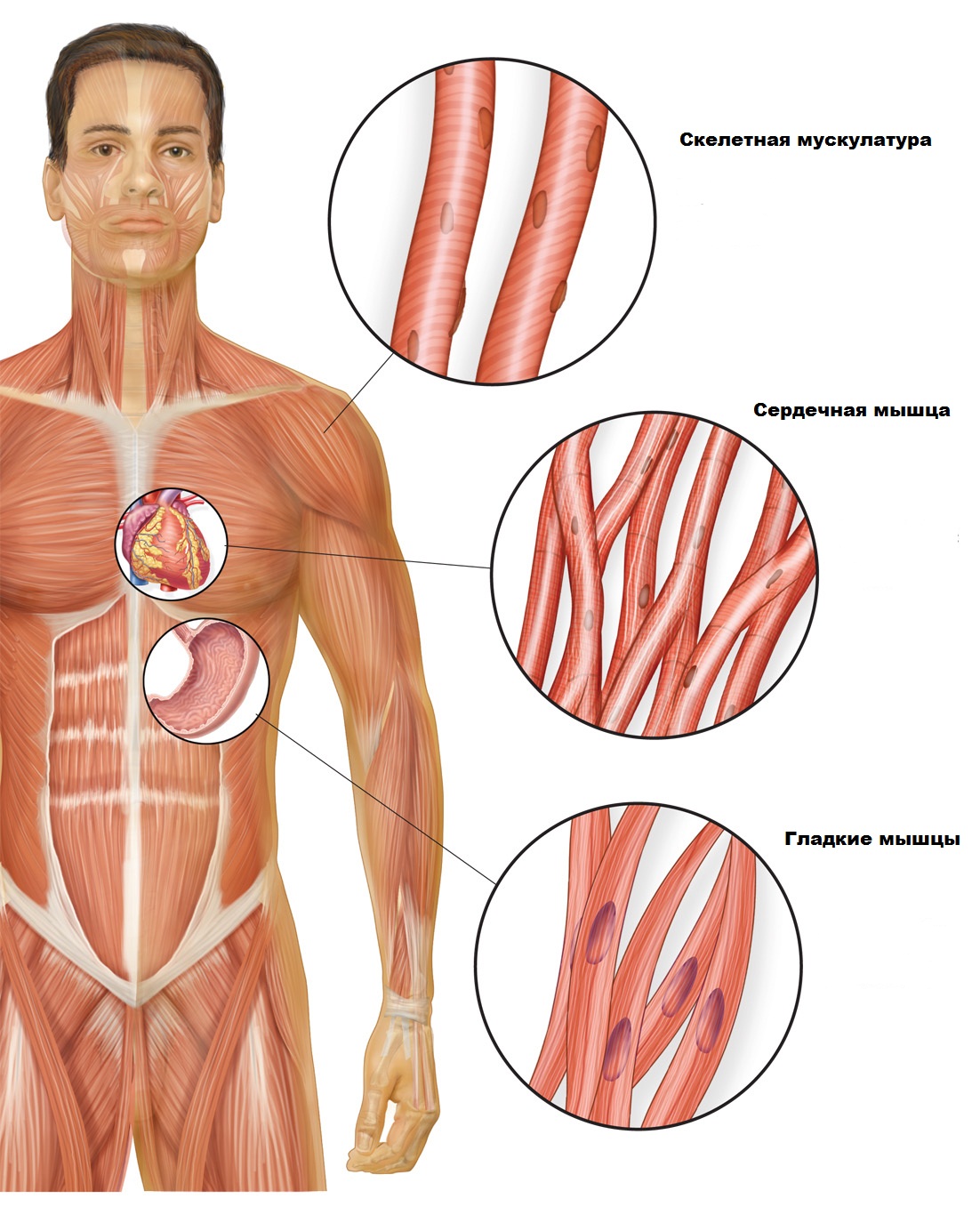
Тогда как гипертрофировать мышцы с преобладанием медленных волокон?
Все просто, нужно выполнять упражнения в многоповторном режиме. Для примера возьмем икры (в них много медленных волокон). Хорошим подходом к тренировке этих мышц будут упражнения, которые можно выполнять неспеша в течение минуты (или более, в зависимости от вашей тренированности). Для примера возьмем подъёмы на носочки. За минуту получится примерно 30-40 повторений — это по сути тренировка на выносливость.
А что тогда насчет обычных силовых тренировок? Ведь в икрах все еще остаются быстрые волокна, которые тоже хочется гипертрофировать.
Хотя многоповторные нагрузки и оказывают на икры наибольший эффект (в отличие от, например, на грудных мышц), для достижения максимального эффекта можно разбавлять их редкими, но «тяжелыми» тренировками с числом повторов от 8 до 20. В таком случае можно использовать утяжелители или просто выполнять позитивную фазу (вставать на носочки) в максимально быстром темпе. Такой подход поможет максимально включить быстрые волокна.
А как обеспечить рост мышц с быстрыми волокнами?
Например, вы хотите гипертрофировать трицепс (помним, что в нем много быстрых волокон). Это значит, что эффективными будут подходы с малым, и средним числом повторов и большой нагрузкой (50-80% от одноповторного максимума). При этом, длительность подхода не должна превышать 25-30 секунд, так как к этому времени уже успевает закончится АТФ и потихоньку подходят к концу запасы креатин фосфата (еще один вид топлива для быстрых волокон). После этого необходим отдых в 60-120 секунд (этого хватает, на ресинтез запасов топлива для быстрых волокон). С другими мышцами, с преобладанием быстрых волокон примерна такая же картина.
В довесок скажу, что с распределением волокон все не так просто. Есть еще ряд факторов (таких как пол, возраст и т.д.), которые могут оказать существенное влияние на соотношение мышечных волокон в мышцах человеческого тела.
Подробнее об этих и других аспектах, связанных с соотношением типов мышечных волокон в теле мы поговорим в следующей статье.
P. S. Вы уже наверное поняли, что эта тема достаточно сложная и применять эти знания не так уж просто. Но мы с друзьями заморочились и недавно запилили фитнесс приложение на основе ИИ, и написали об этом небольшую статью. Оно в самом начале оценивает точку старта человека и на основе его физических особенностей создает индивидуальные тренировки.
Если влезть под капот, то мы увидим, что алгоритм учитывает сколько времени должны длиться подходы, чтобы привести именно к гипертрофии, при этом нагрузка калибруется так, чтобы человек реально мог все выполнить. И да, он не выплёвывает легкие после первой тренировки, и на завтра может ходить + еще куча интересных механизмов на базе спортивной физиологии, о которых мы немного расскажем позже.
Не проходящая ноющая боль в мышцах
Мышечная боль или миалгия представляет собой довольно неприятный симптом, приносящий неудобства и мучения ее обладателю. Она имеет тянущий или спастический характер. Такие боли могут быть вызваны излишними физическими нагрузками или длительным неправильным положением тела и быть неопасными, либо сигнализировать о развитии серьезной болезни. Кунцевский лечебно-реабилитационный центр специализируется на диагностике и лечении заболеваний, вызывающих подобно рода симптомы.
Причины возникновения
Среди основных причин возникновения мышечной боли выделяют:
- тяжелые физические нагрузки;
- травмы;
- нахождение в положении, нарушающем кровообращение;
- плоскостопие или лишний вес;
- хождение на высоких каблуках;
- болезни суставов;
- нарушения опорно-двигательной системы;
- нарушения обменных процессов;
- воспалительные процессы;
- травмы: ушибы, растяжения, повреждения костей и суставов;
- бактерии, вирусы и паразиты, как источник интоксикации;
- хроническая усталость;
- недостаток витаминов В.
Внимание! Точные причины возникновения непроходящих ноющих болей в мышцах помогут выяснить квалифицированные специалисты Кунцевского лечебно-реабилитационного центра, обладающего современным диагностическим оборудованием.
Классификация
Различают несколько видов мышечных болей:
- фибромиалгия — спазмы в волокнистых мышцах, связках и сухожилиях;
- миалгия — боли по всему объему мышц;
- миозит — тупая боль в мышцах после перенапряжения или вследствие воспаления;
- болевые ощущения после спортивных тренировок.
Патологии
Патологические болевые ощущения в мышцах, требующие обязательного посещения специалиста, выражаются в следующем:
- постоянная или хроническая боль;
- дергающий характер недуга;
- болевые ощущения появились после тренировки, но не проходят после отдыха;
- наблюдаются покраснения мягких тканей или отеки;
- наблюдаются сопутствующие симптомы в виде повышенной температуры, одышки, сыпи, задержки мочеиспускания.
Важно! При появлении хотя бы одного из вышеперечисленных состояний рекомендуется обращение к специалисту.
Диагностика
Диагностика причин болевых ощущений в мышцах состоит из следующих основных этапов:
Кунцевский центр предлагает все виды современной диагностики: МРТ, УЗИ, лабораторные исследования, способные выявить причины патологических болей в мышцах и назначить адекватное лечение.
Какой врач лечит
Лечением и диагностикой подобных симптомов занимаются врачи-неврологи, ревматологи или хирурги.
У всех людей иногда возникает боль в мышцах. Если мышцы болят после занятий спортом или после длительного периода активности, в этом нет ничего странного, и беспокоиться не стоит.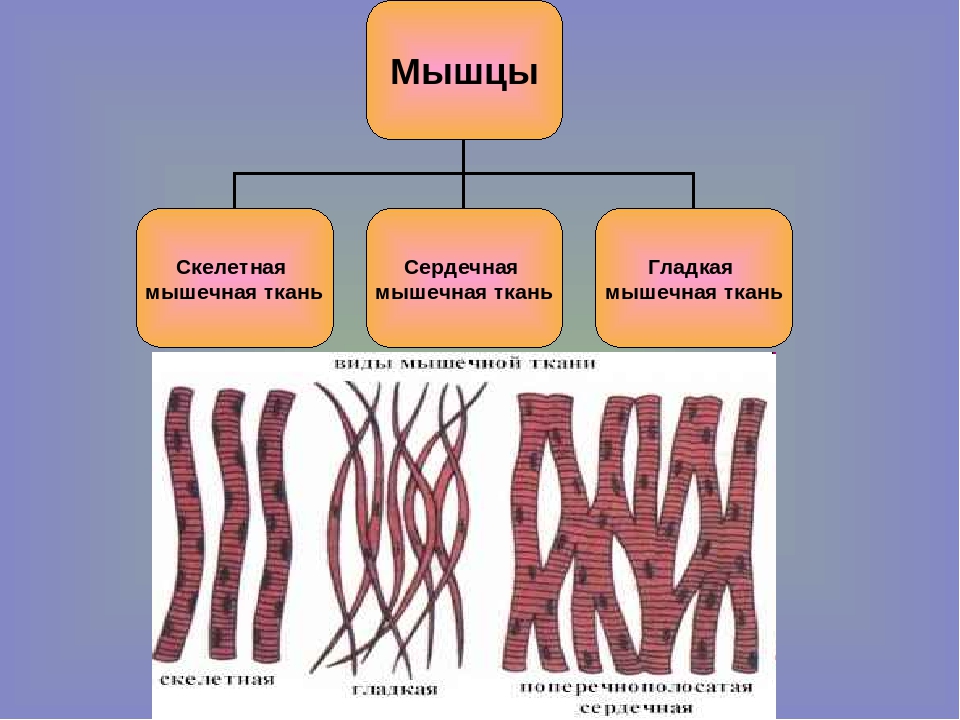
ВАЖНО! Однако, если вы отмечаете длительную ноющую боль в мышцах, которая в течение длительного промежутка времени не проходит, это повод обратиться к специалисту.
Самолечение — не выход из ситуации, потому что причин развития симптома может быть множество, начиная от травмы, заканчивая хроническими воспалительными процессами. Запишитесь на прием к неврологу нашего центра, чтобы разобраться в причинах возникновения симптома, выбрать правильную и эффективную тактику лечения!
Виды лечения
В зависимости от причины возникновения болей в мышцах применяют следующие виды лечения:
- медикаментозное;
- массаж;
- ЛФК;
- оперативное лечение в тяжелых случаях.
Реабилитация
Реабилитационный период характеризуется прохождением процедур массажа, выполнения лечебных упражнений, корректировки питания при наличии лишнего веса.
Рекомендации по образу жизни
В качестве профилактических мер миалгий различного происхождения важно избегать тяжелых физических нагрузок и стрессов, не переохлаждаться, долечивать до конца инфекционные заболевания и вести здоровый образ жизни.
Ноющая боль в мышцах достаточно частое явления, порой оно носит безобидный характер и в скором времени проходит, но иногда является следствием серьезных заболеваний, требующих контроля и лечения. Высококвалифицированные специалисты Кунцевского лечебно-реабилитационного центра готовы оказать своевременную и качественную помощь в решении данного вопроса.
Искусственные мышцы обогнали по мощности настоящие
Shuguang Li et al. / PNAS, 2017
Разработана технология создания недорогих искусственных мышц на основе жесткого каркаса, заключенного в мягкую камеру.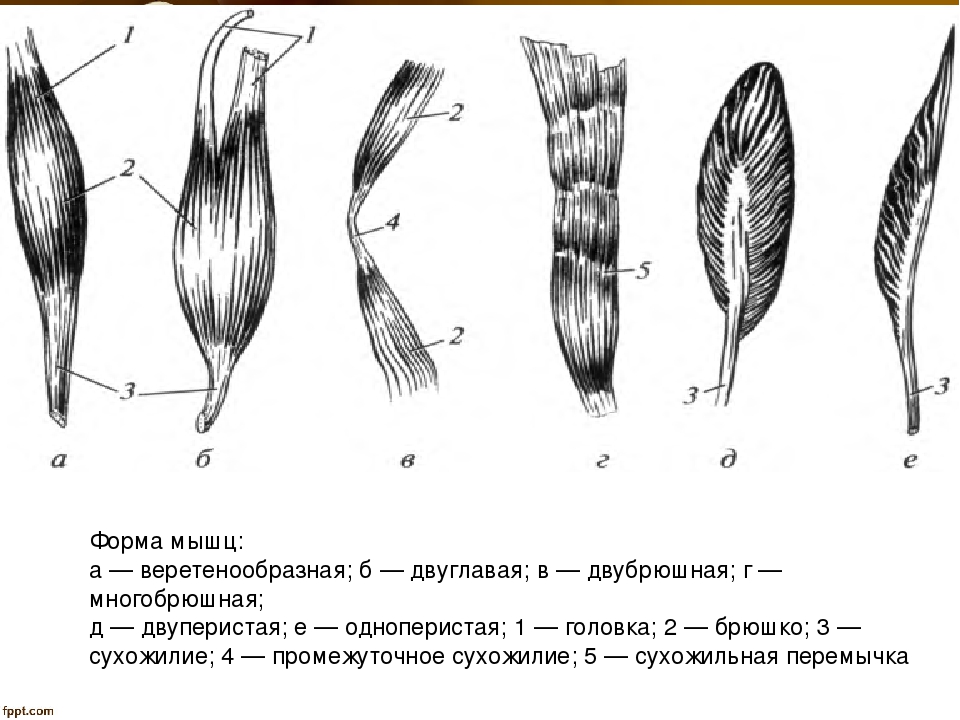 Мышцы сокращаются за счет уменьшения в них давления, причем их можно создавать, используя разные материалы. Статья опубликована в журнале Proceedings of the National Academy of Sciences.
Мышцы сокращаются за счет уменьшения в них давления, причем их можно создавать, используя разные материалы. Статья опубликована в журнале Proceedings of the National Academy of Sciences.
Инженеры, разрабатывающие роботов, нередко используют в своих изобретениях конструкции, напоминающие по функциям живых существ. Несмотря на это, для движения роботы все равно чаще всего используют электромоторы или двигатели внутреннего сгорания, соединенные со сложными механическими передачами. Некоторые исследователи придерживаются другого подхода и разрабатывают источники движения, более близкие по своему устройству к мышцам. Уже существует немало прототипов искусственных мышц, которые могут сокращаться подобно настоящим мышцам, но почти все они требуют дорогих материалов и технологических процессов, при этом эффективность многих из них все еще низка.
Исследователи под руководством Роберта Вуда (Robert Wood) из Гарвардского университета разработали простую и недорогую технологию создания эффективных искусственных мышц, которые можно создавать из большого количества разных материалов. Принципиальная схема создания таких актуаторов довольно проста. В качестве основы используется каркас заданной формы, который может складываться и раскладываться. Затем вокруг этого каркаса склеиваются или сплавляются два фрагмента пленки из полимера или другого воздухонепроницаемого и мягкого материала. Таким образом формируется мягкая камера с жестким каркасом внутри, которая подключается к источнику разницы давления.
Принцип действия искусственных мышц
Shuguang Li et al. / PNAS, 2017
Управление актуатором происходит за счет уменьшения или увеличения давления жидкости или газа внутри камеры. В результате актуатор начинает менять форму: складываться или наоборот увеличиваться в размерах, а в случае с каркасом сложной формы, совершать другие движения — например, изгибаться в определенную сторону.
Пример захватывающего устройства
Shuguang Li et al. / PNAS, 2017
С помощью такой технологии исследователи создали несколько прототипов актуаторов, и измерили их эффективность. Один из этих прототипов, представляющий собой десятисантиметровый линейный актуатор весом менее трех грамм, смог поднять груз массой более трех килограмм. Исследователи подсчитали, что пиковая мощность таких актуаторов составляет около двух киловатт на килограмм массы, что делает их мощнее настоящих скелетных мышц млекопитающих.Ранее ученые представляли множество прототипов искусственных мышц, работающих на основе разных принципов. Некоторые также работают за счет давления, например, робогусеница, основную часть которой занимает полимерная пена, покрытая силиконом, а также мягкие вакуумные актуаторы из множества полых ячеек. Другие используют для своей работы нагревание: таким образом работают мышцы на основе нейлоновой лески и недавно представленный материал, наполненный пузырьками с этанолом, который при нагревании превращается в газ и расширяется. Помимо этого недавно был представлен актуатор из множества слоев двумерного материала, который расширяется при внедрении в него сторонних ионов. Кстати, не всегда искусственные мышцы сделаны полностью из искусственных материалов. Тайваньские ученые сделали мышцы из тонкой пленки из кожицы лука, которая сокращается под действием электричества.
Григорий Копиев
6 лучших женских упражнений для мышц тазового дна
6 лучших женских упражнений для мышц тазового дна!
Эта программа позволит вам укрепить мышцы тазового дна, раскрыть таз, улучшить кровообращение и даже усилить ощущения от секса!Наш эксперт: Александра Панферова, мастер-тренер направления групповых программ федеральной сети фитнес-клубов X-Fit.

«Кошка»
«Плечевой пост»
Исходное положение лежа на спине, ноги согнуты в коленях, голени перпендикулярны полу, стопы на ширине таза. Акцент на работу ягодиц и мышц тазового дна, а так же артикуляцию(вытяжение) позвоночника.
На выдохе, подкручивая таз, напрягайте ягодицы, втягивайте глубокие мышцы живота, создавая «эффект вакума» и поднимайте таз, оставляя опору на лопатки. Шея расслаблена. На вдохе возвращайте таз на пол, вытягивая весь позвоночник по полу.
«Перекат»
Акцент на работу мышц тазового дна и артикуляцию позвоночника.
Сгруппировавшись, втяните мышцы живота и, сохраняя эту силу, выполните перекат, касаясь лопатками пола, а потом возвращайтесь обратно. Старайтесь выполнять упражнение плавно.
Раскрываем мышцы внутренней поверхности бедра
Исходное положение сидя на полу, прямые ноги максимально широко открыты в стороны. Ноги активные, создавайте вытяжение через пятки. Начинайте наклон вперед с прямой спиной , создавая спокойное вытяжение мышц внутренней линии бедра. Удерживайте это положение 30-40 секунд.
Наклоны к одной прямой ноге
Исходное положение сидя, одна нога вытянута в сторону.
На вдохе, сохраняя ягодицы прижатыми к полу, выполните наклон в сторону, к прямой ноге.
Перевернутое положение
Сгруппируйтесь, поднимите таз и вытяните ноги вверх до вертикального положения.
Напрягите ягодицы и мышцы живота. Ощущайте ноги активными, тянитесь пальцами ног вверх.
 Удерживайте положение от 30 сек.
Удерживайте положение от 30 сек.Источник:http://www.goodhouse.ru/
|
Многие полагают, что физическая активность зимой должна ограничиваться «утренней» домашней зарядкой и занятиями в теплых стенах фитнес-центра. Это заблуждение. Несмотря на то, что за окном стало холоднее, тренировки на улице все равно приносят пользу: они помогают укрепить иммунитет, что немаловажно в период эпидподъема заболеваемости ОРВИ, побороть бессонницу, снимают стресс и улучшают настроение. Кроме того, на свежем морозном воздухе быстрее «тают» жировые отложения. Зимой следует как можно чаще бывать на улице и заниматься спортом на свежем воздухе, ведь это намного эффективнее, чем тренировки в помещении. Зимние виды спорта не только дарят массу положительных эмоций, но и укрепляют здоровье. Лыжи. Лыжные прогулки подразумевают движение без остановок в течение длительного периода времени, тем самым они дают аэробную нагрузку организму. Во время таких тренировок повышается выносливость, улучшается работа сердечнососудистой системы, органов дыхания, опорно-двигательного аппарата, повышается иммунитет и в целом происходит общее оздоровление организма. Кроме того, ходьба на лыжах позволяет сжечь до 500–600 калорий в час в зависимости от прилагаемых усилий, а также укрепить мышцы спины, бедер, живота, ягодиц, ног и рук. По мнению врачей, зимой достаточно устраивать часовые прогулки на 3–5 км 2–3 раза в неделю, чтобы поддерживать себя в прекрасной форме. Сноубординг с каждым годом становится все более популярным благодаря положительному воздействию на организм человека. Во время занятий на сноуборде задействованы все группы мышц, особенно ног, ягодиц, живота и бедер, а нагрузка позволяет за час избавиться от 400 и более калорий. Кроме того, сноубординг укрепляет вестибулярный аппарат, нормализует работу сердечнососудистой и дыхательной системы, а также насыщает кровь кислородом. Коньки. Катание на коньках прекрасно тренирует гибкость, быстроту, ловкость и умение держать равновесие. Конькобежный спорт помогает развивать мышцы в основном нижней части тела, а фигурное катание и выполняющие различных трюков и поддержки — иметь более сильный «верх». Кроме того, регулярные занятия на льду укрепляют здоровье, повышают тонус организма и его сопротивляемость различным инфекциям, а также улучшают настроение благодаря повышенной выработке эндорфинов – «гормонов счастья». За час катания на коньках можно сжечь до 400–500 калорий и укрепить мышцы ног, бедер и ягодиц. Для поднятия тонуса организма достаточно кататься на коньках 2–3 раза в неделю по часу. Хоккей на льду. Любительские игры безопасны и даже полезны для организма. Во время тренировок задействована как нижняя, так и верхняя часть тела, благодаря чему отлично прорабатываются мышцы ног, рук, живота и спины. В зависимости от интенсивности игры за час сжигается до 500–600 калорий. Хоккей также благоприятно влияет на вестибулярный и опорно-двигательный аппарат, повышает иммунитет. Этот командный вид спорта тренирует и характер, вырабатывая такие качества, как выносливость, упорство, ловкость и реакция. Чтобы получить максимальную отдачу от игры в хоккей, следует играть 1–2 раза в неделю по часу, делая небольшие перерывы в игре. Санки — это не только детская забава, но и прекрасный способ поддерживать тело в тонусе. Во время спусков и подъемов организм получает умеренную кардионагрузку, благодаря чему происходит укрепление сердечной мышцы, нормализуется кровяное давление, а также эффективно сжигаются калории и уходят лишние килограммы. Кроме того, катание на санках – это настоящий «антидепрессант», стимулирующий выработку гормонов, отвечающих за хорошее настроение. Занимайтесь спортом, укрепляйте свое здоровье и заряжайтесь позитивом! |
 Еще одной его особенностью является то, что он дарит массу «острых» ощущений, незаменимых для психического здоровья. Для укрепления здоровья следует вставать на сноуборд не реже 1–2 раз в неделю и кататься не менее часа.
Еще одной его особенностью является то, что он дарит массу «острых» ощущений, незаменимых для психического здоровья. Для укрепления здоровья следует вставать на сноуборд не реже 1–2 раз в неделю и кататься не менее часа.
Растяжения связок, сухожилий и мышц | Макаров С.А., Сергиенко С.А.
Повреждение сухожильно–связочного аппарата является одной из наиболее актуальных и часто встречающихся причин, ограничивающих физическую деятельность людей, ведущих активный образ жизни. Основным этиологическим фактором служит острое травматическое повреждение, называемое растяжением, либо избыточная циклическая нагрузка в течение длительного времени – перенапряжение. Растяжения связок, сухожилий и мышц – это острые и травматические по причине повреждения. Существуют три степени тяжести растяжения связок:
I степень – небольшая боль из–за разрыва нескольких волокон связки.
II степень – умеренная боль, отек и нетрудоспособность.
III степень – сильная боль из–за разрыва связки и последующая нестабильность сустава.
Растяжение мышцы в свою очередь является травматическим повреждением самих мышечных волокон или соединения мышцы и сухожилия и классифицируется также по трем степеням тяжести:
I – умеренная.
II – средняя степень повреждения, связана со слабостью пораженной мышцы, ее болезненным сокращением.
III – полный разрыв соединения мышцы и сухожилия, проявляющийся сильной болью и невозможностью сокращения поврежденной мышцы.
Перенапряжение – это не острое, повторяющееся поражение мягкотканных структур вследствие постоянно случающихся микротравм, которое ведет к изменению локальной микроциркуляции и, как следствие, к дегенеративным процессам в мягких тканях, где при микроскопии отмечается нарушение структуры местных тканей с их лизисом, лейкоцитарной инфильтрацией и экстравамацией крови.
Повторяющиеся движения за время длительной работы приводят к возникновению повреждения от «переиспользования» у людей некоторых профессий. Приблизительно 10–20% музыкантов, машинисток, кассиров и рабочих конвейера жалуются на рецидивы синдрома растяжения, среди спортсменов этот процент колеблется от 30 до 50.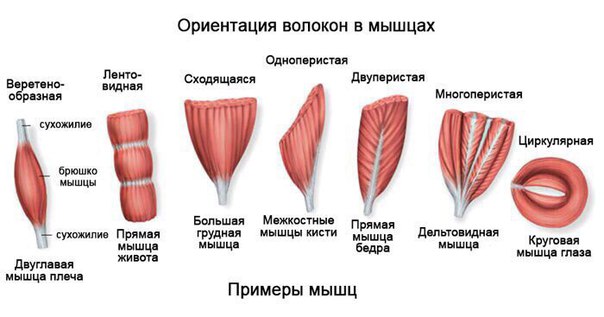
Повреждения из–за перенапряжения (перетренировки) подразделяются на четыре степени:
I степень – боль только после физической активности.
II степень – боль во время и после физической нагрузки, не влияющая на результат работы.
III степень – боль во время и после физической нагрузки, влияющая на результат работы.
IV степень – постоянная боль, нарушающая ежедневную физическую активность.
Следует также отметить, что поражение сухожильного аппарата может протекать в виде «тендинита», «тендиноза» и «теносиновита».
Тендинит возникает из–за травмы сухожилия и связанного с нею разрушения сосудов и острого, подострого или хронического воспаления.
Тендиноз – это невоспалительной природы атрофия и дегенерация волокон внутри сухожилия, часто связанная с хроническим тендинитом, которая может привести к частичному или полному разрыву сухожилия.
Теносиновит – это воспаление паратендона, который является наружным влагалищем некоторых сухожилий и выстлан синовиальной мембраной (например, поражение сухожилия разгибателей большого пальца кисти при теносиновите де Курвена).
В данном обзоре имеет смысл остановиться именно на нагрузочных повреждениях, вызванных часто повторяющимися однообразными движениями, то есть на перенапряжении, а также учесть легкие степени растяжения связок, сухожилий и мышц, поскольку травма, сопровождающаяся разрывом этих структур, в большинстве случаев требует хирургического лечения (повреждение передней крестообразной связки и коллатеральных связок коленного сустава, разрыв ахиллова сухожилия).
Наиболее часто встречающиеся виды повреждения из–за перенапряжения (перетренировки):
связок – «локоть бейсболиста», «колено пловца», синдром трения илиотибиального тракта, «колено прыгуна», подошвенный фасциит;
сухожилий – тендинит ахиллова сухожилия, верхненадколенниковый тендинит, тендинит двуглавой мышцы плеча, тендинит задней большеберцовой мышцы, латеральный эпикондилит («локоть теннисиста»), тендинит надостной мышцы (вращающей манжеты).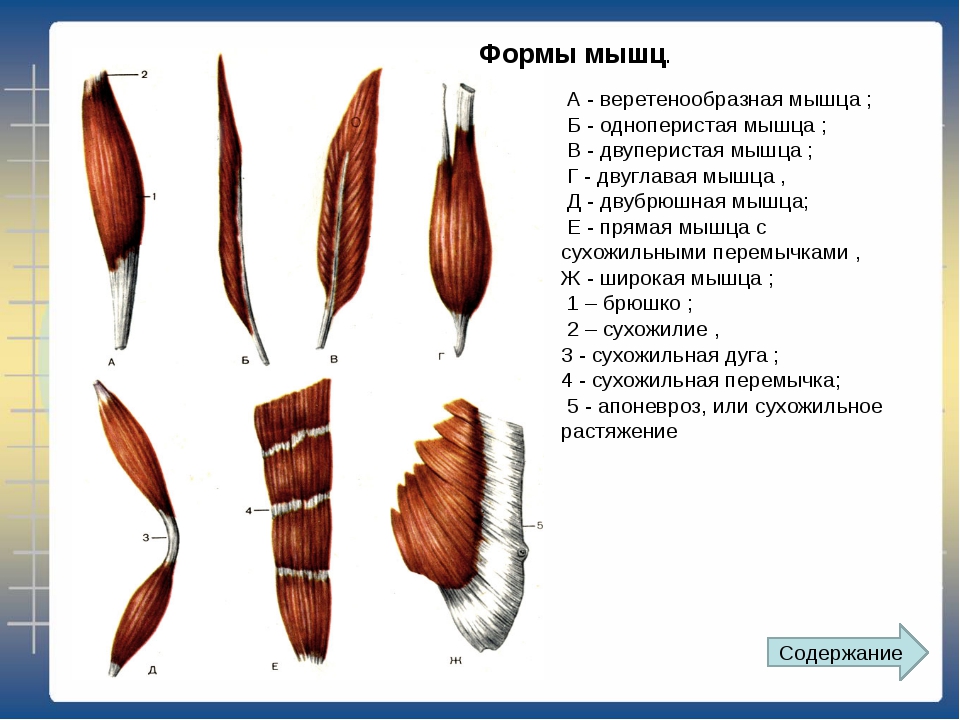
«Локоть теннисиста» – это латеральный эпикондилит. Этот синдром возникает вследствие перетренировки и проявляется болью по латеральной поверхности локтевого сустава. Пациенты обычно связывают его возникновение с игрой в теннис. На сегодняшний день латеральный эпикондилит считается воспалительным заболеванием и/или микроразрывом в месте прикрепления короткого лучевого разгибателя кисти (m. extensor carpi brevis). Провоцирующее движение – форсированное разгибание среднего пальца кисти против сопротивления – вызывает боль, так как мышца прикрепляется к основанию пясти среднего пальца.
«Локоть игрока в гольф» – медиальный эпикондилит, возникает при повреждении от перетренировки сухожилий мышц пронаторов и сгибателей предплечья в месте их прикрепления к медиальному надмыщелку. Эта область подвергается вальгусно направленному воздействию на вершине замаха, боль отмечается над медиальным надмыщелком и усиливается при сгибании и пронировании предплечья на фоне сопротивления.
«Локоть бейсболиста» (воспаление медиального апофиза) – это заболевание возникает из–за вальгусно направленной силы при частом движении руки по кривой броска мяча. У пострадавшего отмечают микроразрывы сухожилий мышц пронаторов и сгибателей, а в тяжелых случаях – отрыв и раздробление медиального апофиза.
Синдром трения илиотибиального тракта (СТИТ) – это боль по латеральной поверхности коленного сустава вследствие раздражения и воспаления дистальной части илиотибиального тракта, когда он проходит над латеральном мыщелком бедренной кости. Боль усиливается при пальпации дистальной части тракта в момент разгибания ноги в коленном суставе. СТИТ возникает при чрезмерно интенсивном беге, беге по пересеченной местности.
«Колено пловца» – состояние, возникающее в коленном суставе при вальгусно направленной на колено силе из–за резких движений ногой во время плавания брассом. Обычно это наблюдается при растяжении медиальной коллатеральной связки коленного сустава, что и вызывает боли.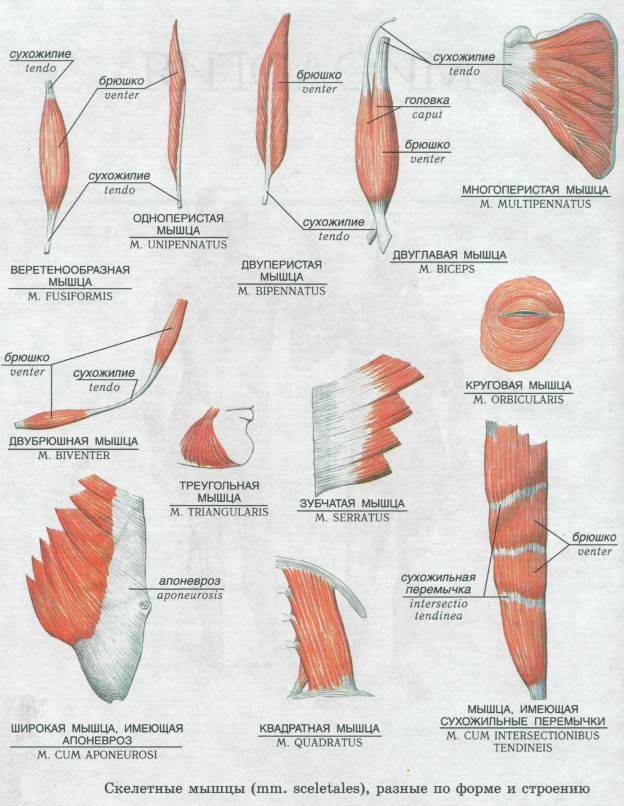
«Колено прыгуна» – так называемый тендинит надколенника. Часто встречается у прыгунов в высоту, баскетболистов и волейболистов. Характеризуется болями в нижнем полюсе надколенника, в месте прикрепления связки надколенника. Развивается из–за постоянного повреждения данной области, когда не происходит восстановления и заживления травмы.
Тендинит двуглавой мышцы плеча проявляется болью в передней части плечевого сустава, которая усиливается при активных движениях в плечевом суставе и менее выражена или отсутствует при пассивных движениях, а также сопровождается локальной болезненностью при пальпации области над длинной головкой сухожилия двуглавой мышцы. В случае сопутствующего миозита тендинит двуглавой мышцы сопровождается выраженной болезненностью мышцы.
Бурсит надколенника сопровождается болью, отеком и местным повышением температуры в сумке надколенника, которая расположена поверхностнее надколенника. Вызывается бурсит повторной травмой или нагрузкой, как при стоянии на коленях.
Воспаление ахиллова сухожилия проявляется болью в пятке, иногда болью по задней поверхности ноги. Дорсальное и подошвенное сгибание стопы усиливает боль, область наибольшей болезненности находится на 2–3 см проксимальнее места соединения сухожилия с пяточной костью. Сухожилие может быть отечно и утолщено, причиной чаще является спондилоартропатия с поражением периферических суставов (болезнь Рейтера, анкилозирующий спондилоартрит), а также травма.
Подошвенный фасциит или пяточная шпора анатомически возникает из места прикрепления короткого сгибателя пальцев, локализующегося по переднемедиальному краю бугристости пяточной кости несколько глубже места прикрепления подошвенной фасции. Перенапряжение одной из этих структур, как считается, приводит к возникновению реактивной воспалительной продукции костной ткани или формированию шпоры вторично из–за тракции этих структур. Однако остается неясным, какой из механизмов ответственен за это. В любом случае шпора вторична по отношению к перенапряжению.
В любом случае шпора вторична по отношению к перенапряжению.
Также следует упомянуть о такой патологии, как «расколотая голень» – это повреждение от перетренировки, вызываемое хронической тракцией надкостницы большеберцовой кости. При этом поражаются либо берцовые мышцы, либо m. soleus, что характеризуется постепенно начинающейся болью по переднемедиальной или заднемедиальной поверхности голени. Боль возникает у спортсменов на старте забега, во время бега стихает и вновь усиливается после окончания забега. При пальпации определяется болезненность по заднемедиальному краю большеберцовой кости, обычно на границе средней и нижней трети. Боль усиливается при дорсальном сгибании стопы против сопротивления.
Лечение
Терапевтическое лечение повреждения связок, сухожилий и мышц включает в себя первичную и вторичную терапию.
Первичная терапия:
- Защита от нагрузки
- Отдых
- Лед
- Давящая повязка
- Возвышенное положение
- Поддержка повязкой
Вторичная терапия
- Физиотерапия
- Реабилитация
- Инъекции
- Лечебная физкультура
- Обследование и повторное обследование
- Салицилаты.
Таким образом, основой лечения является раннее обезболивание и противовоспалительная терапия травмы мягких тканей, особенно в случаях сопутствующего миозита. Длительно текущее воспаление ухудшает процесс заживления мягких тканей, что приводит к их детренированности и функциональной недостаточности. При активном воспалительном процессе важен относительный отдых для области поражения. Лед эффективен как противовоспалительный агент только в первые часы после травмы, затем предпочтительней использовать тепло. Иммобилизация шиной или повязкой может применяться для усиления защиты травмированной конечности или части ее от нагрузки.
Инъекции кортикостероидов при хронических процессах не дают полного излечения, к тому же они увеличивают скорость деградации коллагена, снижают синтез нового коллагена, уменьшают прочность сухожилия при растяжении, что приводит к его разрыву, если инъекции выполнять неправильно или очень часто. В связи с этим их применение оправдано только в остром периоде и не чаще 1 раза в 2–3 недели.
Необходимо также использовать нестероидные противовоспалительные препараты (НПВП) per os, причем в данных ситуациях препараты различных групп примерно равноэффективны. Длительное их применение рекомендуется при хронических состояниях перенапряжения, при острой травме они действенны в течение 72 часов.
Весьма эффективным методом лечения травматических повреждений мягких тканей является локальная терапия с использованием мазей и гелей, содержащих НПВП. Применение местных средств особенно эффективно у лиц пожилого и старческого возраста с отягощенным язвенным анамнезом. Эти средства способны при необходимости заменить системно применяемые НПВП. При растяжении связок и мышц, сопутствующих миозитах, сопровождающихся раздражением нервных корешков и периферических нервов, используются комплексные препараты, содержащие НПВП и растительные компоненты. В России одним из наиболее известных и хорошо зарекомендовавших себя препаратов этого ряда является Эфкамон, обладающий отвлекающим, анальгезирующим, согревающим, рассасывающим и противовоспалительным действием. Входящий в состав препарата метилсалицилат оказывает противовоспалительное и анальгезирующее действие за счет подавления синтеза простагландинов в очаге воспаления. Настойка перца стручкового содержит капсаицин, обладающий сильным раздражающим действием на рецепторы кожи. Уникальное сочетание природных компонентов препарата, сочетанное действие входящих в его состав эфирных масел, обеспечивающих выраженное анальгетическое действие за счет взаимодействия с чувствительными рецепторами кожи, делает препарат особо эффективным при травматическом повреждении мягких тканей (ушибах), миозитах, ганглионитах, а также артритах и шейно-крестцовом радикулите, что особенно важно у лиц пожилого и старческого возраста. Следует отметить тот факт, что препарат не обладает иммуносупрессивным эффектом, свойственным практически всем синтетическим препаратам.
Мазь втирают в кожу пораженной области в количестве 3–4 г. 2–3 р/сут и покрывают сухой согревающей повязкой. Длительность лечения зависит от характера и тяжести заболевания.
Таким образом, использование современных препаратов, особенно местного действия (Эфкамон), является эффективным методом купирования боли и сопутствующего воспаления при травматическом повреждении сухожильно–связочного аппарата, что способствует скорейшему восстановлению физической активности и возвращению к прежнему качеству жизни пациентов.
Литература:
1. Михеев С.М. Применение местных средств в ревматологии. Русский медицинский журнал 2000: т.8 №7. – с. 300–302.
2. Насонова В.А., Фоломеева О.М., Амирджанова В.Н. Ревматические болезни в свете международной статистической классификации болезней и проблем, связанных со здоровьем (десятый пересмотр). Тер. арх. 1998: №5. – с.5–8.
3. Руководство по ревматологии. По ред. В.А. Насоновой, Н.В. Бунчука. – М. – 1997.
4. Силин Л.Л., Бровкин С.В. Применение гелей в комплексном лечении закрытых повреждений мягких тканей. Медицинская помощь 2001: №2.
5. Чичасова Н.В. Локальная терапия хронических заболеваний опорно–двигательного аппарата. Русский медицинский журнал 2001: т.9 №7–8. – с. 286–288.
6. Data from American Association of Retired Persons, the Administration on Aging, and the U.S. Bureau of the Sensus. A profile of older Americans, 1994.
.
Физиотерапия – основные физиотерапевтические процедуры
Автор
Пилат Людмила НиколаевнаВедущий врач
Физиотерапевт
Физиотерапевтические процедуры широко используются в качестве вспомогательного средства при лечении заболеваний различных систем и органов. Они используются в послеоперационный период и в период реабилитации пациента после перенесенного заболевания или травмы. Благодаря физиотерапии эффективность лечения возрастает, процесс выздоровления протекает более быстро.
Физиотерапевты «Семейного доктора» располагают современными аппаратными средствами, позволяющими проводить необходимые и наиболее востребованные физиотерапевтические процедуры, среди которых:
-
TENS-терапия – транскутанная (то есть осуществляемая через кожу) электрическая нервная стимуляция – используется для восстановления сократительной способности пострадавших нервов и мышц, устранения болей неврологического или посттравматического происхождения.
-
Магнитотерапия – метод физиотерапии, основанный на действии магнитного поля. Стимулирует кровоснабжение тканей, способствует уменьшению отеков, воспаления, болевых ощущений. Используется при лечении последствий травм, неврологических и кардиологических заболеваний, заболеваний опорно-двигательного аппарата. Позволяет повысить уровень физических нагрузок.
-
Миостимуляция – воздействие на мышцы слабым импульсным током, вызывающее их сокращение. Достигается укрепление мышц при отсутствии активных физических нагрузок. Восстанавливается тонус тканей, улучшается обмен веществ, снимается мышечное перенапряжение.
-
Импульсная коротковолновая терапия — помогает в период постравматических состояний (при растяжениях, вывихах, ушибах, переломах, гематомах, ссадинах и т.п.), способствует заживлению ран, часто используется после операций на челюсти и конечностях и при некоторых типах воспалений. Данная терапия также показана пациентам с нарушениями периферического кровообращения.
-
СМТ-терапия (амплипульстерапия) – разновидность электротерапии, при котором воздействие на организм осуществляется с помощью амплитудных пульсаций переменных среднечастотных токов (частотой от 2 до 5 кГц). Улучшает лимфо- и кровообращение в пораженной области, способствует снижению болевых ощущений.
-
Лазеротерапия — воздействие с помощью направленного светового потока (лазера). В результате в организме активизируются важнейшие биохимические процессы, обновляются ткани, восстанавливается их жизнедеятельность, включаются механизмы саморегуляции.
-
Ультразвуковая терапия.
-
Фонофорез – сочетание ультразвукового воздействия с действием лекарственных или косметических средств. Ультразвук повышает эффективность местного применения лекарственных препаратов.
-
Электрофорез — введение лекарственных препаратов с помощью постоянного электрического тока.
-
УВЧ-терапия – физиотерапия с помощью ультравысокочастотного электромагнитного поля. Используется для лечения воспалений, травм, радикулита, невралгии.
-
УФ-терапия – воздействие на организм ультрафиолетовым излучением. Повышает сопротивляемость организма инфекциям, активизирует деятельность эндокринной системы и обмен веществ. Используется для восполнения дефицита витамина D, лечения дерматологических заболеваний.
Не занимайтесь самолечением. Обратитесь к нашим специалистам, которые правильно поставят диагноз и назначат лечение.
Оцените, насколько был полезен материал
Спасибо за оценку
видов мидий | Sciencing
Обновлено 22 ноября 2019 г.
Автор: Raine Chasing
В мире существует множество различных видов мидий. Мидия очень похожа на моллюска; он живет в раковине и процветает в соленых или пресных реках, ручьях, приливных водах и озерах. Различные виды мидий также являются любимым блюдом многих любителей моллюсков, а также ненавистными вредителями в озерах и реках.
Зебровая мидия (Dreissena ploymorpha)
Зебровая мидия — пресноводный вид мидий, обитающий в озерах и реках Америки.Они родом из Польши и Советского Союза и впервые появились в Америке на озере Сен-Клер в 1988 году. У них есть полосатый узор на раковине, дающий им название зебра. Они маленькие, но некоторые могут вырасти до 2 дюймов и жить от четырех до пяти лет.
Подробнее о характеристиках мидий.
Зебра является вредителем озер из-за огромного потребления фитопланктона и зоопланктона. Поступая таким образом, они изнуряют голодом других местных рыбных животных, пришедших из этого района.Они производят от 300 000 до 1 миллиона яиц в год, но лишь 2 процента доживают до взрослого возраста.
Молодые мидии свободно плавают по рекам и озерам, переходя по водным течениям из одного места в другое. Более старые мидии неподвижны, прикрепляясь к камням, лодкам, трубам, черепахам или другим мидиям.
Голубые мидии (Mytilus edulis)
Голубые мидии водятся по всему миру в умеренных и полярных водах. Они прикрепляются к сваям и камням вдоль пляжа в приливных зонах.Это корпуса с жесткими шарнирами, которые различаются по цвету: синему, пурпурному и коричневому. Внутренняя часть ракушки жемчужно-белая с синей или фиолетовой каймой по краям.
Подробнее о характеристиках ракушек.
Они вырастают до 10 сантиметров в длину, а некоторые могут вырасти до 20 сантиметров, однако это бывает редко. Голубые мидии также известны под другими названиями, такими как заливные мидии, выращенные на фермах мидии и мидии Острова Принца Эдуарда (PEI).
Мидии кроличьи лапки (четырехугольные цилиндрические)
Мидии кроличьи лапки — это пресноводные моллюски, получившие свое название от формы раковины; форма кроличьей лапки.Раковины у них шарнирные, толстые, прямоугольной формы, удлиненной формы с выступами и выступами по внешней стороне. Внутренняя часть раковины белого цвета, а внешняя — желтовато-коричневого или оливкового цвета, которая может достигать 4 дюймов в длину. Мидия кроличья лапка является вымирающим видом и встречается в чистых текущих ручьях, таких как река Вердигрис.
Мидии Snuffbox (Epioblasma triquetra)
Мидии Snuffbox — это мидии среднего размера, вырастающие только до 2 дюймов.У них есть раковина треугольной формы, желтого цвета и очень толстая. Их раковины имеют сплошные и рваные темно-зеленые полосы по ширине, а один конец откидной.
Мидии Табакерки — исчезающий вид и охраняются законом. Они обитают в быстро движущихся реках с булыжником, песком или гравием, так что они могут зарыться глубоко в отложениях русла.
Конская мидия (Modiolus modiolus)
Конская мидия может вырасти до 20 сантиметров, что намного больше, чем другие виды мидий.Они живут на глубине от 10 до 25 метров в воде; некоторые были обнаружены на глубине до 280 метров под водой. Они прикрепляются к твердым поверхностям, таким как камни, черепахи и другие мидии.
мышцы | Системы, типы, ткани и факты
Мышца , сократительная ткань животных, функция которой заключается в движении.
поперечнополосатая мышца; двуглавая мышца человекаСтроение поперечно-полосатой или скелетной мышцы. Поперечно-полосатая мышечная ткань, такая как ткань двуглавой мышцы человека, состоит из длинных тонких волокон, каждое из которых, по сути, представляет собой пучок более тонких миофибрилл.Внутри каждой миофибриллы находятся филаменты белков миозина и актина; эти нити скользят друг мимо друга по мере того, как мышца сокращается и расширяется. На каждой миофибрилле можно увидеть регулярно возникающие темные полосы, называемые Z-линиями, где перекрываются актиновые и миозиновые филаменты. Область между двумя линиями Z называется саркомером; саркомеры можно рассматривать как первичную структурную и функциональную единицу мышечной ткани.
Encyclopædia Britannica, Inc.Британская викторина
Человеческое тело: факт или вымысел?
Могут ли люди выбрать правшу или левшу? От нервов и генов до мышц и органов — посмотрите, насколько вы владеете обеими руками, выбирая между правильным — и неправильным — в этой викторине.
Движение, сложное взаимодействие мышечных и нервных волокон — это средство, с помощью которого организм взаимодействует с окружающей средой. Иннервация мышечных клеток или волокон позволяет животному вести нормальную жизнедеятельность. Организм должен двигаться, чтобы найти пищу, или, если он ведет малоподвижный образ жизни, должен иметь средства, чтобы приносить пищу самому себе. Животное должно уметь перемещать питательные вещества и жидкости по своему телу, а также реагировать на внешние или внутренние раздражители.Мышечные клетки подпитывают свои действия, преобразовывая химическую энергию в форме аденозинтрифосфата (АТФ), которая получается в результате метаболизма пищи, в механическую энергию.
Мышца — это сократительная ткань, сгруппированная в скоординированные системы для большей эффективности. У людей мышечные системы классифицируются по внешнему виду и расположению клеток. Три типа мышц — поперечно-полосатая (или скелетная), сердечная и гладкая (или гладкая). Поперечно-полосатая мышца почти исключительно прикреплена к скелету и составляет основную часть мышечной ткани тела.Многоядерные волокна находятся под контролем соматической нервной системы и вызывают движение за счет сил, действующих на скелет, подобно рычагам и шкивам. Ритмическое сокращение сердечной мышцы регулируется синоатриальным узлом, кардиостимулятором сердца. Хотя сердечная мышца — это специализированная поперечно-полосатая мышца, состоящая из удлиненных клеток с множеством центрально расположенных ядер, она не находится под произвольным контролем. Гладкие мышцы выстилают внутренние органы, кровеносные сосуды и дерму, и, как и сердечная мышца, их движения управляются вегетативной нервной системой и, следовательно, не находятся под произвольным контролем.Ядро каждой коротко сужающейся клетки расположено по центру.
Одноклеточные организмы, простые животные и подвижные клетки сложных животных не имеют обширных мышечных систем. Скорее, движение в этих организмах вызывается волосковидными расширениями клеточной мембраны, называемыми ресничками и жгутиками, или цитоплазматическими расширениями, называемыми псевдоподиями.
Получите подписку Britannica Premium и получите доступ к эксклюзивному контенту. Подпишись сейчасЭта статья представляет собой сравнительное исследование мышечных систем различных животных, включая объяснение процесса сокращения мышц.Для описания мышечной системы человека, связанной с вертикальной позой, см. мышечная система человека.
Общие характеристики мышц и движений
Мышцы обеспечивают движения многоклеточных животных и поддерживают осанку. По внешнему виду он напоминает мясо или плоть рыбы. Мышцы — самая многочисленная ткань у многих животных; например, он составляет от 50 до 60 процентов массы тела у многих рыб и от 40 до 50 процентов у антилоп. Некоторые мышцы находятся под сознательным контролем и называются произвольными мышцами.Другие мышцы, называемые непроизвольными мышцами, сознательно не контролируются организмом. Например, у позвоночных мышцы стенок сердца ритмично сокращаются, перекачивая кровь по телу; мышцы стенок кишечника продвигают пищу за счет перистальтики; мышцы стенок мелких кровеносных сосудов сжимаются или расслабляются, контролируя приток крови к различным частям тела. (Эффекты мышечных изменений кровеносных сосудов проявляются в покраснении и побледнении из-за увеличения или уменьшения притока крови к коже, соответственно.)
Мышцы — не единственное средство передвижения животных. Многие протисты (одноклеточные организмы) передвигаются вместо этого, используя реснички или жгутики (активно преодолевая процессы на поверхности клетки, которые продвигают организм через воду). Некоторые одноклеточные организмы способны к амебоидному движению, при котором содержимое клетки перетекает в отростки, называемые псевдоподиями, из тела клетки. Некоторые простейшие с ресничками передвигаются с помощью стержней, называемых мионемами, которые способны быстро сокращаться.
Немышечные способы передвижения важны и для многоклеточных животных.Многие микроскопические животные плавают за счет биения ресничек. Некоторые мелкие моллюски и плоские черви ползают, используя реснички на нижней стороне тела. Некоторые беспозвоночные, которые питаются путем фильтрации частиц из воды, используют реснички для создания необходимых водных потоков. У высших животных лейкоциты используют амебоидные движения, а реснички клеток, выстилающих дыхательные пути, удаляют инородные частицы с нежных мембран.
Мышцы состоят из длинных тонких клеток (волокон), каждая из которых представляет собой пучок более тонких волокон (рис. 1).Внутри каждой фибриллы находятся относительно толстые нити белкового миозина и тонкие нити актина и других белков. Когда мышечное волокно удлиняется или укорачивается, волокна остаются практически постоянной по длине, но скользят друг мимо друга, как показано на рисунке 2. Напряжение в активных мышцах создается поперечными мостиками (т. Е. Выступами толстых волокон, которые прикрепляются к тонким и приложить к ним силы). По мере того, как активная мышца удлиняется или укорачивается, а волокна скользят друг мимо друга, поперечные мосты постоянно отсоединяются и снова прикрепляются в новых положениях.Их действие похоже на натягивание веревки из рук в руки. Некоторые мышечные волокна имеют длину несколько сантиметров, но большинство других клеток составляют лишь доли миллиметра. Поскольку эти длинные волокна не могут адекватно обслуживаться одним ядром, по их длине распределены многочисленные ядра.
миофиламенты в поперечно-полосатой мышцеРис. 2: Расположение миофиламентов в поперечно-полосатой мышце. На верхней диаграмме мышца растянута, а на нижней — сокращена. Толстые волокна — 1.Длина 6 микрометров (0,0016 миллиметра) у поперечно-полосатых мышц позвоночных, но до шести микрометров у некоторых членистоногих.
Encyclopædia Britannica, Inc.Работа, выполняемая мышцами, требует химической энергии, полученной в результате метаболизма пищи. Когда мышцы сокращаются при приложении напряжения и выполнении механической работы, часть химической энергии преобразуется в работу, а часть теряется в виде тепла. Когда мышцы удлиняются при приложении напряжения (например, при медленном опускании веса), используемая химическая энергия вместе с механической энергией, поглощаемой действием, преобразуется в тепло.Выработка тепла — важная функция мышц у теплокровных животных. Дрожь — это мышечная активность, которая выделяет тепло и согревает тело. Точно так же некоторые насекомые перед полетом некоторое время вибрируют крыльями, нагревая мышцы до температуры, при которой они работают лучше всего.
USFWS: американские мидии
Живые породы как важный природный ресурс |
Почему мидии в опасности | Почему мы заботимся? | Есть надежда? | Как вы можете помочь?
Америки Мидии или моллюски — это группа животных, которые настолько неприметны, что их часто принимают за камни.Лежа на На дне озер, рек и ручьев они редко перемещаются и питаются, фильтруя воду для микроскопических частиц пищи. Они могут даже размножаться, не двигаясь. Самец рассеивает сперму, а поток воды несет это к самке, где происходит оплодотворение. Но на протяжении большей части В Северной Америке, и особенно на Среднем Западе, эти похожие на скалы существа отправляют срочное сообщение.
Северная Америка имеет самое большое разнообразие пресноводных мидий в мире.И внутри Северная Америка, исторически на Среднем Западе были одни из самых высоких показателей видов мидий. Однако в настоящее время на Среднем Западе (Миннесота, Висконсин, США) Айова, Миссури, Иллинойс, Индиана и Огайо) более половины из 78 известные виды классифицируются как находящиеся под угрозой исчезновения на федеральном уровне, находящиеся под угрозой исчезновения или Особое беспокойство вызывают государственные виды. Никакая другая группа животных на Среднем Западе находится в столь серьезной опасности.Чтобы представить это в перспективе, The Nature Conservancy сообщает, что около 70 процентов мидий в Северной Америке вымерли. или находятся под угрозой, по сравнению с 16,5% видов млекопитающих и 14,6%. процент видов птиц.
Мидии не в беде просто потому, что они хрупкие существа, которые выход в любом случае. Хотя в основном они ведут сидячий образ жизни, они могут двигаться.Многие виды адаптировались к постоянно меняющимся условиям в ручьях. и реки. Они также могут закрывать свои раковины, чтобы избежать кратковременного воздействия. к токсинам или другим неблагоприятным условиям окружающей среды. Таким образом, мидии суровые существа, которые могут противостоять суровым условиям, если эти условия временные. Тот факт, что так много мидий находится под угрозой на Среднем Западе показывает, что произошли значительные долгосрочные изменения к нашим озерам и водным путям.И эти изменения были настолько драматичными что у этих водных животных проблемы с выживанием.
Жилая Камни как важные природные ресурсы
В Раньше мидии использовались коренными американцами при изготовлении инструментов и посуда. Мидии также были важным источником пищи. После европейского поселение, из ракушек заготавливали жемчужные пуговицы.К началу 1900-х годов этот многомиллионный бизнес был полностью завершен. качаться по реке Миссисипи. Потому что не было никаких элементов управления в этой отрасли мидии были чрезмерно уловлены, и количество многих видов истощился. Это, наряду с использованием пластиковых заменителей, положило конец индустрии жемчужных пуговиц.
В В 1960-х годах японская промышленность культивированного жемчуга начала расширяться.Если крошечный кусочки песка или раковины мидий вставляются в окружающих устриц нарушение силикатом, создающее жемчужину. Тысячи тонн живых мидий собирают из рек Северной Америки каждый год и поставляется в Японию, где кусочки ракушек используются в качестве «семенного жемчуга». Спрос и высокие цены в этой отрасли привели к увеличению случаи незаконного промысла мидий.
Мидии играют важную роль в водной экосистеме. Много разных видов диких животных едят мидий, в том числе енотов, выдр, цапель и цапель. Мидии фильтруют воду для еды и, таким образом, являются системой очистки. Они обычно присутствуют в группах, называемых грядками. Грядки из мидий могут варьироваться размером от менее квадратного фута до многих акров; эти мидии Грядки могут быть твердым «булыжником» на озере, реке или ручье. дно, которое поддерживает другие виды рыб, водных насекомых и червей.
Хотя их жизнь кажется скучной, их репродуктивные стратегии весьма увлекательны. После того, как мужчина разошелся сперматозоидами, которые токами переносятся в самка, где происходит оплодотворение, оплодотворенные яйца трансформируются в личиночное состояние внутри самки. Затем она упаковывает личинок в заманчивую приманку, которая привлечет определенную рыбу.Когда цель Рыба приближается, она изгоняет личинок на рыбу. Личинки прикрепить к жабрам или плавникам рыбы и прокатиться на ней несколько недель пока они продолжают свое превращение в молодую мидию. Когда превращение завершено, они выпадают из целой рыбы, и начинают свою жизнь как молодые взрослые мидии. Мало того, что этот метод репродукции, интересной биологам и исследователям природы, Тот факт, что мидиям для размножения требуются определенные виды рыб, означает что мидии также являются хорошим индикатором здоровья рыбы-хозяина населения.
Почему Неужели мидии так опасны?
Визуализировать река Миссисипи (или река Огайо, или любая из многих рек и ручьи, пересекающие нашу сельскую местность), как это должно было показаться первые европейские исследователи. Берега реки были засажены деревьями, были много отмелей, периодически обнаруживались заторы, а вода была полностью без антропогенных загрязнителей.Рыбы, вероятно, было много, и если вы пробовали песок и камни на дне, вы бы обнаружили изобилие насекомых, червей и других водных беспозвоночных. Затем визуализируйте Миссисипи или другой водный путь, каким он выглядит сегодня. С этими визуализациями имея в виду, нетрудно поверить, что многие животные живущим в этих водных путях сегодня будет нелегко.Но давайте посмотрим на изменения, которые были внесены в наши реки и ручьи и как эти изменения повлияли на мидии.
ПлотиныНа большинстве наших рек построено множество плотин. Будь то гидроэнергетика, рекреация или навигация, плотины — это физический барьер, который является основным угроза для мидий. В жизненный цикл молодых мидий входит: прикрепление к жабрам определенного вида рыб.Эти молодые мидии, которые на самом деле являются личинками, разносятся рыбой в разные районы реки. В конце концов личинка мидии выпадает с жабр, приземляется. снизу, и если это подходящая область, мидия продолжает расти. Если вы ведете оседлый образ жизни, это отличная стратегия для заселения озер и рек. Однако, поскольку плотины препятствие для рыб, они также предотвращают попадание мидий в верхнем течении от движения вниз по течению и наоборот.
Раковина из черного дерева ( Fusconaia ebena ) и ухо слона ( Elliptio crassidens ) мидии резко рассмотреть влияние плотин на распространение мидий. Основным хозяином этих мидий является сельдь скипджека. Скипджек жил в реке Миссисипи. водораздел от городов-побратимов на юг до Мексиканского залива. Это было на самом деле мигрирующий вид в верхнем бассейне реки Миссисипи, где он был известен нерестом в озере Пепин и, вероятно, использовался в других районах водораздел.Но после того, как на Миссисипи была возведена плотина гидроэлектростанции Река в Кеокеке, штат Айова, в 1913 г. мигрируют к северу от плотины. Таким образом, этот вид рыб был истреблен. от верхнего течения реки Миссисипи к северу от плотины в Кеокеке. Вместе с искоренение скипджека, полное (или большинство) воспроизведение черного дерева прекратились ракушечные и слоновьи мидии вверх по течению от плотины.Физическое лицо живые экземпляры этих видов до сих пор встречаются в верховьях Миссисипи. водораздел, потому что эти мидии очень долгожители и были так обильно перед плотиной. Однако размножения у этих видов нет. дольше встречается к северу от плотины в Кеокеке.
Дополнительно выступая в качестве барьера для движения рыбы, плотины воздействуют на мидий, изменяя проточная водная среда рек.Площадь водохранилища выше по течению плотины представляет собой озероподобную среду на участке реки. В большинстве случаев, речные виды мидий в озерах не выживают. Так что все мидии русла в верхнем течении водохранилища плотины ликвидированы. В зависимости от в зависимости от типа плотины, сбросы с плотины не могут имитировать естественные условия, вместо этого неестественно высокие выбросы воды чередуются с неестественно низкие попуски воды.Низкий уровень воды особенно неблагоприятен мидиям (и большинству других речных животных).
Седиментация и загрязнениеСельское хозяйство Производство заставляет эрозию почвы стекать в реки и озера. Мосты также являются участками высокой эрозии и отложений. Большое количество осадка вход в ручьи и реки может похоронить гравий и каменистое дно и задушить моллюски.Многие виды мидий не могут жить на илистой или рыхлой основе. песчаное дно, им нужно, чтобы дно реки было каменным, гравийным или твердым пески. Осадок в сточных водах часто несет с собой пестициды, которые еще больше загрязняет воду. Другие загрязнители, такие как ПХД, ртуть и свинец оседает в водотоках из-за сбросов промышленных предприятий. Широкий с промышленных площадок выделяется множество других токсичных веществ.
С навигацией на многих реках и промышленных комплексах, расположенных на берегах рек, там всегда есть угроза разлива нефти или токсичного химического вещества. Также многие общественные и частные септические системы опорожняются в наши водные пути. В 1970-х годах НЕТ живые мидии были найдены в нижних пятнадцати милях Миннесоты. и реки Миссисипи, от городов-побратимов до озера Пепин.Это было называют пустыней мидий. Плохое качество воды стало причиной того, что эти районы не могли поддерживать мидий. С тех пор выборка указывает что качество воды улучшается. Фактически, недавняя съемка мидий на севере озера Пепин задокументировали присутствие большого количества особей и количество видов мидий.
Экзотика ВидыЭкзотические виды представляют постоянно растущую угрозу для наших местных растений и животные.Что касается местных мидий, заражение мидиями зебры произошло. близкие к катастрофическим последствиям. Скорее всего, мидии были завезены в Великие озера случайно, в результате сброса балластных вод крупными океанскими судами из Европы. Из Великих озер баржи и лодки переносят взрослых особей и личинок в реки Иллинойс и Миссисипи.
Они увеличиваются в количестве быстрее, чем неместные мидии, и прикрепляются к почти любая твердая поверхность, в том числе родные мидии.Они так быстро размножаются и в таком изобилии, что движение, питание и размножение местных мидий поведение подавлено. У одной мидии было более 10000 зебр мидии на нем.
Почему нам не все равно?
- Мониторы здоровья водной среды: наличие разнообразных и воспроизводящихся популяций мидий указывают на здоровую водную систему, что означает хорошую рыбалку, хорошее качество воды для водоплавающих птиц и других диких животных, а также как гарантия того, что наша вода безопасна.И наоборот, когда популяции мидий подвержены риску, это указывает на проблемы для других видов рыб и диких животных, и люди тоже.
- Экологический Ценность: мидии являются естественными фильтрами, питающимися водорослями, планктоном и илы, они помогают очистить водную систему. Мидии также являются важным источник пищи для многих видов диких животных, включая выдр, енотов, Ондатра, цапли, белые цапли и немного рыбы.
- Экономический ценность: пресноводные мидии были и остаются основным экономическим ресурс; сначала в индустрии пуговиц, а теперь и в культивированном жемчуге промышленность. Мидии из Северной Америки составляют ядро культивируемых жемчужная промышленность в Азии. и эстетическая ценность: изучение мидий, их естественная история, и требования к среде обитания дают интересные и важные уроки о взаимосвязанности водной системы и о том, как виды приспосабливаются в их экосистему.
- Культурный значение: Мидии сыграли важную роль в культурной истории доисторические и недавние коренные народы Огайо и Миссисипи Бассейны рек. Их использовали в пищу, а снаряды — для украшения, инструменты и как товар для торговли. Индийская ракушка (груды снарядов, оставленные коренными американцами) расширяются на мили вдоль старых деревень и лагерей вдоль Миссисипи и реки Огайо.
- Биоразнообразие: мидии играли и, надеюсь, будут продолжать играть важную роль в наших водных экосистемах. Учитывая, что менее 20 видов мидий находятся в большинстве других стран мира, в наших североамериканских реки и ручьи действительно «богаты» почти 300 видами !! Мидии не болеют раком. Исследователи хотят знать, почему и мидии могут иметь дополнительные ценности в будущем, которые мы не можем сейчас предсказать.Утрата любого из этих видов обязательно будет иметь последствия. от того, как функционирует водная экосистема.
Пассаж
и выполнение Закона об исчезающих видах и чистой воде
Закон способствовал восстановлению некоторых видов. Находящиеся под угрозой исчезновения
Закон о видах обеспечивает защиту существующих сообществ мидий, содержащих
находящихся под угрозой исчезновения и находящихся под угрозой исчезновения видов и способствовал усилиям по восстановлению
исчезающие мидии.Применение Закона о чистой воде привело к
в улучшенном качестве воды, что позволило повторно заселить мидии
и ловить рыбу в некоторых районах.
Чем вы можете помочь?
Для защиты мидий: можно сделать.
- Консервировать энергии в попытке ограничить строительство новой гидроэлектростанции электростанции.
- Предел или прекратите использование пестицидов, чтобы сохранить почву и предотвратить сток в близлежащие районы. озера и ручьи. Стратегии комплексной борьбы с вредителями (IPM) могут замените использование пестицидов — узнайте об этом больше.
- Помогите контролировать эрозию почвы путем посадки деревьев и растений, чтобы предотвратить сток почвы в пресную воду области.
- Удалить водные сорняки с лодочных прицепов и двигателей перед повторным использованием предотвратить распространение таких вещей, как мидии зебры.НЕ бросайте сорняки обратно в воду. поддержите программы управления водосбором, позвонив в местный ресурс управляющее агентство.
- Поддержка и соблюдать карантин, осмотр и обеззараживание мидий-зебр программы.
Мидии реки Санта-Крус | Моллюски реки Огайо | Угроза зебр-мидий
Мидии Дом
Дом, находящийся под угрозой исчезновения
типов мышечной ткани | Биология I
Мышечные клетки специализируются на сокращении.Мышцы позволяют совершать движения, такие как ходьба, а также облегчают процессы в организме, такие как дыхание и пищеварение. Тело состоит из трех типов мышечной ткани: скелетных мышц, сердечных мышц и гладких мышц (рис. 1).
Рис. 1. Тело состоит из трех типов мышечной ткани: скелетных мышц, гладких мышц и сердечных мышц, визуализированных здесь с помощью светового микроскопа. Гладкомышечные клетки короткие, суженные на каждом конце и имеют только одно пухлое ядро на каждом. Клетки сердечной мышцы разветвленные и поперечно-полосатые, но короткие.Цитоплазма может ветвиться, и у них есть одно ядро в центре клетки. (кредит: модификация работы NCI, NIH; данные шкалы от Мэтта Рассела)
Ткань скелетных мышц образует скелетные мышцы, которые прикрепляются к костям или коже и контролируют передвижение и любое движение, которое можно контролировать сознательно. Скелетную мышцу также называют произвольной мышцей, поскольку ею можно управлять с помощью мысли. Скелетные мышцы длинные и цилиндрические на вид; при рассмотрении под микроскопом ткань скелетных мышц имеет полосатый или полосатый вид.Строчки вызваны регулярным расположением сократительных белков (актина и миозина). Актин представляет собой глобулярный сократительный белок, который взаимодействует с миозином для сокращения мышц. Скелетная мышца также имеет несколько ядер, присутствующих в одной клетке.
Гладкая мышечная ткань встречается в стенках полых органов, таких как кишечник, желудок и мочевой пузырь, а также вокруг проходов, таких как дыхательные пути и кровеносные сосуды. Гладкая мышца не имеет бороздок, не находится под произвольным контролем, имеет только одно ядро на клетку, сужается с обоих концов и называется непроизвольной мышцей.
Ткань сердечной мышцы находится только в сердце, а сердечные сокращения перекачивают кровь по всему телу и поддерживают кровяное давление. Как и скелетная мышца, сердечная мышца имеет поперечно-полосатую форму, но в отличие от скелетных мышц, сердечная мышца не может контролироваться сознательно и называется непроизвольной мышцей. Он имеет одно ядро на клетку, разветвлен и отличается наличием вставочных дисков.
Структура волокон скелетных мышц
Каждое волокно скелетных мышц представляет собой клетку скелетных мышц.Эти клетки невероятно большие, диаметром до 100 мкм и длиной до 30 см. Плазматическая мембрана волокна скелетных мышц называется сарколеммой . Сарколемма — это место проведения потенциала действия, которое вызывает сокращение мышц. Внутри каждого мышечного волокна находится миофибрилл, — длинные цилиндрические структуры, расположенные параллельно мышечному волокну. Миофибриллы проходят по всей длине мышечного волокна, и, поскольку их диаметр составляет всего около 1,2 мкм, внутри одного мышечного волокна можно найти от сотен до тысяч.Они прикрепляются к сарколемме своими концами, так что по мере укорачивания миофибрилл сокращается вся мышечная клетка (рис. 2).
Рис. 2. Клетка скелетных мышц окружена плазматической мембраной, называемой сарколеммой, с цитоплазмой, называемой саркоплазмой. Мышечное волокно состоит из множества фибрилл, собранных в упорядоченные единицы.
Поперечно-полосатый вид ткани скелетных мышц является результатом повторяющихся полос белков актина и миозина, которые присутствуют по длине миофибрилл.Темные полосы A и светлые полосы I повторяются вдоль миофибрилл, а выравнивание миофибрилл в клетке приводит к тому, что вся клетка выглядит полосатой или полосчатой.
Рис. 3. Саркомер — это область от одной Z-линии до следующей Z-линии. Многие саркомеры присутствуют в миофибриллах, что приводит к полосатости, характерной для скелетных мышц.
Каждая полоса I имеет плотную линию, проходящую вертикально через середину, называемую диском Z или линией Z. Z-диски обозначают границу единиц, называемых саркомеров , которые являются функциональными единицами скелетных мышц.Один саркомер — это пространство между двумя последовательными Z-дисками и содержит одну целую полосу А и две половины полосы I. Миофибриллы состоят из множества саркомеров, расположенных по ее длине, и когда саркомеры индивидуально сокращаются, миофибриллы и мышечные клетки укорачиваются (рис. 3).
Миофибриллы состоят из более мелких структур, называемых миофиламентами . Существует два основных типа волокон: толстые волокна и тонкие волокна; у каждого свой состав и расположение. Толстые нити встречаются только в полосе А миофибриллы. Тонкие нити прикрепляются к белку в Z-диске, называемому альфа-актинином, и встречаются по всей длине I-полосы и частично в A-полосе. Область, в которой перекрываются толстые и тонкие волокна, имеет более плотный вид, так как между ними мало места. В середине темной полосы есть вертикальная линия, называемая линией М, на которой дополнительные белки удерживают вместе толстые волокна. И Z-диск, и линия M удерживают миофиламенты на месте, чтобы поддерживать структурное расположение и наслоение миофибрилл.
Толстые и тонкие нити сами состоят из белков. Толстые нити состоят из белкового миозина. Хвост молекулы миозина соединяется с другими молекулами миозина, образуя центральную область толстой нити около линии M, тогда как головки выравниваются по обе стороны от толстой нити, где тонкие нити перекрываются. Основным компонентом тонких филаментов является белок актин. Два других компонента тонкой нити — тропомиозин и тропонин. Актин имеет сайты связывания для прикрепления миозина.Нити тропомиозина блокируют сайты связывания и предотвращают актин-миозиновые взаимодействия, когда мышцы находятся в состоянии покоя. Тропонин состоит из трех глобулярных субъединиц. Одна субъединица связывается с тропомиозином, одна субъединица связывается с актином, а одна субъединица связывает ионы Ca 2+ .
Посмотрите это видео, демонстрирующее организацию мышечных волокон.
Единая клональная линия трансмиссивного рака идентифицирована у двух видов морских мидий в Южной Америке и Европе
Существенные изменения:
1) Секвенирование дополнительных локусов необходимо, чтобы окончательно показать, что BTN2 в Чили / Аргентине на самом деле является тем же штаммом рака, который обнаружен в Европе.Кроме того, секвенирование менее 1 кб по 3 локусам затрудняет выводы о происхождении и гибридизации. Например, как они определили, что «раковые» аллели EF1a не были вариантами числа копий у хозяина? Были ли проверены доброкачественные ткани на отсутствие этих аллелей у носителей? Наконец, при таком небольшом секвенировании, почему авторы уверены, что они не являются числами секвенирования, которые являются обычными для многих видов морских беспозвоночных. Чтобы отреагировать на это, потребуется дополнительное секвенирование.
Теперь мы включили дополнительные данные о последовательности, которые подтверждают эти утверждения. У нас есть данные о последовательности из дополнительного митохондриального локуса (mt COI ), которые подтверждают, что аллели из M. edulis и M. chilensis идентичны и происходят от M. trossulus , и что они отличаются от . Mytilus BTN1 (рисунок 4). Мы обнаружили, что два образца из Чили показали свидетельство рекомбинации (единственное свидетельство рекомбинации, обнаруженное до сих пор во всех протестированных локусах).Мы также использовали массив SNP на раковых мидиях, а также большую выборку здоровых людей, чтобы идентифицировать наличие SNP, уникальных для M. trossulus (рис. 6). Это позволило нам изучить многие независимые ядерные локусы, чтобы подтвердить, что раковые клетки имеют происхождение M. trossulus и что этот сигнал из SNP M. trossulus не может быть обнаружен ни у одного нормального животного из M. chilensis . или M. edulis популяций.
Относительно вопроса о том, могут ли дополнительные аллели быть просто вариацией числа копий, авторы обзора правы в том, что одного только обнаружения множественных аллелей недостаточно для подтверждения заявления о трансмиссивном раке — ключевой вывод состоит в том, что существует множество аллелей и что некоторые из них являются связанными с раком аллелями, которые идентичны у независимых людей.Например, можно ожидать, что дупликация генома у индивидуума M. chilensis приведет к возникновению множественных аллелей, филогенетически связанных с нормальными аллелями M. chilensis . Не ожидается, что это приведет к возникновению второго аллеля, который почти идентичен аллелю другого вида. И не ожидается, что будет получена одна и та же последовательность у нескольких особей двух разных видов.
Кроме того, мы не обнаружили свидетельств наличия множественных аллелей в нормальных образцах.При проведении ПЦР и секвенирования общей ДНК невозможно отличить истинный митохондриальный ген от переноса ядра в митохондрии (numt). Рецензенты правы, что мы не можем этого исключить, и мы разъяснили это в тексте. Однако, если бы один из связанных с раком аллелей «mtCR» имел номер, например, это не изменило бы вывод о том, что существует последовательность, полученная из митохондриальной ДНК M. trossulus , которая почти идентична при раке у нескольких индивидуумов в два вида.
2) Генные деревья и деревья видов часто противоречат друг другу, и авторы не выбрали достаточное количество локусов и достаточного количества сливающейся истории, чтобы разрешить это несоответствие. Несоответствие генов в геноме теперь скорее правило, чем исключение. Единая слитная история бесполой линии передачи BTN должна помочь улучшить некоторые из них, но необходимо более трех маркеров. Кроме того, в том же духе, подчеркивают сами авторы, их «дерево» несовместимо с аллелями G и H, происходящими от BTN1S.Но это основано на одном локусе, чего недостаточно.
Генные деревья и деревья видов часто противоречат друг другу, но, как отмечают обозреватели, клональный трансмиссивный рак является бесполым, поэтому мы не ожидаем увидеть несоответствия между деревом генов и деревом родословной рака, за исключением случая рекомбинации. Мы действительно видим некоторое несоответствие в образцах нормальных половых мидий, как и ожидалось (например, M. chilensis интрогрессировали последовательность M. trossulus в локусе EF1alpha , и небольшое количество особей оказались гибридами), но мы видим отсутствие несоответствий между какими-либо деревьями генов в любом из четырех локусов, за исключением митохондриальной рекомбинации в одном локусе в двух чилийских образцах.Кроме того, во всех локусах деревья генов поддерживают клональную экспансию почти идентичной бесполой линии, происходящей от M. trossulus и отличной от Mytilus BTN1. Мы специально выделили доказательства того, что ни у аллелей G, ни у H не может быть общего предка рака с аллелем S, как уже отмечалось, но мы специально не упомянули, что каждый другой локус предоставляет аналогичные доказательства. Например, в h5 нормальные образцы M. trossulus были идентифицированы с аллелями, которые были точно идентичны аллелям, обнаруженным как в Mytilus BTN1, так и в 2.Поэтому маловероятно, что BTN1 и 2 произошли от одного человека, и несколько локусов разошлись, чтобы стать более близкими к другим нормальным последовательностям в популяции. Мы разъяснили это в тексте. Добавление четвертого локуса и данные из 13 SNP, специфичных для M. trossulus , также должны предоставить дополнительные доказательства этих утверждений.
Проблемы, влияющие на интерпретацию данных: ответ должен включать либо существенное уточнение, либо переписывание разделов:
1) Что подразумевается под «дробью» М.trossulus «SNP? Сколько? Может быть, это ошибка секвенирования?
Это было краткое объяснение ранее опубликованных данных, в которых массив SNP использовался для анализа разнообразия в популяции из особей M. edulis , и это было сформулировано нечетко. Авторы этого исследования отметили, что в некоторых локусах присутствует сигнал флуоресценции, соответствующий аллелю M. trossulus , который не должен присутствовать в популяции. Однако более неожиданным оказался тот факт, что аллели не были ожидаемой долей от общего числа аллелей.Ожидается, что один SNP будет присутствовать в 0% (гомозиготный по аллелю 1), 50% (гетерозиготный по обоим аллелям 1 и 2) или 100% (гомозиготный по аллелю 2) ДНК в одном локусе в образце, но в этих образцах доля аллелей M. trossulus находилась между этими числами (например, 30%). Это предполагает, что тестируемый образец ДНК может представлять собой комбинацию двух геномов, один из которых представляет собой нормальный геном M. edulis , а другой — рак из M. trossulus . Это само по себе явно неубедительно и привело к текущему изучению этих и других образцов.Мы попытались расширить и прояснить наше объяснение этого в тексте.
2) Что подразумевается под: Мы секвенировали… и обнаружили больше, чем два обычных аллеля в «большинстве» из них… Что такое «большинство»? Таких примеров много.
В каждом случае неоднозначных слов, таких как «многие» или «большинство», мы заменяли их конкретными числами. В специально упомянутом случае «большая часть» из четырех образцов составляла три образца.
3) Результаты количественной ПЦР при попытке клонирования аллелей болезни сбивают с толку.Обнаружение того, что у двух из четырех животных, пораженных M. chilensis, есть высокие уровни аллеля, ассоциированного с раком, а у двух других нет — предполагают ли авторы еще одну передающуюся линию рака?
Это вполне вероятно. Основываясь на полном отсутствии аллелей Mytilus BTN2 во всех четырех локусах в этих двух образцах M. chilensis , мы заключаем, что эти два образца не имеют Mytilus BTN2. Таким образом, они либо 1) были неправильно диагностированы, 2) имеют обычные виды рака, которые не передаются, или 3) имеют еще одну трансмиссивную линию рака, которая еще не описана.Мы разъяснили это в тексте.
4) Учитывая невозможность последовательной амплификации аллелей рака в различных образцах, неясно, как можно сделать какие-либо выводы относительно вклада канадских образцов и M. trossulus.
Мы не можем последовательно клонировать аллели из некоторых образцов рака, но мы можем последовательно амплифицировать связанные с раком аллели с помощью кПЦР из всех образцов, диагностированных как злокачественные, из M. edulis и M.chilensis , за исключением двух образцов M. chilensis , которые отрицательны по всем четырем локусам (и поэтому не имеют Mytilus BTN2). Мы включили анализ канадских образцов M. trossulus , чтобы определить, можно ли обнаружить аллели Mytilus BTN2 в нормальной популяции или в образцах, положительных по Mytilus BTN1. Мы не нашли ни одного образца M. trossulus с аллелями, которые соответствуют Mytilus BTN2 по всем локусам (действительно, канадский M.trossulus , вообще амплифицированный с помощью специфичных для рака праймеров EF1alpha ). Это показало, что эти диагностические праймеры последовательно идентифицируют Mytilus BTN2 и что Mytilus BTN2 не был обнаружен в этой популяции M. trossulus и отличается от Mytilus BTN1.
5) Также неясно, как интерпретировать тот факт, что аллели, связанные с раком h5, обнаружены у нескольких нормальных животных M. trossulus. Сколько это «некоторые»?
Мы прояснили неоднозначные слова в тексте, например «некоторые.В частности, поскольку все трансмиссивные раковые заболевания произошли от одного нормального человека, у которого был первый рак, который дал начало клону, ожидается, что аллели, почти соответствующие раку, могут быть обнаружены у нормальных животных в популяции. Следовательно, ожидается, что некоторые нормальные животные M. trossulus будут иметь аллели h5 , почти идентичные Mytilus BTN1, а некоторые другие будут иметь аллели, почти идентичные Mytilus BTN2. Примечательно, что нормальных животных с Mytilus, BTN2 или BTN1-подобными аллелями во всех локусах пока не обнаружено.Два человека, у которых впервые возникли эти линии рака, могли бы, но эти два человека давно мертвы. Мы подробно остановились на этом в тексте.
6) Секвенирование менее 1kb по 3 локусам затрудняет выводы о происхождении и гибридизации. Например, как они определили, что «раковые» аллели EF1a не были вариантами числа копий у хозяина? Были ли проверены доброкачественные ткани на отсутствие этих аллелей у носителей? В частности, если рассматривать только нормальные аллели, существует четкая группировка, основанная на видах происхождения в филогенетическом дереве. Есть три исключения из этой кластеризации, от одного нормального M.trossulus (MW59) и два больных M. chilensis (Mch51 и Castro26) с некоторыми отчетливо подобными M. edulis аллелями. Они могут отражать интрогрессию или истинные гибридные особи — MW59 также имеет аллель EF1α, соответствующий M. edulis, и, вероятно, представляет собой гибрид этого вида, который был завезен в Канаду (Crego-Prieto et al., 2015). шум, хотя и ожидаемый, становится проблематичным, когда он охватывает треть данных. Пожалуйста, объясните более подробно ваше обоснование ваших выводов и включите обсуждение предостережений.
Как отмечалось выше, генных деревьев, показанных в этом исследовании, было бы недостаточно для разрешения слияния и полной эволюционной истории трех половых видов Mytilus , тестируемых здесь.
У нас действительно есть доказательства как интрогрессии (в локусе EF1alpha ), так и гибридных особей (таких как MW59, отмеченных выше). Однако данные об аллелях, связанных с раком, убедительно подтверждают клональные, бесполые линии, и все четыре дерева полностью согласуются (за исключением обнаружения митохондриальной рекомбинации у двух индивидуумов).
https://doi.org/10.7554/eLife.47788.025Древние саркомерные миозины, обнаруженные в специализированных мышцах | Скелетная мышца
Хартман М.А., Спудич Я.А. Краткий обзор суперсемейства миозинов. J Cell Sci. 2012; 125: 1627–32.
Артикул CAS PubMed PubMed Central Google Scholar
Маскарелло Ф., Тониоло Л., Канчеллара П., Реджиани С., Маккатроццо Л. Экспрессия и идентификация 10 саркомерных изоформ MyHC в скелетных мышцах человека различного эмбриологического происхождения.Разнообразие и сходство видов млекопитающих. Анн Анат. 2016; 207: 9–20.
Артикул Google Scholar
Скьяффино С., Росси А.С., Смерду В., Лейнванд Л.А., Реджиани С. Миозины развития: паттерны экспрессии и функциональное значение. Скелетная мышца. 2015; 5: 1–14.
Артикул CAS Google Scholar
Horton MJ, Rosen C, Close JM, Sciote JJ. Количественная оценка РНК тяжелой цепи миозина в мышцах гортани человека: дифференциальная экспрессия в вертикальном и горизонтальном задних перстневидных мышцах и тиреоаритеноидах.Ларингоскоп. 2008. 118: 472–7.
Артикул CAS Google Scholar
Horton MJ, Brandon CA, Morris TJ, Braun TW, Yaw KM, Sciote JJ. Обильная экспрессия РНК тяжелой цепи миозина IIB в подмножестве волокон жевательных мышц человека. Arch Oral Biol. 2001; 46: 1039–50.
Артикул CAS PubMed PubMed Central Google Scholar
Harrison BC, Allen DL, Leinwand LA.IIb или не IIb? Регуляция экспрессии гена тяжелой цепи миозина у мышей и мужчин. Скелетная мышца. 2011; 1: 1–9.
Артикул CAS Google Scholar
Stedman HH, Kozyak BW, Nelson A, Thesier DM, Su LT, Low DW, et al. Мутация гена миозина коррелирует с анатомическими изменениями человеческого происхождения. Природа. 2004; 428: 415–8.
Артикул CAS Google Scholar
Скьяффино С., Реджиани С. Типы волокон в скелетных мышцах млекопитающих. Physiol Rev.2011; 91: 1447–531.
Артикул CAS Google Scholar
Hoh JF. Сверхбыстрый или жевательный миозин и эволюция мышц, закрывающих челюсти, у позвоночных. J Exp Biol. 2002; 205: 2203–10.
PubMed Google Scholar
Ресницоу Д., Дикон Дж. К., Уоррик Х. М., Спудич Дж. А., Лейнванд Л. А..Функциональное разнообразие в семействе миозиновых моторов скелетных мышц человека. Proc Natl Acad Sci. 2010; 107: 1053–8.
Артикул CAS Google Scholar
Аксель Т., Чой Ю Э, Саттон С., Руппель К.М., Спудич Дж. Изменения силы ансамбля, которые являются результатом мутаций сердечного миозина человека и низкомолекулярного эффектора. Cell Rep. 2015; 11: 910–20.
Артикул CAS PubMed PubMed Central Google Scholar
Harris DE, Work SS, Wright RK, Alpert NR, Warshaw DM. Сила миозина гладких, сердечных и скелетных мышц и генерация движения оцениваются с помощью механических взаимодействий поперечных мостиков in vitro. J Muscle Res Cell Motil. 1994; 15: 11–9.
Артикул CAS Google Scholar
Барани М. АТФазная активность миозина коррелировала со скоростью укорачивания мышц. J Gen Physiol. 1967; 50: 197–218.
Артикул PubMed PubMed Central Google Scholar
Walklate J, Дживз Массачусетс. Температурный коллектор для машины с остановленным потоком, позволяющий проводить измерения от -10 до + 40 ° C. Анальная биохимия. 2015; 476: 11–6.
Артикул CAS Google Scholar
Weiss A, McDonough D, Wertman B, Acakpo-Satchivi L, Montgomery K, Kucherlapati R, et al. Организация кластеров генов тяжелой цепи скелетного миозина человека и мыши является высококонсервативной. Proc Natl Acad Sci. 1999; 96: 2958–63.
Артикул CAS Google Scholar
Saez LJ, Gianola KM, McNally EM, Feghali R, Eddy R, Shows TB, et al. Гены тяжелой цепи сердечного миозина человека и их связь в геноме. Nucleic Acids Res. 1987; 15: 5443–59.
Артикул CAS PubMed PubMed Central Google Scholar
Desjardins PR, Burkman JM, Shrager JB, Allmond LA, Stedman HH. Эволюционные последствия трех новых членов семейства генов тяжелой цепи саркомерного миозина человека.Mol Biol Evol. 2002; 19: 375–93.
Артикул CAS Google Scholar
Тониоло Л., Канчеллара П., Маккатроццо Л., Патруно М., Маскарелло Ф., Реджиани С. Обнародование жевательного миозина: первое определение сократительных параметров мышечных волокон челюстных мышц хищников. Am J Physiol Cell Physiol. 2008; 295: C1535–42.
Артикул CAS Google Scholar
McGuigan K, Phillips PC, Postlethwait JH. Эволюция генов тяжелой цепи саркомерного миозина: данные рыб. Mol Biol Evol. 2004; 21: 1042–56.
Артикул CAS Google Scholar
Икеда Д., Оно Ю., Снелл П., Эдвардс Ю.К., Элгар Дж., Ватабе С. Дивергентная эволюция семейства генов тяжелой цепи миозина у рыб и четвероногих: данные сравнительного геномного анализа. Physiol Genomics. 2007; 32: 1–15.
Артикул CAS Google Scholar
Rossi AC, Mammucari C, Argentini C, Reggiani C, Schiaffino S. Два новых / древних миозина в скелетных мышцах млекопитающих: MYh24 / 7b и MYh25 экспрессируются в экстраокулярных мышцах и мышечных веретенах. J Physiol. 2010. 588: 353–64.
Артикул CAS Google Scholar
Роулерсон А., Маскарелло Ф., Веггетти А., Карпен Э. Волокнистый состав мышц первой жаберной дуги у Carnivora и Primates. J Muscle Res Cell Motil.1983; 4: 443–72.
Артикул CAS Google Scholar
Роулерсон А., Поуп Б., Мюррей Дж., Уэлен Р. Б., Weeds AG. Новый миозин присутствует в мышцах кошачьих, закрывающих челюсти. J Muscle Res Cell Motil. 1981; 2: 415–38.
Артикул CAS Google Scholar
Hoh JFY, Lim JHY, Kang LDH, Lucas CA. Экспрессия сверхбыстрого миозина в мышцах, закрывающих челюсти, Crocodylus porosus .В: Григг Г.С., Сибахер Ф., Франклин К.Э., редакторы. Биология и эволюция крокодилов: Суррей Битти и сыновья; 2001. с. 156–64.
Шмидт-Реза А. Эволюция систем органов. Оксфорд: издательство Оксфордского университета; 2007.
Google Scholar
Mackie GO, Singla CL. Распространение импульса и сокращение в оболочке сложной асцидии. Biol Bull. 1987. 173: 188–204.
Артикул CAS Google Scholar
Schuchert P, Reber-Müller S, Schmid V. Специфическая экспрессия тяжелой цепи миозина на жизненной стадии у гидрозоа Podocoryne carnea. Дифференциация. 1993; 54: 11–8.
Артикул CAS Google Scholar
Зайпель К., Шмид В. Эволюция поперечно-полосатой мускулатуры: медузы и происхождение триплобластики. Dev Biol. 2005. 282: 14–26.
Артикул CAS Google Scholar
Steinmetz PRH, Kraus JEM, Larroux C, Hammel JU, Amon-Hassenzahl A, Houliston E, et al. Независимая эволюция поперечнополосатых мышц у книдарий и билатерий. Природа. 2012; 487: 231–4.
Артикул CAS PubMed PubMed Central Google Scholar
Гудсон Х., Спудич Дж. Молекулярная эволюция семейства миозинов: взаимосвязи, полученные из сравнения аминокислотных последовательностей. Proc Natl Acad Sci. 1993; 90: 659–63.
Артикул CAS Google Scholar
OOta S, Saitou N. Филогенетическая взаимосвязь мышечных тканей, выведенная из наложения генов деревьев. Mol Biol Evol. 1999; 16: 856–67.
Артикул CAS Google Scholar
Брюнет Т., Фишер А.Х., Штейнмец П.Р., Лаури А., Бертуччи П., Арендт Д. Эволюционное происхождение двунаправленных гладких и поперечно-полосатых миоцитов.Элиф. 2016; 5: 1–24.
Артикул Google Scholar
Sciote JJ, Horton MJ, Rowlerson AM, Link J. Специализированные черепные мышцы: чем они отличаются от мышц конечностей и брюшного пресса? Клетки Тканевые Органы. 2003. 174: 73–86.
Артикул PubMed PubMed Central Google Scholar
Портер Дж. Д., Бейкер Р. С., Рагуза Р. Дж., Брюкнер Дж. К.. Экстраокулярные мышцы: основные и клинические аспекты структуры и функции.Surv Ophthalmol. 1995; 39: 451–84.
Артикул CAS PubMed PubMed Central Google Scholar
Спенсер РФ, Портер Дж. Д.. Биологическая организация экстраокулярных мышц. Prog Brain Res. 2006; 151: 43–80.
Артикул PubMed PubMed Central Google Scholar
Purves D, Augustine G, Fitzpatrick D, Hall W, LaMantia A, White L. Действия и иннервация экстраокулярных мышц.В кн .: Неврология. 5-е изд: Sinauer Associates, Inc; 2012. с. 436–40.
Портер Дж. Д.. Экстраокулярная мышца: клеточные адаптации для разнообразного функционального репертуара. Ann N Y Acad Sci. 2002; 956: 7–16.
Артикул PubMed PubMed Central Google Scholar
Васики Р., Зия-Газвини Ф, Блумер Р., Лукас Дж. Р., Майр Р. Типы мышечных волокон экстраокулярных мышц человека: гистохимическое и иммуногистохимическое исследование.Исследование Ophthalmol Vis Sci. 2000; 41: 980–90.
CAS Google Scholar
Джейкоби Дж., Ко К., Вайс К., Рашбрук Дж. Систематические вариации экспрессии миозина вдоль экстраокулярных мышечных волокон взрослой крысы. J Muscle Res Cell Motil. 1990; 11: 24–40.
Артикул Google Scholar
Бриггс М.М., Шахат Ф. Сверхбыстрый экстраокулярный миозин (MYh23) локализован в зоне иннервации как в глобальном, так и в глазничном слоях экстраокулярной мышцы кролика.J Exp Biol. 2002; 205: 3133–42.
CAS PubMed Google Scholar
Рубинштейн Н.А., Портер Дж.Д., Хох Дж.Ф. Развитие продольной изменчивости изоформ миозина в орбитальных волокнах экстраокулярных мышц крыс. Исследование Ophthalmol Vis Sci. 2004. 45: 3067–72.
Артикул Google Scholar
Лим С.Дж., Юнг Х.Х., Чо Я. Постнатальное развитие изоформ тяжелой цепи миозина в экстраокулярных мышцах крыс.Mol Vis. 2006; 12: 243–50.
CAS PubMed Google Scholar
Zhou Y, Liu D, Kaminski HJ. Экспрессия тяжелой цепи миозина в экстраокулярной мышце мыши: более сложная, чем ожидалось. Исследование Ophthalmol Vis Sci. 2010. 51: 6355–63.
Артикул Google Scholar
Парк К., Лим Дж., Сон С., Йеул С. Экспрессия изоформы тяжелой цепи миозина в экстраокулярных мышцах человека: продольные вариации и паттерны экспрессии в глобальном и орбитальном слоях.Мышечный нерв. 2012; 45: 713–20.
Артикул CAS Google Scholar
Wieczorek DF, Periasamy M, Butler-Browne GS, Whalen RG, Nadal-Ginard B. Коэкспрессия множества генов тяжелой цепи миозина в дополнение к тканеспецифической в экстраокулярной мускулатуре. J Cell Biol. 1985; 101: 618–29.
Артикул CAS PubMed PubMed Central Google Scholar
Уинтерс Л.М., Бриггс М.М., Шахат Ф. Ген тяжелой цепи миозина экстраокулярных мышц человека (MYh23) отображается в кластере быстрых генов миозина развития на хромосоме 17. Геномика. 1998. 54: 188–9.
Артикул CAS Google Scholar
Бриггс М.М., Шахат Ф. Ранняя специализация сверхбыстрого миозина в экстраокулярных и гортанных мышцах. J Exp Biol. 2000; 203: 2485–94.
CAS PubMed PubMed Central Google Scholar
Шахат Ф, Бриггс ММ. Филогенетические последствия сверхбыстрого миозина в экстраокулярных мышцах. J Exp Biol. 2002; 205: 2189–201.
CAS PubMed Google Scholar
Кьеллгрен Д., Торнелл Л., Андерсен Дж., Педроса-Домеллоф Ф. Изоформы тяжелой цепи миозина в экстраокулярных мышцах человека. Исследование Ophthalmol Vis Sci. 2003. 44: 1419–25.
Артикул Google Scholar
Будак М.Т., Богданович С., Визен М.Х., Лозинская О., Хурана Т.С., Рубинштейн Н.А. Слоистые различия экспрессии генов в экстраокулярных мышцах, идентифицированные с помощью лазерной микроскопии. Physiol Genomics. 2004. 20: 55–65.
Артикул CAS Google Scholar
Кханна С., Мерриам А.П., Гонг Б., Лихи П., Портер Дж.Д. Комплексное профилирование экспрессии по классам мышечной ткани и определение молекулярной ниши экстраокулярной мышцы.FASEB J. 2003; 17: 1370–2.
Артикул CAS Google Scholar
Портер Дж. Д., Ханна С., Рао Дж. С., Ли Дж., Андраде Ф. Х. Экстраокулярная мышца определяется принципиально отличным профилем экспрессии генов. Proc Natl Acad Sci. 2001; 98: 12062–7.
Артикул CAS Google Scholar
Zhou Y, Cheng G, Dieter L, Hjalt TA, Andrade FH, Stahl JS, et al. Измененный фенотип при условном нокауте Pitx2 в экстраокулярной мышце.Исследование Ophthalmol Vis Sci. 2009; 50: 4531–41.
Артикул Google Scholar
Китамура К., Миура Х., Миягава-Томита С., Янадзава М., Катох-Фукуи Ю., Сузуки Р. и др. Дефицит Pitx2 мыши приводит к аномалиям вентральной стенки тела, сердца, экстра- и периокулярной мезодермы и изомерии правого легкого. Разработка. 1999; 126: 5749–58.
CAS PubMed Google Scholar
Diehl AG, Zareparsi S, Qian M, Khanna R, Angeles R, Gage PJ. Морфогенез экстраокулярных мышц и экспрессия генов регулируются дозой гена Pitx2. Исследование Ophthalmol Vis Sci. 2006; 47: 1785–93.
Артикул Google Scholar
Zhou Y, Liu D, Kaminski HJ. Pitx2 регулирует экспрессию изоформы тяжелой цепи миозина и множественную иннервацию в экстраокулярной мышце. J Physiol. 2011; 589: 4601–14.
Артикул CAS PubMed PubMed Central Google Scholar
Hebert SL, Daniel ML, McLoon LK. Роль Pitx2 в поддержании фенотипа миогенных клеток-предшественников в экстраокулярных мышцах. PLoS One. 2013; 8
Conti-Fine BM, Milani M, Kaminski HJ. Миастения гравис: прошлое, настоящее и будущее. J Clin Invest. 2006; 116: 2843–54.
Артикул CAS PubMed PubMed Central Google Scholar
Ю Вай Ман CY, Chinnery PF, Griffiths PG. Экстраокулярные мышцы обладают совершенно разными свойствами, которые делают их избирательно уязвимыми для определенных заболеваний.Нервно-мышечное расстройство. 2005; 15: 17–23.
Артикул CAS Google Scholar
Prabhakar BS, Bahn RS, Smith TJ. Современные взгляды на патогенез болезни Грейвса и офтальмопатии. Endocr Rev.2003; 24: 802–35.
Артикул CAS Google Scholar
Камински Х.Дж., Мазен А., Ли Р.Дж., Катирджи МБ, Рафф Р.Л. Экстраокулярные мышцы защищены от распространенной дистрофии Дюшенна.Энн Нейрол. 1992; 32: 586–8.
Артикул CAS Google Scholar
Андраде Ф. Х., Портер Дж. Д., Камински Х. Дж. Сохранение глазных мышц при мышечных дистрофиях: уроки, которые следует усвоить? Microsc Res Tech. 2000; 48: 192–203.
Артикул CAS Google Scholar
Портер Дж. Д., Мерриам А. П., Кханна С., Андраде Ф. Х., Ричмондс С. Р., Лихи П. и др. Конститутивные свойства, а не молекулярные адаптации, опосредуют сохранение экстраокулярных мышц у дистрофических мышей MDX.FASEB J. 2003; 17: 893–5.
Артикул CAS Google Scholar
Kallestad KM, Hebert SL, Mcdonald AA, Daniel ML, Cu SR, Mcloon LK. Сохранение экстраокулярной мышцы при старении и мышечных дистрофиях: гипотеза миогенных клеток-предшественников. Exp Cell Res. 2011; 317: 873–85.
Артикул CAS PubMed PubMed Central Google Scholar
Verma M, Fitzpatrick KR, McLoon LK.Восстановление и регенерация экстраокулярных мышц. Curr Ophthalmol Rep. 2017; 5: 207–15.
Артикул PubMed PubMed Central Google Scholar
Хант С. Мышечное веретено млекопитающих: периферические механизмы. Physiol Rev.1990; 70: 643–63.
Артикул CAS Google Scholar
Matthews PBC. Развитие взглядов на внутреннюю работу и функциональную роль мышечного веретена.J Physiol. 1981; 320: 1–30.
Артикул CAS PubMed PubMed Central Google Scholar
Hulliger M. Мышечное веретено млекопитающих и его центральный орган управления. Rev Physiol Biochem Pharmacol. 1984; 101: 1–110.
CAS PubMed Google Scholar
Tourtellotte WG, Milbrandt J. Сенсорная атаксия и агенез мышечного веретена у мышей, лишенных фактора транскрипции Egr3.Нат Жене. 1998. 20: 87–91.
Артикул CAS Google Scholar
Blum KP, Lamotte D’Incamps B, Zytnicki D, Ting LH. Кодирование силы в мышечных веретенах при растяжении пассивной мышцы. PLoS Comput Biol. 2017; 13: 1–24.
Артикул CAS Google Scholar
Бэнкс Р., Стейси М. Количественные исследования мышечных веретен млекопитающих. Бостон: Спрингер; 1988 г.
Google Scholar
Банки RW. Аллометрический анализ количества мышечных веретен в скелетных мышцах млекопитающих. J Anat. 2006. 208: 753–68.
Артикул CAS PubMed PubMed Central Google Scholar
Matthews PBC. Там, где шла анатомия, за ней следовала физиология: обзор нашего развивающегося понимания мышечного веретена, того, что оно делает и как работает.J Anat. 2015; 227: 104–14.
Артикул PubMed PubMed Central Google Scholar
Ovalle WK. Тонкая структура интрафузальных мышечных волокон крыс. J Cell Biol. 1971; 51: 83–103.
Артикул PubMed PubMed Central Google Scholar
Thornell LE, Carlsson L, Eriksson PO, Liu JX, Österlund C, Stål P, et al. Типирование интрафузальных волокон.J Anat. 2015; 227: 136–56.
Артикул PubMed PubMed Central Google Scholar
Лю Дж., Эрикссон П., Торнелл Л., Педроса-Домеллоф Ф. Содержание клетчатки и состав тяжелых цепей миозина в мышечных веретенах в двуглавой мышце пожилого человека Brachii. J Histochem Cytochem. 2005. 53: 445–54.
Артикул CAS Google Scholar
Николс Т., Коуп Т. Механизмы поперечного моста, лежащие в основе зависимых от истории свойств мышечных веретен и рефлексов растяжения.Может J Phys Pharm. 2004. 82: 569–76.
Артикул CAS Google Scholar
Миллер А.Дж. Краниомандибулярные мышцы: их роль в функциях и формах: CRC Press; 1991.
Bredman JJ, Wessels A, Weijs WA, Korfage JA, Soffers CA, Moorman AF. Демонстрация «кардиоспецифической» тяжелой цепи миозина в жевательных мышцах человека и кролика. Histochem J. 1991; 23: 160–70.
Артикул CAS Google Scholar
Korfage JA, Van Eiiden TM. Изоформный состав миозина медиальной и латеральной крыловидных мышц человека. J Dent Res. 2000. 79: 161–169.
Артикул Google Scholar
Korfage JA, Koolstra JH, Langenbach GEJ, van Eijden TMGJ. Волокнистый состав мышц челюсти человека (часть 2), роль гибридных волокон и факторов, ответственных за индивидуальные различия. J Dent Res. 2005; 84: 784–93.
Артикул CAS Google Scholar
English AW, Eason J, Pol M, Schwartz G, Shirley A. Различные фенотипы среди волокон жевательной мышцы кролика, содержащих тяжелые цепи медленного / бета-миозина: новый тип разнообразия во взрослых мышцах. J Muscle Res Cell Motil. 1998. 19: 525–35.
Артикул CAS Google Scholar
Исон Дж.М., Шварц Г.А., Павлат Г.К., англ. AW. Половая диморфная экспрессия тяжелых цепей миозина у взрослых жевательных мышей. J Appl Physiol.2000; 89: 251–8.
Артикул CAS Google Scholar
Boom HPW, van Spronsen PH, van Ginkel FC, van Schijndel RA, Castelijns JA, Tuinzing DB. Сравнение площади и объема поперечного сечения мышц челюсти у людей с длинным и коротким лицом с помощью МРТ. Arch Oral Biol. 2008. 53: 273–81.
Артикул CAS Google Scholar
Китаи Н., Крейборг С., Бакке М., Паулсен Х.У., Мёллер Е., Дарванн Т.А. и др.Трехмерное магнитно-резонансное изображение нижней челюсти и жевательных мышц в случае ювенильного хронического артрита, леченного с помощью аппарата Herbst. Угол Ортод. 2002; 72: 81–7.
PubMed Google Scholar
Sciote JJ, Raoul G, Ferri J, Close J, Horton MJ, Rowlerson A. Функция массетера и неправильный прикус скелета. Преподобный Стоматол Чир Максиллофак Чир Орале. 2013; 114: 79–85.
CAS PubMed Google Scholar
Скьяффино С., Реджиани С. Молекулярное разнообразие миофибриллярных белков: регуляция генов и функциональное значение. Physiol Rev.1996; 76: 371–423.
Артикул CAS Google Scholar
Равоса М.Дж., Нин Дж., Костли Д.Б., Дэниел А.Н., Сток С.Р., Стэк МС. Жевательная биомеханика и пластичность жевательного волокна. J Musculoskelet Neuronal Interact. 2010; 10: 46–55.
CAS PubMed Google Scholar
Чочон Р.Л., Нисбетт Р.А., Корруччини Р.С. Последовательность питания и черепно-лицевое развитие, связанные с жевательной функцией мини-свиней. J Craniofac Genet Dev Biol. 1997; 17: 96–102.
CAS PubMed Google Scholar
Whiteley AT, Kendrick GS, Matthews JL. Влияние функции на костные и мышечные ткани черепно-лицевой области крысы. Угол Ортод. 1966; 36: 13–7.
CAS PubMed Google Scholar
Килиаридис С., Шю до н.э. Изометрическое мышечное напряжение, создаваемое стимуляцией жевательного аппарата после длительного изменения режима питания растущих крыс. Arch Oral Biol. 1988; 33: 467–72.
Артикул CAS Google Scholar
Сато И., Кониси К. Влияние мягкой диеты на развитие митохондрий жевательных мышц крыс. Okajimas Folia Anat Jpn. 2004. 81: 67–74.
Артикул Google Scholar
Langenbach G, van de Pavert S, Savalle W., Korfage H, van Eijden T. Влияние консистенции пищи на жевательные мышечные волокна кролика. Eur J Oral Sci. 2003; 111: 81–4.
Артикул Google Scholar
He T. Черепно-лицевая морфология и рост у хорьков: эффекты от изменения жевательной функции. Swedish Dent J Suppl. 2004; 165: 1–72.
Google Scholar
Китагава Ю., Митера К., Огасавара Т., Нодзё Ю., Мияути К., Сано К. Изменения в гистохимических характеристиках ферментов жевательной мышцы, вызванные длительной мягкой диетой у растущих кроликов. Oral Dis. 2004; 10: 271–6.
Артикул CAS Google Scholar
Miehe B, Fanghänel J, Kubein-Meesenburg D, Nägerl H, Schwestka-Polly R. Жевательная мускулатура в измененных окклюзионных отношениях — модельное исследование с экспериментальными животными.Анн Анат. 1999; 181: 37-40.
Артикул CAS Google Scholar
Сайто Т., Охнуки Ю., Ямане А., Саеки Ю. Влияние последовательности диеты на мРНК тяжелой цепи миозина жевательной мышцы крысы во время постнатального развития. Arch Oral Biol. 2002; 47: 109–15.
Артикул CAS Google Scholar
Смит А., Вебер К.М., Ньютон Дж., Денни М. Развитие и возрастные изменения рефлексов системы закрытия челюсти человека.Электроэнцефалогер Клин Нейрофизиол. 1991. 81: 118–28.
Артикул CAS Google Scholar
Косиони А.Е., Карказис ХК. ЭМГ-исследование влияния старения на рефлекс жевательного рефлекса человека. Геродонтология. 1994; 11: 30–8.
Артикул CAS Google Scholar
Newton JP, Yemm R, Abel RW, Menhinick S. Изменения в мышцах челюсти человека с возрастом и состоянием зубов.Геродонтология. 1993; 10: 16–22.
Артикул CAS Google Scholar
Monemi M, Eriksson PO, Eriksson A, Thornell LE. Неблагоприятные изменения в составе типа волокон жевательной мышцы человека по сравнению с двуглавой мышцей плеча во время старения. J Neurol Sci. 1998. 154: 35–48.
Артикул CAS Google Scholar
Eriksson PO, Thornell LE. Гистохимические и морфологические характеристики мышечных волокон жевательной мышцы человека, медиального крыловидного отростка и височных мышц.Arch Oral Biol. 1983; 28: 781–95.
Артикул CAS Google Scholar
Monemi M, Eriksson PO, Dubail I, Butler-Browne GS, Thornell LE. Тяжелая цепь миозина плода увеличивается в жевательных мышцах человека при старении. FEBS Lett. 1996; 386: 87–90.
Артикул CAS Google Scholar
Monemi M, Liu JX, Thornell L, Eriksson P. Состав тяжелых цепей миозина боковых крыловидных и двубрюшных мышц человека у молодых людей и пожилых людей.J Muscle Res Cell Motil. 2000; 21: 303–12.
Артикул CAS Google Scholar
Österlund C, Lindström M, Thornell LE, Eriksson PO. Выдающаяся неоднородность в составе тяжелых цепей миозина у молодых жевательных мышц человека по сравнению с молодыми двуглавыми мышцами плеча. Histochem Cell Biol. 2012; 138: 669–82.
Артикул CAS Google Scholar
Рикеттс РМ.Основа цефалометрической коммуникации. Am J Orthod. 1960. 46: 330–57.
Артикул Google Scholar
Ингерваль Б., Хелькимо Э. Сила жевательных мышц и морфология лица у человека. Arch Oral Biol. 1978; 23: 203–6.
Артикул CAS Google Scholar
Килиаридис С. Влияние жевательных мышц на черепно-лицевой рост. Acta Odontol Scand.1995; 53: 196–202.
Артикул CAS Google Scholar
Weijs WA, Hillen B. Взаимосвязь между поперечным сечением жевательных мышц и формой черепа. J Dent Res. 1984; 63: 1154–7.
Артикул CAS Google Scholar
Килиаридис С., Келебо П. Толщина жевательной мышцы, измеренная с помощью УЗИ, и ее связь с морфологией лица. J Dent Res.1991; 70: 1262–5.
Артикул CAS Google Scholar
Van Spronsen PH, Weijs WA, Valk J, Prahl-Andersen B, Van Ginkel FC. Сравнение поперечных сечений челюстных мышц длиннолицых и нормальных взрослых. J Dent Res. 1992; 71: 1279–85.
Артикул Google Scholar
Benington PC, Gardener JE, Hunt NP. Объем жевательной мышцы, измеренный с помощью УЗИ, и его связь с морфологией лица.Eur J Orthod. 1999; 21: 659–70.
Артикул CAS Google Scholar
Proffit WR, Fields HW, Nixon WL. Окклюзионные силы у взрослых с нормальным и удлиненным лицом. J Dent Res. 1983; 62: 566–70.
Артикул CAS Google Scholar
Hunt NP, Cunningham SJ. Влияние ортогнатической хирургии на окклюзионную силу у пациентов с вертикальными деформациями лица.Int J Oral Maxillofac Surg. 1997; 26: 87–91.
Артикул CAS Google Scholar
Piancino MG, Isola G, Merlo A, Dalessandri D, Debernardi C, Bracco P. Тип жевания и мышечная активация у пациентов с открытым прикусом. J Electromyogr Kinesiol. 2012; 22: 273–9.
Артикул Google Scholar
Роулерсон А., Рауль Дж., Даниэль Й, Клоуз Дж., Маурадж, Калифорния, Ферри Дж. И др.Различия в типах волокон жевательных мышц, связанные с различной морфологией лица. Am J Orthod Dentofac Orthop. 2005; 127: 37–46.
Артикул Google Scholar
Boyd SB, Gonyea WJ, Finn RA, Woodard CE, Bell WH. Гистохимическое исследование жевательной мышцы у пациентов с вертикальным избытком верхней челюсти. J Oral Maxillofac Surg. 1984. 42: 75–83.
Артикул CAS Google Scholar
Бреннан М., Антонишин О. Влияние манипуляции височной мышцей на рост черепа: экспериментальное исследование. Plast Reconstr Surg. 1996; 97: 13–24.
Артикул CAS Google Scholar
Hohl TH. Транспозиция жевательных мышц у приматов: влияние на черепно-лицевой рост. J Maxillofac Surg. 1983; 11: 149–56.
Артикул CAS Google Scholar
Zhu M, Zhao W, Jia L, Lu J, Qiao T., Qu Q. Самый старый сочлененный остейхтиан показывает мозаичные персонажи гнатомоя. Природа. 2009; 458: 469–74.
Артикул CAS Google Scholar
Forey P, Janvier P. Agnathans и происхождение челюстных позвоночных. Природа. 1993; 361: 129–34.
Артикул Google Scholar
Brazeau MD, Friedman M. Происхождение и ранняя филогенетическая история челюстных позвоночных.Природа. 2015; 520: 490–7.
Артикул PubMed PubMed Central Google Scholar
Канг ЛХД, Ругани А, Уокер М.Л., Бестак Р, Хох JFY. Экспрессия жевательных специфических изоформ тяжелой цепи миозина, миозин-связывающего протеина-С и тропомиозина в мышечных волокнах и культурах сателлитных клеток жевательных мышц кошек. J Histochem Cytochem. 2010. 58: 623–34.
Артикул CAS PubMed PubMed Central Google Scholar
van Rooij E, Liu N, Olson EN. МикроРНК сгибают мышцы. Тенденции Genet. 2008. 24: 159–66.
Артикул CAS Google Scholar
van Rooij E, Quiat D., Johnson BA, Sutherland LB, Qi X, Richardson JA, et al. Семейство микроРНК, кодируемых генами миозина, регулирует экспрессию миозина и работу мышц. Dev Cell. 2009; 17: 662–73.
Артикул CAS PubMed PubMed Central Google Scholar
Нагасе Т, Кикуно Р., Исикава К., Хиросава М., Охара О. Прогнозирование кодирующих последовательностей неидентифицированных генов человека. IX. Полные последовательности 100 новых клонов кДНК из мозга, которые могут кодировать большие белки in vitro. ДНК Res. 2000; 7: 143–50.
Артикул CAS Google Scholar
Bell ML, Buvoli M, Leinwand LA. Разделение экспрессии интронной микроРНК и ее гена-хозяина миозина путем пропуска экзона. Mol Cell Biol.2010; 30: 1937–45.
Артикул CAS PubMed PubMed Central Google Scholar
Warkman AS, Whitman SA, Miller MK, Garriock RJ, Schwach CM, Gregorio CC, et al. Экспрессия развития и регуляция сердечной транскрипции Myh7b, третьей тяжелой цепи миозина в сердце позвоночных. Цитоскелет. 2012; 69: 324–35.
Артикул CAS Google Scholar
Rubio MD, Johnson R, Miller CA, Huganir RL, Rumbaugh G. Регулирование структуры и функции синапсов с помощью различных двигателей миозина II. J Neurosci. 2011; 31: 1448–60.
Артикул CAS PubMed PubMed Central Google Scholar
Haraksingh RR, Jahanbani F, Rodriguez-Paris J, Gelernter J, Nadeau KC, Oghalai JS, et al. Секвенирование экзома и картирование вариантов числа копий по всему геному выявляют новые ассоциации с нейросенсорной наследственной потерей слуха.BMC Genomics. 2014; 15: 1–19.
Артикул CAS Google Scholar
Эспозито Т., Сампаоло С., Лимонгелли Г., Вароне А., Формикола Д., Диодато Д. и др. Дигенное мутационное наследование генов интегрина альфа 7 и тяжелой цепи миозина 7B вызывает врожденную миопатию с некомпактной кардиомиопатией левого желудочка. Orphanet J Rare Dis. 2013; 8: 1–13.
Артикул Google Scholar
Hanchard NA, Swaminathan S, Bucasas K, Furthner D, Fernbach S, Azamian MS, et al. Полногеномное исследование ассоциации врожденных сердечно-сосудистых левосторонних поражений показывает ассоциацию с локусом на хромосоме 20. Hum Mol Genet. 2016; 25: 2331–41.
Артикул CAS PubMed PubMed Central Google Scholar
Браун К.М., МакГрегор С., Монтгомери Г.В., Крейг Д.В., Чжао З.З., Иядурай К. и др. Общие варианты последовательностей на 20q11.22 указывают на предрасположенность к меланоме. Нат Жене. 2008; 40: 838–40.
Артикул CAS PubMed PubMed Central Google Scholar
Chatzinasiou F, Lill CM, Kypreou K, Stefanaki I., Nicolaou V, Spyrou G, et al. Всесторонний полевой синопсис и систематический метаанализ исследований генетической ассоциации меланомы кожи. J Natl Cancer Inst. 2011; 103: 1227–35.
Артикул CAS PubMed PubMed Central Google Scholar
Antonopoulou K, Stefanaki I, Lill CM, Chatzinasiou F, Kypreou KP, Karagianni F, et al. Обновленный полевой синопсис и систематические мета-анализы исследований генетических ассоциаций меланомы кожи: база данных MelGene. J Invest Dermatol. 2015; 135: 1074–9.
Артикул CAS Google Scholar
Мачида С., Нода С., Такао А., Наказава М., Мацуока Р. Экспрессия гена тяжелой цепи 2 медленного скелетного миозина в клетках волокна Пуркинье в сердце цыпленка.Biol Cell. 2002; 94: 389–99.
Артикул CAS Google Scholar
Nasipak BT, Kelley DB. Геном диплоидного бесхвостого амурана Xenopus tropicalis содержит новый набор генов тяжелой цепи саркоплазматического миозина, экспрессируемых в личиночных мышцах и гортани. Dev Genes Evol. 2008; 218: 389–97.
Артикул CAS PubMed PubMed Central Google Scholar
Garriock RJ, Meadows SM, Krieg PA. Экспрессия развития и сравнительный геномный анализ генов тяжелой цепи сердечного миозина Xenopus. Dev Dyn. 2005; 233: 1287–93.
Артикул CAS Google Scholar
Лонсдейл Дж., Томас Дж., Сальваторе М., Филлипс Р., Ло Е., Шад С. и др. Проект экспрессии генотипа в ткани (GTEx). Нат Жене. 2013; 45: 580–5.
Артикул CAS Google Scholar
Меле М., Феррейра П., Ревертер Ф, ДеЛука Д., Монлонг Дж., Саммет М. и др. Человеческий транскриптом в тканях и у людей. Наука. 2015; 348: 660–5.
Артикул CAS PubMed PubMed Central Google Scholar
Miao X, Luo Q, Zhao H, Qin X. Анализ коэкспрессии и идентификация связанных с плодовитостью длинных некодирующих РНК в яичниках овец. Научный доклад 2016; 6: 1–10.
Артикул CAS Google Scholar
Miao X, Luo Q, Zhao H, Qin X. Комплексный анализ miRNA и метилированных генов, кодирующих мРНК и lncRNA, у пород овец с разной плодовитостью. Front Physiol. 2017; 8: 1–14.
Артикул Google Scholar
Hansel NN, Paré PD, Rafaels N, Sin DD, Sandford A, Daley D, et al. Полногеномное ассоциативное исследование, выявление новых локусов, связанных с реактивностью дыхательных путей при хронической обструктивной болезни легких.Am J Respir Cell Mol Biol. 2015; 53: 226–34.
Артикул CAS PubMed PubMed Central Google Scholar
Бэр Л.А., Моррисон А.С., Роуленд С.М., Шиффман Д., Люк М.М., Якубова О.А. и др. Пять распространенных вариантов генов определяют повышенный генетический риск ишемической болезни сердца. Genet Med. 2007; 9: 682–9.
Артикул CAS Google Scholar
Люк М.М., Лалушек В., Роуленд С.М., Катанезе Дж. Дж., Болоник Дж. И., Буй Н. Д. и др.Полиморфизмы, связанные как с некардиоэмболическим инсультом, так и с ишемической болезнью сердца: Венский регистр инсульта. Cerebrovasc Dis. 2009. 28: 499–504.
Артикул PubMed PubMed Central Google Scholar
Morrison AC, Bare LA, Chambless LE, Ellis SG, Malloy M, Kane JP, et al. Прогнозирование риска ишемической болезни сердца с использованием шкалы генетического риска: исследование риска атеросклероза в сообществах. Am J Epidemiol. 2007. 166: 28–35.
Артикул Google Scholar
Йошино С., Киллуффо Р., Бест П., Аткинсон Е., Аоки Т., Каннингем Дж. И др. Полиморфизм одиночных нуклеотидов, связанный с аномальной функцией микрососудов коронарных сосудов. Coron Artery Dis. 2014; 25: 281–9.
Артикул PubMed PubMed Central Google Scholar
Нири М.Т., Нири Дж. М., Лунд Г. К., Холт Т. Н., Гарри Ф. Б., Мохун Т. Дж. И др.Тяжелая цепь 15 миозина связана с давлением в легочной артерии крупного рогатого скота. Pulm Circ. 2014; 4: 496–503.
Артикул CAS PubMed PubMed Central Google Scholar
Цинь Х., Хсу МХ, Моррис Б.Дж., Хох JFY. Отдельный подкласс поперечно-полосатых миозинов млекопитающих: структура и молекулярная эволюция «сверхбыстрой» или жевательной тяжелой цепи миозина. J Mol Evol. 2002; 55: 544–52.
Артикул CAS Google Scholar
Zhu J, Sanborn JZ, Diekhans M, Lowe CB, Pringle TH, Haussler D. Сравнительный геномный поиск утраты давно установленных генов в человеческом происхождении. PLoS Comput Biol. 2007; 3: 2498–509.
CAS Google Scholar
Канг Л., Хьюз С., Петтигрю Дж., Хох Дж. Экспрессия гена тяжелой цепи миозина, специфичная для челюсти, в мышцах овец, собак, обезьян, летучих лисиц и микробов, закрывающих челюсти. Basic Appl Myol. 1994; 4: 381–92.
Google Scholar
Hoh JF, Kim Y, Sieber LG, Zhong WW, Lucas CA. Мышцы, закрывающие челюсти, кенгуру экспрессируют тяжелую цепь альфа-сердечного миозина. J Muscle Res Cell Motil. 2000; 21: 673–80.
Артикул CAS Google Scholar
Reiser PJ, Bicer S, Chen Q, Zhu L, Quan N. Жевательная («сверхбыстрая») тяжелая цепь миозина и легкая цепь 1 миозина эмбриона / предсердия в мышцах, закрывающих челюсти грызунов. J Exp Biol. 2009; 212: 2511–9.
Артикул CAS Google Scholar
Брюнет М., Гай Ф, Пилбим Д., Маккей Х. Т., Ликиус А., Ахунта Д. и др. Новый гоминид из верхнего миоцена Чада, Центральная Африка. Природа. 2002; 418: 145–51.
Артикул CAS PubMed PubMed Central Google Scholar
Уокер А., Лики Р. Э., редакторы. Скелет нариокотома Homo erectus. Кембридж: издательство Гарвардского университета; 1993.
Google Scholar
Тобиас П.В., Ущелье О. Том 4: черепа, эндокасты и зубы homo habilis. Кембридж: Издательство университета; 1991.
Google Scholar
Като С., Саеки Ю., Янагисава К. Чувствительность к Ca 2+ и временные реакции натяжения на растяжение длины шага в препаратах челюстных мышц кошачьих с механической зачисткой. Arch Oral Biol. 1985; 30: 429–32.
Артикул CAS PubMed PubMed Central Google Scholar
Perry GH, Verrelli BC, Stone AC. Сравнительный анализ раскрывает сложную историю молекулярной эволюции MYh26 человека. Mol Biol Evol. 2005. 22: 379–82.
Артикул CAS PubMed PubMed Central Google Scholar
Прюфер К., Расимо Ф., Паттерсон Н., Джей Ф., Санкарараман С., Сойер С. и др. Полная последовательность генома неандертальца из Горного Алтая. Природа. 2014; 505: 43–9.
Артикул CAS PubMed PubMed Central Google Scholar
Perry GH, Kistler L, Kelaita MA, Sams AJ. Понимание фенотипа и эволюции рациона гомининов на основе данных о древних последовательностях ДНК. J Hum Evol. 2015; 79: 55–63.
Артикул PubMed PubMed Central Google Scholar
Макколлум М.А., Шервуд С.К., Виньярд С.Дж., Лавджой СО, Шахат Ф. Об эволюции черепа с мышцами и эволюции человеческого мозга: история, стоящая за заголовками MYh26. J Hum Evol. 2006; 50: 232–6.
Артикул PubMed PubMed Central Google Scholar
Карри П. Генетика человека: изучение эволюции гоминидов. Природа. 2004; 428: 373–4.
Артикул CAS PubMed PubMed Central Google Scholar
Oh HJ, Choi D, Goh CJ, Hahn Y. Потеря функции генов и эволюция фенотипов человека. BMB Rep. 2015; 48: 373–9.
Артикул CAS PubMed PubMed Central Google Scholar
Для некоторых видов мышца означает воду
Исследователи из Университета штата Аризона обнаружили, что детские питоны превращают мышцы в воду для себя и своего потомства, но за это приходится платить.© Сандра Леандер / ASUДля животных в засушливом климате поиск воды является ключом к выживанию. Группа исследователей нашла одно место, где его находят некоторые виды, — это их мышцы.
«У большинства животных нет передвижной столовой», — сказал Джордж Бруш IV, докторант Университета штата Аризона и ведущий автор статьи в Proceedings of the Royal Society B. «Вопрос в том, не двигаются ли они. и они не пьют, как они сами выживают? А что насчет их потомства? »
Бруш возглавлял группу исследователей из Университета штата Калифорния и Центра биологических исследований в Шизе во Франции, изучающих, как крупные селекционеры — виды, использующие накопленную энергию для поддержания воспроизводства, — могут обеспечивать водой себя и свое потомство.В число крупных заводчиков входят гремучие змеи недалеко от дома в Аризоне, но команда Бруша вместо этого предпочла изучить Австралийского детского питона ( Antaresia childreni ), который размножается в засушливый сезон, когда природных водных ресурсов особенно мало, и в это время ничего не потребляет.
Бруш знал, что для крупных селекционеров жир важен для преобразования в энергию. А как насчет воды? А что с мышцами? По его словам, в то время как жир состоит всего лишь на 10 процентов из воды, мышцы на 75 процентов состоят из воды.
Его команда обнаружила, что во время размножения мышцы играют важную роль не только в обеспечении энергией, но и в производстве воды, когда в противном случае они были недоступны.
Изучая змей в лаборатории, исследователи сравнили репродуктивных и непродуктивных питонов.
