
Сгибание ног в тренажере лежа — SportWiki энциклопедия
- Похожая статья: Сгибание ног на тренажере для женщин
Инвентарь:тренажёр.
Мышцы, работающие при сгибании ног на тренажёре лёжа:1 — полуперепончатая; 2 — двуглавая мышца бедра; 3 — полусухожильная;4 — икроножнаяОсновные мышцы:задняя поверхность бедра.
Дополнительные мышцы: икроножная.
Уровень подготовки: от начального до продвинутого.
Основная нагрузка приходится на заднюю поверхность бедра.
Шаг 1. Лягте лицом вниз на тренажёр, чтобы валики были чуть выше пяток.
Шаг 2. На выдохе медленно согните ноги в коленях с максимальной амплитудой.
Шаг 3. На вдохе медленно вернитесь в начальное положение.
Следите за тем, чтобы во время движения корпус не отрывался от поверхности тренажёра, так как это снимет нагрузку с мышц бедра, перераспределив её на ягодичные мышцы. Вариант, в котором сгибания выполняются одной ногой, отлично подойдет для изолированного тренинга и при негармоничном развитии бёдер.
Сгибания голени vs тяга на прямых ногах[править | править код]
Принято считать, что тренировки со свободными весами более эффективны, чем занятия на тренажёрах. Однако из всех правил бывают исключения. Тренировка бицепса бедра в сгибаниях голени лёжа обеспечит больший мышечный рост, чем тяга штанги на прямых ногах. Такие результаты опубликовал в Journal of Strength and Conditioning американский учёный Брэд Шенфельд [1].
В эксперименте принимали участие 10 мужчин, ранее занимавшихся с отягощениями. Спортсмены выполняли тягу штанги на прямых ногах и сгибания голени в тренажёре лёжа. Мышечная активность и электропроводимость измерялись во время выполнения упражнений прикреплёнными к мышцам электродами. Чем больше электрическая активность в мышце, тем больше мышца напряжена и больший стимул к росту она получает.
Сгибания ног вызвали больше электрической активности в нижней части поверхности бедра, чем тяга на прямых ногах. Также сгибания мобилизовали больше мышечных волокон во внешней и внутренней частях двуглавой мышцы бедра.
Верхняя часть задней поверхности бедра одинаково реагировала как на сгибания, так и на тягу на прямых ногах. Таким образом, по мере выполнения повторений в сгибаниях голени, степень напряжения мышц была в совокупности выше, что говорит о большей изоляции бицепса бедра в данном упражнении. Однако это не означает, что тяга на прямых ногах — неэффективное упражнение.
«Достаточно будет несколько увеличить время длительности подхода, например замедлением темпа, и выявленные недостатки будут компенсированы появлением большего количества факторов роста в мышечных клетках, а также дополнительным рекрутированием новых мышечных волокон.»
| Сгибания голени | Тяга на прямых ногах | |
|---|---|---|
| Нижняя внешняя часть | 110 | 40 |
| Нижняя внутренняя часть | 79 | 42 |
| Верхняя внешняя часть | 82 | 75 |
| Верхняя внешняя часть | 120 | 122 |
- ↑ J Strength Cond Res.
 2015 Jan; 29(1):159-64
2015 Jan; 29(1):159-64
Сгибание ног в тренажере сидя
6 минут на освоение. 345 просмотров
AtletIQ — приложение для бодибилдинга
600 упражнений, более 100 программ тренировок на массу, силу, рельеф для дома и тренажерного зала. Это фитнес-револиция!
Общая информация
Тип усилияДругоеЖимНетСтатическиеТяга
Вид упражнения
СиловоеРастяжкаКардиоПлиометрическоеStrongmanКроссфитПауэрлифтингТяжелая атлетикаСтрейчингово-силовое упражнениеЙогаДыханиеКалланетика
Тип упражнения
БазовоеИзолирующееНет
Сложность
НачинающийПрофессионалСредний
Целевые мышцы
Сгибание ног в тренажере сидя видео
Как делать упражнение
- Отрегулируйте тренажёр в соответствии с ростом. Спина должна плотно прижиматься к спинке.
- Валики должны оказаться под ногами, чуть ниже икр. Закрепите удерживающую платформу на бёдрах немного выше коленей. Держитесь за боковые ручки тренажёра. Носки смотрят вверх (или же вы можете выбрать другое положение). Ноги вытянут перед собой. Это исходное положение.
- На выдохе, сгибая колени, опускайте ноги к задней части бедра как можно сильнее. Туловище не подвижно. Задержитесь на секунду.
- На вдохе медленно вернитесь в исходное положение.
- Повторите необходимое количество раз.
Внимание: Работайте с адекватным весом. Тело не должно подниматься или раскачиваться. Это может вызвать травмы нижней части спины и подколенного сухожилия.
Варианты: Это упражнение можно выполнять с различным положением стоп.
Фото с правильной техникой выполнения
Какие мышцы работают?
При соблюдении правильной техники выполнения упражнения «Сгибание ног в тренажере сидя» работают следующие группы мышц: Бедра, а также задействуются вспомогательные мышцы:Вес и количество повторений
Количество повторений и рабочий вес зависит от вашей цели и других параметров. Но общие рекомендации могут быть представлены в виде таблицы:
| Цель | Подходы | Повторений | Вес, %1Rm | Отдых м/у подходами |
|---|---|---|---|---|
| Развитие силы | 2-6 | 1-5 раз | 100-85% | 3-7 мин |
| Набор массы | 3-6 | 6-12 раз | 85-60% | 1-4 мин |
| Сушка, рельеф | 2-4 | 13-25 раз | 60-40% | 1-2 мин |
Сделать тренинг разнообразнее и эффективнее можно, если на каждой тренировке изменять количество повторений и вес снаряда.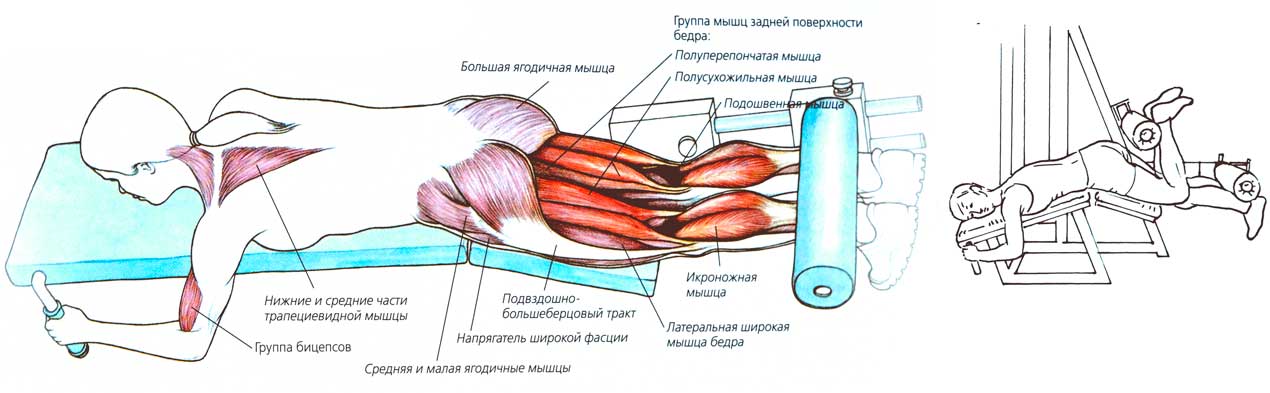
*Укажите вес снаряда и максимальное количество повторений, которое можете выполнить с этим весом.
Не хотите считать вручную? Установите приложение AtletIQ!
- Электронный дневник тренировок
- Помнит ваши рабочие веса
- Считает нагрузку под вас
- Контролирует время отдыха
Лучшие программы тренировок с этим упражнением
Среди программ тренировок, в которых используется упражнение «Сгибание ног в тренажере сидя» одними из лучших по оценкам спортсменов являются эти программы:
Чем заменить?
Вы можете попробовать заменить упражнение «Сгибание ног в тренажере сидя» одним из этих упражнений.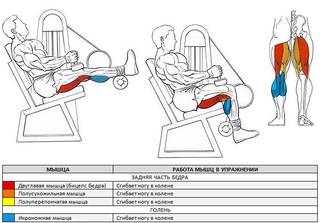 Возможность замены определяется на основе задействуемых групп мышц.
Возможность замены определяется на основе задействуемых групп мышц.Сгибание ног в тренажере сидя Author: AtletIQ: on Сгибание ног в тренажере сидя — польза упражнения, как правильно выполнять и сколько подходов делать.. Rating: 5
Сгибание ног в тренажере сидя: техника, мышцы, альтернатива
Сгибание ног в тренажере сидя — упражнение для изолированной проработки бицепса бедра. Это упражнение подойдет как для новичков, так как не требует специальной подготовки и обладает простой техникой выполнения, так и для профессионалов и спортсменов со стажем, желающим приобрести красивый рельеф и придать правильную форму мышцам ног.
Рекомендуется выполнять сгибание ног в тренажере сидя после выполнения тяжелых базовых упражнений, например приседаний со штангой или жима ногами. Этим вы сможете добиться того самого чувства жжения в мышцах благодаря эффекту пампинга, то есть накачки мышц кровью.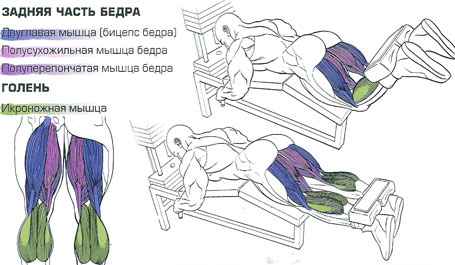 К тому же упражнение на сгибание ног просто обязано присутствовать в вашей программе тренировки ног, потому что оно развивает зачастую отстающую группу мышц-сгибателей коленного сустава. Сбалансированное и пропорциональное развитие как мышц-сгибателей, так и мышц-разгибателей (по сути эти группы мышц являются антагонистами) снижает риск возникновения травмы колена!
К тому же упражнение на сгибание ног просто обязано присутствовать в вашей программе тренировки ног, потому что оно развивает зачастую отстающую группу мышц-сгибателей коленного сустава. Сбалансированное и пропорциональное развитие как мышц-сгибателей, так и мышц-разгибателей (по сути эти группы мышц являются антагонистами) снижает риск возникновения травмы колена!
Какие мышцы работают
- вся группа седалищно-подколенных мышц бедра: бицепс бедра (длинная и короткая головки), полуперепончатая мышца, полусухожильная мышца
- икроножные мышцы (латеральная и медиальная головки)
Техника выполнения
Сгибание ног в тренажере сидя, исходное положение- Настройте положение подушек рычага тренажера для сгибания ног в соответствии с вашим ростом. Для этого сначала настройте положение спинки сидения так, чтобы она поддерживала спину прямой во время выполнения упражнения. Сядьте в тренажер, вытяните ноги и положите голени на подушки рычага (валики должны оказаться примерно на 5-10 см ниже икр).
- Зафиксируйте бедра специальной подушкой-упором расположенной чуть выше колен.
- Ухватитесь руками за рукоятки тренажера, чтобы оставаться в одном положении во время выполнения упражнения.
- На выдохе начинайте подтягивать вес к себе, выполняя сгибание ног в коленях. Старайтесь держать корпус неподвижно. Максимально, на сколько это возможно, согните ноги и подержите напряжение в мышца секунду.
- На вдохе медленно возвращайтесь в исходное положение, разгибая ноги и опуская вес. Повторите упражнение.

Положение стоп
Меняя положение стоп при выполнении упражнения вы меняете и акцент нагрузки:
- Носки вытянуты вперед. Сгибая ноги с вытянутыми носками стоп вперед, вы акцентируете основную долю нагрузки на седалищно-подколенных мышцах.
- Носки повернуты к себе. Если носки стоп повернуть к себе, то часть нагрузки перейдет на икроножные мышцы.
- Носки смотрят в стороны.
 Если при сгибании ног носки развернуть в стороны, нагрузка сместится на внешнюю часть бицепса бедра.
Если при сгибании ног носки развернуть в стороны, нагрузка сместится на внешнюю часть бицепса бедра. - Носки повернуты внутрь. Повернув носки внутрь, а пятки наружу, вы сосредоточите основное усилие на проработке внутренней части бицепса бедра.
Альтернатива
Список аналогичных упражнений, которыми можно заменить сгибание ног в тренажере сидя:
- Сгибание ноги в тренажере стоя
- Сгибание ног в тренажере лежа
- Гиперэкстензии на бицепса бедра
- Сгибание ноги в кроссовере лежа
- Сгибание ног с резинкой
Советы
- Не используйте чрезмерно большой вес в этом упражнении, чтобы не получить травму коленей и сухожилий. Старайтесь делать контролируемое сгибание и разгибание, ни в коем случае не бросайте вес.
- Для концентрической проработки мышц можно выполнять это упражнение одной ногой, то есть поочередно делать подходы для каждой ноги в отдельности.
Сгибание ног в тренажере какие мышцы работают.
 Сгибание ног лежа в тренажере: все секреты упражнения
Сгибание ног лежа в тренажере: все секреты упражненияЗадняя поверхность бедра зачастую оказывается без должного внимания. Да, базовые приседания прорабатывают все группы мышц, расположенные на ногах.
Однако, если вы хотите достичь высоких результатов во формировании красивого и пропорционального тела, то без сгибаний ног лёжа это сделать довольно трудно.
Ведь оно относится к классу изолирующих упражнений, которые позволяют выделить и проработать отдельную мышечную группу.
В этой статье мы подробно разберём сгибания ног лёжа, правильную технику их выполнения и распространённые ошибки.
Популярное упражнение на сгибание ног в положении лежа позволяет эффективно проработать, в отличие от других, именно бицепс (заднюю поверхность) бедра.
Сгибание ног лежа, будучи формирующим упражнением, не наращивает мышечную массу, а лишь укрепляет ее и подчеркивает рельеф.
Поэтому выполнять его стоит после основного комплекса упражнений в качестве завершающего этапа тренировки.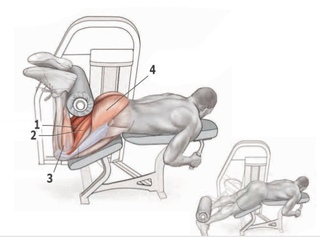
Какие мышцы работают
Основная работа мышц в упражнении заключается в сгибании/разгибании ног в колене.
В большей степени задействованы мышцы:
- Двуглавая (находится на внешней стороне задней поверхности бедра – бицепс).
- Полуперепончатая (находится на внутренней стороне задней поверхности бедра).
- Полусухожильная (находится на внутренней стороне задней поверхности бедра, покрывает полуперепончатую мышцу).
- Икроножная (верхняя и серединная часть задней поверхности голени).
Стабилизаторами выступают мышцы верхней части туловища (нижние и средние части трапециевидных и передние зубчатые мышцы, бицепсы).
Благодаря регулярному выполнению упражнения проявляется рельеф задней поверхности бедра, выравниваются визуальные пропорции (особенно сзади), а также снижается риск травматизма этой группы мышц при выполнении других упражнений.
Варианты выполнения упражнения
Сгибание ног в положении лежа не имеет каких-либо вариаций для выполнения, однако, существуют смежные изолирующие упражнения для сгибания и разгибания ног, различающиеся лишь расположением тела.
К ним относятся сгибания ног сидя и сгибания ног стоя. Основным (базовым) упражнением для наращивания бицепса бедра является становая тяга на прямых ногах.
Техника выполнения упражнения
Подготовка к выполнению и исходное положение
Перед началом упражнения необходимо отрегулировать положение валика в зависимости от длины ног. Он должен упираться в заднюю часть лодыжек на расстоянии примерно трех пальцев поперек ноги от пятки.
В исходном положении надо лечь лицом вниз и взяться за рукояти тренажера для обеспечения устойчивости.
Важный момент: колени при этом должны немного выходить за края скамьи, а не упираться в нее. Ягодицы и пресс следует держать напряженными.
Выполнение упражнения
На вдохе необходимо быстро поднять валик ногами так, чтобы голени составляли с бедрами прямой угол (как минимум). В этом пиковом положении задержаться на пару секунд и напрячь бицепс бедра для увеличения работы мышц.
На выдохе плавно не до конца распрямить ноги и сразу начать следующее повторение.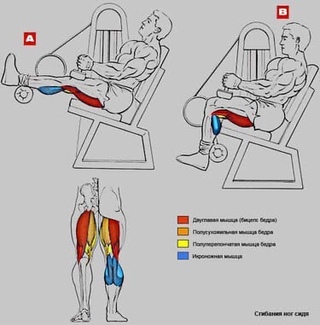
- Следует начинать с 12-15 повторений по 2-4 подхода.
- Корпус тела должен быть плотно прижат к скамье для хорошей опоры во время выполнения упражнения.
- Предпочтение лучше отдавать тренажерам с изогнутой скамьей, которая обеспечивает большее растяжение тренируемых мышц.
- Можно попробовать выполнять упражнение отдельно для каждой ноги по очереди, чтобы лучше концентрироваться на проработке мышц.
Не стоит приподнимать или двигать таз, а также использовать другие мышцы (например, спины или рук, опирающихся на рукояти), так как это снижает эффективность упражнения для бицепса бедра, увеличивает риск травмы других мышц. При этом, если скамья прямая, бедра могут немного приподниматься в естественном движении, поэтому не нужно просить кого-то прижимать их в процессе выполнения упражнения.
- Не стоит выполнять упражнения рывками, используя силу инерции, так как от этого снижается нагрузка на бицепс бедра и увеличивается вероятность травмы других мышц и суставов.

- Опускать ноги в конце упражнения лучше плавно и не до конца – это сохраняет напряжение и эластичность мышц, защищает коленные суставы от повреждения.
- Ноги и стопы должны быть параллельны друг другу, однако, иногда можно немного менять их расположение. Чтобы увеличить нагрузку на внешнюю часть задней поверхности бедра, следует держать вместе пятки, на внутреннюю часть – носки.
- Необходимо правильно индивидуально подобрать вес. Не стоит выбирать слишком большую нагрузку, особенно если имеются проблемы со спиной.
- После выполнения каждого подхода нужно растягивать мышцы.
- После окончания упражнения уставшими должны быть мышцы именно бедра, а не спины. Если чувствуется обратное, значит упражнение выполняется неправильно.
Подводим итоги
В этом посте вы узнаете, как правильно делать сгибание ног на тренажере. Это упражнение весьма популярно, как у мужчин, так и у женщин. Почему? Благодаря тому, что задействуется огромное количество мышц, эффективно сжигается жир и прорисовывается рельеф.
Как выполняется сгибание ног
На самом деле, это не одно упражнение, а просто механика — сгибание ноги в колене. Имеет большое значение, каким образом этом происходит. Сгибание ног, сидя на тренажере, возможно в спортивном зале, на специально оборудованном тренажере. При этом вы прокачаете и заднюю часть бедра.
Сгибание ног стоя, как следует из названия, выполняется в стоячем положении. позволит вам прокачать и подколенные мышцы и, конечно же, мышцы бедра.
Сгибание ног на тренажере нужно выполнять после базовых упражнений. Не имеет значения, как давно вы тренируетесь, эти упражнения хороши как для новичков, так и для мастеров. Сколько раз нужно делать сгибание? Выполняйте упражнения по три-четыре подхода от десяти до пятнадцати раз.
Выполняем сгибание сидя
Сядьте на тренажер, ноги зафиксируйте валиком. Колени должны вылезать за край сидения, позволяя максимально распрямляться коленям.
Приступая к упражнению, ваши колени должны быть либо совершенно выпрямленными, либо немного согнутыми. Сделайте вдох, задержите дыхание и согните ноги. Доведя угол в колене до девяносто градусов, остановитесь и максимально напрягите ноги. Именно в этот момент вы работаете над мышцами. После вернитесь в исходное положение — так правильно выполняется сгибание ног сидя в тренажере.
Сделайте вдох, задержите дыхание и согните ноги. Доведя угол в колене до девяносто градусов, остановитесь и максимально напрягите ноги. Именно в этот момент вы работаете над мышцами. После вернитесь в исходное положение — так правильно выполняется сгибание ног сидя в тренажере.
Сгибание стоя
Это упражнение позволит вам проработать все мышцы бедра и икроножную мышцу. Возьмитесь за рукоятку тренажера руками. Ногу, которой будете выполнять упражнение, нужно прижать к опоре — верхнему валику. Голень подставьте под нижний валик. Выпрямляйте ногу и заводите щиколотку под нижний валик. Делая вдох, сгибайте ногу в колене. Выполнив сгибание, выдыхайте.
Можно увеличить нагрузку на мышцы во время выполнения этого упражнения — нужно всего лишь сгибать стопу при сгибании голени. Если же вам дается выполнение тяжело, то держите стопу выпрямленной — это снизит нагрузку на икроножную мышцу.
Сгибание ног на тренажере лежа
Еще одно полезное упражнение, которое позволит проработать подколенные мышцы, мышцы бедра и в меньшей степени — икроножную мышцу.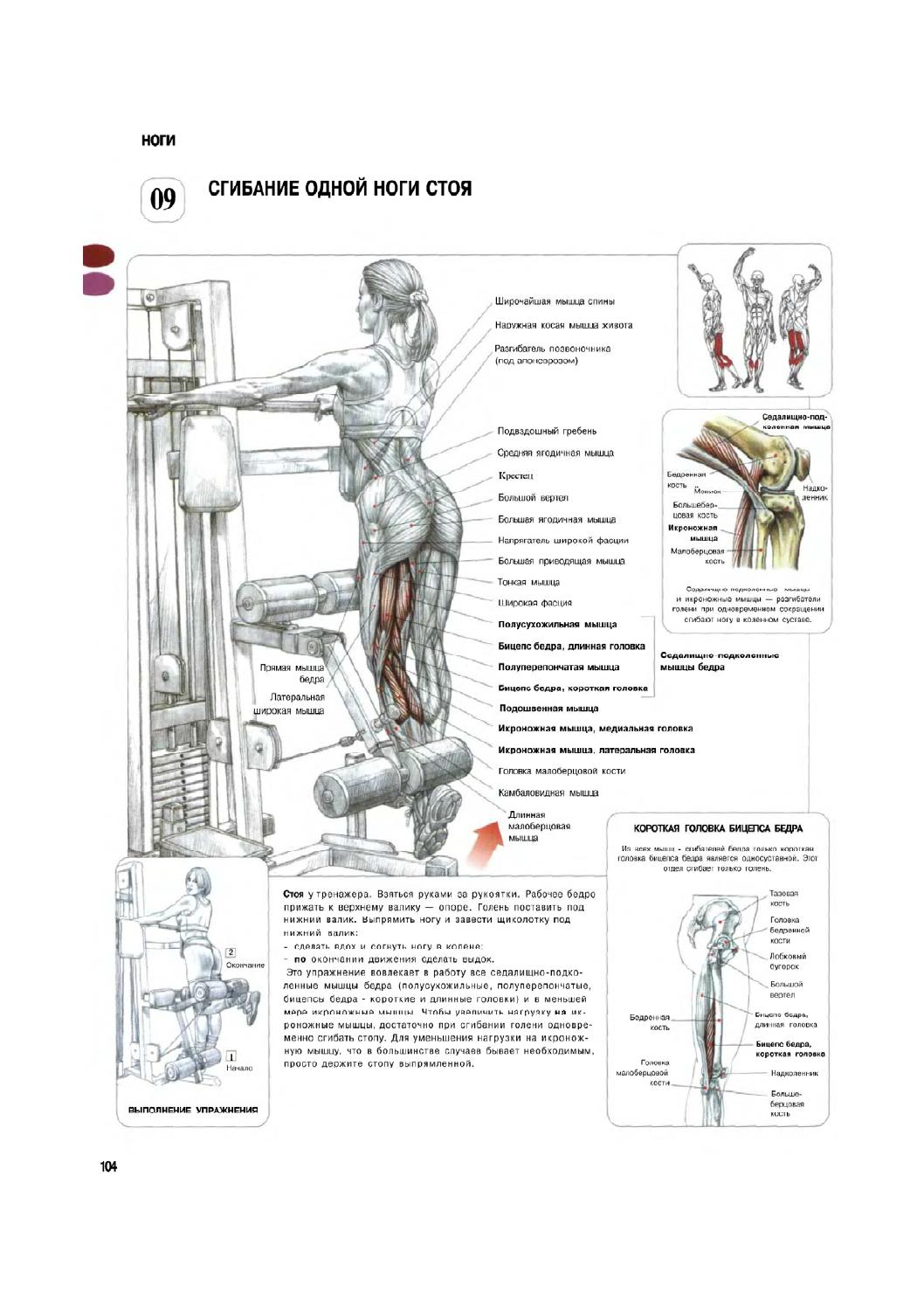 Как выполнять: лягте на тренажер лицом вниз, возьмитесь руками за поручни. Ноги нужно распрямить и положить их под валики. Выполняем — делайте вдох и сгибайте оба колена. Заводите валик, стараясь коснуться ногами ягодиц. Сделайте задержку. Медленно возвращайте ноги в исходное положение. Делайте выдох.
Как выполнять: лягте на тренажер лицом вниз, возьмитесь руками за поручни. Ноги нужно распрямить и положить их под валики. Выполняем — делайте вдох и сгибайте оба колена. Заводите валик, стараясь коснуться ногами ягодиц. Сделайте задержку. Медленно возвращайте ноги в исходное положение. Делайте выдох.
Тренинг для тех, кто работает на массу
Если ваша цель — наработка мышечной массы, то есть особые нюансы как в выполнении упражнений, так и в питании. Касательно работы в зале вам следует делать меньше подходов с максимально возможным весом.
Правильно подберите питание. Вам необходимо съедать в день белка из расчета два грамма на один килограмм идеального веса и употреблять правильные углеводы — каши, овощи. Конечно же, речь о сладком не идет, иначе вы наберете вместе с мышечной массой жировую. Во время тренировок пейте гейнеры.
Если вы хотите похудеть
Занимаясь для похудения, нужно в первую очередь помнить о питании.
Даже эффективный тренинг окажется бесполезным перед страстью к вкусненькому. Основой вашего питания должен стать белок (куриное мясо, творог, яйцо, рыба всех сортов), углеводы в виде зеленых овощей. Каши полезны, но если вы хотите потерять ненужные жировые отложения, то пока от них откажитесь.
Основой вашего питания должен стать белок (куриное мясо, творог, яйцо, рыба всех сортов), углеводы в виде зеленых овощей. Каши полезны, но если вы хотите потерять ненужные жировые отложения, то пока от них откажитесь.
Что же касается тренировок, то имеет смысл выполнять упражнения с небольшим весом, однако делая большее количество повторений. Если средний показатель — 10 раз за подход, то делайте не менее 20.
Владыка всея сайта и фитнес-тренер | подробнее >>
Род. 1984 г Тренируется с 1999 г. Тренирует с 2007.. КМС по пауэрлифингу. Чемпион России и Юга России по версии AWPC. Чемпион Краснодарского края по версии IPF. 1 разряд по тяжёлой атлетике. 2-х кратный призёр чемпионата Краснодарского края по т/а. Автор более 700 статей по фитнесу и любительскому атлетизму. Автор и соавтор 5 книг.
Место в : вне конкурса ()
Дата: 2012-05-29 Просмотры: 1 256 804 Оценка: 4.8
За что статьям даются медали:
Дополнительные —
Сложность выполнения — лёгкая
Сгибание ног в тренажёре лёжа — видео
Вес и количество повторений для новичков
Для мужчин: 10 — 15 повторений.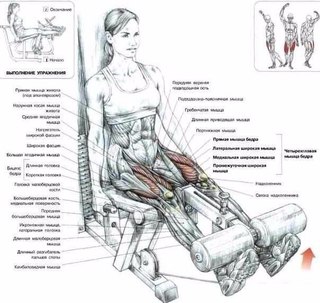 2 — 3 подхода.
2 — 3 подхода.Для женщин: 10 — 15 повторений. 2 — 3 подхода.
Нагрузка по группам мышц
Нагрузка указана по 10-ти бальной шкале (общая нагрузка суммируется)Описание упражнения
Рекомендую в финальной фазе немного поднимать колени вверх. От этого выиграет и задняя поверхность бедра и ягодицы, так как это заставит их сильнее сокращаться. Опускать ноги лучше до конца.Основные фишки
1. Ложиться надо так, чтобы колени не упирались в подставку, а немного свешивались. Так более безопасно для них. 2. Валик устанавливаем так, чтобы он упирался в щиколотку, ближе к пятке. Чем длиннее рычаг, тем лучше. 3. Сгибать ноги желательно до конца. Чтобы валик касался ягодиц. Разгибать же можно как до конца, так и оставляя небольшой угол в колене. 4. Если в верхней фазе движения вы будете немного колени отрывать от спинки, то немного повысите эффективность упражнения. Так будут включаться в работу ягодицы, и будет сильнее сокращаться задняя поверхность бедра. 5. Это упражнение не является базовым. Поэтому делайте его после становых тяг, наклонов или гиперэкстензии. Чтобы «добить» заднюю поверхность бедра. 6. Как вариант, можно делать это упражнение одной ногой по очереди. Так вы сможете лучше сосредоточится на одной ноге.
5. Это упражнение не является базовым. Поэтому делайте его после становых тяг, наклонов или гиперэкстензии. Чтобы «добить» заднюю поверхность бедра. 6. Как вариант, можно делать это упражнение одной ногой по очереди. Так вы сможете лучше сосредоточится на одной ноге.Сгибание ног в тренажёре – это изолированное упражнение на бицепс бедра , которое хорошо работает в связке с такими упражнениями, как тяга гантелей на прямых ногах или разгибание ног в тренажёре. А также сгибания ног могут заменить базовые упражнения на заднюю поверхность бедра, если их нельзя выполнять, например, из-за лёгких травм спины. Если есть травмы, то для начала необходимо получить разрешение врача на занятия. Если же всё в порядке и заниматься спортом можно, то нужно потратить немного времени на изучение техники выполнения этого упражнения.
Виды упражнений и техника выполнения
Основные виды упражнений на сгибание
В спортивных клубах чаще всего можно встретить три основных разновидности тренажёров. Различаются они по исходному положению тела:
В некоторых тренажёрах нет регулировщика веса и вместо этого нужно самостоятельно навешивать блины на штыри, расположенные на валике или сбоку от тренажёра.
Техника выполнения
Перед выполнением упражнения необходимо тщательно изучить технику его выполнения. В независимости от положения тела (лёжа, сидя или стоя) рекомендации к правильному выполнению упражнения будут следующими:
Как разнообразить тренировку: чем заменить сгибания, пример тренировки
Альтернативные варианты
Пример тренировки
Эта тренировка включает в себя упражнение на сгибание ног.
Сгибания ног можно включить и в домашние тренировки, и в тренировки в спортивном зале. Это упражнение подходит как для начинающих, так и для опытных спортсменов. Новичкам следует сначала внимательно изучить технику выполнения и не гнаться за большими весами, так как не величина веса, а правильное выполнение упражнение быстрее приведёт к результату.
По статистике, упражнение сгибание ног лежа пользуется наибольшей популярностью у прекрасной половины человечества. И это вполне объяснимо: данное упражнение позволяет привести в форму, а также продемонстрировать свои стройные, подтянутые ножки.
Сгибание ног в тренажере лежа как раз и акцентировано на разработке мускулатуры задней стороны ног и ягодиц, а также придании рельефности и объемности бицепсам бедер.
Оно относится не к базовым, а к изолирующим упражнениям и является очень важным для обеспечения баланса между мускулатурой передней и задней части ноги, придавая ногам эстетичный вид.
Сгибание ног лежа для девушек как раз и ценно тем, что формирует красивые ноги, не слишком наращивая мышечную массу.
Сгибание ног лежа мышцы задействует на задней поверхности бедра, а если точнее, то бицепсы бедра, полусухожильные и полуперепончатые мышцы и частично еще икроножные мышцы. Соответствующий тренажер можно найти в любом тренажерном зале.
Само по себе сгибание ног в тренажере в лежачем положении – упражнение довольно простое по технике выполнения, но, по мнению мужчин, является достаточно сложной тренировкой.
Прежде всего, с помощью нижнего валика необходимо выставить положение тренажера, соответствующее вашему росту.
Упражнение выполняется в положении лежа на животе, с параллельно расположенными ногами, заведенными под валик, который упирается в заднюю часть лодыжек.
Талия должна находиться на изломе скамьи, руки – прочно держаться за поручни, а колени чуть-чуть выходить за пределы скамьи во избежание травматизма. При этом весь корпус прижат к скамье. Такова исходная позиция, в которой выполняется сгибание ног лежа на животе. Далее:
- На глубоком вдохе, задержав дыхание и не отрываясь от скамейки, валик, напрягая мышцы, подтягивают к ягодицам.
- Выдох и задержка в самой верхней позиции на несколько секунд.
- Вдох и очень медленное возвращение в исходное положение.
Казалось бы, сгибания ног в положении лежа – очень простое упражнение. У упражнения сгибание ног лежа техника действительно не слишком сложная, однако тем важнее ее точное соблюдение:
- крепко держаться за ручки;
- плотно прижимать таз к скамейке;
- заводить ноги так высоко, чтобы они почти соприкасались с ягодицами;
- чтобы сохранять нагрузку, не распрямлять колени, находящиеся в самой низкой точке, полностью;
- движение вниз должно происходить медленнее, чем вверх.

Это упражнение лучше делать после основных базовых упражнений для ног, нацеленных на наращивание мышечной массы, в то время как оно является формирующим.
техника выполнения, особенности и замечания.
Сгибание ног на тренажере для девушекСгибание ног на тренажере для девушек: лежа на животе
Сгибание ног на тренажере для девушек: сидя
Тренажеров для сгибания ног всего 3 вида — для сгибания ног лежа, для сгибания ног лежа поочередно, и для сгибания ног сидя. Все они направлены на изолированную проработку мышц задней поверхности бедра и активно применяются в женском тренинге. Уровень технической сложности этого движения невысок, между тем в современном функциональном фитнесе его часто называют травмоопасным. Сгибание ног в тренажере — одно из самых распространенных упражнений.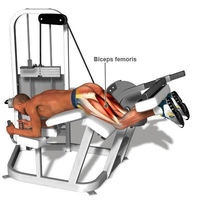
Цель упражнения
В фитнесе это обычно гипертрофия задней поверхности бедер, особенно для тех людей, у которых нет возможности выполнять вертикальные жимы ногами, махи гирей, тяги на прямых или чуть согнутых ногах. В силовом тренинге упражнение используется как специально-вспомогательное движение при недостаточно развитом сгибателе бедра. Есть мнение, что упражнение может активно применяться в тренинге, направленном на реабилитацию связок и суставов, но оно оспаривается.
Обычно упражнение в фитнесе используют в конце силовой тренировки, в бодибилдинге встречается и вариант с «предварительным утомлением» группы мышц при помощи этого движения в начале силовой тренировки.
Работающие мышцы
Бицепс бедра и мышцы голени
Техника
В тренажере на обе ноги
Нужно отрегулировать подушку-фиксатор так, чтобы не было избыточного разгибания в коленном суставе. В исходном положении ноги чуть подсогнуты в коленях, что обеспечивает безопасность движения. Вес выставляется относительно небольшой, с предельными весами односуставные изолирующие движения лучше не выполнять.
Вес выставляется относительно небольшой, с предельными весами односуставные изолирующие движения лучше не выполнять.
Далее необходимо лечь лицом вниз на скамью тренажера, разместить пятки под подушкой, подтянуть живот, и за счет одновременного сгибания обеих ног активно привести подушку тренажера к ягодицам, затем медленно выпрямить ноги в исходное положение.
В этом тренажере можно осуществлять и подъем одной ногой, тогда следует особое внимание уделить тому, чтобы подушка была стабильна, когда вы принимаете упор с одной стороны и нога двигалась без перекосов в коленном и тазобедренном суставах.
В тренажере с одной ноги
В этом случае машина сконструирована так, что имеются 2 отдельные подушки для каждой ноги своя. И вес навешивается непосредственно на весовые стеки, а не через блочную систему. Этот тренажер требует внимательного подбора весов. Требуется также «освободить» колени, и не упираться ими в скамью, сгибая ноги. В остальном техника и механика движения полностью повторяют предыдущий вариант.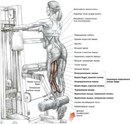
Сидя
В тренажере сидя необходимо обеспечить упор так, чтобы голени были перпендикулярны полу в исходном положении. Стоит отказаться и от избыточного «силового» сгибания, когда колени становятся остырыми в крайней точке амплитуды
Упражнение может выполняться как в 8-12 повторениях, так и в большем количестве повторов, все зависти от цели тренирующегося и его приоритетов.
работающие мышцы и техника выполнения
Лягте на тренажёр лицом вниз, валики на голенях чуть ниже икр. Тяните носки на себя, тело и бедра плотно прижаты к скамье. На выдохе сгибайте ноги. В точке наибольшего напряжения задержитесь на секунду. На вдохе опустите ноги обратно в исходное положение.
Сгибание ног в тренажере лежа: видео
Правила выполнения упражнения
- Отрегулируйте тренажёр в соответствии со своим ростом.
Лягте на скамью лицом вниз, ноги положив под валики (чуть ниже икроножных мышц). Совет: Желательно заниматься на тренажёре, где скамья под углом, в таком случае лучше прорабатывается бицепс бедра.
- Тело плотно прижато к скамье. Ноги выпрямлены. Держитесь за боковые ручки тренажёра. Тяните носки на себя (или же можно использовать другое положение ног). Это исходное положение.
- На выдохе сгибайте ноги как можно сильнее. Бёдра при этом не отрываются от скамьи. В точке наибольшего напряжения задержитесь на секунду.
- На вдохе опустите ноги обратно в исходное положение. Повторите необходимое количество раз.
Предупреждение: Не берите чрезмерно большой вес. Вы не должны раскачивать или дёргать телом, иначе это может привести к травме нижней части спины и подколенного сухожилия.
Варианты: Это упражнение предполагает три положения стоп. Фактически, это три разных упражнения. Помимо этого, вы можете работать с гантелью между ног (попросите напарника правильно положить её). Такой вариант подходит для опытных. И, наконец, для лучшей изоляции, можно работать каждой ногой по очереди.
Такой вариант подходит для опытных. И, наконец, для лучшей изоляции, можно работать каждой ногой по очереди.
Альтернативные упражнения
10 упражнений, на которые вы зря тратите время, и 10 эффективных альтернатив
1. Использование тренажера для сгибания ног сидя
Почему? Кажется, что это должно помочь вам избавиться от жира на внешней и внутренней части бедер. Но увы, это не так. Упражнения, направленные только на одну часть тела, не помогут похудеть.
Чем заменить? Выпады вперед, назад и в стороны. Эти упражнения приведут в тонус и сделают более сильным все тело, в том числе и бедра. К тому же они более практичны и делают вас выносливее для повседневной жизни (в отличие от тренажера, по словам Грега Джастиса, владельца и основателя фитнес-центра AYC Health & Fitness в Канзас-Сити).
2. Разгибание ног на тренажере
Почему? Упражнение должно быть направлено на усиление ваших квадрицепсов. Но когда вы разгибаете ноги на тренажере в положении сидя, ваши колени получают слишком большую нагрузку.
Чем заменить? Приседания или выпады. Они работают сразу на квадрицепсы, подколенные сухожилия и ягодицы.
3. Упражнения на пресс
Почему? Просто потому, что они не настолько эффективны. Исследования подтвердили, что упражнения на пресс менее эффективны для стимуляции мышечных волокон, чем упражнения, направленные на стабилизацию позвоночника (планка).
Чем заменить? Планка (читайте «Держим планку: комплекс простых и эффективных упражнений для красивого тела»). Выполняя это упражнение, вы задействуете больше мышц кора, чем при качании пресса. Кроме того, делая планку, вы не сможете потянуть спину.
4. Наклоны в сторону с гимнастической палкой или повороты
Почему? Вам кажется, что таким образом вы укрепляете мышцы кора или сжигаете жир, но на самом деле нет. В реальности, когда вы работате с весом, расположенным у вас за шеей, и начинаете крутить и вращать штангой, сопротивление не такое уж и большое. Лучше тренироваться, превозмогая гравитацию.
В реальности, когда вы работате с весом, расположенным у вас за шеей, и начинаете крутить и вращать штангой, сопротивление не такое уж и большое. Лучше тренироваться, превозмогая гравитацию.
Чем заменить? Боковая планка в статике или динамике. Это как раз упражнение, направленное на сопротивление гравитации, которое эффективно прорабатывает ваши мышцы.
5. Сведение рук на блочном тренажере
Почему? Вы сидите, из-за этого ваша более сильная часть тела получает большую нагрузку, чем более слабая. Возникает дисбаланс.
Чем заменить? Отжимания. Во время отжиманий вы вынуждены полагаться и на руки, и на ноги, иначе упадете. Кроме того, отжимания включают плюсы планки и работают также на мышцы пресса.
6. Сгибание ног на тренажере из положения лежа
Почему? Когда вы выполняете упражнения лежа, ваш позвоночник не задействован, вы тренируете наименьшее количество мышц. К тому же такое упражнение оказывается бесполезным в реальной жизни: вряд ли кто-то сможет назвать хоть одну причину, по которой нам необходимо сгибать ноги из положения лежа.
К тому же такое упражнение оказывается бесполезным в реальной жизни: вряд ли кто-то сможет назвать хоть одну причину, по которой нам необходимо сгибать ноги из положения лежа.
Чем заменить? Подъем ноги назад из положения стоя. Это упражнение активизирует заднюю часть вашего тела, которая включает поясницу, ягодицы, мышцы бедра и икры, одновременно заставляя работать и мышцы туловища для сохранения баланса. Самое главное, что это упражнение развивает необходимые в повседневной жизни гибкость и умение балансировать.
7. Жим лежа
Почему? Из-за того что вы лежите, упражнение не позволяет нагрузке равномерно распределяться по всему телу. Передняя часть ваших ног оказывается под большим напряжением, которое выливается в огромную нагрузку на колени.
Чем заменить? Приседания с гантелей или штангой небольшого веса. Упражнение направлено на большую группу мышц ног, в том числе ягодицы, мышцы бедер и квадрицепс.
8. Разгибание руки с гантелей
Почему? У вас не получится поднимать достаточно тяжелый вес и при этом сохранять правильную позицию рук. Обычно плечо начинает провисать, эффективность упражнения теряется.
Чем заменить? «Череподробилка», или французский жим лежа на трицепс. Упражнение позволит вам прокачать мышцы трицепса без потери формы рук. Положение «череподробилка» позволяет вашим локтям оставаться там, где следует, и получить таким образом необходимую нагрузку.
9. Подъем на носки, стоя на тренажере
Почему? Тренажер опускает весь вес на ваши плечи, что дает нагрузку сначала на вашу спину, а потом уже на икроножные мышцы.
Чем заменить? Спринт. Это лучше, чем обычный бег, для развития ваших икр. Спринт активирует больше быстросокращающихся мышечных волокон, которые отвечают за рост и развитие мышц.
Если спринт не для вас, попробуйте подъем на носки, используя вес тела. Если и это покажется вам слишком легким, то попробуйте делать это упражнение на одной ноге.
Если и это покажется вам слишком легким, то попробуйте делать это упражнение на одной ноге.
10. Жим ногами
Почему? Люди склонны добавлять слишком большой вес, что потенциально может привести к травме. К тому же это упражнение не такое эффективное, если сравнивать его с аналогами.
Чем заменить? Приседания с гантелями. Вы по-прежнему качаете мышцы ног, но при выполнении этого упражнения вы более стабильны и меньше рискуете получить травму.
Источник: msn.com
Как эффективно и безопасно сбросить вес:
Мышечные модели динамики движения вперед для изучения нормальной и патологической походки | Журнал NeuroEngineering и реабилитации
Yamaguchi GT: Динамическое моделирование Musculoskeletal движения. Нью-Йорк, Спрингер; 2001.
Google ученый
 org/ScholarlyArticle»> 2.
org/ScholarlyArticle»> 2.Zajac FE: Мышцы и сухожилия: свойства, модели, масштабирование и применение в биомеханике и моторном управлении. Crit Rev Biomed Eng 1989, 17: 359-411.
CAS PubMed Google ученый
Zajac FE, Neptune RR, Kautz SA: Биомеханика и координация мышц при ходьбе человека. Часть I: введение в концепции, передачу мощности, динамику и моделирование. Походка 2002, 16: 215-232. 10.1016 / S0966-6362 (02) 00068-1
Артикул PubMed Google ученый
Zajac FE, Neptune RR, Kautz SA: Биомеханика и координация мышц при ходьбе человека: часть II: уроки динамического моделирования и клинические последствия. Походка 2003, 17: 1-17. 10.1016 / S0966-6362 (02) 00069-3
10.1016 / S0966-6362 (02) 00069-3
Артикул PubMed Google ученый
Neptune RR: Компьютерное моделирование и симуляция движений человека. Приложения в спорте и реабилитации. Phys Med Rehabil Clin N Am 2000, 11: 417-34, viii.
CAS PubMed Google ученый
Hatze H: Фундаментальные вопросы, последние достижения и будущие направления миодинамики. J Electromyogr Kinesiol 2002, 12: 447-454. 10.1016 / S1050-6411 (02) 00038-X
CAS Статья PubMed Google ученый
Pandy MG: Компьютерное моделирование и симуляция движений человека. Annu Rev Biomed Eng 2001, 3: 245-273. 10.1146 / annurev.bioeng.3.1.245
Annu Rev Biomed Eng 2001, 3: 245-273. 10.1146 / annurev.bioeng.3.1.245
CAS Статья PubMed Google ученый
Shelburne KB, Pandy MG, Anderson FC, Torry MR: Картина силы передней крестообразной связки при нормальной ходьбе. Дж. Биомех 2004, 37: 797-805. 10.1016 / j.jbiomech.2003.10.010
Статья PubMed Google ученый
Shelburne KB, Pandy MG, Torry MR: Сравнение поперечных сил и нагрузки на связки в здоровом колене и колене с дефицитом ACL во время ходьбы. Дж. Биомех 2004, 37: 313-319. 10.1016 / j.jbiomech.2003.07.001
Статья PubMed Google ученый
 org/ScholarlyArticle»> 10.
org/ScholarlyArticle»> 10.Piazza SJ, Delp SL: Трехмерное динамическое моделирование движения полного протезирования коленного сустава во время выполнения задачи повышения. J Biomech Eng 2001, 123: 599-606. 10.1115 / 1.1406950
CAS Статья PubMed Google ученый
Caruntu DI, Hefzy MS: Трехмерное анатомическое динамическое моделирование человеческого колена с включением тибио-бедренного и пателло-бедренного суставов. J Biomech Eng 2004, 126: 44-53. 10.1115 / 1.1644565
Артикул PubMed Google ученый
Halloran JP, Petrella AJ, Rullkoetter PJ: Явное конечно-элементное моделирование механики полного эндопротезирования коленного сустава. Дж. Биомех 2005, 38: 323-331.10.1016 / j.jbiomech.2004.02.046
Статья PubMed Google ученый
Bei Y, Fregly BJ: Многотельное динамическое моделирование механики контакта с коленом. Med Eng Phys 2004, 26: 777-789. 10.1016 / j.medengphy.2004.07.004
PubMed Central Статья PubMed Google ученый
Фрегли Б.Дж., Сойер В.Г., Харман М.К., Бэнкс С.А.: Расчетное прогнозирование износа при полной замене коленного сустава на основе кинематики in vivo. Дж. Биомех 2005, 38: 305-314. 10.1016 / j.jbiomech.2004.02.013
Статья PubMed Google ученый
Neptune RR, Sasaki K: Создание силы подошвенного сгибателя голеностопного сустава является важным фактором, определяющим предпочтительную скорость перехода от ходьбы к бегу. J Exp Biol 2005, 208: 799-808. 10.1242 / jeb.01435
Артикул PubMed Google ученый
Sasaki K, Neptune RR: Механическая работа мышц и использование упругой энергии во время ходьбы и бега с близкой к предпочтительной переходной скорости походки. Gait Posture 2005, в печати.
Google ученый
Сасаки К., Нептун RR: Различия в мышечной функции при ходьбе и беге с одинаковой скоростью. J Biomech 2005, в печати.
Google ученый
Hreljac A: Предпочтительные и энергетически оптимальные переходные скорости походки при передвижении человека. Med Sci Sports Exerc 1993, 25: 1158-1162.
CAS Статья PubMed Google ученый
Hreljac A: Детерминанты скорости перехода походки при движении человека: кинематические факторы. Дж. Биомех 1995, 28: 669-677. 10.1016 / 0021-9290 (94) 00120-S
CAS Статья PubMed Google ученый
Прилуцкий Б.И., Грегор Р.Дж.: Действия мышц, связанные с махами и опорой, по-разному запускают переходы человека-бега-бега и бега-ходьбы. J Exp Biol 2001, 204: 2277-2287.
CAS PubMed Google ученый
Райт И.К., Нептун Р.Р., ван ден Богерт А.Дж., Нигг Б.М.: Влияние податливости и гибкости голеностопного сустава на растяжение связок. Med Sci Sports Exerc 2000, 32: 260-265.10.1097 / 00005768-200002000-00002
CAS Статья PubMed Google ученый
Райт И.К., Нептун Р.Р., ван ден Богерт А.Дж., Нигг Б.М.: Влияние положения стопы на растяжение связок голеностопного сустава. Дж. Биомех 2000, 33: 513-519. 10.1016 / S0021-9290 (99) 00218-3
CAS Статья PubMed Google ученый
Маклин С.Г., Хуанг X, Су А., Ван ден Богерт А.Дж.: Биомеханика в сагиттальной плоскости не может повредить ПКС во время обрезания в сторону. Clin Biomech 2004, 19: 828-838. 10.1016 / j.clinbiomech.2004.06.006
Артикул Google ученый
Андерсон Ф.К., Пэнди MG: Динамическая оптимизация ходьбы человека. J Biomech Eng 2001, 123: 381-390. 10.1115 / 1.1392310
CAS Статья PubMed Google ученый
Дэви Д.Т., Оду М.Л .: Метод динамической оптимизации для прогнозирования мышечных сил в фазе качания походки. Дж. Биомеханика 1987, 20: 187-201. 10.1016 / 0021-9290 (87) -1
CAS Статья Google ученый
Yamaguchi GT, Zajac FE: Восстановление естественной походки без посторонней помощи при параплегии с помощью функциональной нервно-мышечной стимуляции: исследование с компьютерным моделированием. IEEE Trans Biomed Eng 1990, 37: 886-902. 10.1109 / 10.58599
CAS Статья PubMed Google ученый
Chow CK, Jacobson DH: Исследования передвижения человека с помощью оптимального программирования. Math Biosci 1971, 10: 239-306. 10.1016 / 0025-5564 (71) -9
Артикул Google ученый
Герритсен KG, ван ден Богерт AJ, Hulliger М, Цернике РФ: Внутренние свойства мышцы облегчают управление двигательным — исследование компьютерного моделирования. Управление двигателем 1998, 2: 206-220.
CAS PubMed Google ученый
Saunders JBDCM, Inman VT, Eberhart HD: Основные детерминанты нормальной и патологической походки. JBJS 1953, 35-A: 543-558.
CAS Google ученый
Гард С.А., Чайлдресс DS: Влияние сгибания колена в фазе опоры на вертикальное смещение туловища при нормальной ходьбе. Arch Phys Med Rehabil 1999, 80: 26-32. 10.1016 / S0003-9993 (99)
-9
CAS Статья PubMed Google ученый
Ortega JD, Farley CT: Минимизация вертикального движения центра масс увеличивает метаболические затраты при ходьбе. J Appl Physiol 2005.
Google ученый
Пэнди М.Г., Берме N: Количественная оценка детерминант походки во время одиночной стойки с помощью трехмерной модели — Часть 1.Походка нормальная. Дж. Биомех 1989, 22: 717-724. 10.1016 / 0021-9290 (89)
CAS Статья PubMed Google ученый
Neptune RR, Zajac FE, Kautz SA: Требования к механической работе мышц во время нормальной ходьбы: энергетические затраты на поднятие центра масс тела значительны. Дж. Биомех 2004, 37: 817-825. 10.1016 / j.jbiomech.2003.11.001
CAS Статья PubMed Google ученый
Mochon S, McMahon TA: Баллистическая ходьба. Дж. Биомеханика 1980, 13: 49-57. 10.1016 / 0021-9290 (80)
-X
CAS Статья Google ученый
Александр РМ: Энергосберегающие механизмы при ходьбе и беге. J Exp Biol 1991, 160: 55-69.
CAS PubMed Google ученый
Neptune RR, Kautz SA, Zajac FE: Вклад отдельных подошвенных сгибателей голеностопного сустава в поддержку, продвижение вперед и начало движения во время ходьбы. Дж. Биомех 2001, 34: 1387-1398. 10.1016 / S0021-9290 (01) 00105-1
CAS Статья PubMed Google ученый
Андерсон Ф.К., Пэнди М.Г.: Вклад отдельных мышц в поддержку нормальной ходьбы. Походка 2003, 17: 159-169.10.1016 / S0966-6362 (02) 00073-5
Артикул PubMed Google ученый
Neptune RR, Zajac FE, Kautz SA: Мышечная сила перераспределяет сегментарную силу для движения тела во время ходьбы. Походка 2004, 19: 194-205. 10.1016 / S0966-6362 (03) 00062-6
CAS Статья PubMed Google ученый
Райли П.О., Делла Кроче У., Керриган, округ Колумбия: Пропульсивная адаптация к изменению скорости ходьбы. Дж. Биомех 2001, 34: 197-202. 10.1016 / S0021-9290 (00) 00174-3
CAS Статья PubMed Google ученый
Neptune RR, Kautz SA, Zajac FE: Комментарии к «Движущей адаптации к изменению скорости походки». Дж. Биомех 2001, 34: 1667-1670. 10.1016 / S0021-9290 (01) 00093-8
CAS Статья PubMed Google ученый
Piazza SJ, Delp SL: Влияние мышц на сгибание в коленях во время фазы качания походки. Дж. Биомех 1996, 29: 723-733. 10.1016 / 0021-9290 (95) 00144-1
CAS Статья PubMed Google ученый
Anderson FC, Goldberg SR, Pandy MG, Delp SL: Вклад мышечных сил и кинематики отталкивания пальцев в максимальное сгибание колена во время фазы качания при нормальной походке: анализ индуцированного положения. Дж. Биомех 2004, 37: 731-737. 10.1016 / j.jbiomech.2003.09.018
Статья PubMed Google ученый
Goldberg SR, Anderson FC, Pandy MG, Delp SL: Мышцы, которые влияют на скорость сгибания колена при двойной опоре: последствия для походки с скованным коленом. Дж. Биомех 2004, 37: 1189-1196. 10.1016 / j.jbiomech.2003.12.005
Статья PubMed Google ученый
Райли П.О., Керриган, округ Колумбия: Кинетика походки с жесткими ногами: анализ индуцированного ускорения. IEEE Trans Rehabil Eng 1999, 7: 420-426. 10.1109 / 86.808945
CAS Статья PubMed Google ученый
Райли П.О., Керриган, округ Колумбия: Крутящее действие двусуставных мышц в период качания при походке с неподвижными ногами: анализ прямой динамической модели. Дж. Биомех 1998, 31: 835-840.10.1016 / S0021-9290 (98) 00107-9
CAS Статья PubMed Google ученый
Goldberg SR, Ounpuu S, Delp SL: Важность начальных условий в фазе качания при походке с жесткими коленями. Дж. Биомех 2003, 36: 1111-1116. 10.1016 / S0021-9290 (03) 00106-4
Артикул PubMed Google ученый
Higginson JS, Zajac FE, Neptune RR, Kautz SA, Delp SL: Вклад мышц в поддержку во время ходьбы у человека с постинсультным гемипарезом. J Biomech 2005, в печати.
Google ученый
Siegel KL, Kepple TM, Stanhope SJ: Использование индуцированного ускорения для понимания стабильности колена во время походки людей с мышечной слабостью. Gait Posture 2005, в печати.
Google ученый
Арнольд А.С., Андерсон Ф.К., Панди М.Г., Делп С.Л.: Мышечный вклад в разгибание бедра и колена во время фазы опоры на одну конечность при нормальной походке: основа для исследования причин походки приседания. Дж. Биомех 2005, 38: 2181-2189. 10.1016 / j.jbiomech.2004.09.036
Статья PubMed Google ученый
Thelen DG: Регулировка параметров модели механики мышц для имитации динамических сокращений у пожилых людей. J Biomech Eng 2003, 125: 70-77. 10.1115 / 1.1531112
Артикул PubMed Google ученый
Lloyd DG, безье TF: EMG привод костно-мышечная модель для оценки мышечных сил и моментов коленного сустава в естественных условиях. Дж. Биомех 2003, 36: 765-776. 10.1016 / S0021-9290 (03) 00010-1
Артикул PubMed Google ученый
Манал К., Бьюкенен TS: Однопараметрическая модель нейронной активации к мышечной активации: оценка изометрических суставных моментов по электромиограммам. Дж. Биомех 2003, 36: 1197-1202.10.1016 / S0021-9290 (03) 00152-0
Артикул PubMed Google ученый
Гарнер Б.А., Пэнди М.Г.: Оценка свойств сухожилий в верхней конечности человека. Энн Биомед Энг 2003, 31: 207-220. 10.1114 / 1.1540105
Артикул PubMed Google ученый
Asakawa DS, Blemker SS, Rab GT, Bagley A, Delp SL: Трехмерная геометрия мышцы-сухожилия после пересадки сухожилия прямой мышцы бедра. J Bone Joint Surg Am 2004, 86-A: 348-354.
PubMed Google ученый
Блемкер С.С., Пинский П.М., Delp SL: Трехмерная модель мышцы выявляет причины неравномерного деформирования двуглавой мышцы плеча. Дж. Биомех 2005, 38: 657-665. 10.1016 / j.jbiomech.2004.04.009
Статья PubMed Google ученый
Blemker SS, Delp SL: Экскурсии волокон прямой мышцы бедра и широкой мышцы бедра, спрогнозированные с помощью трехмерных моделей мышц. J Biomech 2005.
Google ученый
Blemker SS, Delp SL: Трехмерное представление сложной архитектуры и геометрии мышц. Энн Биомед Энг 2005, 33: 661-673. 10.1007 / s10439-005-1433-7
Артикул PubMed Google ученый
Thelen DD, Riewald SA, Asakawa DS, Sanger TD, Delp SL: Аномальное соединение моментов колена и бедра во время максимальных нагрузок у лиц с церебральным параличом. Мышечный нерв 2003, 27: 486-493. 10.1002 / mus.10357
Артикул PubMed Google ученый
Riewald SA, Delp SL: Действие прямой мышцы бедра после переноса дистального сухожилия: генерирует ли оно момент сгибания колена? Dev Med Child Neurol 1997, 39: 99-105.
CAS Статья PubMed Google ученый
Lundberg A: Кинематика голеностопного сустава и стопы. Рентгеновская стереофотограмметрия in vivo. Acta Orthop Scand Suppl 1989, 233: 1-24.
CAS Статья PubMed Google ученый
Reinbolt JA, Schutte JF, Fregly BJ, Koh BI, Haftka RT, George AD, Mitchell KH: Определение кинематических моделей многосуставных суставов для конкретных пациентов с помощью двухуровневой оптимизации. Дж. Биомех 2005, 38: 621-626. 10.1016 / j.jbiomech.2004.03.031
Статья PubMed Google ученый
Chen G: Вклады индуцированного ускорения в динамику движения физически не определены. Gait Posture 2005, в печати.
Google ученый
Оптимальная координация горизонтальных и вертикальных прыжковых движений с максимальным усилием — компьютерное моделирование | BioMedical Engineering OnLine
Целью этого исследования было изучить стратегию координации горизонтального прыжкового движения с максимальным усилием в сравнении с вертикальным прыжком.Горизонтальный прыжок и вертикальный прыжок с противодвижением были произведены с использованием техники компьютерного моделирования и численной оптимизации (рисунок 2, таблица 2). Выяснилось, что движение тазобедренного сустава больше при горизонтальном прыжке, чем при вертикальном (рис. 2, 3). Это наблюдение согласуется с результатами, полученными в [12], а также в [18]. В вертикальных прыжковых движениях ориентация сегмента туловища должна быть почти прямой, а его угловой момент должен быть уменьшен почти до нуля в момент взлета.Это условие необходимо для того, чтобы человеческое тело подпрыгивало вертикально при прямой позе [33, 34]. Следовательно, при вертикальном прыжке допускается меньшая активность тазобедренного сустава, чем при горизонтальном.
Оптимальное время движения для горизонтального прыжка было больше, чем для вертикального (0,92 с и 0,65 с соответственно). Этот результат согласуется с выводом, представленным в [12], в котором наблюдалась большая продолжительность движения сегмента туловища при горизонтальном прыжке, чем при вертикальном.Эти времена движения были аналогичны тем, которые наблюдались в экспериментальном исследовании [12], что также предполагает, что имитационная модель и метод оптимизации, использованные в этом исследовании, отражают фундаментальную природу прыжковых движений человека.
Величина сгибания тазобедренного сустава во время контрдвижения была больше при горизонтальном прыжке, чем при вертикальном (рис. 3). Голеностопный сустав при горизонтальном прыжке принимал позу с тыльным сгибом раньше, чем при вертикальном.При сочетании этих двух условий все тело было наклонено вперед в горизонтальном прыжке больше, чем в вертикальном (рис. 2). Этот результат полностью согласуется с экспериментальным наблюдением, приведенным в [12]. Это разумно, учитывая, что требуется создать импульс как вперед, так и вверх к моменту взлета при горизонтальном прыжке. С другой стороны, при вертикальном прыжке требуется генерировать только восходящий импульс. Чтобы прыгнуть вверх в прямой позе, при вертикальном прыжке центр масс тела должен удерживаться над ступнями.Движения тазобедренных, коленных и голеностопных суставов при вертикальном прыжке согласовывались, чтобы соответствовать этому требованию (Рисунок 2).
При изучении профилей активации мышц (рис. 4) и развития силы (рис. 5) можно заметить, что мышцы-сгибатели ноги задействованы для создания более сильных сгибательных движений в суставах во время фазы противодействия при горизонтальных прыжках. Это явление было ярко выражено при действии m. iliopsoas, m. rectus femoris и m. tibialis anterior.Это было видно с момента начала движения. Это действие привело к перемещению центра масс тела вперед во время встречного движения. Такая конфигурация сегментов тела помогла увеличить горизонтальный импульс, передаваемый центру масс тела за счет противодействия. Продолжительность активации мышц-разгибателей тазобедренного сустава (ягодичных мышц и подколенных сухожилий) при горизонтальном прыжке была больше, чем при вертикальном. Это наблюдение согласуется с выводом о том, что тазобедренный сустав при горизонтальном прыжке использовался более активно, чем при вертикальном.Было падение активации мм. vasti во время фазы отталкивания, тогда как была сильная активация m. biceps femoris в фазе отталкивания в горизонтальном прыжке. Такая координация была необходима для сохранения позы с наклоном вперед во время фазы отталкивания.
Результаты работы отдельных мышц (таблица 3) в целом были одинаковыми при горизонтальном и вертикальном прыжках. М. подвздошно-поясничная мышца, внешние вращающие мышцы бедра и подколенные сухожилия были единственными исключениями, в которых наблюдалась разница более чем на 10 Дж в выходной мощности.Производительность работы м. iliopsoas была больше при горизонтальном прыжке, чем при вертикальном, потому что эта мышца была активирована на большем уровне во время контрдвижения при горизонтальном прыжке (Рисунки 4 и 5). С другой стороны, выходная мощность внешних вращающих мышц бедра и подколенных сухожилий была меньше при горизонтальном прыжке, чем при вертикальном. В частности, нагрузка на подколенные сухожилия была отрицательной при горизонтальных прыжках (-4,8 Дж, Таблица 3). Это говорит о том, что подколенные сухожилия испытали эксцентрическое действие, при котором эта мышца была растянута одновременно с приложением мышечной силы.Это связано с тем, что во время контрдвижения в направлении сгибания тазобедренного сустава был придан большой импульс туловищному сегменту, а подколенные сухожилия использовались для противодействия импульсу. Таким образом, выходное усилие этой мышцы было большим (Рисунок 5), а выходная мощность — отрицательным (Таблица 3). Подобный механизм, по-видимому, вызвал меньшую мощность работы внешних вращающих мышц бедра при горизонтальном прыжке (5,3 Дж), чем при вертикальном прыжке (19,3 Дж).
Интересно, что количество поступательной механической энергии, полученной центром масс тела во время прыжкового движения, было больше в горизонтальном прыжке (258.9 Дж), чем в вертикальном прыжке (180,3 Дж; 70% по сравнению с горизонтальным прыжком). Этот результат согласуется с тем, что было сообщено в [12], в которой увеличение поступательной энергии центра масс тела во время вертикального прыжка составляло целых 64% от увеличения поступательной энергии центра масс тела во время горизонтального прыжка. Прыгать. Поскольку разница в общей мощности мышечной работы была незначительной (общее значение составляло 357,3 Дж для горизонтального прыжка и 375,8 Дж для вертикального прыжка; разница составляла всего 5%), было предложено передать мышечную работу на механическая энергия центра масс тела более эффективна при горизонтальном прыжке, чем при вертикальном.Объяснение этого открытия заключается в различии передачи механической энергии во время встречного движения. В горизонтальных прыжках снижение потенциальной энергии при перемещении сегментов тела в нижнее положение было связано с увеличением кинетической энергии этих сегментов, движущихся в прямом направлении (рис. 2). Следовательно, во время противодействия в горизонтальных прыжках потери механической энергии были меньше. Однако в вертикальном прыжке весь нисходящий импульс, генерируемый во время противодействия, должен был нейтрализоваться мышечными усилиями, прежде чем тело начало двигаться вверх (Рисунок 2).2) в момент отталкивания от прыжков в обоих видах прыжков был невелик. Величина была менее 6% для горизонтальных прыжков и менее 3% для вертикальных прыжков. Другими словами, переводческие компоненты внесли гораздо больший вклад. Это может быть связано с тем, что значение момента инерции сегментов человеческого тела обычно невелико.
В этом исследовании результирующая скорость центра масс тела в момент отталкивания была больше при горизонтальном прыжке, чем при вертикальном (таблица 2).Этот результат кажется несовместимым с результатами, представленными в [18], в которых результирующая скорость центра масс тела в момент взлета была почти одинаковой при горизонтальном и вертикальном прыжках. Это различие можно объяснить наличием / отсутствием противодействия. Поскольку в прыжке, смоделированном в этом исследовании, использовалось противодействие, и поскольку при горизонтальном прыжке наблюдалась эффективная передача механической энергии, центр масс тела испытывал больший прирост механической энергии к моменту взлета.Предполагается, что этот механизм передачи энергии был менее очевиден в горизонтальном прыжковом движении, изученном в [18], поскольку это движение было прыжком на корточках, а не прыжком против движения.
Оптимальный угол проекции, полученный в этом исследовании, составлял 34 градуса, тогда как этот параметр, полученный в предыдущем экспериментальном исследовании, составлял 48 градусов [12]. Это несоответствие может быть объяснено различием в скелетно-мышечных свойствах модели и субъектов, т.е. субъекты, которые участвовали в работе [12] были «сильнее», чем модели, используемых в данном исследовании.Это очевидно при сравнении высоты вертикального прыжка, полученной в этом исследовании (38,5 см), с высотой вертикального прыжка испытуемых (41,0 см), хотя обе эти цифры находятся в пределах диапазона экспериментальных наблюдений, о которых сообщалось в многочисленных предыдущих исследованиях вертикальных прыжков. . Это различие кажется разумным, учитывая, что испытуемые были подготовленными спортсменами (австралийский футбол) в [12]. Как обсуждалось ранее, горизонтальное расстояние, пройденное при горизонтальном прыжке, рассчитывалось как
Xlanding = Xt.о. + VXt.o.⋅2⋅VYt.o.gMathType @ СПР @ 5 @ 5 @ + = feaafiart1ev1aaatCvAUfKttLearuWrP9MDH5MBPbIqV92AaeXatLxBI9gBaebbnrfifHhDYfgasaacH8akY = wiFfYdH8Gipec8Eeeu0xXdbba9frFj0 = OqFfea0dXdd9vqai = hGuQ8kuc9pgc9s8qqaq = dirpe0xb9q8qiLsFr0 = vr0 = vr0dc8meaabaqaciaacaGaaeqabaqabeGadaaakeaacqWGybawdaWgaaWcbaGaemiBaWMaemyyaeMaemOBa4MaemizaqMaemyAaKMaemOBa4Maem4zaCgabeaakiabg2da9iabdIfaynaaBaaaleaacqWG0baDcqGGUaGlcqWGVbWBcqGGUaGlaeqaaOGaey4kaSIaemOvay1aaSbaaSqaaiabdIfayjabdsha0jabc6caUiabd + gaVjabc6caUaqabaGccqGHflY1daWcaaqaaiabikdaYiabgwSixlabdAfawnaaBaaaleaacqWGzbqwcqWG0baDcqGGUaGlcqWGVbWBcqGGUaGlaeqaaaGcbaGaem4zaCgaaaaa @ 5502 @Анализируя правую часть этой формулы, можно вывести, что второй член максимизируется, когда угол проекции составляет 45 градусов.Однако первый член (X t.o. ) также внес существенный вклад в это исследование. Компьютерная имитационная модель и численная оптимизация выбрали стратегию увеличения X т.о. с меньшим углом проекции. Хотя предполагается, что оптимальный угол проекции станет ближе к 45 градусам при усилении мышечных параметров модели, настройка компьютерной имитационной модели для конкретной группы пациентов требует очень сложных и сложных процедур.Этот вопрос необходимо рассмотреть в будущих исследованиях.
В этом исследовании оптимальный образец мышечной активации, включая начальный уровень мышечной активации, был найден с использованием численной оптимизации. Это привело к разумным движениям как для горизонтальных, так и для вертикальных прыжковых движений. Замечено, что исходный уровень мышечной активации и мышечные силы не были идентичными между этими движениями (рис. 4 и 5). Такой результат, т.е. расхождения в начальных условиях моделирования, на первый взгляд может показаться спорным.Однако эти начальные условия не были и не были присвоены модели : вместо этого процедура численной оптимизации показала, что эти начальные условия являются наиболее подходящими для модели для выполнения прыжков, начиная с идентичного вертикального положения. Мы полагаем, что общее сходство между моделируемой динамикой тела и динамикой людей предполагает обоснованность подхода, принятого в этом исследовании. Поскольку целевая функция, использованная в этом исследовании, учитывала только поступательные движения центра масс тела, вращательная составляющая механической энергии не анализировалась явно.Не обсуждалась и техника постановки стопы при приземлении. Однако они могут стать более важными при выполнении более точных сравнений между смоделированными и экспериментально зафиксированными движениями или при применении методов и результатов этого исследования к спортивным сценам. Более сложное моделирование и симуляция, включающие этап приземления, будут полезны в будущих исследованиях с целью снижения риска травм у спортсменов.
% PDF-1.3 % 180 0 объект > endobj xref 180 172 0000000016 00000 н. 0000003792 00000 н. 0000003982 00000 н. 0000004013 00000 н. 0000004072 00000 н. 0000005011 00000 н. 0000005436 00000 н. 0000005502 00000 н. 0000005708 00000 н. 0000005798 00000 н. 0000005887 00000 н. 0000005997 00000 н. 0000006100 00000 н. 0000006200 00000 н. 0000006300 00000 н. 0000006415 00000 н. 0000006531 00000 н. 0000006646 00000 н. 0000006762 00000 н. 0000006877 00000 н. 0000006993 00000 п. 0000007107 00000 н. 0000007223 00000 н. 0000007339 00000 н. 0000007453 00000 п. 0000007567 00000 н. 0000007681 00000 н. 0000007777 00000 н. 0000007872 00000 н. 0000007968 00000 п. 0000008064 00000 н. 0000008160 00000 н. 0000008256 00000 н. 0000008352 00000 п. 0000008448 00000 н. 0000008544 00000 н. 0000008640 00000 н. 0000008734 00000 п. 0000008827 00000 н. 0000008922 00000 н. 0000009016 00000 н. 0000009111 00000 п. 0000009207 00000 н. 0000009303 00000 н. 0000009397 00000 н. 0000009493 00000 п. 0000009588 00000 н. 0000009682 00000 н. 0000009778 00000 п. 0000009873 00000 н. 0000009968 00000 н. 0000010063 00000 п. 0000010158 00000 п. 0000010254 00000 п. 0000010350 00000 п. 0000010446 00000 п. 0000010541 00000 п. 0000010637 00000 п. 0000010733 00000 п. 0000010829 00000 п. 0000010924 00000 п. 0000011020 00000 п. 0000011116 00000 п. 0000011212 00000 п. 0000011307 00000 п. 0000011403 00000 п. 0000011499 00000 п. 0000011595 00000 п. 0000011690 00000 п. 0000011786 00000 п. 0000011882 00000 п. 0000011978 00000 п. 0000012073 00000 п. 0000012169 00000 п. 0000012265 00000 п. 0000012361 00000 п. 0000012456 00000 п. 0000012552 00000 п. 0000012648 00000 п. 0000012744 00000 п. 0000012839 00000 п. 0000012935 00000 п. 0000013031 00000 п. 0000013127 00000 п. 0000013222 00000 п. 0000013318 00000 п. 0000013414 00000 п. 0000013510 00000 п. 0000013605 00000 п. 0000013701 00000 п. 0000013797 00000 п. 0000013893 00000 п. 0000013988 00000 п. 0000014084 00000 п. 0000014180 00000 п. 0000014276 00000 п. 0000014371 00000 п. 0000014467 00000 п. 0000014563 00000 п. 0000014659 00000 п. 0000014754 00000 п. 0000014850 00000 п. 0000014946 00000 п. 0000015042 00000 п. 0000015137 00000 п. 0000015233 00000 п. 0000015329 00000 п. 0000015425 00000 п. 0000015520 00000 п. 0000015616 00000 п. 0000015712 00000 п. 0000015808 00000 п. 0000015903 00000 п. 0000015999 00000 н. 0000016095 00000 п. 0000016191 00000 п. 0000016286 00000 п. 0000016382 00000 п. 0000016478 00000 п. 0000016574 00000 п. 0000016668 00000 п. 0000016763 00000 п. 0000016858 00000 п. 0000016954 00000 п. 0000017189 00000 п. 0000017230 00000 п. 0000017254 00000 п. 0000028159 00000 п. 0000028183 00000 п. 0000039943 00000 н. 0000039967 00000 н. 0000050793 00000 п. 0000050817 00000 п. 0000062496 00000 п. 0000062520 00000 п. 0000074759 00000 п. 0000074783 00000 п. 0000085578 00000 п. 0000085833 00000 п. 0000086188 00000 п. 0000086957 00000 п. 0000087612 00000 п. 0000087704 00000 п. 0000087931 00000 п. 0000088296 00000 п. 0000088530 00000 п. 0000088799 00000 н. 0000089038 00000 п. 00000
00000 п. 00000 00000 п. 0000092403 00000 п. 0000092851 00000 п. 0000093663 00000 п. 0000094346 00000 п. 0000111795 00000 н. 0000112340 00000 н. 0000112789 00000 н. 0000113274 00000 н. 0000113592 00000 н. 0000113907 00000 н. 0000114079 00000 п. 0000120499 00000 н. 0000120523 00000 н. 0000131528 00000 н. 0000131552 00000 н. 0000144361 00000 н. 0000144440 00000 н. 0000150295 00000 н. 0000150639 00000 н. 0000153314 00000 н. 0000004113 00000 п. 0000004989 00000 н. трейлер ] >> startxref 0 %% EOF 181 0 объект > endobj 182 0 объект [ 183 0 руб. ] endobj 183 0 объект > / Ж 206 0 Р >> endobj 184 0 объект > endobj 350 0 объект > ручей HTILa ~ vj-DB «№7- \Сгибание и разгибание: записывайте антагонистические мышцы
Фон
Как объяснялось в предыдущем эксперименте, когда вы решаете совершить движение, ваши верхние мотонейроны из моторной коры головного мозга посылают потенциалы действия (всплески!), Перемещающиеся по вашему кортикоспинальному тракту (спинному мозгу), где они синапсируются с нижними мотонейронами, которые продолжают действовать. распространяют сигнал на обозначенные мышечные волокна.Здесь каждый отдельный мотонейрон и мышечные волокна, которые они иннервируют, создают «двигательную единицу». В этом нервно-мышечном соединении высвобождается нейротрансмиттер, называемый ацетилхолином, который приводит к генерации потенциала действия в мышечном волокне. При получении этого возбуждающего сигнала волокна мышц сокращаются. Движение!
Ваше тело использует этот каскад электрических и химических сигналов для управления движением, но важно отметить, что каждое движение зависит не от одной мышцы, а от многих! В человеческом теле более 600 скелетных мышц! Большинство этих мышц функционируют в «антагонистических парах», что означает, что когда одна мышца сокращается (укорачивается), другая в паре расслабляется при тщательном контроле, чтобы позволить удлинение (растяжение).Подобная координация мышц дает нам возможность двигаться грациозно, например, касаясь кончиком носа, не ударяя себя по лицу. Также, чтобы делать точные движения, например, заправлять нить в иглу.
Поговорим подробнее об этих «антагонистических парах». Прекрасный пример пары — двуглавая мышца плеча и трицепс плеча.
Прежде чем мы пойдем дальше, нам нужно сначала рассмотреть несколько анатомических терминов движения. «Сгибание» — это изгибающее движение, при котором угол между двумя частями уменьшается.Сокращение бицепса демонстрирует сгибание, то есть приближает предплечье к предплечью и уменьшает угол между ними. Итак, ваш бицепс описывается как мышца-сгибатель. На рисунке ниже изображение справа показывает сгибание бицепса. Противоположная мышца сгибателя называется мышца-разгибатель. Ваш трицепс — разгибатель. Когда вы сокращаете трицепс, ваша рука выпрямляется, и угол между предплечьем и плечом увеличивается. Возможно, вы уже догадались, но это называется «расширением», и вы можете видеть это на левой иллюстрации ниже.Эти обозначения являются внутренними, то есть они являются неизменным свойством мышцы. Это означает, что при сокращении мышцы-сгибателя всегда происходит сгибание и никогда не происходит разгибание (и наоборот, при сокращении мышц-разгибателей).
Итак, теперь, когда мы установили наши условия движения, мы можем обсудить эти антагонистические пары как следует! Две мышцы антагонистической пары находятся в оппозиции. То есть, если одна конечность разгибает во время ее сокращения, другая вернет конечность в исходное положение при сгибании.В каждой паре, в зависимости от движения, одна мышца играет роль «агониста», а другая — «антагониста». Агонист — это мышца, которая сокращается, вызывая движение. Антагонист — это противоположная мышца, которая расслабляется относительно растяжения. Эти две роли, агониста и антагониста, можно менять местами. Чтобы визуализировать это, вернемся к нашему примеру с бицепсами и трицепсами. Изображение, машущее лучшему другу: когда ваша рука отодвигается от вас, ваш трицепс действует как агонист, сокращаясь, чтобы вытянуть руку.Ваш бицепс — антагонист, расслабляющий, чтобы позволить удлинение, и, возможно, очень легкое сокращение, чтобы контролировать скорость движущегося предплечья.
Когда ваша рука движется обратно во время вашего махового движения, ваш бицепс является агонистом, сгибая руку к вам. В этом случае ваш трицепс является антагонистом и должен расслабиться, чтобы растянуться, чтобы позволить движение. Итак, вы можете видеть, что в отличие от внутренних обозначений сгибателей и разгибателей, две роли антагонистических пар зависят от движения.Если движение меняется на противоположное, агонист и антагонист меняются ролями.Но не всегда все так просто! В некоторых движениях, таких как отжимания, агонист не меняется с направлением движения. Во время отжиманий трицепс является агонистом независимо от того, опускаете ли вы свое тело по направлению к земле или отжимаетесь. Это потому, что приложена нагрузка от силы тяжести. Если вас постоянно тянет вниз постоянная сила, ваши трицепсы должны оставаться в сокращении, чтобы удерживать вас, независимо от того, двигаетесь ли вы вверх или вниз.А теперь брось и дай мне 20!
Сделав еще один шаг вперед, важно понимать, что даже когда вы не двигаетесь, мышцы постоянно работают к вам, чтобы поддерживать осанку. Да, даже когда вы сутулитесь в своем La-Z-Boy, ваш мозг приказывает мышцам сокращаться! Таким образом, ключевым моментом является то, что, когда вы думаете сделать какое-либо движение или даже просто остановиться, наш мозг (в частности, моторная кора) посылает потенциалы действия (шипы!) По нашим нервам не только в одну, но и в несколько мышц, чтобы координировать движение. с контролем или просто чтобы оставаться в равновесии и в вертикальном положении.
Видео
Эксперимент
Теперь давайте воспользуемся этим 2-канальным Muscle SpikerBox, исследуя нашу любимую антагонистическую пару.
- Поместите два наклеенных электрода на бицепс и подсоедините 2 красных зажима типа «крокодил» из канала 1 (правая сторона устройства) к металлическому выступу электродов. Затем поместите две наклейки на электроды на трицепс и подключите к ним красные кабели канала 2.Наконец, поместите один электрод-наклейку на тыльную сторону руки и подсоедините оба черных зажима типа «крокодил» (от каналов 2 и 3) к этому электроду.
- Подключите стереодинамик к одному из 2-канальных аудиоразъемов Muscle SpikerBox. Подключите (с помощью синего кабеля) линейный вход вашего компьютера (он же микрофон) к другому аудиоразъему. Затем откройте программу SpikeRecorder. Вам нужно будет нажать кнопку «Конфигурация» в верхнем левом углу SpikeRecorder, чтобы включить левый и правый входы.
Кроме того, вы можете подключить второй аудиоразъем к своему смартфону (с помощью зеленого кабеля) и открыть приложение Backyard Brains. Бесплатно для: Android или iPhone. Обратите внимание, что оба сигнала будут отображаться вместе с текущим приложением.
- Включите оба канала ЭМГ, вращая каждое колесико, пока не услышите активность во время сокращения бицепсов и трицепсов. Посмотрите на приложение и убедитесь, что вы видите свои сигналы! Если сигналы отсекаются (попадают в верхнюю и нижнюю часть окна), уменьшите усиление, повернув регулировочное кольцо вниз.
- А теперь давайте начнем эксперименты!
А) Начнем с размахивания. Вы можете заставить кого-нибудь помахать вам в ответ? Как вы думаете, какие мышцы вы используете для выполнения этого действия? Чтобы исследовать свою гипотезу, проведите эксперимент, изменив положение наклеенных электродов, пока не увидите ЭМГ, полную потенциалов действия. Помните, что будьте хорошим ученым, документируя свой метод и собирая данные!
Поднимите свое приветствие жестами с помощью дополнительных мышечных усилий, чтобы вы могли лучше визуализировать / слышать ЭМГ.Можете ли вы использовать свои две ЭМГ, чтобы проверить, какая мышца является агонистом при сгибании руки? Как насчет того, чтобы протянуть руку?B) Мышца-антагонист тоже может показывать шипы, как вы думаете, почему? Постарайтесь изучить движения своей руки, пока не сможете полностью расслабить антагониста (без шипов).
- Пойдем еще дальше, сделаем отжимание! (Если пол грязный или вы просто не любите отжиматься, вы также можете сделать отжимание на краю прочного стола, прислонившись к нему.) Можете ли вы проверить с помощью ЭМГ, что трицепс является агонистом, поднимаетесь вы или опускаетесь?
Вот наши результаты (ЭМГ бицепса вверху оранжевым, ЭМГ трицепса внизу желтым):
На этом скриншоте выше показаны две ЭМГ во время фазы отжимания отжимания. Бицепсы выглядят расслабленным антагонистом, и эти трицепсы определенно делают больше шипов! На снимке экрана выше показаны две ЭМГ во время фазы опускания отжимания. Мы видим, что, хотя направление движения изменилось, трицепсы все еще несут нагрузку и играют роль агониста.
Предложения по дальнейшим экспериментам:
Можете ли вы вспомнить другие антагонистические пары в вашем теле? Проверьте свою гипотезу, записав ЭМГ этих двух мышц, чтобы убедиться, что вы правы. Помните, у одного должно быть много спайков (агонист), в то время как у другого должно быть мало или вообще не должно быть (антагонист) и наоборот. Еще одно интересное исследование — попытаться найти две мышцы, которые отображают потенциал действия в своих ЭМГ, даже когда вы стоите или сидите неподвижно. 2-канальный Muscle SpikerBox также позволяет вам сравнивать свои мышцы с мышцами ваших друзей.Наблюдайте за своими ЭМГ во время армрестлинга!
Или попробуйте поединок по армрестлингу без помощи рук. Победитель достается тому, кто сможет дольше держать шипы!Воспользуйтесь преимуществами этих двух каналов и расскажите нам о результатах ваших экспериментов! Отправьте нам свой процесс и результаты по электронной почте [email protected]!
Банкноты
Если у вас есть наш оригинальный 2-канальный Muscle SpikerBox с 6 отдельными выводами (а не 2 оранжевыми кабелями), цвета зажимов типа «крокодил» будут другими. Красный и черный зажимы должны охватывать интересующую мышцу, в то время как белый используется в качестве заземляющего электрода.Механическое поведение мышц при сгибании и разгибании нижней конечности в различных возрастных группах с использованием BRG.LifeMod
- Нитин Сахаи
- Рави П. Тевари
- Локеш Сингх
Документ конференции
- 3 Цитаты
- 1,6 км Загрузки
Abstract
Основная цель этой статьи — выяснить мышцу, которая играет важную роль во время сгибания и разгибания на моделях разных возрастных групп, а также степень механической деформации, которая имеет место в мышцы нижней конечности с помощью БРГ.Программное обеспечение Life Mod, которое работает в среде ADAM. Момент мышц получается с помощью симуляции обратной и прямой динамики модели человека, которая показывает нам, какая мышца активна во время сгибания и разгибания и какой момент (крутящий момент) эта мышца прилагает, что вызывает деформацию мышц. Согласно механической модели мышцы Хилла, она состоит из трех элементов: один сократительный элемент, соединенный последовательно с эластичным элементом, и оба они прикреплены параллельно эластичному элементу.Во время сокращения и расширения мышц происходит деформация мышц, и в течение одного полного цикла ходьбы они восстанавливают свою первоначальную форму и размер благодаря своей вязкоупругой природе. Деформация мышц получается путем определения изменения длины мышц во время расширенного сгибания и разгибания в полном цикле походки на моделях различных возрастных групп. После проведения моделирования на моделях разной возрастной группы было замечено, что деформация мышц нижней конечности максимальна в Rectus femoris, которая является одной из четырех четырехглавых мышц тела человека, остальные — Vastus medialis, Vastus intermediateus, и Vastus lateralis.На основании нашего эксперимента сделан вывод, что Rectus femoris играет важную роль в регулировании сгибания и разгибания колена.
Ключевые слова
Сгибание и разгибание модели мышц холма Цикл походкиЭто предварительный просмотр содержания подписки,
войдите в систему, чтобы проверить доступ.
Предварительный просмотр
Невозможно отобразить предварительный просмотр. Скачать превью PDF.
Информация об авторских правах
© Springer-Verlag Berlin Heidelberg 2010
Авторы и филиалы
- Nitin Sahai
- Ravi P.Тевари
- Локеш Сингх
- 1. Кафедра прикладной механики, Национальный технологический институт Мотилала Неру, Университет Дидмед, Аллахабад, Индия
Сгибание в локтевом суставе — обзор
3 Знать среду использования
Разработчики устройств должны знать, какое устройство будет использоваться он должен соответствовать всем аспектам среды использования, чтобы гарантировать безопасность и эффективность конструкции. По сути, конструкции устройств зависят от контекста; он может не подходить для всех приложений или мест.
Это требует исследования среды использования, чтобы определить те ограничения, которые конструкция устройства должна учитывать, такие как вспомогательное оборудование в рабочем пространстве, рабочая высота и ориентация, освещение, шум, температура, температура, а также возможные отвлекающие факторы. Каждый из них вносит свой вклад в конкретные требования к дизайну, например конструкция должна обеспечивать тепловую защиту во всех точках взаимодействия при использовании из-за экстремальных температур.
Особые экологические соображения включают:
- •
Используйте рабочее пространство
Устройства используются в различных рабочих местах (например,g., больницы, дома и мобильные сценарии), а также могут быть развернуты в нескольких средах. Каждый уникальный сценарий использования должен быть проанализирован на предмет уникальных характеристик. Среды многократного использования могут иметь конфликтующие факторы, которые необходимо уравновесить во время разработки.
- •
Использование высоты и ориентации рабочего пространства
Высота, ориентация и расположение устройства в окружающей среде напрямую влияют на то, как оно используется. Например, при лапароскопии пользователи могут иметь серьезные сгибания плеча и локтя в результате конструкции пистолетной рукоятки, в результате положения пользователя относительно высоты хирургического стола, и, хотя это не обязательно проблематично, это вызовет дискомфорт у пользователей и уменьшит возможности в силах, необходимых для активации средств управления.На использование устройства также влияют другие объекты в среде в отношении хранения и доступа.
- •
Освещение
Освещение включает окружающее освещение, а также прямые источники света. В темноте или в условиях низкой освещенности требуются специальные приспособления, такие как экраны с подсветкой и светящиеся точки касания для облегчения идентификации и улучшения читаемости. Устройства, которые находятся в средах с экстремальным освещением, часто требуют мониторов, которые уменьшают / контролируют блики, чтобы обеспечить достаточный контраст маркировки.Решения для освещения зависят от задачи и риска в общей конструкции устройства.
- •
Звуковой шум
Окружающий шум влияет на все звуковые сигналы, которые может подавать устройство. Кроме того, условия срабатывания сигнализации могут вызвать утомление от сигнализации и заставить пользователей игнорировать или игнорировать важные предупреждения. Устройства, обеспечивающие чрезмерную звуковую обратную связь, отвлекают, поскольку могут подавлять важную информацию и / или мешать общению между сверстниками. При развитии слуховых функций необходимо учитывать уровень шума в среде предполагаемого использования.
- •
Температура и влажность
Температура и человеческий фактор напрямую влияют на работу устройства и пользователя. Они также влияют на одежду, которую носят пользователи, например им может потребоваться защитная одежда.
- •
Отвлекающие факторы
Отвлекающие факторы исходят из нескольких источников и принимают различные формы. Пользователи могут отвлекаться на окружающий шум, фоновую музыку, общение между коллегами, многозадачность, усталость и стресс.
Чтобы лучше понять среду использования с целью определения конкретных требований к дизайну, контекстный запрос (глава 5) следует использовать как часть надежной стратегии человеческого фактора.
Для получения дополнительной информации о защите окружающей среды см. ANSI / AAMI HE75 Экологические, организационные и структурные вопросы. Дополнительную информацию о конструктивных ограничениях, связанных с окружающей средой, можно найти в ANSI / AAMI HE75 «Проектирование переносных медицинских устройств и устройств, используемых в неклинических условиях непрофессионалами».
Нейромеханическое моделирование прыжка саранчи
ВВЕДЕНИЕ
Способность убегать от хищников имеет большое эволюционное значение. Многие животные, в том числе саранча, в процессе эволюции специализируются на прыжках как способе бегства и передвижения. Прыжок саранчи опосредуется нервными и биомеханическими механизмами, которые сначала подготавливают животное к прыжку, запасая механическую энергию в ногах, а затем внезапно выпускают ее, чтобы катапультировать животное в воздух. Из-за невозможности произвести необходимые измерения во время прыжка, механизмы в первую очередь изучались во время другого поведения, толчка, когда тело саранчи удерживается неподвижно (Heitler, 1988; Heitler and Burrows, 1977a; Heitler and Burrows, 1977b). ).И удар, и прыжок включают быстрое разгибание ноги, но, поскольку удар не нагружен, он происходит гораздо быстрее, чем прыжок. Несмотря на это различие, сходная нервно-мышечная активность до и во время обоих видов поведения привела к предположению, что двигательные программы удара и прыжка одинаковы (Heitler and Burrows, 1977a).
Двигательная программа саранчи «толчок / прыжок» состоит из трех фаз (Берроуз, 1995; Хейтлер и Берроуз, 1977а). Первый этап — взведение ноги. Саранча готовится к прыжку, активируя сгибатель большеберцовой кости, чтобы привести большеберцовую кость в полностью согнутое положение.Вторая фаза — это период совместного сокращения, при котором задействованы как разгибатели, так и сгибатели большеберцовой кости. Мышца-разгибатель большеберцовой кости медленно сокращается и накапливает энергию в аподеме разгибателя, в полулунном отростке (SLP) и в кутикуле ноги. По мере того как в сгибателе большеберцовой кости нарастает напряжение, его дистальное сухожилие проходит над кутикулярной инвагинацией в вентральной части дистального вентрального отдела бедра, называемой шишкой Гейтлера (Bennet-Clark, 1975). Это позволяет сухожилию сгибателя большеберцовой кости прикрепляться к большеберцовой кости под углом почти 90 градусов.когда большеберцовая кость полностью согнута, тем самым максимизируя ее механическую эффективность в поддержании сгибания большеберцовой кости против возрастающего напряжения гораздо более крупной мышцы-разгибателя большеберцовой кости (Heitler, 1974). Шишка также действует как захват или фиксатор на сухожилии, что помогает поддерживать большеберцовую кость согнутой, в то время как мышца-сгибатель находится в напряжении. В третьей фазе скачок запускается, когда и мышца-сгибатель, и ее двигательные нейроны подавлены. Когда напряжение сгибателя опускается ниже порогового значения, сухожилие соскальзывает с захвата, и большеберцовая кость быстро растягивается для выполнения прыжка (Heitler, 1974).
Технические ограничения мешают четкому пониманию роли SLP в ударах и прыжках. SLP — это сильно склеротизированная часть кутикулы бедренно-большеберцового сустава заднегруди. При подготовке к удару или прыжку SLP изгибается, как лук для стрельбы из лука, во время медленного сокращения мощной мышцы-разгибателя, чтобы накапливать энергию, которая позже быстро высвобождается для усиления удара или прыжка (Bennet-Clark, 1975). Функция SLP была выведена из его движения во время ударов ногами, записанного на высокоскоростном видео, и из расчетов энергии, запасенной в аподеме разгибателя, SLP и кутикуле бедренной кости (Bennet-Clark, 1975; Burrows and Morris, 2001 ).Энергия, накопленная в SLP, и время ее высвобождения, по-видимому, важны для выполнения прыжка. Высокоскоростная видеозапись ударов саранчи показывает, что SLP не начинает разворачиваться, пока большеберцовая кость не повернется более чем на 30 градусов. (Берроуз и Моррис, 2001). Причина этой задержки и ее роль в динамике прыжка неизвестны. Повреждение SLP, который является неотъемлемой частью сустава ноги, делает саранчу неспособной прыгать, и поэтому сравнение прыжковых характеристик животных с функциональной SLP и без нее практически невозможно.
Развивающаяся область нейромеханического моделирования может помочь решить эти проблемы (Pearson et al., 2006). Мы использовали новый нейромеханический симулятор AnimatLab (www.animatlab.com), чтобы построить и протестировать модель саранчи на основе текущих описаний нейронных и биомеханических процессов, управляющих прыжком. Затем модель саранчи была помещена в физически реалистичную среду в AnimatLab, чтобы изучить как способность программы толкательного мотора производить реалистичный прыжок, так и влияние SLP на поведение прыжков.
МАТЕРИАЛЫ И МЕТОДЫ
Животные
Взрослые особи саранчи, Schistocerca americana L., были получены из колонии в Колледже Агнес Скотт, Декейтер, Джорджия, США, содержались в клетках небольшими группами при 27 ° C в течение 12 ч: 12 ч цикла L: D и кормили свежим органическим салатом и смесью 2/1 свежих зародышей пшеницы и сухого молока. Особей переводили из клетки в комнату для видеозаписи и помещали на платформу для прыжков. Платформа содержала нагревательный элемент, который мог регулировать местную температуру, и была покрыта очень мелкой наждачной бумагой, чтобы саранча не скользила по поверхности для прыжков.Желтая деревянная мишень размером 25 см × 30 см помещалась в 30 см от платформы, и прыжки к цели вызывались либо легкими прикосновениями к животу ручной палочкой, либо повышением температуры платформы. Животные были извлечены после прыжка и возвращены на платформу для еще одной попытки. Прыжки вызывались примерно с 5-минутными интервалами; После 10 прыжков особи возвращались в клетку. Скачки саранчи регистрировались при 500 кадрах с −1 и разрешении 512 × 240 пикселей двумя видеокамерами Photron PIC R2 Fastcam (Сан-Диего, Калифорния, США) с временем экспозиции 0.5 мс.
Locust модель
Чтобы отличать ссылки на модель и ее части от ссылок на саранчу, названия частей модели были выделены курсивом и обозначены соответствующими частями тела саранчи, тогда как ссылки на саранчу и ее части сделаны обычным шрифтом.
Трехмерная графическая модель тела саранчи была разработана на основе трехмерной многоугольной сетки, которая была куплена в Интернете (www.turbosquid.com), а затем разделена на отдельные сегменты тела с помощью графической программы Blender (www.blender.org). Многоугольная сетка — это набор вершин и треугольных граней, которые определяют объем этого сегмента. Размеры каждого сегмента были изменены в соответствии с опубликованными анатомическими измерениями (Bennet-Clark, 1975; Heitler, 1974). Предполагалось, что все сегменты имеют однородную плотность, и распределение массы по всему объему ячейки определяет момент инерции для этого сегмента . Модель имеет длину тела 48 мм и общую массу 2,5 г, с заднегруди, и голени, длиной 26 мм (Bennet-Clark, 1975).Отдельные части тела и конечности , сегменты были соединены статическими или плоскими шарнирами в AnimatLab для сборки модели тела саранчи. Угловые ограничения на шарнирные соединения были установлены, чтобы ограничить движение каждого сустава до нормального диапазона соответствующего сустава животного. Чтобы гарантировать, что центр масс (COM) всего саранчи был расположен надлежащим образом, небольшие утяжеленные массы были размещены вдоль оси тела грудной клетки
и брюшных сегментов , чтобы отрегулировать распределение массы в пределах body (Беннет-Кларк, 1975).Затем определяли COM, прикрепляя корпус к шарнирному соединению и позволяя ему свободно вращаться. Массу перераспределяли до тех пор, пока саранча не сбалансировалась как по вертикали, так и по горизонтали в желаемом месте. Модель саранчи, использованная для этих симуляций, можно найти на сайте www.animatlab.com/locust.Биомеханика
Геометрия и биомеханические свойства бедренно-большеберцового сустава заднегруди играют решающую роль в накоплении энергии для прыжка (рис.1). Разгибатель большеберцовой кости / аподема показан в виде красной линии, которая прикрепляется к большеберцовой кости (рис. 1a), тогда как сгибатель большеберцовой кости мышца / аподема показаны зеленым цветом. Он охватывает шишку Гейтлера (рис. 1h) и прикрепляется к большеберцовой кости (рис. 1c). Бедренная кость была соединена с небольшим блоком массой 1,6 мг, который представлял SLP (рис. 1f) (Bennet-Clark, 1975). Скользящий призматический шарнир соединял SLP с бедренной костью (рис.1г). Во время нормального совместного сокращения дистальный конец SLP (где прикрепляется большеберцовая кость) перемещается на 0,3 мм вентрально и на 0,4 мм проксимально (Burrows and Morris, 2001). Шарнир скольжения был ориентирован так, чтобы позволить массе SLP двигаться в том же направлении (рис. 1b – g). Пружина прикрепляла массу SLP к бедренной кости и была ориентирована вдоль направления движения скользящего сустава (рис. 1e). Жесткость полулунной пружины рассчитывалась по кривой зависимости напряжения от деформации, полученной для SLP (Bennet-Clark, 1975).Для растяжения отростка параллельно аподеме разгибателя на 0,4 мм требуется приблизительно 14,2 Н силы (Bennet-Clark, 1975). Однако SLP перемещается как проксимально, так и вентрально, и эта величина проксимальной деформации соответствует 0,3 мм вентральной деформации для 0,5 мм общей деформации. Исходя из этой деформации, мы рассчитали жесткость SLP как 28,4 кН м -1 . Для этого значения была задана константа полулунной пружины .
Рис.1.Модель бедренно-большеберцовой кости ( FT ) сустава заднегруди .(a) Точка прикрепления аподемы разгибателя на голени . (b) шарнирный шарнир FT и соединение пружины полулунного отростка ( SLP ). (c) прикрепление аподема сгибателя точки на голени . (d) Более дистальная точка большеберцовой кости. (e) Пружина SLP прикреплена между бедренной костью и большеберцовой костью . (f) Масса SLP перемещается по скользящему шарниру (g), ориентированному между точками b и g, с наклоном 36.9 град. (з) Комок Гейтлера . Сгибающая мышца охватывает эту опухоль, чтобы изменить ее ориентацию относительно голени при перемещении ноги . (i) Сухожильный фиксатор моделируется как пружина, расположенная между точками c и i (пурпурная линия). Он доступен только тогда, когда большеберцовая кость полностью согнута и сгибающая мышца имеет натяжение более 0,15 Н (Bennet-Clark, 1975; Heitler, 1974). Расстояние между a, b равно 0,76 мм, b, c равно 1.64 мм. Угол a, b, c равен 144 градусам. а b, c, d — 143 град. (Heitler, 1974). Показана модель мышц сгибателей и разгибателей. Он состоит из пружины ( K pe), подключенной параллельно генератору натяжения (T), и демпфера (B), подключенного последовательно с другой пружиной ( K se). (1) Мышца активируется за счет возбуждения двигательного нейрона (МН). (2) Это деполяризует мышечную мембрану. (3) Изменения мембранного напряжения ( В м ) преобразуются в значение натяжения с использованием сигмоидальной функции.(4) Значение натяжения масштабируется в зависимости от длины мышцы. (5) Масштабное натяжение прилагается к мышце генератором силы для создания сокращения. Мышца-разгибатель большеберцовой кости / аподема показана красной линией, а мышца-сгибатель большеберцовой кости / аподема показана зеленым.
Голень была соединена с массой SLP с помощью шарнирного соединения, которое позволяло большеберцовой кости поворачиваться на 5 градусов. и 160 град. (Рис. 1б). Шарнирный сустав бедра и большеберцовой кости соединен с массой SLP, так что во время совместного сокращения и разгибания большеберцовой кости шарнирный сустав будет перемещаться вдоль ползунка с массой SLP , чтобы приблизительно соответствовать движению сустава, наблюдаемому у саранчи. (Берроуз и Моррис, 2001).Расстояния и углы, которые определяют отношения между насадкой-разгибателем , шарнирным суставом бедра и большеберцовой кости и насадкой-сгибателем , были установлены на опубликованные измеренные значения (расстояние между a, b составляет 0,76 мм, а между b, c — 1,64 мм. Угол a, b, c составляет 144 градуса, а угол b, c, d равен 143 градусам) (Heitler, 1974).
Мышца представлена в AnimatLab линейной моделью мышц Хилла (Hill, 1970; McMahon, 1984; Shadmehr and Wise, 2005a; Shadmehr and Wise, 2005b).Каждая мышечная модель состоит из последовательной пружины (Kse), соединенной последовательно с параллельной комбинацией параллельной пружины (Kpe), демпфера (B) и силового привода (T). Свойства модели мышцы определяются длиной мышцы в состоянии покоя, константами пружины и рывка, кривой «стимул – напряжение» и кривой «длина – напряжение». Кривая «стимул – напряжение» — это сигмоидальная функция, которая связывает уровень силы исполнительного механизма с деполяризацией мышечной мембраны. Кривая «длина – натяжение» представляет собой обратную параболу, которая определяет процент силы привода, прилагаемой к данной длине мышцы.
В этом моделировании моделируются только две мышцы каждой из задних ног: сгибатель голени и разгибатель голени. Максимальная сила, которую может создать разгибатель, составляет 15 Н, что достигается при деполяризации после латентного периода в 300-800 мс (Bennet-Clark, 1975). Серийная жесткость пружины разгибателя была рассчитана с использованием модуля Юнга 18,9 кН мм -2 , найденного для аподема разгибателя Беннет-Кларком (Bennet-Clark, 1975). Средний размер образцов аподема составлял 3 мм в длину × 0.Ширина 25 мм и толщина 40 мкм, поэтому они имеют площадь 0,01 мм 2 и длину 3 мм. Эти значения позволили нам рассчитать жесткость пружины как 63 кН м −1 из модуля Юнга с использованием уравнения K = YA / L , где Y — модуль, A — площадь и L — длина. В отсутствие опубликованных измерений, которые позволили бы рассчитать жесткость параллельной пружины, мы использовали значение 20 Н · м -1 , потому что оно создавало небольшое, но заметное напряжение при растяжении мышцы-разгибателя .Коэффициент демпфирования мышцы-разгибателя был вручную установлен на 700 Н · м -1 , чтобы обеспечить время нарастания до пикового напряжения примерно 400 мс. Кривая «стимул-напряжение» и характеристики отклика нейрона без шипов , который представляет мышечную мембрану, были настроены так, чтобы воспроизводить подергивающую реакцию мышцы-разгибателя на одиночный спайк FETi под углом бедра к большеберцовой кости 90 градусов. (Heitler, 1988). Кривая длина – напряжение была также воспроизведена из значений мышечных сокращений, которые были сняты при различных углах бедра и голени путем стимуляции мышцы (Bennet-Clark, 1975).Стимуляция мышцы-разгибателя вызывала судорожные реакции, очень похожие на те, которые были зарегистрированы для мышцы-разгибателя в ответ на спайк FETi. Результирующая кривая «длина – натяжение» мышцы-разгибателя достигла максимума в полностью согнутом положении и уменьшалась при разгибании ноги .
Записи сгибающей мышцы показали, что она производит максимальное напряжение около 0,75 Н в ответ на тетанизирующую стимуляцию и что максимальное напряжение достигает 35-40 мс после задержки в 15 мс (Bennet-Clark, 1975).Следующие значения параметров позволили модели сгибающей мышцы воспроизвести записанный временной ход пикового напряжения и напряжения; жесткость последовательной пружины K se составляла 100 Н · м −1 , жесткость параллельной пружины K pe составляла 20 Н · м −1 , а коэффициент демпфирования B составлял 10 Н · м -1 . Кривая «стимул – напряжение» была настроена для получения желаемого максимального напряжения. Как и в случае модели разгибателя , кривая длины-натяжения сгибателя была близка к своему максимальному значению, когда нога была полностью согнута, и почти к своему минимальному значению, когда нога была разогнута.
Сухожилие сгибающей мышцы акации содержит карман. Когда большеберцовая кость полностью согнута, а напряжение сгибателя превышает 0,15 Н, этот карман захватывается шишкой Гейтлера, что помогает удерживать сухожилие на месте (Bennet-Clark, 1975; Heitler, 1974). Это свойство фиксации сухожилий играет важную роль в прыжке после совместного сокращения, когда мышца-сгибатель и двигательные нейроны тормозятся. Замок помогает сохранять большеберцовую кость полностью согнутой даже при снижении напряжения сгибателей.Это предотвращает преждевременное разгибание большеберцовой кости и инициирует прыжок, когда натяжение сгибателя падает ниже порогового значения для сохранения блокировки. В модели фиксатор сухожилия был представлен пружиной, которая соединяет аттачмент сгибателя с точкой бедра (рис. 1, пурпурная линия между точками c и i). Пружина была отключена и не создавала натяжения, если голень не была полностью согнута и натяжение сгибателя не превышало 0,15 Н.
Нейронная модель
В этом моделировании использовалась основанная на проводимости модель нейрона интеграции и возбуждения. Нейроны были смоделированы как отдельные эквипотенциальные отсеки, каждый из которых характеризовался набором определяемых пользователем параметров, включая постоянную времени мембраны, размер (т.е. входную проводимость), напряжение мембраны, шум тока, начальный порог спайков, аккомодацию частоты спайков, спайк после- проводимость гиперполяризации и проводимость кальция с переменными активации и инактивации (MacGregor and Lewis, 1977).
Нейронная сеть, используемая для генерации моторных программ толчка и прыжка была разработана для применения правильных моторных сигналов в последовательности и продолжительности, которые имитируют моторную программу, наблюдаемую при пинании саранчи (Heitler and Burrows, 1977a; Heitler and Burrows, 1977b). Начальное сгибание голени начинается, когда девять быстрых мотонейронов сгибателя большеберцовой кости стимулируются к возбуждению (рис. 2A, зеленые нейроны FLTi ) (Burrows, 1995; Burrows, 1996).Эти нейроны синапсируют с мембраной сгибающей мышцы (рис. 2F, светло-голубой узел FM ), вызывая деполяризацию мышцы и сокращение мышцы-сгибателя . Быстрый разгибатель моторного нейрона большеберцовой кости (рис. 2B, красный нейрон FETi ) синапсирует с мембраной мышцы-разгибателя (рис. 2E, светло-голубой узел EM ), вызывая его сокращение. Центральный возбуждающий синапс соединяет нейрон FETi с быстрыми сгибательными двигательными нейронами (от B до A) (Burrows, 1996; Heitler and Burrows, 1977b).Есть также два тормозных интернейрона , которые участвуют в запуске скачка . Мультимодальный ‘M’ интернейрон (рис. 2C, золотой M нейрон ) ингибирует возбуждающие двигательные нейроны-сгибатели , в то время как ингибиторный двигательный нейрон-сгибатель (рис. 2D, желтый нейрон FI ) синапса на сгибающую мышцу и подавляет ее напрямую (Burrows, 1995; Pearson et al., 1980). Узел управления замком сухожилия (рис.2G, светло-голубой) отвечает за включение в сухожилия запирающей пружины , когда голень достаточно согнуты и сгибателей имеет напряжение выше порога блокировки. Сеть, которая управляет правой заднегруди , показана на рис. 2; идентичная сеть управляет левой заднегруди .
Нейроны были сконфигурированы для воспроизведения наблюдаемых частот срабатывания во время удара. Пик срабатывания нейронов FETi находился в диапазоне от 60 Гц до 100 Гц (Heitler and Burrows, 1977a), в то время как нейроны FLTi срабатывали около 60 Гц (Heitler and Burrows, 1977A).Центральный возбуждающий синапс , соединяющий FETi с FLTi нейронами , был настроен путем воспроизведения эксперимента, в котором FETi стимулировали срабатывание с частотой примерно 10 Гц, а синаптический ответ FLTi отслеживался (Heitler and Burrows, 1977b). Первый спайк FETi произвел 20 мВ EPSP (возбуждающий постсинаптический потенциал) во всех мотонейронах FLTi ; ВПСП распадались примерно за 100 мс (Burrows, 1996; Heitler and Burrows, 1977b).Ответы на последующие спайков были уменьшены синаптической депрессией аналогично тому, что наблюдали экспериментально (Heitler and Burrows, 1977b).
Все нейронов имели случайный тонический шум 0,3 мВ, добавленный к их мембранным потенциалам на каждом временном шаге. Псевдослучайное число Генератор, который контролировал шум, был инициализирован с использованием случайного начального значения в начале каждого моделирования. Это привело к тому, что каждая симуляция с другим семенем давала немного разные результаты, потому что изменения напряжения нейрона приводили к изменениям во времени моторной программы и временах нарастания и спада напряжения в каждой из мышц .
Рис.2.Модель нейронной сети для создания моторных программ kick и jump . Показанная сеть предназначена для правой ноги. (A) Девять быстрых мотонейронов сгибателей большеберцовой кости (зеленый FlTi ). FlT — это синапс на мембране сгибающей мышцы (голубой FM ). (B) Один быстрый двигательный нейрон-разгибатель большеберцовой кости (красный FETi ). FETi синапсов на мембрану мышцы-разгибателя (голубой EM ).(C) Мультимодальный интернейрон (золото M ) ингибирует FlTis . (D) Ингибитор сгибателей (желтый FI ) ингибирует сгибающих мышц . (E) Деполяризация мембраны мышцы-разгибателя вызывает сокращение мышцы-разгибателя . (F) Деполяризация мембраны сгибающей мышцы заставляет сгибающуюся мышцу сокращаться. (G) Узел управления фиксацией сухожилия (голубой) управляет включением пружины фиксации сухожилия на основе вращения голени и натяжения сгибающей мышцы .(H) Когда запускается прыжок , сустав бедро-большеберцовая кость быстро вращается, чтобы произвести прыжок или прыжок .
Порядок моделирования экспериментов
Во время моделирования kick , саранча была подвешена над землей и повернулась так, чтобы ее нижняя поверхность была наверху, и закреплена на месте так, чтобы она не могла упасть. Все сустава ног , за исключением суставов бедра и голени и суставов большеберцовой кости и предплюсны из заднегруди были заблокированы для предотвращения вращения.Программа толкающего двигателя заставляла большеберцовую кость сначала сгибаться, а затем толкаться с высокой скоростью. Это позволило нам измерить движение SLP и большеберцового вращения . Крутящий момент SLP относительно прикрепления разгибателя был рассчитан путем записи координат сустава бедренно-большеберцовой кости, прикрепления разгибателя и вектора силы SLP . Эти значения использовались для расчета плеча момента вектора силы SLP относительно крепления разгибателя , и это было использовано для расчета крутящего момента, прикладываемого SLP.Скорость удара была измерена как пиковая скорость между началом удара и концом удара , когда большеберцовая кость полностью повернулась на 160 градусов. Удар — это время от начала удара до полного поворота голени .
Тот же образец стимула, который использовался для удара ногой , был применен для создания двигательного рисунка для прыжка . Моделирование саранчи началось с саранчи, проведенной 4.5 см над землей , а затем упал на землю . Изначально только бедренно-большеберцовые суставы из задних ног могли свободно вращаться. Все остальные шарнира были заблокированы и неподвижны, а задние ноги были подняты в воздух, чтобы голень могла свободно вращаться без столкновения с землей . После того, как большеберцовая кость была полностью согнута, коаксиальный коаксиально-бедренный сустав был отрегулирован так, чтобы зафиксировать угол опоры относительно земли и обеспечить угол отрыва примерно 55 градусов.Этот угол был выбран, потому что он производил прыжка , что минимизировало кувырок во время взлета. суставы для всех остальных ног оставались заблокированными на протяжении всей программы прыжковой моторики , чтобы поддерживать стабильную и последовательную осанку, а положение передних ног было скорректировано, чтобы зафиксировать первоначальный шаг тела животного на 2 град. Замки на шарнирах передних и средних ног были отключены при срабатывании прыжка .Это позволило всем опорам свободно перемещаться во время взлета и баллистической фазы прыжка . SLP был отключен для испытаний путем блокировки скользящего призматического шарнира SLP , чтобы предотвратить его перемещение, и путем отключения пружины SLP , чтобы предотвратить возникновение напряжения. Натяжение разгибателя контролировали путем изменения частоты возбуждения мотонейронов FETi .
Прогнозируемое расстояние прыжка было рассчитано по формуле: d = 2⋅E⋅sin (2⋅θ) m⋅g, (1) где d — расстояние, E — энергия, θ — средний угол взлета для всех смоделированных прыжков, м, — масса и г, — сила тяжести (Bennet-Clark, 1975).Мощность скачка была рассчитана как скалярное произведение силы, действующей на тело во время скачка импульса и скорости тела (Bennet-Clark, 1975). Энергия скачка была рассчитана путем интегрирования кривой мощности за период времени импульса. Начало прыжка или Удар всегда отсчитывался от момента, когда фиксатор сухожилия был отключен. Прыжок Продолжительность — это время от начала до тех пор, пока либо тело , либо одна из задних ног не коснется земли .Расстояние прыжка было измерено как разница между положениями саранчи в конце и в начале прыжка . Продолжительность импульса прыжка — это время от начала прыжка до тех пор, пока одна из ног не потеряла контакт с землей . Скорость и ускорение прыжка были максимальными из значений, полученных во время импульса прыжка . Все данные анализировались в Matlab (Matlab R2007a, Mathworks Inc., Натик, Массачусетс, США), а статистические сравнения были выполнены с использованием одностороннего анализа дисперсионной функции Anova1.
Влияние момента сгибания SLP на сгибающую мышцу было определено путем сравнения уровня напряжения, при котором разгибатель смог преодолеть напряжение в сгибателе , когда пружина SLP была цела и когда он был отключен. Натяжение разгибателя было установлено на 5 Н, а фиксатор сухожилия был отключен для обоих тестов.Первый тест проводился с неповрежденной SLP , тогда как во втором тесте пружина SLP была отключена. Как только разгибатель достиг уровня натяжения 5 Н, сгибатель был подавлен, и его натяжение уменьшалось до тех пор, пока не достигло точки, в которой разгибатель смог преодолеть его и разогнуть ногу .
РЕЗУЛЬТАТЫ
Моторная программа и прыжки
Смоделированная двигательная программа и паттерны активности мышц , ответственные за толчок , показаны на рис.3. Программа толчкового двигателя началась с стимуляции девяти мотонейронов FlTi с правой и левой стороны (рис. 2) для возбуждения с частотой около 60 Гц, что вызвало напряжение в мышцы-сгибатели , которые привели к полному сгибанию левой и правой голеней (рис. 3A, F). Последовательность токовых стимулов, приложенных к моторнейронам FETi с обеих сторон, начала фазу совместного сокращения (рис. 3B, E). Каждый текущий стимул вызвал соответствующий всплеск в FETi .В дополнение к движению мышцы-разгибателя , FETi возбуждал FlTi с той же стороны (Heitler, 1988), чтобы позволить сгибающей мышце сохранять сгибающуюся большеберцовую кость , несмотря на натяжение установленного разгибателя . . Пик был инициирован стимуляцией FI (рис. 3D) и M ингибирующих нейронов (рис. 3C) приложенным током в течение 80 мс, что привело к их срабатыванию приблизительно с частотой 200 Гц.Нейрон FI подавлял сгибающую мышцу , вызывая быстрое снижение напряжения сгибателя . Одновременно M нейрон ингибировал FlTi s, чтобы удалить привод на сгибающей мышце . Быстрое ингибирование FlTi s и сгибателя запускало толчок , снижая натяжение сгибателя ниже уровня, необходимого для поддержания фиксации сухожилия (рис. 3F, G) (Heitler and Burrows, 1977a ; Пирсон и др., 1980). При отключенном фиксаторе сухожилия сгибателя большеберцовая кость начала очень быстро разгибаться, завершая разгибание за 4 мс, то же время, которое было записано с помощью высокоскоростной видеосъемки (Burrows and Morris, 2001).
Рис.3.Нейронный выход сети скачкового двигателя. Девять сгибающих мотонейронов были стимулированы к срабатыванию (A) во время фазы взведения, что увеличивало напряжение сгибающей мышцы (F) и поворачивало большеберцовую кость в полностью согнутое положение (H).Моторный нейрон-разгибатель FETi (B) начал активироваться, чтобы произвести совместное сокращение и увеличить частоту сгибателя через центральную возбуждающую синаптическую связь от FETi к FLTi . Ингибирующие интернейроны M (C) и ингибитор сгибателей ( FI ) (D) затем начали срабатывать, как только разгибатель достиг желаемого уровня напряжения (E). Это привело к тому, что натяжение сгибателя (F) упало ниже порога фиксации сухожилия (G), что отключило сухожильную пружину.Безальтернативное натяжение привело к быстрому расширению большеберцовой кости (H). Каждая диаграмма соответствует выходу помеченного элемента на рис. 2.
Чтобы проверить гипотезу о том, что одна и та же нейронная схема и моторная программа могут производить и толчок, и прыжок, мы использовали модель цепи толчка и моторную программу, чтобы вызвать имитацию прыжка. Расширенный вид данных для удара и прыжка показан на рис. 4. Хотя одна и та же моторная программа контролировала оба моделируемых поведения, ненагруженная нога выдвинулась менее чем за 5 мс, чтобы произвести толчок (Инжир.4A), тогда как нагрузка, создаваемая телом , заставляла ногу выдвигаться намного более плавно, создавая прыжок (Рис. 4B). Сравнение смоделированных перемещений саранчи с серией кадров прыжка саранчи ( S. americana ) высокоскоростной камерой, работающей на 500 кадрах с −1 (см.Материалы и методы; рис. 5) показывает, что моделирование уловило наиболее характерные особенности результирующего поведения прыжка.При этом, как и в большинстве зарегистрированных нами прыжков саранчи, ноги животного отрывались от земли примерно через 30 мс после начала прыжка, а голова и грудная клетка сохраняли свою ориентацию в течение 45 мс, пока животное оставалось в поле зрения (рис. 5, врезка). Моделирование прыжков саранчи продемонстрировали большинство из этих поведений. Саранча , показанная прыгающая на рис.5, отображает ту же серию движений ног в те же моменты времени, что и саранча, показанная на вставке, хотя ее тело начало вращаться вперед к последнему кадру моделирования. .
Рис.4.Расширенный вид прыжка или удар данные. Kick Данные на рис. 3 расширены и сравниваются с данными из прыжка . Выходные данные моторной программы были одинаковыми как для толчка , так и для прыжка , поэтому здесь не указывались. Каждая диаграмма показывает напряжение в разгибателе и сгибателе мышцы левой заднегрудной ноги , вращение бедренно-большеберцовой кости ( FT ) сустава и состояние фиксации сухожилия .(A) Чтобы произвести толчок , большеберцовая кость начала очень быстро вращаться после отключения фиксатора сухожилия и завершила полное разгибание за 4,1 мс. (B) Прыжок использовал ту же программу двигателя, но нога вращалась медленнее, потому что большеберцовая кость была загружена, и поэтому достигла максимального значения через 28,35 мс. Цветные линии графика имеют тот же цвет, что и соответствующие линии графиков тех же переменных на рис. 3.
Опубликованные измерения прыжкового поведения саранчи ( Schistocerca gregaria ) (Таблица 1) предоставляют эталоны, с которыми можно сравнить поведение прыжка саранчи саранчи .Модель саранчи. прыжка были выполнены со случайным посевом шума, добавленным к мембранным потенциалам всех нейронов модели и мышц , в то время как все остальные параметры модели оставались постоянными. Случайно посеянный мембранный потенциал Шум гарантировал, что саранча вела себя немного по-разному для каждого прыжка , моделируемого при заданном наборе значений параметров, исключительно из-за случайности в нейронах и их влияния на биомеханику.Это предоставило метод для измерения дисперсии поведения , когда все остальные начальные условия были идентичны. Изменяя частоту срабатывания FETi , напряжение разгибателя , можно было охватить весь диапазон поведенческих данных, которые наблюдались у живых животных. Несколько показателей прыжка производительности были получены и сопоставлены с опубликованными контрольными значениями для живой саранчи (Таблица 1) и оказались практически такими же. FETi с частотами от 44 Гц до 100 Гц произвело разгибателя напряжения от 9,4 Н до 15 Н, что привело к прыжкам между 46 см и 120 см длиной (таблица 1; круги на рис. 6A, R 2 = 0,96). Моделирование наиболее близко соответствовало данным в реальном времени, когда частота FETi была установлена на 50 Гц, что вызывало разгибателя напряжений 12 Н и прыжка длиной 64 см. Энергия для прыжка также изменялась линейно и очень хорошо соответствовала прогнозируемому расстоянию для этого уровня энергии и угла взлета (рис.6B, R 2 = 0,98). Эти сходства демонстрируют, что производительность jump у саранчи с использованием моторной программы kick близко соответствует таковой у живой саранчи.
Рис.5.Скриншоты смоделированного и реального прыжка саранчи. На вставках — изображения настоящего прыжка саранчи. Смоделированная саранча дает скачка , очень похожий на те, которые зарегистрированы для живой саранчи. Изображения живой саранчи представляют собой последовательные кадры, снятые с помощью высокоскоростной камеры с частотой 500 кадров -1 .
Роль SLP
Вклад SLP в прыжок саранчи был проанализирован путем сравнения прыжков , сделанных с SLP в неповрежденном состоянии, с прыжками , сделанными с отключенным. Без SLP расстояние прыжка на оставалось пропорциональным натяжению разгибателей , но наклон линии регрессии был уменьшен на 40%, а прогнозируемое пересечение X (минимальное натяжение разгибателей , необходимое для создания скачок ) был больше (рис.6A квадраты, R 2 = 0,95). Разница в наклоне указывает на то, что SLP увеличивает эффект изменения напряжения разгибателя на расстояние прыгнул . Разница в интервале X обеспечивает измерение того, как накопление энергии SLP снижает минимальное напряжение разгибателя , необходимое для выполнения прыжка . Разница между двумя линиями регрессии использовалась для определения вклада SLP в прыжка на расстояния для разных напряжений разгибателей .Когда SLP был отключен, расстояние прыжка было уменьшено на 45% при 15 Н и на 55% при 10 Н. Пиковая мощность импульса прыжка при неповрежденном SLP была значительно выше, чем когда SLP был отключен (1,94 ± 0,05 мВт с, 1,10 ± 0,04 мВт без, P <10 −30 ; рис.7). Более высокая пиковая мощность также привела к значительному увеличению полной энергии для скачка (15,3 ± 0,4 мДж с, 8,58 ± 0.4 мДж без, P <10 −30 ). Кроме того, когда SLP был отключен, питание прекратилось значительно раньше, чем когда SLP был исправен (24,7 ± 0,3 мс с, 24,1 ± 0,3 мс без, P <10 −10 ).
Высокоскоростное видео ударов саранчи показало, что SLP не разворачивается до тех пор, пока большеберцовая кость не расширится более чем на 30 градусов. (Берроуз и Моррис, 2001). Чтобы проанализировать этот результат с помощью моделирования, саранча была воспроизведена саранчой .На рис. 8A показано, как деформация вращения SLP и FT вращения шарнира изменяется со временем во время скачка . Закрашенными черными квадратами отмечены значения деформации SLP с интервалами в 1 мс, как если бы они были записаны высокоскоростной видеокамерой, работающей с частотой 1000 кадров с -1 , как показано на рис. 3B Берроуза и Морриса (Берроуз и Моррис, 2001). Значительное разворачивание SLP сначала будет обнаружено через 2 мс после начала скачка при фотографировании на 1000 кадров с -1 (рис.8A, B, пунктирная линия i ), когда угол поворота FT достиг 38,1 градуса.
Чтобы понять, что вызвало эту задержку, полезно взглянуть на крутящий момент SLP , создаваемый силой SLP относительно насадки разгибателя (рис. 8B). Усилие SLP прикладывается к суставу FT , и поэтому не создается прямого крутящего момента вокруг этого сустава , но он создает крутящий момент на голени вокруг крепления разгибателя . SLP крутящий момент выполняет двойную роль: он удерживает ногу согнутой до тех пор, пока нога не начнется, когда она меняет знак и ускоряет разгибание ноги . Это изменение знака происходит в результате изменений в конфигурации шарнира FT во время разгибания ветви (рис. 8; ролик 1 в дополнительном материале). Время, необходимое для смены знака, учитывает задержку.
Таблица 1.Сравнение реальных и смоделированных прыжков саранчи
Инжир.6.Прыжки. (A) Прыжки были выполнены с полулунным процессом и без него ( SLP ) для множества различных значений натяжения разгибателей . Напряжение Extensor было изменено путем изменения времени между всплесками FETi от 10 мс до 23 мс с интервалами 1 мс. На каждом временном интервале было выполнено двадцать прыжка . Расстояние прыжка изменялось линейно с напряжением разгибателя для обоих случаев ( R 2 = 0.96 с SLP , R 2 = 0,95 без SLP ). Без SLP расстояние прыжка было сокращено на 45% при 15 Н, и это сокращение увеличилось до 55% при 10 Н. (B) Расстояние прыжка также линейно изменялось с энергией прыжка и соответствовало предсказал прыжка на расстояние ( R 2 = 0,98). Это согласуется с предполагаемой взаимосвязью между энергией прыжка и расстоянием (Bennet-Clark, 1975).
Изменение знака крутящего момента SLP произошло в результате изменения положения разгибателя относительно вектора силы SLP . В начале толчка , когда голень все еще согнута, вектор силы SLP был направлен выше крепления разгибателя , и поэтому крутящий момент SLP был отрицательным и действовал, чтобы удерживать ногу изогнутый (рис. 8B, C и ).Более того, напряжение в аподеме разгибателя поддерживало напряжение SLP . Когда отрицательный крутящий момент, создаваемый сгибающей мышцей на суставе FT , был устранен путем торможения, положительный крутящий момент, создаваемый разгибающей мышцей на суставе FT, начал разгибать ногу . Как начальное вращение ноги , так и укорочение аподемы разгибателя уменьшили силу, приложенную к SLP , что начало уменьшать ее деформацию (рис.8A, между i и ii ). Крепление разгибателя протягивалось через вектор силы SLP , в результате отрицательный (сгибающий) крутящий момент SLP уменьшался и изменялся на положительный (разгибающий) крутящий момент (рис. 8C ii и iii ). Во время этого перехода (фиг. 8A, пунктирная линия ii ) деформация в SLP снизилась менее чем на 10%. После того, как крутящий момент SLP стал положительным, он усилил вращение бедренно-большеберцового сустава , а натяжение разгибателя больше не могло поддерживать деформацию SLP , которая затем быстро уменьшалась (рис.8A iii –C iii ).
Отрицательный крутящий момент SLP в начале прыжка помогает сгибающей мышце поддерживать согнутое положение голени . Чтобы определить объем помощи, оказываемой SLP , мы сравнили величину натяжения сгибателя , необходимого для сохранения сгибания ноги , когда пружина SLP была цела, и когда она была отключена. При неповрежденной пружине SLP , сгибающая мышца удерживала голень полностью согнутой, пока натяжение сгибателя не упало до 0.26 Н. Когда пружина SLP была отключена, минимальное натяжение сгибателя , необходимое для удержания ноги от разгибания, составляло 0,56 Н. При наличии пружины SLP требовалось на 54% меньше напряжения в мышце сгибателя . , чтобы ножка не выдвигалась, когда блокировка голени не была активна.
ОБСУЖДЕНИЕ
Моторная программа и прыжки
Нейронный контроль и биомеханика пинания саранчи были хорошо описаны, потому что пинок можно вызвать, пока саранча рассекается и сдерживается; таким образом, позволяя одновременно делать электромиограммы мышц (ЭМГ) и внутриклеточные записи от центральных нейронов.Хотя в настоящее время невозможно сделать эти записи у прыгающей саранчи, сходство между толчком и прыжком привело к предположению, что одна и та же двигательная программа может вызывать оба поведения. Вычислительная нейромеханическая модель саранчи позволила нам определить, достаточны ли нейронная цепь, двигательная программа и биомеханическая конфигурация ног саранчи для учета прыжка, а также удара. Наши результаты демонстрируют, что программа толчковой моторики способна воспроизвести полный диапазон прыжкового поведения, описанного в литературе, и что контроль ключевой переменной напряжения разгибателей дает саранче возможность изменять важные характеристики прыжка, такие как расстояние прыжка. и скорость взлета.Кроме того, мы также смогли убедиться, что энергия прыжка и угол взлета являются хорошими предикторами конечной дистанции прыжка, когда саранча взлетает с минимальным кувырком. Эти результаты убедительно подтверждают гипотезу о том, что программа толчковой моторики используется как для ударов ногой, так и для прыжков.
Рис. 7.Скачок мощности с и без полулунного процесса ( SLP ). Было использовано только разгибателя, натяжения в пределах 0,25 Н от 15 Н. Перейти на мощности с неповрежденным SLP (черный) и с выключенным SLP (серый).Наблюдается значительная разница в величине пиковой мощности (1,94 ± 0,05 мВт с, 1,10 ± 0,04 мВт без, P <10 −30 ), полной энергии во время скачка импульса (15,3 ± 0,4 мДж с, 8,58 ± 0,4 мДж без, P <10 −30 ) и длительностью импульса (24,7 ± 0,3 мс с, 24,1 ± 0,3 мс без, P <10 −10 ).
Рис. 8.Полулунный процесс ( SLP ) перемещение и крутящий момент во время толчка .До удара большеберцовая кость была согнута, натяжение разгибателя составляло 15 Н, крутящий момент SLP составлял -4,7 мН м -1 , а деформация SLP составляла почти 4,5 мм. (A) бедро-большеберцовая кость ( FT ) сустав вращался с максимальной скоростью 63 градуса. м с −1 и достиг полного раскрытия 160 град. через 4 мс (пунктирная серая линия, правая ось). Деформация SLP быстро уменьшалась во время толчка (черная линия, левая ось).Заштрихованные черные квадраты представляют позиции SLP с интервалами времени 1 мс, когда высокоскоростная камера со скоростью 1000 кадров с -1 будет делать изображения [сравните с рис. 3B Берроуза и Морриса (Берроуз и Моррис, 2001)]. Начало удара показано пунктирной вертикальной линией ( i ). Первая точка, где заметное уменьшение SLP будет видно на 1000 кадров с -1 , показана пунктирной вертикальной линией ( iii ), что соответствует повороту на FT на 38.12 град. (B) Крутящий момент SLP вокруг точки прикрепления разгибателя (черная линия, левая ось) был изначально отрицательным, что помогло сохранить сгибание ноги . Значительное перемещение SLP и разгибание ноги произошло после того, как крутящий момент стал положительным, на пунктирной вертикальной линии ( ii ). (C) Отрицательный крутящий момент SLP возник, когда сила, приложенная к SLP , вызвала крутящие моменты, которые замедляли поворот голени на ( i ), в то время как положительный крутящий момент усиливал поворот голени на .Положительный крутящий момент возник только после того, как нога повернулась достаточно, чтобы переместить точку прикрепления разгибателя к противоположной стороне вектора силы SLP ( iii ). Светло-серый шарнир — это положение шарнира FT в то время, а черный шарнир — это положение шарнира FT в начале выступа . Рисунки в части C предназначены только для иллюстративных целей и не в масштабе.
Роль SLP
Прыжок является важным поведением саранчи как для передвижения, так и для спасения от хищников.SLP — это эволюционная адаптация, которая, как считалось, позволяет саранче прыгать значительно дальше, чем без нее. Однако из-за биомеханики заднегрудиального фиброзного сустава прямые тесты влияния SLP на прыжковые характеристики живой саранчи затруднены или невозможны. Представленная здесь нейромеханическая модель саранчи позволила нам определить вклад SLP в скачок. Моделирование показывает, что SLP может помочь сохранить ногу согнутой до тех пор, пока вращение большеберцовой кости не изменит знак крутящего момента SLP в пользу разгибания.Затем SLP увеличивает мощность прыжка примерно вдвое.
Основная роль мышцы-сгибателя метаторцической ноги во время прыжков и ударов ногами заключается в том, чтобы удерживать большеберцовую кость в полностью согнутом положении при одновременном сокращении. Сгибающая мышца должна быть достаточно большой, чтобы выполнять эту роль, но любой избыточный объем, посвященный сгибающей мышце, является расточительным, и этот объем лучше использовать разгибающей мышце для обеспечения большей мощности для прыжка. Мы обнаружили, что SLP помогает поддерживать сгибание ноги во время совместного сокращения и, таким образом, обеспечивает момент сгибания, который в противном случае должен был бы обеспечивать сгибатель .Когда пружина SLP была отключена, любой создаваемый ею момент сгибания был снят, и требовалось на 54% больше натяжения сгибателя , чтобы не дать большеберцовой кости разгибаться. Мы предполагаем, что наличие SLP позволяет мышце-сгибателю быть меньше, чем было бы в противном случае, и что экономия в объеме большеберцовой кости может позволить относительно более крупную мышцу-разгибатель и более мощный прыжок.
Высокоскоростная видеозапись борьбы саранчи показала, что разворачивание SLP задерживается до тех пор, пока большеберцовая кость не повернется более чем на 30 градусов.(Берроуз и Моррис, 2001). Модель саранчи помогает объяснить, почему происходит такая задержка в развертывании SLP и как это влияет на прыжковые способности саранчи. Как показано на рис.8, задержка, по-видимому, зависит от движения точки прикрепления разгибателя большеберцовой кости через вектор силы SLP во время начального периода разгибания ноги, изменяя крутящий момент SLP , вызвавший сгибание, на тот, который продвигает удлинение ноги . Наконец, моделирование показывает, что SLP позволяет саранче с по прыгнуть на более чем на 45% дальше, чем без него, в зависимости от силы, прикладываемой разгибателем голени .
Накопление энергии за счет деформации кутикулы или деформации в аподемах — распространенные методы преодоления ограничений скелетных мышц, которые необходимы членистоногим для быстрых движений, таких как прыжки и щелчки (Gronenberg, 1996). Некоторые из этих животных развили сложную специализацию, позволяющую им выполнять эти быстрые движения. Кроличья блоха Spilopsyllus cuniculus накапливает энергию для своего прыжка в резиновой подушечке во внутреннем скелете грудной клетки, но высвобождается, когда вектор силы проходит через сустав ноги и перенаправляется от сгибания ноги к разгибанию, как SLP саранчи ( Беннет-Кларк и Люси, 1967).Froghopper Philaenus spumarius накапливает энергию для прыжка, сгибая дугообразное образование кутикулы в плевральной дуге. Система блокировки трения не позволяет ногам двигаться до тех пор, пока напряжение в депрессорах не превысит удерживающую силу замка (Burrows, 2003; Burrows, 2006; Burrows et al., 2008). И муравей-ловушка Odontomachus bauri , и креветка-щелкун Alpheus californiensis накапливают энергию для быстрого закрытия своей нижней челюсти или когтя в аподеме мышцы и за счет деформации кутикулы экзоскелета (Gronenberg, 1995a; Gronenberg). , 1995b; Ритцманн, 1973).Креветка Alpheus heterochelis имеет механизм, похожий на механизм блохи, где блокировка доводчика происходит, когда вектор силы перемещается выше точки поворота, а отпускание происходит, когда вспомогательная мышца доводчика тянет его назад, чтобы создать большой закрывающий момент ( Рицманн, 1974).
Хотя все эти механизмы схожи, есть различие, которое кажется уникальным для SLP-механизма саранчи. Энергия прыжка в основном исходит от аподема разгибателя и SLP.У саранчи SLP и аподема создают силы, которые действуют в разных местах и в разных направлениях, и это ключевая особенность, вызывающая задержку развертывания SLP. Механизмы накопления энергии, используемые блохами (Bennet-Clark and Lucey, 1967) и креветками (Ritzmann, 1974), имеют сходства и различия с системой, описанной здесь. Они похожи в том, что движение вектора силы вызывает разворот в направлении крутящего момента. У креветок это происходит, когда более близкая мышца протягивается от одной стороны сустава к другой.У саранчи это происходит, когда аподема разгибателя протягивается через вектор силы SLP . Разница заключается в том, что креветка и блоха используют небольшую вспомогательную мышцу, чтобы тянуть главную сократительную мышцу над суставом, тогда как у саранчи именно биомеханика движения сустава вызывает изменение крутящего момента SLP на противоположное.
Заключение
Эта работа демонстрирует полезность нейромеханического моделирования для анализа взаимодействия нейронных механизмов управления с биомеханическими процессами, которые опосредуют и ограничивают движения животного.Нейромеханическое моделирование позволяет понять работу нервных цепей, которые были описаны у животных, подвергнутых вскрытию, удержанию и анестезии, в чем-то подобном их естественному контексту, где последствия их активности для движения становятся очевидными.
Одной из областей будущих исследований, выявленных в ходе моделирования, является роль кувырка во время прыжка. Угол взлета можно контролировать с помощью ориентации задних ног (Sutton and Burrows, 2008). Мы отметили при моделировании, что изменение ориентации может привести к углу взлета, при котором вектор тяги не совпадает с COM Locust.В тех случаях саранча демонстрировала кувыркающееся поведение во время прыжка , которое качественно отличалось от поведения живых животных, прыгающих с аналогичными углами взлета. Мы полагаем, что у саранчи может быть несколько механизмов предотвращения нежелательного кувырка при разных углах взлета, в том числе установка начального подъема тела, изменение положения тела во время взлета и контроль передачи импульса между ногами и телом. Модель саранчи может оказаться полезной при изучении этих механизмов и их важности.
Вторая область — это роль референции в управлении прыжком. Направление и расстояние прыжка контролируются исходной позой животного и натяжением сухожилия разгибателя (Santer et al., 2005; Sutton and Burrows, 2008). Было идентифицировано несколько наборов проприоцепторов, которые предоставляют сенсорную информацию, полезную для этого контроля, и описаны роли, которые некоторые играют во время ударов ногами (Burrows and Pfluger, 1988; Heitler, 1977; Heitler, 1995). Однако неясно, как реафферентные ответы или их использование могут быть изменены для прыжков, когда и условия нагрузки, и ограничения, налагаемые начальной настройкой, различны.Учитывая большие трудности в экспериментальном решении этих вопросов, может оказаться полезным моделирование реафферентных ответов в условиях замкнутого цикла, которые приводят к скачку.
.