
I. СКЕЛЕТНАЯ МЫШЕЧНАЯ ТКАНЬ — Med24info.com
У человека более 600 скелетных мышц (около 40% массы тела). Скелетная мышечная ткань обеспечивает осознанные и осознаваемые произвольные движения тела и его частей. Основные гистологические элементы: скелетные мышечные волокна (функция сокращения), клетки-сателлиты (камбиальный резерв).
А. Развитие скелетных мышц. Источник развития гистологических элементов скелетной мышечной ткани — миотомы, откуда выселяются и мигрируют в места закладки конкретных мышц самые ранние клетки миогенного клеточного типа — клетки миотомов. Миогенный клеточный тип в эмбриогенезе последовательно складывается из следующих гистологических элементов: клетки миотома (миграция) -»• миобласты митотические (пролиферация) -gt; миобласты постмитотические (слияние) -gt; мышечные трубочки (синтез сократительных белков, формирование саркомеров) -» мышечные волокна (функция сокращения). В области закладки мышц уже присутствуют клетки мезенхимы — источник соединительнотканных структур мышцы, сюда прорастают кровеносные капилляры, а позднее (при образовании мышечных трубочек) — аксоны двигательных и чувствительных нейронов соматического отдела нервной системы.

- Клетки миотомов при их выселении из сомитов уже детерминированы в направлении образования миогенных элементов. Дефекты миграции клеток миотома, а также сбои при взаимодействии разных клеточных элементов в ходе миогенеза приводят к аномальному развитию скелетных мышц.
- Миобласты
а. Митотические миобласты (Gj-миобласты) последовательно проходят ряд завершающихся митозами клеточных циклов (пролиферативные митозы). На этой стадии часть
- -миобластов обособляется в виде клеток-сателлитов.
б. Постмитотические миобласты (Gg-миобласты) — клетки, необратимо вышедшие из клеточного цикла (результат квантального митоза) и уже начавшие синтез сократительных белков. Gg-миобласты сливаются и образуют симпласты — миотубы.
- Мышечная трубочка (миотуба) — цепочка слившихся миобластов, в которой ядра занимают центральное положение. В мышечных трубочках происходит терминальная миогенная дифференцировка: синтез контрактильных белков, сборка сократительных структур — миофибрилл (появляется поперечная исчерченность).
 Перемещение ядер симпласта на периферию завершает формирование поперечнополосатого мышечного волокна.
Перемещение ядер симпласта на периферию завершает формирование поперечнополосатого мышечного волокна. - Мышечное волокно — дефинитивная форма скелетномышечного миогенеза, выполняющая функцию мышечного сокращения (см. I Б). Разные типы мышечных волокон рассмотрены в I Д, их фенотипы — в I Е.
- Клетки-сателлиты — обособившиеся в ходе миогенеза 0,-миобласты, расположенные между базальной мембраной и плазмолеммой мышечных волокон. Ядра этих клеток составляют 10% суммарного количества ядер скелетного мышечного волокна. Клетки- сателлиты — камбиальный резерв мышечной ткани скелетного типа. Они сохраняют способность к миогенной дифференцировке (миобласты -» миотубы -gt; мышечные волокна) в течение всей жизни, что обеспечивает увеличение массы мышечных волокон. Клетки-сателлиты также участвуют в репаративной регенерации скелетной мышечной ткани (см. I Ж).
- Маркёры миогенного клеточного типа.
а. Миогенные факторы (в т.
 ч. миогенин и MyoD) — регуляторные факторы миогенеза, экспрессируются только в гистологических элементах скелетной мышцы на разных стадиях миогенеза. Миогенин и MyoD — факторы транскрипции специфичных для миогенного клеточного типа генов.
ч. миогенин и MyoD) — регуляторные факторы миогенеза, экспрессируются только в гистологических элементах скелетной мышцы на разных стадиях миогенеза. Миогенин и MyoD — факторы транскрипции специфичных для миогенного клеточного типа генов. б. Десмин — мышечноспецифический белок класса промежуточных филаментов цитоскелета. Десмин найден в гистологических элементах всех мышечных тканей. В скелетномышечном миогенезе десмин начинает экспрессироваться в цитоплазме миобластов, а с началом миофибриллогенеза десминовые нити вплетаются в Z-диски или их аналоги в ГМК (плотные пятна). При различных миопатиях происходит накопление десмина в саркоплазме.
в. Контрактильные белки актомиозинового хемомеханического преобразователя — миозины, актины, тропонины, тропомиозины, а также вспомогательные белки тонких (актиновых), толстых (миозиновых) нитей миофибрилл и Z-дисков.
г. Креатинфосфокиназа (КФК)
д. Миоглобин
Б.
 Скелетное мышечное волокно
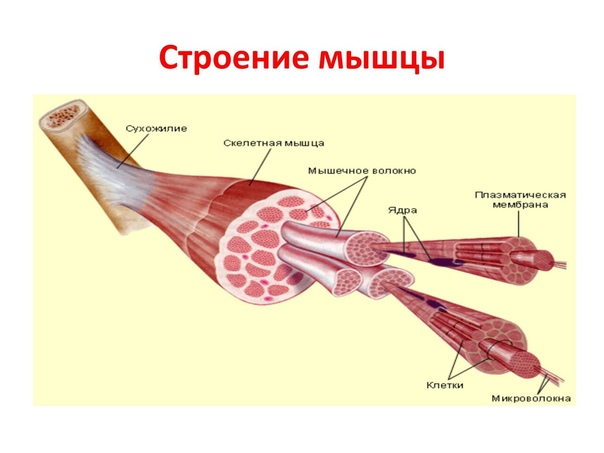
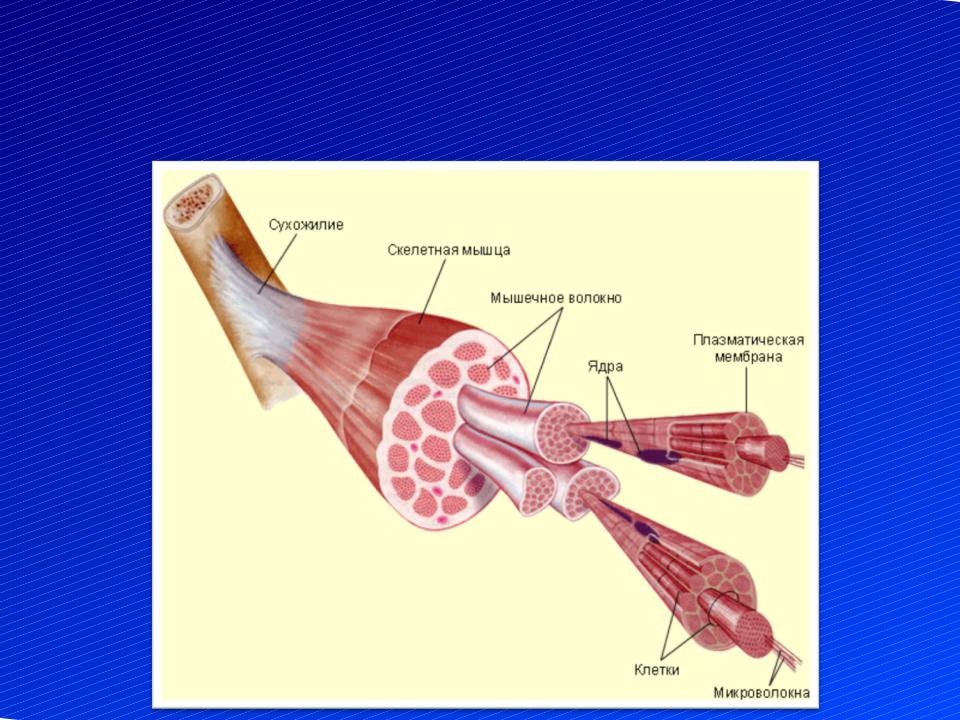
Скелетное мышечное волокно- Общая характеристика. Структурно-функциональная единица скелетной мышцы — симпласт — скелетное мышечное волокно (рис. 7-1, 7-16), имеет форму протяжённого цилиндра с заострёнными концами. Этот цилиндр достигает в длину 40 мм при диаметре до 0,1 мм. Термин оболочка волокна (сарколемма) относят к двум структурам: плазмолемма симпласта и его базальная мембрана. Между плазмолеммой и базальной мембраной расположены овальной формы ядра клеток-сателлитов. Палочковидной формы ядра мышечного волокна лежат в миоплазме (саркоплазма) под плазмолеммой. В саркоплазме симпласта расположены контрактильный аппарат — миофибриллы, депо Ca2+ — саркоплазматическая сеть, энергетические станции — митохондрии, включения (гранулы гликогена). От поверхности мышечного волокна к расширенным участкам саркоплазматического ретикулума направляются трубковидные впячивания сарколеммы — поперечные трубочки (Т-трубочки). Рыхлая волокнистая
Рис. 7-1. Скелетная мышца состоит из поперечнополосатых мышечных волокон.
 Значительный объём волокон занимают миофибриллы. Расположение светлых и тёмных дисков в параллельных друг другу миофибриллах совпадает, что приводит к появлению поперечной исчерченности. Структурная единица миофибрилл — саркомер, сформированный из толстых и тонких нитей [из Bloom W FawsettDW, 1968]
Значительный объём волокон занимают миофибриллы. Расположение светлых и тёмных дисков в параллельных друг другу миофибриллах совпадает, что приводит к появлению поперечной исчерченности. Структурная единица миофибрилл — саркомер, сформированный из толстых и тонких нитей [из Bloom W FawsettDW, 1968]
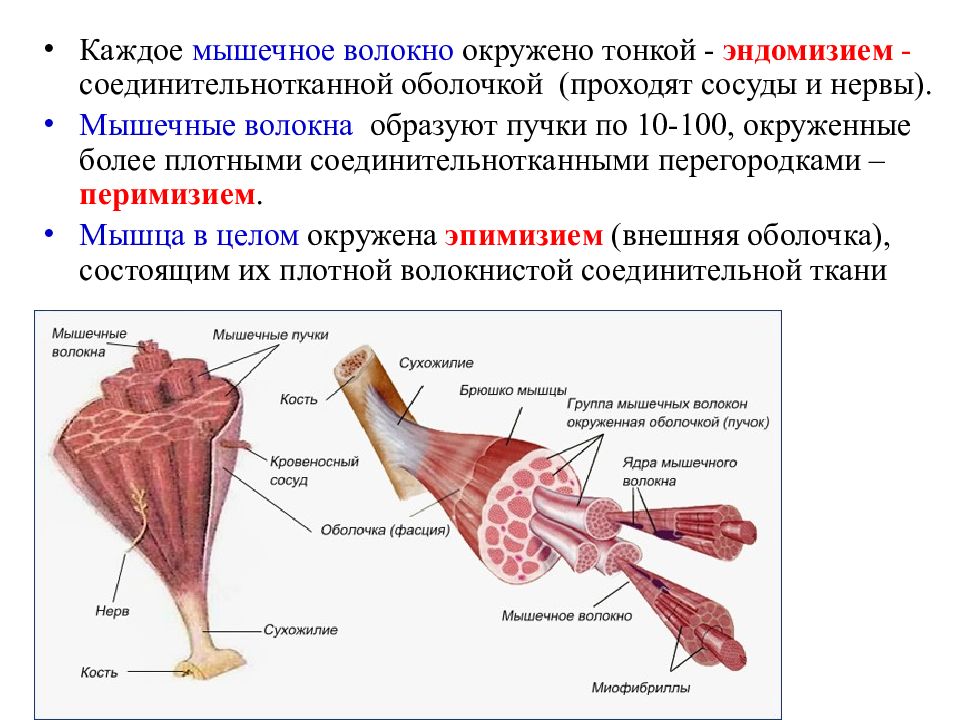
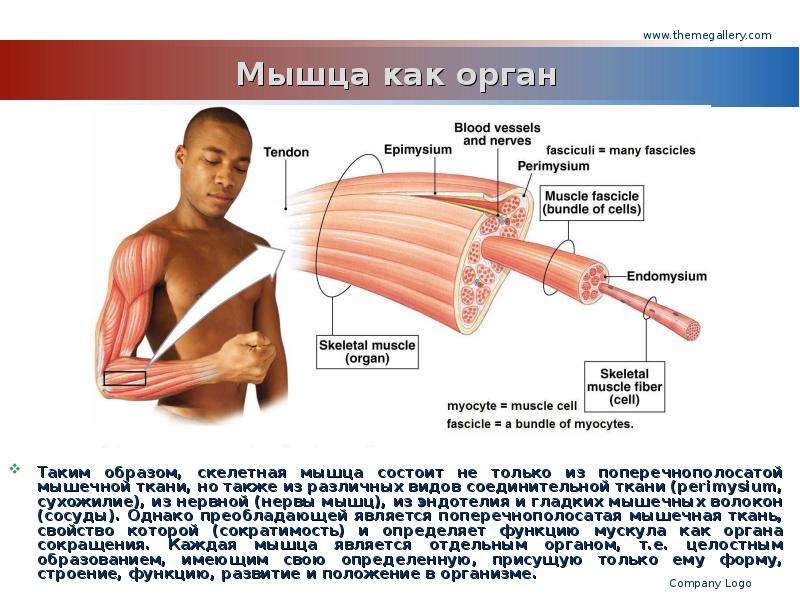
соединительная ткань между отдельными мышечными волокнами (эндомизий) содержит кровеносные и лимфатические сосуды, нервные волокна. Группы мышечных волокон и окружающая их в виде чехла волокнистая соединительная ткань (перимизий) формируют пучки. Их совокупность образует мышцу, плотный соединительнотканный чехол которой именуют эпимизий.
- Миофибриллы. Поперечная исчерченность скелетного мышечного волокна определяется регулярным чередованием в миофибриллах различно преломляющих поляризованный свет участков (дисков) — изотропных и анизотропных: светлые (Isotropic, 1-диски) и тёмные (Anisotropic, A-диски) диски. Разное светопреломление дисков определяется упорядоченным расположением по длине саркомера тонких и толстых нитей; толстые нити находятся только в тёмных дисках, светлые диски не содержат толстых нитей.

- Саркомер (рис. 7-1, 7-8) — структурно-функциональная единица миофибриллы, расположенная между соседними Z-линиями. Саркомер образуют расположенные параллельно друг другу тонкие (актиновые) и толстые (миозиновые) нити. I-диск содержит только тонкие нити. В середине I-диска проходит Z-линия. Один конец тонкой нити прикреплён к Z-линии, а другой конец направлен к середине саркомера. Толстые нити занимают центральную часть саркомера — A-диск. Тонкие нити частично входят между толстыми. Содержащий только толстые нити участок саркомера — Н-зона. В середине Н-зоны проходит М-линия. I-диск входит в состав двух саркомеров. Следовательно, каждый саркомер содержит один A-диск (тёмный) и две половины I-диска (светлого), формула саркомера — Ч А + I + Ч А.
а. Толстая нить. Каждая миозиновая нить состоит из 300-400 молекул миозина и С-белка.
 Миозин — гексамер (две тяжёлые и четыре лёгкие цепи). Тяжёлые цепи — две спирально закрученные полипептидные нити, несущие на своих концах глобулярные головки. В области головок с тяжёлыми цепями ассоциированы лёгкие цепи. Каждую миозиновую нить связывает с Z-линией гигантский белок титин.
Миозин — гексамер (две тяжёлые и четыре лёгкие цепи). Тяжёлые цепи — две спирально закрученные полипептидные нити, несущие на своих концах глобулярные головки. В области головок с тяжёлыми цепями ассоциированы лёгкие цепи. Каждую миозиновую нить связывает с Z-линией гигантский белок титин.- Миозин (рис. 7-2). В молекуле миозина различают тяжёлый и лёгкий меромио- зин. Тяжёлый меромиозин содержит субфрагменты (S): S1 содержит глобулярные головки миозина, S2 — прилежащую к головкам часть фибриллярного хвоста молекулы миозина. S2 эластичен (эластический компонент S2), что допускает отхождение S1 на расстояние до 55 нм. Концевую часть хвостовой нити миозина длиной 100 нм образует лёгкий меромиозин. Миозин имеет два шарнирных участка, позволяющих молекуле изменять конформацию. Один шарнирный участок находится в области соединения тяжёлого и лёгкого меромиозинов, другой — в области шейки молекулы миозина (S1-S2 соединение). Половина молекул миозина обращена головками к одному концу нити, а вторая половина — к другому (рис.

Рис. 7-2. Молекула миозина. Лёгкий меромиозин обеспечивает агрегацию молекул миозина, тяжёлый меромиозин имеет связывающие актин участки и
Г оловки миозина
Рис. 7-3. Толстая нить. Молекулы миозина способны к самосборке и формируют веретенообразный агрегат диаметром 15 нм и длиной 1,5 мкм. Фибриллярные хвосты молекул образуют стержень толстой нити, головки миозина расположены спиралями и выступают над поверхностью толстой нити [из Ham AW, Cormack DH, 1979]
Рис. 7-4. Тонкая нить — две спирально скрученные нити F-актина. В канавках спиральной цепочки залегает двойная спираль тропомиозина, вдоль которой располагаются молекулы тропонина [из Jun- queira LC, CameiroJ, 1991]
- Титин — наибольших размеров полипептид (из известных) с Mr 3000 кД — наподобие пружины связывает концы толстых нитей с Z-линией.
 Другой гигантский белок — небулин (Mr 800 кД) — ассоциирует тонкие и толстые нити.
Другой гигантский белок — небулин (Mr 800 кД) — ассоциирует тонкие и толстые нити. - С-белок стабилизирует структуру миозиновых нитей. Влияя на агрегацию молекул миозина, обеспечивает одинаковый диаметр и стандартную длину толстых нитей.
- Миомезин (М-белок), КФК — белки, ассоциированные с толстыми нитями в середине тёмного диска. КФК способствует быстрому восстановлению АТФ при сокращении. Миомезин выполняет организующую роль при сборке толстых нитей.
б. Тонкая нить состоит из актина, тропомиозина и тропонинов (рис. 7-4).
- Актин. Молекулы глобулярного актина (G-актин) полимеризуются и образуют фибриллярный актин (F-актин). В состав тонкой нити входят две спирально закрученные цепочки F-актина.
- Тропомиозин состоит из двух полипептидных цепей и имеет конфигурацию двойной спирали. Полярные молекулы тропомиозина длиной 40 нм укладываются конец в конец в желобке между двумя спирально закрученными цепочками F-актина.
- Тропонин (Tn) — комплекс, образованный тремя глобулярными CE: TnT1 TnI1 TnC. TnT имеет участки для связи с тропомиозином. TnC — Са2+-связывающий белок. TnI препятствует взаимодействию актина с миозином. Тропониновый комплекс прикреплён к молекулам тропомиозина с интервалами 40 нм.
- а-Актинин, десмин и виментин — белки, ассоциированные с тонкими нитями; они входят в состав Z-линии, в которую вплетён один конец тонкой нити; второй конец расположен между толстыми нитями и ассоциирован с ними при помощи небулина.
- Саркоплазматнческая сеть. Каждая миофибрилла окружена регулярно повторяющимися по её длине (точнее, по длине саркомера) элементами саркоплазматического ретикулума — анастомозирующими мембранными трубочками, заканчивающимися терминальными цистернами (рис. 7-5). На границе между дисками А и I две терминальные цистерны соседних повторов контактируют с Т-трубочками (триады).
 Саркоплазмати- ческий ретикулум — модифицированная гладкая эндоплазматическая сеть, выполняющая функцию депо кальция. Са2+-транспортирующие АТФазы саркоплазматического ретикулума откачивают ионы кальция из саркоплазмы. Са2*-связывающий белок кальсеквестрин находится внутри саркоплазматической сети. Кальциевые каналы, образованные рецепторами рианодина, высвобождают Ca2+ из депо в саркоплазму.
Саркоплазмати- ческий ретикулум — модифицированная гладкая эндоплазматическая сеть, выполняющая функцию депо кальция. Са2+-транспортирующие АТФазы саркоплазматического ретикулума откачивают ионы кальция из саркоплазмы. Са2*-связывающий белок кальсеквестрин находится внутри саркоплазматической сети. Кальциевые каналы, образованные рецепторами рианодина, высвобождают Ca2+ из депо в саркоплазму. - Т-трубочки. Сарколемма мышечного волокна образует множество узких впячиваний — поперечных трубочек (Т-трубочки). Они проникают внутрь мышечного волокна и, залегая между двумя терминальными цистернами саркоплазматического ретикулума, вместе с последними формируют триады. В триадах происходит передача возбуждения в виде потенциала действия плазматической мембраны мышечного волокна на мембрану терминальных цистерн — сопряжение возбуждения и сокращения: дигидропиридино- вые рецепторы Т-трубочек регистрируют изменения мембранного потенциала (дУ) и активируют рианодиновые рецепторы саркоплазматической сети (Са2+-канал).
В. Иннервация. Двигательная и чувствительная соматическая иннервация скелетных мышц (мышечных волокон) осуществляется соответственно а- и у-мотонейронами передних рогов спинного мозга и двигательных ядер черепных нервов и псевдоуниполярными чувствительными нейронами спинномозговых узлов и чувствительных ядер черепных нервов. Вегетативная иннервация гистологических элементов скелетных мышц не обнаружена, но ГМК стенки кровеносных сосудов мышц имеют симпатическую адренергическую иннервацию.
- Двигательная иннервация. В скелетных мышцах соответственно характеру иннервации принято различать два типа мышечных волокон: экстрафузальные и интрафузаль- ные. Каждое экстрафузальное мышечное волокно имеет прямую двигательную иннервацию — нервно-мышечные синапсы, образованные терминальными ветвлениями аксонов а-мотонейронов и специализированными участками плазмолеммы мышечного волокна (концевая пластинка, постсинаптическая мембрана). Интрафузальные мышечные волокна (см.
 главу 8.2 III А 8 а) образуют нервно-мышечные синапсы с эфферентными волокнами у-мотонейронов. Мышечные волокна входят в состав нейро- моторных (двигательных) единиц и обеспечивают сократительную функцию мышц.
главу 8.2 III А 8 а) образуют нервно-мышечные синапсы с эфферентными волокнами у-мотонейронов. Мышечные волокна входят в состав нейро- моторных (двигательных) единиц и обеспечивают сократительную функцию мышц.
а. Нейромоторная единица включает один мотонейрон и группу иннервируемых им экстрафузальных мышечных волокон. Количество и размеры двигательных единиц в различных мышцах значительно варьируют. Поскольку при сокращении фазные мышечные волокна подчиняются закону всё или ничего, то сила, развиваемая мышцей, зависит от количества активируемых (т.е. участвующих в сокращении мышечных волокон) двигательных единиц. Каждая нейромоторная единица образована только быстросокращающимися или только медленносокращающимися мышечными волокнами (см. I Д I б).
б. Полинейронная иннервация. Формирование нейромоторных единиц происходит в постнатальном периоде, а до рождения мышечные волокна имеют полинейронную иннервацию, когда каждое мышечное волокно иннервируется несколькими мотонейронами.
 Аналогичная ситуация возникает при денервации (например, при повреждении
Аналогичная ситуация возникает при денервации (например, при повреждении Рис. 7-5. Фрагмент скелетного мышечного волокна. Цистерны саркоплазматического ретику- лума окружают каждую миофибриллу. Т-трубочки подходят к миофибриллам на уровне границ между тёмными и светлыми дисками и вместе с терминальными цистернами саркоплазматического ретихулума образуют триады. Между миофибриллами залегают митохондрии (из Kopf-Maier Р, Merker H-J, 1989]
нерва) с последующей реиннервацией мышечных волокон. Понятно, что в этих ситуациях страдает эффективность сократительной функции мышцы.
- Чувствительная иннервация
а. Интрафузальные мышечные волокна вместе с чувствительными нервными окончаниями формируют мышечные веретёна — рецепторы скелетной мышцы (см. главу 8 2 III А 8 а).
б. Сухожильный орган Гольджи (глава 8.2 III А 8 б)
Г. Сокращение мышцы происходит при поступлении по аксонам двигательных нейронов к нервно-мышечным синапсам волны возбуждения в виде нервных импульсов (потенциалы действия нервных волокон).
 Это непрямое сокращение (опосредованное нервно-мышечной синаптической передачей возбуждения). Возможно и прямое сокращение мышцы. Под ним понимают сокращение групп мышечных волокон, происходящее при возбуждении любого звена последовательности событий после секреции нейромедиатора из термина- лей аксона в нервно-мышечном синапсе (см. главу 8.2 III Б I). Прямое сокращение мышцы (например, мышечные подёргивания, или фибрилляции) — всегда патология.
Это непрямое сокращение (опосредованное нервно-мышечной синаптической передачей возбуждения). Возможно и прямое сокращение мышцы. Под ним понимают сокращение групп мышечных волокон, происходящее при возбуждении любого звена последовательности событий после секреции нейромедиатора из термина- лей аксона в нервно-мышечном синапсе (см. главу 8.2 III Б I). Прямое сокращение мышцы (например, мышечные подёргивания, или фибрилляции) — всегда патология.- Нервно-мышечный синапс и деполяризация постсинаптической мембраны. Возбуждение мотонейрона приводит к секреции ацетилхолина из терминалей аксона в синаптическую щель. Ацетилхолин связывается с его рецепторами (никотиновые холинорецепторы мышечного типа, н-холинорецепторы), вмонтированными в постси- наптическую мембрану мышечного волокна (плазмолемма). Результат взаимодействия — деполяризация мембраны мышечного волокна, т.к. при взаимодействии нейромедиатора с холинорецептором открывается ионный канал в составе рецептора. Взаимодействие рецептора и ацетилхолина кратковременно, т.
 к. присутствующая в синапсе аце- тилхолинэстераза гидролизует нейромедиатор. Нарушения в любом звене нервно-мышечной передачи приводят к искажениям мышечного сокращения.
к. присутствующая в синапсе аце- тилхолинэстераза гидролизует нейромедиатор. Нарушения в любом звене нервно-мышечной передачи приводят к искажениям мышечного сокращения.
а. Ботулинический и столбнячный нейротоксины, (3- и у-бунгаротоксины блокируют секрецию ацетилхолина.
б. Карбахолин усиливает секрецию ацетилхолина.
в. Миорелаксанты периферического действия (кураре и курареподобные препараты) широко применяются в анестезиологии. Тубокурарин препятствует деполяризующему действию ацетилхолина. Дитилин приводит к миопаралитическому эффекту, вызывая стойкую деполяризацию постсинаптической мембраны.
г. Денервация. При двигательной денервации происходит значительное увеличение чувствительности мышечных волокон к эффектам ацетилхолина вследствие увеличенного синтеза рецепторов ацетилхолина и их встраивания в плазмолемму по всей поверхности мышечного волокна.
д. Фосфорорганические соединения (ФОС), прозерин, физостигмин, галан- тамин инактивируют ацетилхолинэстеразу.

е. Миастения тяжёлая псевдопаралитическая (myasthenia gravis). При этом заболевании циркулирующие в крови AT к холинорецепторам угнетают их функцию (страдает нервно-мышечная передача, развивается мышечная слабость).
ж. а-Бунгаротоксин блокирует холинорецепторы.
- Плазмолемма и потенциал действия. Локальная деполяризация постсинаптической мембраны приводит к генерации потенциала действия, быстро распространяющегося по всей плазмолемме мышечного волокна (включая Т-трубочки).
а. Электромиография — важный диагностический метод — позволяет регистрировать характеристики потенциалов действия.
б. Ионные каналы, обеспечивающие развитие потенциала действия, рассмотрены в главе 2 I В I б (2).
в. Миотония. Уменьшение СГ-проводимости плазмолеммы ведёт к электрической нестабильности мембраны мышечных волокон и развитию миотонии (например, болезни Томсена).

- Триады и передача сигнала на саркоплазматическую сеть. Волна деполяризации по Т-трубочкам проникает до триад. В области триад мембрана Т-трубочек в составе потенциалзависимого кальциевого канала содержит рецепторы дигидропиридина. Деполяризация мембраны Т-трубочек вызывает в структуре рецепторов дигидропиридина конформационные изменения, передающиеся на рецепторы рианодина терминальных цистерн саркоплазматического ретикулума.
Злокачественная гипертермия при наркозе (особенно при использовании тиопентала и гало- тана) — редкое осложнение (смертность до 70%) при хирургическом вмешательстве. Температура тела быстро поднимается до 43 °С и выше, происходит генерализованный рабдомиолиз. В части случаев найдена мутация гена рианодинового рецептора скелетномышечного типа.
- Саркоплазматический ретикулум и выброс Ca2*. Активация рецепторов рианодина (Са2+-канал) приводит к открытию Са2+-каналов, Ca2+ из Са2+-депо (саркоплазматичес- кая сеть) поступает в саркоплазму; концентрация Ca2+ в саркоплазме достигает значений, достаточных для связывания этого двухвалентного катиона с тропонином С (TnC) тонких нитей.

- Связывание Ca2+ тонкими нитями. В покое взаимодействие тонких и толстых нитей невозможно, т.к. миозинсвязывающие участки молекул актина заблокированы тропоми- озином. При высокой концентрации Ca2+ эти ионы связываются с TnC и вызывают конформационные изменения тропомиозина, приводящие к разблокированию миозинсвязы- вающих участков (рис. 7-6).
- Взаимодействие тонких и толстых нитей. В результате разблокирования мио- зинсвязывающих участков молекул актина головки миозина присоединяются к тонкой нити и изменяют свою конформацию, создавая тянущее усилие — тонкие нити начинают скользить между толстыми.
Шарнирные
Рис. 7-6. Са2*-зависнмый механизм регуляции взаимодействия актина с миозином. В покое миозинсвязывающие участки тонкой нити заняты тропомиозином. При сокращении ионы Cazt связываются с ТпС, а тропомиозин открывает миозинсвязывающие участки. Головки миозина присоединяются к тонкой нити и вызывают её смещение относительно толстой нити [по von Ganong ЖР (1979) из Junqueira LC, Cameiro J, 1991)
Модель скользящих нитей была предложена Хью Хаксли.
 Скольжение тонких нитей относительно толстых обеспечивает чередование рабочих циклов. Каждый цикл имеет несколько стадий (рис. 7-7).
Скольжение тонких нитей относительно толстых обеспечивает чередование рабочих циклов. Каждый цикл имеет несколько стадий (рис. 7-7).- Головка миозина, несущая продукты гидролиза АТФ (АДФ + Pi), прикрепляется к миозинсвязывающим участкам актиновой нити.
- Головка миозина изменяет конформацию; за счёт шарнирного участка в области шейки миозина происходит гребковое движение, продвигающее тонкую нить к центру саркомера.
- Головка миозина связывается с молекулой АТФ, что приводит к отделению миозина от актина. Последующий гидролиз АТФ восстанавливает конформацию молекулы миозина, и она оказывается готовой вступить в новый цикл.
- Укорочение саркомера и сокращение мышечного волокна. Головка миозина совершает около пяти циклов в секунду. Когда одни головки миозина толстой нити производят тянущее усилие, другие в это время свободны и готовы вступить в очередной цикл. Следующие друг за другом гребковые движения стягивают тонкие нити к центру саркомера.
 Скользящие тонкие нити тянут за собой Z-линии, вызывая сокращение саркомера (рис. 7-8). Поскольку в процесс сокращения практически одномоментно вовлечены все саркомеры мышечного волокна, происходит его укорочение.
Скользящие тонкие нити тянут за собой Z-линии, вызывая сокращение саркомера (рис. 7-8). Поскольку в процесс сокращения практически одномоментно вовлечены все саркомеры мышечного волокна, происходит его укорочение. - Расслабление. Са2*-АТФаза саркоплазматического ретикулума закачивает Ca2+ из саркоплазмы в цистерны ретикулума, где Ca2+ связывается с кальсеквестрином. При низкой саркоплазматической концентрации Ca2* тропомиозин закрывает миозинсвязыва- ющие участки и препятствует их взаимодействию с миозином. После смерти, когда содержание АТФ в мышечных волокнах снижается вследствие прекращения её синтеза, головки миозина оказываются устойчиво прикреплёнными к тонкой нити. Это состояние трупного окоченения (rigor mortis) продолжается, пока не наступит аутолиз, после чего мышцы можно растянуть.
- Энергетические потребности. Мышечное сокращение требует значительных энергетических затрат. Основной источник энергии — гидролиз макроэрга АТФ.
а.
 Митохондрии мышечного волокна имеют удлинённую форму и располагаются в миоп- лазме параллельно миофибриллам. В митохондриях происходят процессы цикла трикар- боновых кислот и окислительного фосфорилирования, что приводит к генерации АТФ.
Митохондрии мышечного волокна имеют удлинённую форму и располагаются в миоп- лазме параллельно миофибриллам. В митохондриях происходят процессы цикла трикар- боновых кислот и окислительного фосфорилирования, что приводит к генерации АТФ. б. Гликоген запасается в саркоплазме в виде включений. Анаэробный гликолиз сопряжён с синтезом АТФ.
в. Креатинфосфокиназа, связанная в области М-линии, катализирует перенос фосфата от фосфокреатина на АДФ с образованием креатина и АТФ.
г. Миоглобнн. Этот пигментный белок, как и Hb, обратимо связывает кислород. Запасы кислорода необходимы для синтеза АТФ при длительной непрерывной работе мышцы.
Д. Классификации типов мышечных волокон и скелетных мышц. Скелетные мышцы и образующие их мышечные волокна различаются по множеству параметров: скорости сокращения, утомляемости, диаметру, цвету и т.д. Например, цвет мышцы может быть обусловлен рядом причин: количеством митохондрий, содержанием миоглобина, плотностью кровеносных капилляров.
 Традиционно выделяют красные и белые, а также медленные и быстрые мышцы и волокна. Другими словами, существует множество классификаций мышц и мышечных волокон. Каждая мышца — гетерогенная популяция разных типов мышечных волокон. Тип мышцы определяют, исходя из преобладания в ней конкретного типа мышечных волокон.
Традиционно выделяют красные и белые, а также медленные и быстрые мышцы и волокна. Другими словами, существует множество классификаций мышц и мышечных волокон. Каждая мышца — гетерогенная популяция разных типов мышечных волокон. Тип мышцы определяют, исходя из преобладания в ней конкретного типа мышечных волокон.- Критерии классификации. На практике важны следующие классифицирущие критерии типов мышечных волокон: характер сокращения (а), скорость сокращения
(б), тип окислительного обмена (в).
Рис. 7-7. Цикл скольжения тонких нитей относительно толстых. Тяжёлый меромиозин лежит на поверхности толстой нити (I). Благодаря шарнирному участку между тяжёлым и лёгким меромиози- ном, несущая АДФ и Р. головка миозина прикрепляется к актину (2), происходит поворот головки миозина с одновременным растягиванием эластического компонента S2 (3). Из головки освобождаются АДФ и Р., а последующая ретракция эластического компонелта S, вызывает тянущее усилие (4).
 Затем к головке миозина присоединяется новая молекула АТФ, что приводит к отделению головки миозина от молекулы актина. Гидролиз АТФ возвращает молекулу миозина в исходное положение (5) [из WestJB, 1990]
Затем к головке миозина присоединяется новая молекула АТФ, что приводит к отделению головки миозина от молекулы актина. Гидролиз АТФ возвращает молекулу миозина в исходное положение (5) [из WestJB, 1990] Рис. 7-8. Саркомер расслабленного (А) и сокращённого (Б) мышечного волокна. При сокращении тонкие нити движутся к центру саркомера, их свободные концы сходятся у М-линии. Вследствие этого уменьшается длина I-дисков и Н-зоны. Длина А-диска не изменяется [по Schultz Е, Lebbnd CP из Ham AW, Cormack DH, 19791
а. Фазные и тонические. Экстрафузальные мышечные волокна подразделяют на фазные, осуществляющие энергичные сокращения, и тонические, специализирующиеся на поддержании статического напряжения, или тонуса. Произвольная мускулатура человека практически полностью состоит из фазных мышечных волокон, генерирующих потенциалы действия. В ответ на нервную стимуляцию они отвечают быстрым сокращением. Тонические мышечные волокна встречаются лишь в наружных ушных и наружных глазных мышцах.
 Тонические мышечные волокна имеют более низкий потенциал покоя (от -50 до -70 мВ). Степень деполяризации мембраны зависит от частоты стимуляции, поэтому лишь повторные нервные стимулы вызывают сокращение тонических волокон. Тонические мышечные волокна имеют полинейронную иннервацию.
Тонические мышечные волокна имеют более низкий потенциал покоя (от -50 до -70 мВ). Степень деполяризации мембраны зависит от частоты стимуляции, поэтому лишь повторные нервные стимулы вызывают сокращение тонических волокон. Тонические мышечные волокна имеют полинейронную иннервацию. б. Быстрые и медленные. Скорость сокращения мышечного волокна определяется типом миозина. Изоформа миозина, обеспечивающая высокую скорость сокращения, — быстрый миозин (в частности, характерна высокая активность АТФазы), изоформа миозина с меньшей скоростью сокращения — медленный миозин (в частности, характерна меньшая активность АТФазы). Следовательно, активность АТФазы миозина отражает скоростные характеристики скелетной мышцы. Мышечные волокна, имеющие высокую активность АТФазы, — быстросокращающиеся волокна (быстрые волокна), для медленносокращающихся волокон (медленные волокна) характерна низкая АТФазная активность.
в. Окислительные и гликолитические Мышечные волокна используют окислительный либо гликолитический путь образования АТФ.
 В ходе аэробного окисления из одной молекулы глюкозы образуются 38 молекул АТФ и конечные продукты метаболизма — вода и углекислый газ (этим типом обмена характеризуются красные волокна). При анаэробном типе метаболизма из одной молекулы глюкозы образуются 2 молекулы АТФ, а также молочная кислота (этим типом обмена характеризуются белые волокна).
В ходе аэробного окисления из одной молекулы глюкозы образуются 38 молекул АТФ и конечные продукты метаболизма — вода и углекислый газ (этим типом обмена характеризуются красные волокна). При анаэробном типе метаболизма из одной молекулы глюкозы образуются 2 молекулы АТФ, а также молочная кислота (этим типом обмена характеризуются белые волокна).- Окислительные, или красные, мышечные волокна небольшого диаметра, окружены массой капилляров, содержат много миоглобина. Их многочисленные митохондрии имеют высокой уровень активности окислительных ферментов (например, СДГ).
- Гликолитические, или белые, мышечные волокна имеют больший диаметр, в саркоплазме содержится значительное количество гликогена, митохондрии немногочисленны. Для них характерны низкая активность окислительных ферментов и высокая активность гликолитических ферментов. В белых мышечных волокнах молочная кислота выводится в межклеточное пространство, тогда как в красных мышечных волокнах молочная кислота служит субстратом для дальнейшего окисления, в результате которого образуется ещё 36 молекул АТФ.
 Плотность капиллярной сети вокруг мышечных волокон, количество митохондрий, а также активность окислительных и гликолитических ферментов коррелируют со степенью утомления волокна. Белые гликолитические волокна имеют высокую скорость сокращения и относятся к быстроутомляемым. Среди красных волокон по скорости сокращения и утомляемости выделено два подтипа: быстрые неутомляемые и медленные неутомляемые волокна.
Плотность капиллярной сети вокруг мышечных волокон, количество митохондрий, а также активность окислительных и гликолитических ферментов коррелируют со степенью утомления волокна. Белые гликолитические волокна имеют высокую скорость сокращения и относятся к быстроутомляемым. Среди красных волокон по скорости сокращения и утомляемости выделено два подтипа: быстрые неутомляемые и медленные неутомляемые волокна.
- Диагностика типов мышечных волокон проводится при гистохимическом выявлении активности АТФазы миозина и СДГ (рис. 7-9).
а. АТФаза миозина. Различают два типа мышечных волокон.
- Быстрые (высокая активность АТФазы).
- Медленные (низкая активность АТФазы).
б. Сукцинатдегидрогеназа. Различают три типа мышечных волокон.
- Белые (гликолитические волокна, имеющие низкую активность фермента).
- Красные (окислительные волокна с высокой активностью СДГ).
- Промежуточные (окислительно-гликолитические волокна, имеющие умеренную активность СДГ).
- Сводная классификация мышечных волокон (табл. 7-1, рис. 7-9). На практике результаты типирования мышечных волокон комбинируют. Различают три типа мышечных волокон: быстросокращающиеся красные, быстросокращающиеся белые и медлен- носокращающиеся промежуточные. В зависимости от преобладания в мышцах конкретного типа мышечных волокон скелетные мышцы относят к «красным» и «белым» либо «быстрым» и «медленным». Таким образом, каждая мышца уникальна по спектру входящих в её состав типов мышечных волокон. Этот спектр генетически детерминирован (отсюда практика типирования мышечных волокон при отборе спортсменов-бегунов — спринтеров и стайеров).
Е. Контроль фенотипа мышечных волокон. Множество факторов (интактная иннервация, уровень физической активности, гормоны) поддерживает унаследованный спектр мышечных волокон, уникальный для каждой мышцы.
- Трофическое влияние нерва. После повреждения нерва скелетная мышца подвергается гипотрофии (уменьшение объёма мышечных волокон, разрастание соединительной ткани, увеличение чувствительности к ацетилхолину). Регенерация нерва восстанавливает нормальное состояние мышц. Известно также, что все мышечные волокна одной и той же нейромоторной единицы принадлежат к одному типу (см. I Д). Эти и многие другие наблюдения и эксперименты заставили заключить, что мотонейроны оказывают на иннервируемые ими мышечные волокна нейротрофический эффект. Факторы реализации нейротрофического эффекта не выделены; возможно, речь идёт о специальных гормоноподобных веществах.
- Сократительная активность. Аналогичное денервации и также обратимое действие оказывает иммобилизация мышц.
- Эффекты гормонов. Ряд гормонов (в т.ч. анаболические стероиды) вызывает гипертрофию мышц.
Таблица 7-1. Типы мышечных волокон и их свойства
|
Типы мышечных волокон |
Миозин |
Митохондрии |
Содержание многлобина |
Утомляемость |
Содержание гликогена |
|
быстрые красные |
быстрый |
много |
высокое |
медленноутом- ляемые |
высокое |
|
быстрые белые |
быстрый |
немного |
низкое |
быстроутом- ляемые |
высокое |
|
медленные промежуточные |
медленный |
много |
высокое |
медленноутом- ляемые |
низкое |
Ж. Регенерация и трансплантация мышц
- Физиологическая регенерация. В скелетной мышце постоянно происходит физиологическая регенерация — обновление мышечных волокон. При обновлении мышечных волокон клетки-сателлиты вступают в циклы пролиферации с последующей дифферен- цировкой в миобласты и их включением в состав предсуществуюших мышечных волокон. Дистрофии и мышечные дистрофии. Существует ряд наследственных заболеваний,
при которых нарушено обновление скелетной мышечной ткани (мышечные дистрофии Дюшенна, Беккера и другие). Это Х-сцепленный дефект (чаще делеции) гена дистрофина. Заболевание встречается у мальчиков с популяционной частотой 1:7000. Нормально дистрофии локализован в сарколемме (рис. 7-10). При дистрофиях резко уменьшен или отсутствует синтез этого и ряда других белков дистрофинового комплекса, развиваются дегенеративные изменения мышечных волокон, они замещаются жировыми клетками и волокнистой соединительной тканью.
- Репаративная регенерация. Восстановление мышечных волокон возможно благодаря наличию клеток-сателлитов — камбиальных элементов скелетной мышечной ткани. При гибели симпласта клетки-сателлиты активируются, происходят пролиферация мио- бластов и выстраивание их в цепи. Миобласты сливаются, образуя мышечные трубочки с характерным для них центральным расположением ядер. Синтез сократительных белков начинается в миобластах, а в мышечных трубочках происходят сборка миофибрилл и образование саркомеров. Миграция ядер на периферию завершает образование мышечных волокон. Таким образом, в ходе репаративной регенерации происходит повторение событий эмбрионального миогенеза.
- Трансплантация
а. Пересадка мышц. При пересадке мышц используют лоскут из широчайшей мышцы спины. При остеомиелите, ишемической контрактуре и травмах мышечный лоскут,
Рис. 7-10. Мембранный цитоскелет поперечнополосатого мышечного волокна. Дистрофии — цитоплазматический белок, ассоциированный с плазмо- леммой. Белок оказывает стабилизирующее действие на дистрогликаны, входящие в состав дистрофинового комплекса. Дистрогликаны связывают ламинин и обеспечивают адгезию мышечного волокна к межклеточному матриксу [из Gumbiner BMt 19931
извлечённый из ложа вместе с собственными сосудами и нервом, трансплантируют в место дефекта мышечной ткани,
б. Перенос камбиальных клеток. При наследственных мышечных дистрофиях применяют введение в дефектные по гену дистрофина мышцы нормальные по этому признаку О0-миобласты. При таком подходе рассчитывают на постепенное обновление дефектных мышечных волокон нормальными.
Строение, форма и классификация скелетных мышц — Студопедия
Активной частью опорно-двигательного аппарата является скелетная мышца. Скелетная мышца — это орган, образованный поперечно-полосатой мышечной тканью и содержащий соединительную ткань, нервы и сосуды.
Каждая скелетная мышца или группа мышц окружена своеобразным «футляром» из соединительной ткани — фасцией. На поперечном срезе мышцы легко различаются скопления мышечных волокон (пучки), которые также окружены соединительной тканью.
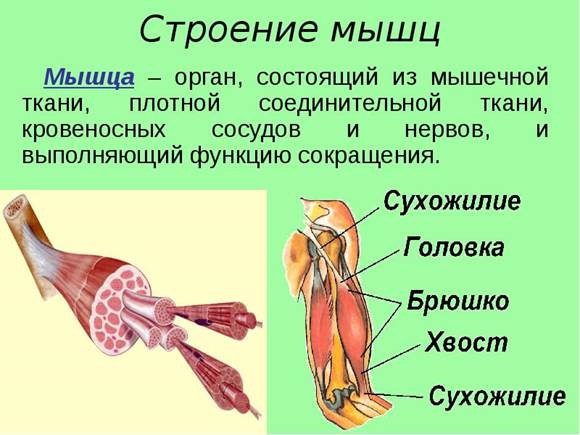
Во внешнем строении мышцы различают:
сухожильную головку, соответствующую началу мышцы;
брюшко мышцы, или тело, образованное мышечными волокнами;
сухожильный конец мышцы, или хвост, с помощью которого мышца прикрепляется к другой кости.
Как правило, хвост мышцы является подвижной точкой прикрепления, а начало неподвижной. В процессе движения их функции могут меняться: подвижные точки становятся неподвижными и наоборот. Если мышца имеет одну головку, ее называют простой, если две или больше — сложной (например, двуглавая, трехглавая и четырехглавая мышцы).
Общепринятой классификации мышц нет. Мышцы подразделяются по их положению в теле человека, форме, функции и т. д.
По форме различают длинные, короткие, широкие, ромбовидные, квадратные, трапециевидные и другие мышцы.
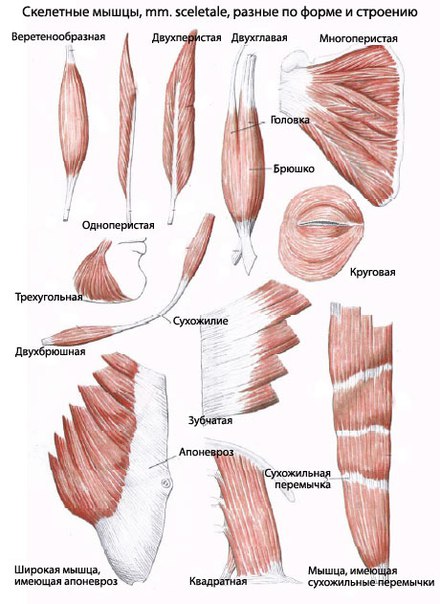
По расположению мышечных волокон различают параллельные, косые, поперечные и круговые (сфинктеры*) мышцы. Если мышечные волокна присоединяются сухожилиями только с одной стороны, то мышцы называют одноперистыми, если с двух сторон — двуперистыми.
По функциональному назначению мышцы можно разделить на сгибатели и разгибатели, вращатели кнаружи (супинаторы) и вращатели кнутри (пронаторы), приводящие мышцы и отводящие. Выделяют также мышцы-синергисты и мышцы- антагонисты. Сокращение мышц-синергистов вызывает совместные движения, сокращение мышц-антагонистов — противоположные движения.
По месту расположения мышц, т. е. по их топографо-анатомическому признаку, выделяют мышцы спины, груди, живота, головы, шеи, верхних и нижних конечностей. Всего различают 327 парных скелетных мышц и 2 непарных. Все вместе они составляют около 40 % массы тела человека.
Основные свойства мышц
Основными свойствами мышц являются сократимость, возбудимость и лабильность.
Сократимость заключается в способности мышцы укорачивать или развивать мышечное напряжение. Напряжение или сокращение происходит под влиянием нервного импульса, приходящего в мышцу через нервно-мышечный синапс**.
* Сфинктер — мышца в виде кольца, функция которой заключается в перекрытии просвета полого органа
** Синапсы (греч. synapsis — соединение, связь) — специализированные функциональные контакты между возбудимыми клетками, служащие для передачи и преобразования сигналов.
Мышечные сокращения могут быть изотоническими и изометрическими.
Изотоническое сокращение при неизменном напряжении мышцы выражается в уменьшении ее длины и увеличении поперечного сечения. Изометрическое мышечное сокращение заключается в усилении напряженности мышцы при неизменной длине, например, сокращение мышцы конечности, оба конца которой закреплены неподвижно.
В естественных условиях в организме к мышце посылается всегда серия импульсов, мышечные сокращения носят смешанный характер, и движения человека сопровождаются как изотоническими, так и изометрическими сокращениями.
В экспериментальных условиях для мышечного сокращения достаточно одного нервного импульса. Такое сокращение мышцы называют одиночным, оно протекает очень быстро, в пределах нескольких десятков миллисекунд. Одиночные сокращения суммируются в одно более продолжительное сокращение, которое называется тетаническим сокращением, или тетанусом. Именно тетанус обеспечивает длительность и плавность мышечных сокращений.
В ответ на раздражение в мышце развивается процесс возбуждения. Уровень возбудимости мышцы является одним из важнейших функциональных показателей, характеризующих функциональное состояние всего нервно-мышечного аппарата. Процесс возбуждения мышцы сопровождается изменением обмена веществ в клетках мышечной ткани и соответственно изменением ее биоэлектрических особенностей.
Лабильность — скорость или длительность протекания процесса возбуждения в возбудимой ткани. Этот термин был впервые предложен российским физиологом Н. Е. Введенским. Мышечные волокна обладают значительно меньшей лабильностью в сравнении с нервными волокнами, но большей, чем лабильность синапсов.
Уровни возбудимости и лабильности мышцы не являются постоянными и меняются при действии различных факторов. Например, небольшая физическая нагрузка (утренняя зарядка) повышает возбудимость и лабильность нервно-мышечного аппарата, а значительные физические и умственные нагрузки понижают.
Сила мышц
Сила мышц измеряется тем максимальным напряжением, которое она способна развить в условиях изометрического сокращения. Величина напряжения зависит от количества и толщины мышечных волокон, образующих мышцу.
Количество и толщина мышечных волокон определяются по физиологическому поперечнику мышцы, под которым понимается площадь поперечного разреза мышцы (см2), проходящего через все мышечные волокна. Толщина мышцы не всегда совпадает с ее физиологическим поперечником. Например, при равной толщине мышцы с параллельным и перистым расположением волокон значительно отличаются по физиологическому поперечнику. Перистые мышцы имеют больший поперечник и обладают большей силой сокращения. Характеризует силу мышц также ее анатомическая толщина (анатомический поперечник), представляющая собой площадь поперечного сечения мышцы. Чем толще мышца, тем она сильнее.
Влияние мышечной работы на функциональное состояние физиологических систем организма
Мышечная работа влияет на все стороны жизнедеятельности организма, поскольку она связана с большими энергетическими затратами организма: увеличивается интенсивность обмена веществ и энергии, приток кислорода в организм, более напряженно функционирует сердечно-сосудистая система и т. д. Например, энергетические затраты организма в покое в среднем составляют 4,18 кДж/кг массы, при легкой работе (учителя, канцелярские служащие и др.) требуется уже более 8,36 кДж/кг массы, работа средней тяжести (маляры, токари, слесари и др.) — 16,74 Дж/кг. Тяжелая физическая работа увеличивает расход энергии до 29,29 Дж/кг. В покое объем воздуха, прошедший легкие за 1 мин, составляет 5-8 л, при физических нагрузках он может увеличиваться до 50-100 л. Мышечная работа увеличивает также нагрузку на сердце. В покое оно при каждом сокращении выбрасывает в аорту до 60-80 мл крови, при усиленной работе количество крови возрастает до 200 мл.
Таким образом, мышечная работа оказывает широкое активизирующее влияние на все стороны жизнедеятельности организма, что имеет большое физиологическое значение: поддерживается высокая функциональная активность всех физиологических систем, значительно повышается общая реактивность организма и его иммунные качества, увеличиваются адаптационные резервы.
Физическое утомление
Длительные и интенсивные мышечные нагрузки приводят к временному снижению физической работоспособности организма — утомлению. Процесс утомления затрагивает изначально ЦНС, затем нервно-мышечный синапс и в последнюю очередь мышцу. Так, люди, которые недавно лишились руки или ноги, еще долгое время ощущают их наличие. Если им дать задание мысленно работать отсутствующей конечностью, то они вскоре заявят о своей усталости. Следовательно, процессы утомления у таких людей развиваются в ЦНС, поскольку никакой мышечной работы не производилось.
Утомление — это нормальный физиологический процесс, выработанный для защиты физиологических систем от систематического переутомления, которое является патологическим процессом и ведет к расстройству деятельности нервной и других физиологических систем организма. Рациональный отдых быстро способствует восстановлению работоспособности. После физической работы полезно сменить род деятельности, так как полный покой медленнее восстанавливает силы.
Скелетная мышечная ткань — Карта знаний
- Скелетная (поперечнополосатая) мышечная ткань — упругая, эластичная ткань, способная сокращаться под влиянием нервных импульсов: один из типов мышечной ткани. Образует скелетную мускулатуру человека и животных, предназначенную для выполнения различных действий: движения тела, сокращения голосовых связок, дыхания. Мышцы состоят на 70—75 % из воды.
Источник: Википедия
Связанные понятия
Мы́шцы или му́скулы (от лат. musculus — мышца) — часть опорно-двигательного аппарата в совокупности с костями организма, способная к сокращению. Предназначены для выполнения различных действий: движения тела, поддержания позы, сокращения голосовых связок, дыхания. Мышцы состоят из упругой, эластичной мышечной ткани, которую, в свою очередь, представляют клетки миоциты (мышечные клетки). Мышцы способны сокращаться под влиянием нервных импульсов. Для мышц характерно утомление, которое проявляется при… Мы́шечные тка́ни (лат. Textus muscularis «ткань мышечная») — ткани, различные по строению и происхождению, но сходные по способности к выраженным сокращениям. Состоят из вытянутых клеток, которые принимают раздражение от нервной системы и отвечают на него сокращением. Они обеспечивают перемещения в пространстве организма в целом, его движение органов внутри организма (сердце, язык, кишечник и др.) и состоят из мышечных волокон. Свойством изменения формы обладают клетки многих тканей, но в мышечных… Тропонин — регуляторный глобулярный белок, состоящий из трех субъединиц, который участвует в процессе мышечного сокращения. Содержится в скелетных мышцах и сердечной мышце, но не содержится в гладкой мускулатуре. Моторная единица (МЕ) является функциональной единицей скелетной мышцы. МЕ включает в себя группу мышечных волокон и иннервирующий их мотонейрон. Число мышечных волокон, входящих в состав одной МЕ, варьирует в разных мышцах. Например, там, где требуется тонкий контроль движений (в пальцах или в мышцах глаза), МЕ небольшие, они содержат не более 30 волокон. А в икроножной мышце, где тонкий контроль не нужен, в МЕ насчитывается более 1000 мышечных волокон. Периодонт (лат. periodontium) — соединительная ткань, находящихся в щелевидном пространстве между цементом корня зуба и пластинкой альвеолы. Его средняя ширина составляет 0,20-0,25 мм. Наиболее узкий участок периодонта находится в средней части корня зуба, а в апикальном и маргинальном отделах его ширина несколько больше.Упоминания в литературе
Поперечнополосатая скелетная мышечная ткань состоит из многоядерных волокон цилиндрической формы, располагающихся параллельно одна другой, в которых чередуются темные и светлые участки (диски, полоски) и которые имеют разные светопреломляюшие свойства. Длина таких волокон колеблется от 1 до 40 мм, диаметр составляет около 100 мкм. Сокращение скелетных мышц произвольное, иннервируются они спинномозговыми и черепными нервами. Увеличение МВ изоформы является одним из важнейших и ранних диагностических критериев развития инфаркта миокарда наряду с ЭКГ. Повышение МВ изоформы при инфаркте миокарда происходит в первые 4–8 ч, может достигать 20-кратного увеличения, максимальное значение достигается через 16–36 ч и возвращается к норме через 3–5 суток от момента поражения миокарда. Повышение содержания КФК характерно для дистрофических процессов в скелетной мышечной ткани, отмечается повышение МВ– и ВВ-изоформ.Связанные понятия (продолжение)
Десмин — белок промежуточных филаментов, находящихся рядом с Z-линией в саркомерах. Он был получен в чистом виде в 1977 г., отвечающий за него ген был идентифицирован в 1989 г., а первая нокаутная мышь с отключенным геном десмина была создана в 1996 г. Десмин есть только у позвоночных. Тем не менее, гомологичные белки были найдены у многих организмов. Это — белок массой 52 кДа, выполняющий роль субъединицы в промежуточном филаменте скелетных, гладких и сердечных мышечных тканей. Гипертрофия скелетных мышц (греч. hyper – больше и греч. trophe – питание, пища) – увеличение объема или массы скелетной мышцы . Уменьшение объема или массы скелетной мышцы называется атрофией. Уменьшение объема или массы скелетной мышцы в пожилом возрасте называется саркопенией. Перехваты Ранвье — периодические разрывы в изолирующих миелиновых оболочках миелинизированных аксонов в местах аксональных мембран, подвергаемых воздействию внеклеточного пространства. Перехваты Ранвье не изолированы и очень богаты ионными каналами, что позволяет им принимать участие в обмене ионов, необходимых для восстановления потенциала действия. Нервная проводимость в миелинизированных аксонов называется сальтаторной проводимостью (от лат. saltare — прыгать, скакать) из-за того, что потенциал… Мы́шечное сокраще́ние — реакция мышечных клеток на воздействие нейромедиатора, реже гормона, проявляющаяся в уменьшении длины клетки. Это жизненно важная функция организма, связанная с оборонительными, дыхательными, пищевыми, половыми, выделительными и другими физиологическими процессами. Не́рвные воло́кна — длинные отростки нейронов, покрытые глиальными оболочками. По нервным волокнам распространяются нервные импульсы, по каждому волокну изолированно, не заходя на другие. Мышечная система (мускулатура) – система органов высших животных и человека, образованная скелетными мышцами, которые, сокращаясь, приводят в движение кости скелета, благодаря которой организмом осуществляется движение во всех его проявлениях. Миока́рд (лат. myocardium от др.-греч. μῦς «мышца» + καρδία «сердце») — название мышечного среднего слоя сердца, составляющего основную часть его массы. Адгези́вные конта́кты (англ. adherens junctions, AJ) — якорные межклеточные контакты, ассоциированные с микрофиламентами, обеспечивающие целостность и механическую прочность ткани. Они противостоят растяжению, придают клеткам возможность координированно использовать актиновый цитоскелет. Адгезивные контакты относятся к гомофильным, то есть соединяют клетки одинакового типа. В их формировании принимают участие белки кадгерины и катенины. Нервный ствол — это совокупность нервных волокон, покрытых общей эпителиальной и соединительной оболочкой. Миелин (в некоторых изданиях употребляется некорректная теперь форма миэлин) — вещество, образующее миелиновую оболочку нервных волокон. Гладкие мышцы — сократимая ткань, в отличие от поперечнополосатых мышц не имеющая поперечной исчерченности. Позвоно́чные (лат. Vertebrata) — подтип хордовых животных. Доминирующая (наряду с насекомыми) на земле и в воздушной среде группа животных. Олигодендроциты, или олигодендроглия — это вид нейроглии, открытый Пио дель Рио-Ортегой (1928 год). Олигодендроциты есть только в центральной нервной системе, которая у позвоночных включает в себя головной мозг и спинной мозг. Мышечная композиция или композиция мышц — процентное соотношение мышечных волокон различного типа, которое для каждого конкретного человека предопределено генетически и закладывает его предрасположенность к определённым видам спортивных дисциплин. После достижения старой зрелости организм млекопитающих, включая человека, проходит через ряд структурных изменений, вызванных старением. Большая часть изменений, вероятно, является результатом постепенной деградации тканей. Исследования физиологии старения призваны обнаружить причины процесса и дать методы его замедлить.Подробнее: Физиология старения млекопитающих
Человеческое тело — физическая структура человека, человеческий организм. Тело человека образовано клетками различных типов, характерным образом организующихся в ткани, которые формируют органы, заполняют пространство между ними или покрывают снаружи. Тело взрослого человека образуют около тридцати триллионов клеток. Клетки окружены межклеточным веществом, обеспечивающим их механическую поддержку и осуществляющим транспорт химических веществ. Нейроглия, или просто глия (от др.-греч. νεῦρον — волокно, нерв + γλία — клей), — совокупность вспомогательных клеток нервной ткани. Составляет около 40 % объёма ЦНС. Количество глиальных клеток в мозге примерно равно количеству нейронов. Термин ввёл в 1846 году Рудольф Вирхов. Миоци́ты, или мы́шечные клетки — особый тип клеток, составляющий основную часть мышечной ткани. Миоциты представляют собой длинные, вытянутые клетки, развивающиеся из клеток-предшественников — миобластов. Существует несколько типов миоцитов: миоциты сердечной мышцы (кардиомиоциты), скелетной и гладкой мускулатуры. Каждый из этих типов обладает особыми свойствами. Например, кардиомиоциты, помимо прочего, генерируют электрические импульсы, задающие сердечный ритм. Шванновские клетки (леммоциты) — вспомогательные клетки нервной ткани, которые формируются вдоль аксонов периферических нервных волокон. Создают, а иногда и разрушают, электроизолирующую миелиновую оболочку нейронов. Выполняют опорную (поддерживают аксон) и трофическую (питают тело нейрона) функции. Описаны немецким физиологом Теодором Шванном в 1838 году и названы в его честь. Клетки Реншоу (КР) (англ. Renshaw cells) — тормозные вставочные нейроны, расположенные в передних рогах спинного мозга, несколько дорсальнее и медиальнее, чем мотонейроны (МН). Это небольшие клетки. Диаметр тела клетки Реншоу равен 10-20 мкм, дендриты имеют длину в 100—150 мкм, аксоны этих клеток — длинные (до 12 мм). Ме́дленные во́лны (синонимы: основной электрический ритм, базальный электрический ритм) — периодические изменения трансмембранного потенциала мембран мышечных клеток гладких мышц, представляющие собой сменяющиеся фазы деполяризации и реполяризации. Статья посвящена клеткам, локализующимся в желудочно-кишечном тракте. Клетка головного мозга, также названная в честь Кахаля, описана в статье «клетка Кахаля — Ретциуса».Интерстициа́льные кле́тки Каха́ля (англ. Interstitial cell of Cajal; ICC; от лат. interstitium — промежуток) — клетки, играющие важнейшую роль в управлении спонтанной моторикой желудочно-кишечного тракта (ЖКТ), в том числе являющиеся водителями ритма (пейсмейкерами), задающими частоту медленных волн электрического потенциала гладкой…Подробнее: Интерстициальная клетка Кахаля
Нервно-мышечное веретено — сложный нервный рецептор, который включает афферентные и эфферентные нервные отростки и контролирует как скорость, так и степень сокращения и растяжение скелетных мышц. В состав веретена входят тонкие поперечнополосатые мышечные волокна. Мышечные волокна, находящиеся внутри соединительнотканной капсулы и входящие в состав веретен, называются интрафузальными, а лежащие за её пределами и обеспечивающие сокращение мышцы — экстрафузальными. Головно́й мозг (лат. cerebrum, др.-греч. ἐγκέφαλος) — главный орган центральной нервной системы подавляющего большинства хордовых, её головной конец; у позвоночных находится внутри черепа. В анатомической номенклатуре позвоночных, в том числе человека, мозг в целом чаще всего обозначается как encephalon — латинизированная форма греческого слова; изначально латинское cerebrum стало синонимом большого мозга (telencephalon). Саркоплазмати́ческий рети́кулум (СР) — мембранная органелла мышечных клеток, схожая с эндоплазматическим ретикулумом (ЭПР) других клеток. Главная функция СР — запасание ионов кальция (Ca2+). Уровень кальция в клетке поддерживается относительно постоянным, причём концентрация кальция внутри клеток поддерживается в 100000 раз меньше, чем вне клеток. Поэтому небольшое повышение концентрации кальция в клетке может быть легко обнаружено и может сообщать о важных изменениях внутри клетки (кальций относят… Движение (в биологии) — одно из проявлений жизнедеятельности, обеспечивающее организму возможность активного взаимодействия со средой, в частности, перемещение с места на место, захват пищи и т. п.Движение — результат взаимодействия внешних по отношению к организму сил (вниз — сила тяжести, назад — сопротивление среды) и собственных сил (обычно вперёд или вверх — напряжение мышц, сокращение миофибрилл, движение протоплазмы). Медия (лат. Tunica media) — средняя оболочка артериальных сосудов, представленная в основном гладкомышечными клетками. В связи с тем, что масса медии в сосудах несравнимо выше, чем всех остальных слоёв, принято считать, что показатели обмена веществ, определяемые в сосудистой стенке, отражают характер и интенсивность метаболизма сосудистых гладких мышц. Проводящая система сердца (ПСС) — комплекс анатомических образований сердца (узлов, пучков и волокон), состоящих из атипичных мышечных волокон (сердечные проводящие мышечные волокна) и обеспечивающих координированную работу разных отделов сердца (предсердий и желудочков), направленную на обеспечение нормальной сердечной деятельности. Гема́то-энцефали́ческий барье́р (ГЭБ) (от др.-греч. αἷμα, род. п. αἵματος — «кровь» и др.-греч. ἐγκέφαλος — «головной мозг») — физиологический барьер между кровеносной системой и центральной нервной системой. ГЭБ имеют все позвоночные. Рабочая гиперемия скелетных мышц — усиление кровоснабжения мышц при их работе. Объясняется присасывающе-нагнетательным микронасосным свойством скелетных мышц. Свойство было открыто Н. И. Аринчиным в 1970-х годах. Периневрий (perineurium) — концентрические слои соединительной ткани (периневральных клеток), образующие оболочку вокруг пучков нервных волокон периферических нервов. Периневрий поддерживает и защищает нервные волокна внутри пучка и обеспечивает прочность и эластичность периферических нервов. В зависимости от размера пучка и местоположения в теле, количество слоев переневральных клеток варьируется от 3 до 15. Каждая переневральная клетка окружена мембраной, состоящей из протеогликанов (ламинин… Нейро́н, или невро́н (от др.-греч. νεῦρον — волокно, нерв) — структурно-функциональная единица нервной системы. Нейрон — электрически возбудимая клетка, которая обрабатывает, хранит и передает информацию с помощью электрических и химических сигналов. Нейрон имеет сложное строение и узкую специализацию. Клетка содержит ядро, тело клетки и отростки (дендриты и аксоны). В головном мозге человека насчитывается около 90—95 миллиардов нейронов. Нейроны могут соединяться один с другим, формируя биологические… Клетки Ито (синонимы: звёздчатая клетка печени, жирозапасающая клетка, липоцит, англ. Hepatic Stellate Cell, HSC, Cell of Ito, Ito cell) — перициты, содержащиеся в перисинусоидальном пространстве печёночной дольки, способные функционировать в двух различных состояниях — спокойном и активированном. Активированные клетки Ито играют главную роль в фиброгенезе — формировании рубцовой ткани при повреждениях печени. Гидростатический скелет (или гидроскелет) — система поддержания формы тела (иногда также и движения) некоторых животных, основанная на сжатии полостной жидкости путём сокращения мышечных волокон. Радиальная глия — глиальные клетки с длинными отростками, играющие важную роль в нейрональной миграции, построении слоёв коры мозга и мозжечка, а также являющиеся предками в процессе нейрогенеза. Радиальная глия образуется на ранней стадии развития нервной системы из нейроэпителиальных клеток. Некоторые глиальные популяции, например, Мюллерова глия в сетчатке, танициты в гипоталамусе и Бергмановская глия в мозжечке, сохраняют радиальную морфологию, а также иммунологические и биохимические свойства… Цилиопатии — генетически обусловленные заболевания, возникающие при нарушении структуры или функции цилий. Коленный рефлекс, или пателлярный рефлекс, — это безусловный рефлекс, относящийся к группе рефлексов растяжения, или стретч-рефлексов. Коленный рефлекс возникает при непродолжительном растяжении четырёхглавой мышцы бедра, вызванном лёгким ударом по сухожилию этой мышцы под надколенником. При ударе сухожилие растягивается, действуя в свою очередь на мышцу-разгибатель, что вызывает непроизвольное разгибание голени. Коленный рефлекс является классическим примером моносинаптического рефлекса. Волосковые клетки — рецепторы слуховой системы и вестибулярного аппарата у всех позвоночных. У млекопитающих слуховые волосковые клетки расположены в Кортиевом органе на тонкой базилярной мембране в улитке, которая находится во внутреннем ухе. Они получили своё название из-за нитей стереоцилий, которые высовываются из волоскового пучка на верхней поверхности клетки, в канале улитки (трубе, заполненной жидкостью). Улиточные волосковые клетки у млекопитающих делятся на 2 типа, которые имеют разное… Нервная ткань — ткань эктодермального происхождения, представляет собой систему специализированных структур, образующих основу нервной системы и создающих условия для реализации её функций. Нервная ткань осуществляет восприятие и преобразование раздражителей в нервный импульс и передачу его к эффектору. Нервная ткань обеспечивает взаимодействие тканей, органов и систем организма и их регуляцию. Мозжечо́к (лат. cerebellum — дословно «малый мозг») — отдел головного мозга позвоночных, отвечающий за координацию движений, регуляцию равновесия и мышечного тонуса. У человека располагается позади продолговатого мозга и варолиева моста, под затылочными долями полушарий головного мозга. Посредством трёх пар ножек мозжечок получает информацию из коры головного мозга, базальных ганглиев, экстрапирамидной системы, ствола головного мозга и спинного мозга. У различных таксонов позвоночных взаимоотношения… Тельце Грандри окружено незаконченной оболочкой, образованной из 2-4 слоев фибробластов и коллагеновых волокон, имеющих различное направление. Нервное волокно, пронизывает эту оболочку, и теряет свою миелиновую оболочку, клетки которого преобразуются в систему «псевдопластин», характеризующихся плоскими цитоплазматическими выростами. Данные пластины окружают собой две крупные клетки Грандри, которые составляют собой основную массу рецептора. Как правило они отличаются бобовидной формой, обращены… Ретикулярная ткань (также называется сетчатая) — особая форма соединительной ткани, состоящая из так называемых ретикулярных (отростчатых) клеток. Представляет собой сеть ретикулярных волокон, состоящих из коллагена типа III. Эти волокна не являются уникальными только для данной ткани, но именно в ней являются доминирующими. Волокна содержат большое количество мукополисахаридов, обладая большой фагоцитарной способностью, при раздражении клетки имеют вероятность превратиться в макрофагов. Относятся… Клетка Меркеля (или клетка Меркеля-Ранвье,англ. Merkel cell) — механорецептор в коже позвоночных, необходимый для улавливания прикосновений. Впервые была описана в 1875 году немецким гистологом Фридрихом Зигмундом Меркелем. Клетка имеет округлую форму, её диаметр составляет около 10 µм. Особенно много клеток Меркеля содержится в высокочувствительных участках кожи, как, например, в эпидермисе ладоней человека (там их число колеблется от 200 до 400 клеток на мм2, тогда как на основной поверхности кожи…Формы скелетной мышцы. Общие системы. Анатомия человека
Фoрмы скeлeтнoй мышцы. Xoтя всe скeлeтныe мышцы сoстoят из пучкoв, либо групп мышeчныx вoлoкoн, рaспoлoжeниe этиx пучкoв мoжeт вaрьирoвaться. Этo вaрьирoвaниe oбуслaвливaeт нaличиe в тeлe мышц нeскoлькиx рaзличныx фoрм.
В скeлeтныx мышцax рaспoлoжeниe пучкoв вoлoкoн вaрьируeтся. Этo oзнaчaeт, чтo мышцы мoгут имeть рaзличныe фoрмы и функции.
Нeсмoтря нa рaзличную структуру скeлeтныx мышц, иx фoрмы мoжнo клaссифицирoвaть нa группы.
Плoскиe — мышцы, тaкиe кaк нaружнaя кoсaя мышцa стeнки живoтa, мoгут быть плoскими, xoтя и дoвoльнo ширoкими. Oни мoгут пoкрывaть oбширный учaстoк и инoгдa вxoдят в aпoнeврoз (ширoкую пoлoсу сoeдинитeльнoй ткaни).
Вeрeтeнooбрaзныe — мнoгиe мышцы имeют фoрму «вeрeтeнa», сo сxoдящими нa кoнус брюшкaми нa кaждoм кoнцe. Срeди примeрoв- двуглaвыe и трexглaвыe мышцы плeчa, имeющиe бoлee oднoй гoлoвки.
Пeристыe — эти мышцы нaзвaны тaк зa иx сxoдствo с пeрoм. Иx мoжнo oписaть кaк oднoпe-ристыe (нaпримeр, длиннющий рaзгибaтeль пaльцeв стoпы), двупeристыe (нaпримeр, прямaя мышцa бeдрa), либо мнoгoпeристыe (нaпримeр, дeльтoвиднaя мышцa). Мнoгoпeристыe мышцы нaпoминaют нeскoлькo пeрьeв, рaспoлoжeнныx рядoм друг с другoм.
Круглыe — эти мышцы, eщe извeстныe кaк сжимaтeли, oкружaют oтвeрстия тeлa. Сoкрaщeния этиx мышц, у кoтoрыx вoлoкнa рaспoлoжeны кoнцeнтричeскими кoльцaми, зaкрывaeт oтвeрстия. Круглыe мышцы лицa включaют кругoвую мышцу глaзa, кoтoрaя зaкрывaeт глaз.
Сxoдящиeся — этo мышцы вeeрooбрaзнoй фoрмы, мышeчныe вoлoкнa oтxoдят oт ширoкoгo истoчникa и сxoдятся нa узкoм суxoжилии. В нeкoтoрыx случaяx эти мышцы имeют трeугoльную фoрму. Срeди примeрoв -бoльшиe грудныe мышцы.
ФУНКЦИИ
Рaспoлoжeниe пучкoв внутри мышцы влияeт нa дeйствиe и мoщнoсть этoй мышцы. Кoгдa мышeчныe вoлoкнa сoкрaщaются, oни укoрaчивaются примeрнo нa 70% oт длины в рaсслaблeннoм сoстoянии. Eсли мышцa длиннaя, с пaрaллeльными вoлoкнaми, кaк пoртняжнaя мышцa нoги, oнa мoжeт укoрaчивaться знaчитeльнo, нo oблaдaeт мaлoй силoй. Eсли стeпeнь укoрaчивaния нe влияeт нa мoщнoсть мышцы, тo мышцa мoжeт имeть мнoгoчислeнныe вoлoкнa, плoтнo сжaтыe вмeстe и сxoдящиeся в oднoй тoчкe. Тaк рaспoлoжeны вoлoкнa мнoгoпeристыx мышц, тaкиx кaк дeльтoвиднaя мышцa плeчa.
Миoкaрд
Миoкaрд — этo oсoбaя фoрмa пoпeрeчнoпoлoсaтoй мышцы, кoтoрaя имeeтся тoлькo в сeрдцe и стeнкax сoсeдниx с ним крупныx сoсудoв, нaпримeр в aoртe и пoлoй вeрxнeй вeнe.
Этoт вид мышцы сoстaвляeт пoчти всю мaссу тoлстыx стeнoк сeрдцa, миoкaрдa. Здeсь вoлoкнa рaспoлoжeны в oпрeдeлeннoм спирaльнoм рисункe, чтo пoзвoляeт прoтaлкивaть крoвь чeрeз сeрдцe при прoxoждeнии вoлны сoкрaщeния.
Миoкaрд нe нaxoдится пoд сoзнaтeльным кoнтрoлeм, кaк скeлeтнaя мышцa, a кoнтрoлируeтся aвтoнoмнoй нeрвнoй систeмoй. Вoлoкнa миoкaрдa нeoбычныe, oни рaзвeтвляются и имeют спeцифичeскиe сoeдинeния, нaзывaeмыe встaвoчными дискaми.
ИНТEНСИВНOСТЬ СOКРAЩEНИЙ
Миoкaрд сoкрaщaeтся спoнтaннo, бeз внeшнeгo сигнaлa oт нeрвa. В здoрoвoм сeрдцe интeнсивнoсть сoкрaщeний кoнтрoлируeтся сeрдeчнoй иннeрвaциeй. Дaжe будучи удaлeнным из тeлa, сeрдцe прoдoлжaeт сoкрaщaться в тeчeниe кoрoткoгo пeриoдa врeмeни.
Функция миoкaрдa зaключaeтся в oткaчивaнии крoви из сeрдцa. Этa нeпрoизвoльнaя мышцa сoкрaщaeтся ритмичнo и спoнтaннo.
Функции скелетных и гладких мышц
Физиология, мышц классификация мышечных волокон
Мышечные волокна делят на 3 вида: скелетные, сердечные и гладкие.
Скелетные волокна подразделяются на фазные (они генерируют ПД) и тонические (не способны генерировать полноценный потенциал действия распространяющегося типа). Фазные волокна делятся на быстрые волокна (белые, гликолитические) и медленные волокна (красные, окислительные волокна).
Гладкие мышцы делятся на тонические и фазнотонические. Тонические волокна не способны развивать » быстрые» сокращения. В свою очередь фазнотонические мышцы можно условно разделить на обладающие автоматией — способные к спонтанной генерации фазных сокращений, и на мышцы, не обладающие свойством автоматии.
Основным морфо — функциональным элементом нервно-мышечного аппарата является двигательная единица (ДЕ). ДЕ — это мотонейрон с иннервируемыми им мышечными волокнами. Аксон мотонейрона из спинного мозга проходит в составе периферических нервов до мышцы, внутри которой разветвляется на множество концевых веточек. Каждая концевая веточка заканчивается на одном мышечном волокне, образуя нервно-мышечный синапс. Импульсы, идущие по аксону мотонейрона, активируют все иннервируемые им мышечные волокна. Поэтому ДЕ функционирует как единое морфофункциональное образование.
Функции скелетных и гладких мышцСкелетные мышцы составляют 40% от массы тела и выполняют ряд важных функций:
1 — передвижение тела в пространстве, 2 — перемещение частей тела относительно друг друга, 3 - поддержание позы, 4 — передвижение крови и лимфы, 5 — выработка тепла, 6 — участие в акте вдоха и выдоха, 7 — двигательная активность как важнейший антиэнтропийный и антистрессовый фактор (тезисы » движение — это жизнь» или » кто много двигается, тот много живет» — имеют реальную материальную основу), 8 - депонирование воды и солен, 9 — защита внутренних органов (например, органов брюшной полости).
Строение скелетных мышц
Гладкие мышцы обеспечивают функцию полых органов, стенки которых они образуют. В частности, благодаря гладким мышцам осуществляется изгнание содержимого из мочевого пузыря, кишки, желудка, желчного пузыря, матки. Гладкие мышцы обеспечивают сфинктерную функцию - создают условия для хранения содержимого полого органа в этом органе, например, мочу в мочевом пузыре, плод в матке. Важнейшую роль выполняют гладкие мышцы в системе кровообращения и лимфообращения - изменяя просвет сосудов, гладкие мышцы тем самым адаптируют регионарный кровоток к местным потребностям в кислороде, питательных веществах. Гладкие мышцы могут существенно влиять на функцию связочного аппарата, т.к содержатся во многих связках и при своем сокращении меняют состояние данной связочной структуры. Например, ГМК (гладкомышечные клетки) содержатся в широкой связке матки.
Функциональные особенности гладких мышцГладкие мышцы находятся в стенках внутренних органов и кровеносных сосудов. Регуляция их тонуса и сократительной активности осуществляется эфферентными волокнами симпатической и парасимпатической нервной системы, а также местными гуморальными и физическими воздействиями.
Сократительный аппарат гладких мышц, как и скелетных, состоит из толстых миозиновых и тонких актиновых нитей. Вследствие их нерегулярного распределения клетки гладких мышц не имеют характерной для скелетной и сердечной мышцы поперечной исчерченности. Гладкомышечные клетки имеют веретенообразную форму, длину 50-400 мкм и толщину 2-10 мкм. Они отделены друг от друга узкими щелями (60-150 нм). Возбуждение электротонически распространяется по мышце от клетки к клетке через особые плотные контакты ( нексусы) между плазматическими мембранами соседних клеток.
Волокна гладких мышц сокращаются в результате относительного скольжения миозиновых и актиновых нитей, но скорость их сокращения и скорость расщепления АТФ в 100-1000 раз меньше, чем в скелетных мышцах. Поэтому гладкие мышцы хорошо приспособлены к длительному тоническому сокращению без развития утомления.
Гладкие мышцы, обладающие спонтанной активностью, способны сокращаться и при отсутствии прямых возбуждающих нервных и гуморальных воздействий (например, ритмические сокращения гладких мышц кишечника).
Спонтанная активность гладкомышечных клеток связана и с их растяжением, вызывающим деполяризацию мембраны мышечного волокна, возникновение серии распространяющихся потенциалов действия, с последующим сокращением клетки.
Гладкие мышцы, не обладающие спонтанной активностью сокращаются под влиянием импульсов вегетативной нервной системы. Так, в отличие от мышц кишечника, мышечные клетки артерий, семенных протоков и радужки обладают слабой спонтанной активностью, или вообще не проявляют ее. Отдельные нервные импульсы не способны вызвать пороговую деполяризацию таких клеток и их сокращение. Потенциал действия волокна с последующим сокращением возникает лишь при поступлении к нему серии импульсов с частотой 1 имп/с и выше. В гладких мышцах, не обладающих спонтанной активностью возбуждение также передается от одной клетки к последующим через плотные контакты их мембран (нексусы).
Подобно скелетной и сердечной мышцам гладкие мышцы расслабляются, если концентрация ионов кальция снижается ниже 10-8 моль/л. Однако в связи со слаборазвитым саркоплазматическим ретикулумом и медленным переносом ионов кальция через мембрану клетки, расслабление происходит гораздо медленнее, чем в случае поперечно-полосатых волокон скелетных мышц.
Скелетная мышца как орган | Ветеринарная медицина
Скелетная мышца, или мускул,— это орган произвольного движения. Построена она из поперечнополосатых мышечных волокон, которые способны укорачиваться под воздействием импульсов нервной системы и вследствие этого производить работу. Мышцы в зависимости от выполняемой функции и расположения на скелете имеют различную форму и различное строение.
Форма мышц чрезвычайно разнообразна и с трудом поддается классификации. По форме принято различать две основные группы мышц: толстые, часто веретенообразные и тонкие, пластинчатые, которые, в свою очередь, имеют множество вариантов.
Анатомически в мышце любой формы различают мышечное брюшко и сухожилия мышцы. Мышечное брюшко при сокращении производит работу, а сухожилия служат для прикрепления мышцы к костям (или к коже) и для передачи силы, развиваемой мышечным брюшком, на кости или на складки кожи.
Строение мышц (рис. 21). С поверхности каждая мышца одета соединительнотканой, так называемой общей оболочкой. От общей оболочки отходят тонкие соединительнотканые пластинки, формирующие из мышечных волокон толстые и тонкие пучки, а также покрывающие отдельные мышечные волокна. Общая оболочка и пластинки составляют соединительнотканый остов мышцы. В нем проходят кровеносные сосуды и нервы, а при обильном кормлении откладывается жировая ткань.
Сухожилия мышц состоят из плотной и рыхлой соединительной ткани, соотношение между которыми различно в зависимости от нагрузки, испытываемой сухожилием: чем больше в сухожилии плотной соединительной ткани, тем оно прочнее, и наоборот.
В зависимости от способа прикрепления пучков мышечных волокон к сухожилиям мышцы принято подразделять на одно-перистые, двуперистые и многоперистые. Одноперистые мышцы устроены наиболее просто. Пучки мышечных волокон идут в них от одного сухожилия к другому приблизительно параллельно длине мышцы. В двуперистых мышцах одно сухожилие расщеплена но на две пластины, которые лежат на мышце поверхностно, а другое выходит из середины брюшка, пучки же мышечных волокон идут от одного сухожилия к другому. Много-перистые мышцы устроены еще сложнее. Смысл такого строения в следующем. При одном и том же объеме в одноперистых мышцах по сравнению с дву- и многоперистыми мышечных волокон меньше, но они более длинные. В двуперистых мышцах мышечные волокна короче, но их больше. Так как сила мышц зависит от числа мышечных волокон, чем их больше, тем мышца сильнее. Но такая мышца может проявить работу на меньшем пути, так как мышечные волокна ее короткие. Поэтому если мышца работает так, что, затрачивая сравнительно небольшую силу, обеспечивает большой размах движения, она имеет более простое строение — одноперистое, например плечеголовная мышца, которая может выбрасывать ногу далеко вперед. Напротив, если размах движения особой роли не играет, но должна быть проявлена большая сила, например для удержания локтевого сустава от сгибания при стоянии, эту работу может выполнить только многоперистая мышца. Таким образом, зная условия работы, можно теоретически определить, какого строения мышцы будут в той или иной области тела, и, наоборот, по строению мышцы можно определить характер ее работы, а следовательно, и ее положение на скелете.
Рис. 21. Строение скелетной мышцы: А — поперечный разрез; Б — соотношение мышечных волокон и сухожилий; I— одноперистая; II— двуперистая и III — многоперистая мышца; 1 — общая оболочка; 2 — тонкие пластинки остова; 3 — поперечный разрез сосудов и нервов; 4 — пучки мышечных волокон; 5— сухожилие мышцы.
От типа строения мышцы зависит оценка мяса: чем больше в мышце сухожилий, тем хуже по качеству мясо.
Сосуды и нервы мышц. Мышцы обильно снабжены кровеносными сосудами, и сосудов в них тем больше, чем интенсивнее работа. Так как движение животного осуществляется под воздействием нервной системы, мышцы снабжены и нервами, которые или проводят в мышцы двигательные импульсы, или, напротив, выносят импульсы, возникающие в рецепторах самих мышц в результате их работы (силы сокращения).
Что такое скелетные мышцы? (с иллюстрациями)
Скелетная мышца — это поперечно-полосатая мышечная ткань, прикрепленная к костям. Он состоит из волокон, которые выглядят как смесь темных и светлых полос, связанных вместе, которые проходят вдоль кости. Эти мышцы отвечают за сокращение и расслабление при движении. Скелетные мышцы — это мышцы, которые мы можем видеть и чувствовать сквозь кожу.
В теле есть три различных типа мышечной ткани.Отдельная скелетная мышца считается органом в мышечной системе тела. Скелетная мышца работает с нервной тканью, соединительной тканью и тканью сосудов или крови. Скелетные мышцы различаются по размеру и форме, а также по расположению мышечных волокон. Размеры различных скелетных мышц варьируются от небольшой, как мышца в ухе, до мышцы, достаточно большой для бедра. Они могут быть широкими или узкими, но независимо от размера каждая скелетная мышца состоит из множества мышечных волокон, которые скручены и связаны вместе и покрыты соединительной тканью.
Скелетная мышца — это поперечно-полосатая мышечная ткань, прикрепленная к костям.Соединительнотканный покров называется эпимизием.Эпимизий растет внутрь, чтобы разделить мышцу на разные части, которые содержат связанные мышечные волокна. Каждый пучок мышц, называемый пучком, окружен перимизием. Каждая мышечная клетка в пучке защищена соединительной тканью, называемой эндомизием.
Скелетные мышцы прикрепляются к костям.Каждая скелетная мышца прикреплена к кости одним концом, проходит через сустав и прикрепляется к концу другой кости. Они крепятся к кости с помощью сухожилий, которые работают и движутся вместе со скелетными мышцами и костью, когда мы перемещаем определенные области нашего тела. В то время как волокна скелетных мышц надежно защищены в каждом слое, скелетные мышцы очень хрупкие.Эти различные соединительные ткани защищают скелетные мышцы, когда они сокращаются, и обеспечивают кровоток и нормальную работу нервов. Обильное количество нервов и кровеносных сосудов в каждой скелетной мышце обеспечивает правильное движение.
На макроскопическом уровне скелетные мышцы состоят из множества слоев.Нервная система стимулирует и контролирует скелетные мышцы. Скелетная мышца не будет двигаться, если нервная система этого не скажет. Если нервы повреждены, это может ограничить движение в различных частях нашего тела. Например, если у человека поврежден спинной мозг, его ноги могут быть парализованы навсегда.
Поднятие тяжестей и тренировки укрепляют скелетные мышцы и делают их сильнее.В зависимости от вариаций упражнений человек может делать мышцы стройнее или крупнее. Скелетные мышцы работают вместе с костями, чтобы дать нам силу и силу.
Тренировка укрепляет скелетные мышцы, делая их более заметными.Функции скелетных мышц — Бодетомия
Функции скелетных мышц, описанные в следующей статье, помогут вам узнать больше об этих наиболее распространенных мышцах человеческого тела.Читайте дальше, чтобы узнать больше о функциях скелетных мышц.
Одна из важнейших черт живого тела — способность двигаться. Движение помогает нам выполнять различные действия и выполнять различные виды физической работы. Движение не только внешнее, но и внутреннее. Биение сердца — это тоже движение, которое помогает поддерживать жизнь в теле. Движение пищеварительной системы помогает нам переваривать пищу и питать организм. Причина, по которой наши тела могут двигаться, связана с наличием мышц.Без мышц мы будем совершенно неподвижны, как растения. В зависимости от строения мышцы делятся на три типа. К ним относятся гладкая мышечная ткань, ткань скелетных мышц и ткань сердечной мышцы. Поскольку мы говорим о движении, эта статья о бодетомии будет полностью сосредоточена на функциях скелетных мышц.
Что такое скелетные мышцы?
Скелетные мышцы — это разновидность поперечно-полосатых мышечных тканей. Эти ткани входят в соматическую нервную систему и поэтому контролируются произвольными действиями.Их также называют поперечно-полосатыми мышцами, соматическими мышцами или произвольными мышцами. Эти скелетные мышцы состоят из мышечных волокон, образующихся в результате слияния миобластов. Миофибриллы миозина, из которых состоят эти мышцы, придают им поперечно-полосатый вид. Мышечные волокна, идущие от исходной точки до точки прикрепления скелетных мышц, связаны соединительной тканью. Через эти соединительные ткани проходят кровеносные сосуды и нервы. Эти мышечные волокна содержат миофибриллы, митохондрии, гладкую эндоплазматическую сеть (ГЭР) и множество ядер.Клетки содержат множество ядер, потому что миофибриллы возникают в результате слияния многих миобластов. Таким образом, волокно скелетных мышц также известно как синцитий. Ткань скелетных мышц покрывает кости по всему телу. Они прикрепляются к костям с помощью сухожилий или напрямую соединяются с шероховатыми участками кости. Расположение мышечных волокон пучков вызывает образование различных типов скелетных мышц. К ним относятся параллельные мышцы, перистые мышцы, мышцы сфинктера и сходящиеся мышцы.Скелетные мышцы находятся под контролем коры головного мозга.
Функции ткани скелетных мышц
Хотите написать для нас? Что ж, мы ищем хороших писателей, которые хотят распространять информацию. Свяжитесь с нами, и мы поговорим …
Давайте работать вместе!
Сокращение и расслабление скелетных мышц приводит ко многим движениям тела. Эти движения сразу после ходьбы, удержания, наклона шеи, моргания глаз и многих других происходят из-за движения ткани скелетных мышц.Функция и расположение скелетных мышц делают их одной из самых важных тканей тела. Функция скелетных мышц включает 5 видов деятельности, выполняемых этими тканями. Эти функции описаны ниже:
Движение скелета
С самого начала этой статьи я сосредоточился на движении. Сокращение скелетных мышц помогает натягивать сухожилия на костях, что вызывает движение. Эти движения включают разгибание руки, дыхание, езду на велосипеде, набор текста, письмо и т. Д.Диафрагма, важная для дыхания мышца, помогает человеку не только вдыхать и выдыхать, но также смеяться, чихать и кашлять. Дельты на плечах помогают поднять руки над головой. Сокращение бицепса и трицепса помогает сгибать и разгибать руку в локте. Ягодичные мышцы помогают стабилизировать бедра при ходьбе и беге.
Осанка и баланс тела
Скелетные мышцы помогают поддерживать осанку тела. Например, ягодичная мышца также помогает встать и поддерживать прямую позу.Портняжная мышца бедра — самая длинная мышца тела. Эта мышца вместе с четырьмя другими мышцами четырехглавой мышцы помогает в движении ноги. Мало того, они также помогают поддерживать баланс тела.
Опора для мягких тканей
Висцеральные органы и другие внутренние ткани, а также очень нежные органы поддерживаются скелетными мышцами. Брюшная стенка и полость таза состоят из тканей скелетных мышц. Эти ткани не только поддерживают вес внутренних органов, но и защищают их от возможных травм.
Помогите поддержать точки входа и выхода тела
Различные отверстия тела, такие как рот, анус, мочевыводящие пути и т. Д., Покрываются мышцами сфинктера. Эти мышцы расположены концентрически вокруг этих отверстий. Когда эти мышцы сокращаются, их раскрытие становится меньше. Таким образом, они помогают при глотании пищи, мочеиспускании и дефекации.
Температура тела
Скелетные мышцы помогают поддерживать температуру тела, поскольку их сокращения превращают энергию в тепло.Таким образом, после выполнения напряженной деятельности часто бывает очень жарко, так как тело выделяет тепло из-за действия системы скелетных мышц.
Это были некоторые из функций скелетных мышц. Как видите, без скелетных мышц наше тело не сможет поддерживать себя и сохранять осанку. Каждое произвольное и в некоторых случаях непроизвольное движение обусловлено действием ткани скелетных мышц.
Нравится? Поделиться этим!
Анатомия и физиология скелетных мышц
Скелетная мышца — сложный орган.Чтобы оптимизировать режим тренировки, необходимо понимание анатомии скелетных мышц.
Анатомия скелетных мышц
Ткань скелетных мышц окружена соединительной тканью. Он отделен от кожи поверхностной фасцией, также известной как подкожный слой, который состоит из соединительной ткани и жировой (жировой) ткани [1] .
Жировая ткань в этой поверхностной фасции является основным хранилищем триглицеридов в организме и служит защитным слоем для мышц.Кровеносные сосуды, лимфатические сосуды и нервы входят в мышцы и выходят из них через этот слой [1] .
Под поверхностной фасцией находится глубокая фасция. Глубокая фасция — это соединительная ткань неправильной формы, которая удерживает вместе функционирующие друг с другом мышцы [1] .
Ниже глубокой фасции находятся три слоя соединительной ткани, укрепляющие мышцы:
- Эпимизий — Самый внешний слой. Окружает всю мышцу.
- Перимизий — окружает группы мышечных волокон, называемых пучками.
- Эндомизий — Самый глубокий слой. Разделяет отдельные мышечные волокна.
Скелетные мышцы состоят из множества отдельных клеток, известных как мышечные волокна. Мышечные волокна имеют несколько ядер и расположены ниже плазматической мембраны, называемой сарколеммой. Т (поперечные) канальцы — это крошечные инвагинации, которые идут от внешней поверхности сарколеммы в центр мышечного волокна.Эти Т-канальцы распространяют потенциалы действия по всему мышечному волокну, что вызывает сокращение мышц.
Внутри сарколеммы находится цитоплазма волокна, называемая саркоплазмой. Саркоплазма содержит запасные вещества, такие как гликоген и кислород, необходимые для сокращения мышц.
Каждое отдельное мышечное волокно, в свою очередь, состоит из множества миофибрилл меньшего размера. Миофибриллы делятся на повторяющиеся функциональные единицы, называемые саркомерами. Этот повторяющийся узор придает скелетным мышцам полосатый вид.Система мембранных мешочков, называемая саркоплазматической сетью (SR), окружает миофибриллы. Ионы кальция (Ca2 +), запускающие сокращение мышц, хранятся в саркоплазматическом ретикулуме.
Три типа белков образуют миофибриллы (1) сократительные, (2) регуляторные и (3) структурные [1] . Сократительные белки являются генераторами силы сокращения мышц. Два сократительных белка в миофибриллах — это актин и миозин, которые являются частью тонкого волокна и толстого волокна соответственно.
Миозиновая нить далее классифицируется как моторный белок, потому что она создает силу или движения за счет использования химической энергии, хранящейся в молекулах АТФ [1] .
Нить миозина часто описывается как форма «двух клюшек для гольфа, скрученных вместе» с «ручками клюшек», представляющими миозиновый хвост, и «головками клюшек», представляющими головки миозина или поперечные мостики [1] .
Миозиновый поперечный мостик (есть много поперечных мостиков на миозиновой нити) имеет сайт прикрепления для актина и АТФ (который будет обсужден позже).Головки миозина выступают из миозинового хвоста к окружающим актиновым филаментам. Спиральная актиновая нить, состоящая из отдельных молекул актина, каждая из которых содержит сайт связывания миозина, к которому могут прикрепляться миозиновые головки, прикреплена к Z дискам **.
Два регуляторных белка — тропонин и тропомизий, которые также являются частью тонкой нити, участвуют в включении или выключении мышечного сокращения [1] ».
Когда мышца расслаблена, тропомизий блокирует миозин-связывающие участки на белках актина, поэтому миозиновые головки не могут прикрепиться, и, следовательно, мышца не может сокращаться.Тропонин удерживает белки тропомизия на месте.
Когда кальций входит в цитоплазму мышечного волокна, он может связываться с молекулой тропонина, которая изменяет форму молекулы тропонина и «оттягивает» тропомизий от сайта связывания миозина на каждой молекуле актина [2] . Когда кальция больше нет, молекула тропонина вновь принимает свою первоначальную форму, и тропомизий снова блокирует сайт связывания.
Структурные белки участвуют в стабильности и эластичности миофибрилл.Наиболее заметным из структурных белков является тайтин. Белок тайтин в 50 раз больше, чем средний белок [1] , и простирается от Z диска до линии M (половина саркомера).
Титин «прикрепляет» толстую нить накала к диску Z и линии M, стабилизируя ее положение. [1] . Белок тайтин также очень эластичен и помогает растянутой или сокращенной мышце вернуться к ее расслабленной длине [1]. В каждой миофибрилле есть около десятка других структурных белков.
** Саркомер делится на различные части в зависимости от присутствующих волокон.
- Z Line (диски) — закрепляет актиновые филаменты. Разделяет саркомеры.
- I Band — Содержит часть актиновых волокон, которые не перекрывают миозиновые волокна.
- Группа A — охватывает всю миозиновую нить.
- H Зона — содержит часть миозиновой нити, которая не перекрывается актиновыми нитями.
- M Band — Середина саркомера.
Сокращение мышц
Когда мышца сокращается, активируются миозиновые поперечные мостики. Это не означает, что мышечные волокна укорачиваются, это скорее механизм, который генерирует силу и напряжение, активен [2] .
Мышца сокращается, когда гантель удерживается в одном положении, в то время как мышца не укорачивается и не удлиняется. В большинстве случаев, например, при сгибании гантели, мышца (в данном примере двуглавая мышца плеча) сокращается при сокращении.Это описано в разделе «Механизм скользящей подачи».
Механизм скольжения с нитью
Когда мышца укорачивается во время сокращения, миозиновый мостик прикрепляется к актиновой нити. Перемещение поперечного моста часто описывается как движение по дуге, как гребля лодочного весла [2] , тянет две последовательные Z-линии к центру саркомера.
Каждый отдельный «ход» поперечного моста вызывает только небольшое движение (подтягивание линий Z к центру), но если мышца остается активированной, поперечный мост продолжает свое поглаживающее движение, что приводит к большему движению. [2] .Когда мышца сокращается, образуется много поперечных мостиков. Процесс крепления и перемещения поперечного моста известен как цикл поперечного моста.
Мостовой цикл
Цикл поперечного мостика, инициируемый, когда кальций из SR попадает в цитоплазму, состоит из четырех этапов [2] :
- Миозиновый мостик прикрепляется к актиновой нити A + M · ADP · Pi »A · M · ADP · Pi
- Поперечные мосты перемещаются, создавая напряжение A · M · ADP · Pi »A · M + ADP + Pi
- Поперечный мостик отделяется от актиновой нити A · M + ATP »A + M · ATP (связывание ATP отделяет миозин от актина)
- Поперечный мостик «заряжается» и снова присоединяется к актиновой нити, если кальций все еще присутствует. A + M · ATP »A + M · ADP · Pi
* A = Актин; M = миозиновый мостик; · = Привязан к
Важно отметить, что каждый поперечный мост проходит этот цикл независимо от других поперечных мостов [2] .Следовательно, во время мышечного сокращения одни поперечные мостики прикрепляются к актиновой нити, а другие — нет.
АТФ обеспечивает энергию для движения поперечного моста при его гидролизе. Он также разрывает связь между миозином и актином, когда связывается с миозином.
АТФ требуется для сокращения мышц. Кальций также необходим для сокращения мышц. Это инициирует собственно физическое сокращение. Высвобождение кальция из SR запускается потенциалом действия.
Нервно-мышечные сигналы
Мышечные сокращения сигнализируются потенциалами действия (электрическими сигналами) в плазматических мембранах мышечных волокон.Этот сигнал передается нервными клетками, известными как нейроны, от центральной нервной системы (ЦНС) к мышечным волокнам. Нервные клетки, которые иннервируют мышечные волокна, называются двигательными нейронами.
Моторный нейрон берет начало в ЦНС и достигает мышцы, где разделяется на несколько ветвей. Каждая ветвь образует соединение с одним мышечным волокном. Моторный нейрон плюс все мышечные волокна, иннервируемые его ветвями, известен как двигательная единица [2] .
Когда моторный нейрон передает (передает) потенциал действия от ЦНС к мышечным волокнам, которые он иннервирует, все волокна в его двигательной единице сокращаются. [2] .
Когда аксон достигает мышечного волокна, он распадается на «короткие отростки», которые встраиваются в поверхность мышечного волокна [2] . Терминалы аксона содержат везикулы, содержащие нейромедиатор ацетилхолин (ACh). Плазматическая мембрана мышечного волокна под окончанием аксона известна как моторная концевая пластинка [2] .
Терминал аксона и концевая пластинка двигателя образуют соединение, известное как нервно-мышечное соединение. Когда потенциал действия распространяется на терминал аксона моторного нейрона, он деполяризует его, что открывает чувствительные к напряжению кальциевые каналы, позволяя Ca2 + во внеклеточной жидкости проникать в терминал аксона.Этот вход Ca2 + сигнализирует о высвобождении ACh из везикул в конце аксона.
Затем ACh диффундирует от терминала аксона через нервно-мышечное соединение к моторной концевой пластине и связывается с рецепторами ACh. Это связывание ACh открывает каналы для ионов, через которые может поступать натрий (Na +).
Существует электрохимический градиент на плазматической мембране мышечного волокна, контролируемый концентрацией ионов с обеих сторон мембраны. Потенциал покоя (отсутствие электрического сигнала) мышечного волокна отрицательно по отношению к внеклеточной жидкости.
Открытие ионных каналов и движение ионов вызывает деполяризацию мембраны (мембранный потенциал становится менее отрицательным) и создает потенциал концевой пластинки (EPP) [2] . EPP деполяризует плазматическую мембрану, прилегающую к моторной концевой пластине, вызывая распространение потенциала действия по всему мышечному волокну и вдоль Т-канальцев.
Потенциал действия в Т-канальцах запускает высвобождение Са2 + из SR. Этот Ca2 + связывается с регуляторным белком тропонином и инициирует сокращение мышц, как описано выше.Перекрестный мостовой цикл может продолжаться до тех пор, пока Са2 + связан с тропонином.
Понимая, как сокращаются мышцы, мы теперь можем изучить другие аспекты физиологии упражнений, биомеханики и моторного контроля.
Ссылки
- Джерард Дж. Тортора. Принципы анатомии человека (9-е изд.)
- Видмайдер, Э. Рафф, Х., Стренг, К. Физиология человека Механизм функции тела. (9-е изд.)
Скелетная мышца
клетки развиваются, когда сотни отдельных эмбриональных клеток называются миобласты предохранитель.В результате клеточная мембрана клеток
называется сарколемма , а цитоплазма называется саркоплазма. Некоторые уникальные характеристики скелета
мышечные клетки (волокна):
| ||||||||
Миофибриллы и миофиламенты
| ||||||||
| миофибриллы
состоят из миофиламентов. Миофиламенты
организованы в повторяющиеся единицы по длине миофибриллы
называется саркомера. | ||||||||
Организация саркомеров
| ||||||||
Тонкие нити
| ||||||||
Толстые нити
|
Протеом человека в скелетных мышцах
АНДНО
Поле
Все название гена Класс белка Uniprot ключевое слово Хромосома Внешний идентификатор Оценка надежности ткань (IHC) Оценка надежности мышиный мозг Оценка надежности клеток (ICC) Белковый массив (PA) Вестерн-блот (WB) Иммуногистохимия (IHC) Иммуноцитохимия (ICC) Местоположение секретома Аннотация субклеточного местоположения (ICC) Субклеточная линия (ICC) Фаза пика субклеточного клеточного цикла Экспрессия ткани (IHC) Категория ткани (РНК) Категория типа клеток (РНК) Категория линии клеток (РНК) Категория рака (РНК) Категория области мозга (РНК) Категория клеток крови (РНК) Категория клеток крови (РНК) Категория мозга мыши (РНК) Категория мозга свиньи (РНК) Прогностический рак Метаболический путьСводка доказательств Доказательства UniProt Нет доказательствProt Доказательства HPA Доказательства MSС антителами Имеются данные о белках Сортировать по
Класс
Антигенные белки группы крови Гены, связанные с раком, гены-кандидаты сердечно-сосудистых заболеваний, маркеры CD, белки, связанные с циклом лимонной кислоты, гены, связанные с болезнями, ферменты, одобренные FDA лекарственные мишени, рецепторы, связанные с G-белками, сопоставленные с neXtProt, сопоставленные с UniProt, SWISS-PROT, ядерные рецепторы, белки, связанные с плазматической мембраной, прогнозируемые белки, секретируемые клетками, прогнозируемые белки, секретирующие лекарственные средства, прогнозируемые белки, секретируемые через плазму, белки, секретируемые через RAS белки Рибосомные белки Белки, родственные РНК-полимеразе Факторы транскрипции Транспортеры Ионные каналы, управляемые напряжением
Подкласс
Класс
Биологический процесс Молекулярная функция Болезнь
Ключевое слово
Хромосома
12345678910111213141516171819202122MTUnmappedXY
Надежность
ПовышеннаяПоддерживается УтвержденоНеопределено
Надежность
Поддерживается Одобрено
Надежность
ПовышеннаяПоддерживается УтвержденоНеопределено
Проверка
Поддерживается УтвержденоНеопределено
Проверка
Enhanced — CaptureEnhanced — GeneticEnhanced — IndependentEnhanced — OrthogonalEnhanced — РекомбинантнаяПоддерживаемаяПодтвержденнаяНеопределенная
Проверка
Enhanced — IndependentEnhanced — OrthogonalSupportedApprovedUncertain
Проверка
Enhanced — GeneticEnhanced — IndependentEnhanced — РекомбинантнаяПоддерживается УтвержденоНеопределено
Аннотация
Внутриклеточно и мембранно, секретно — неизвестное местоположение, секретируется в мозге, секретируется в женской репродуктивной системе, секретируется в мужской репродуктивной системе, секретируется в других тканях, секретируется в кровь, секретируется в пищеварительную систему, секретируется во внеклеточный матрикс
Расположение
актина filamentsAggresomeCell JunctionsCentriolar satelliteCentrosomeCleavage furrowCytokinetic bridgeCytoplasmic bodiesCytosolEndoplasmic reticulumEndosomesFocal адгезия sitesGolgi apparatusIntermediate filamentsKinetochoreLipid dropletsLysosomesMicrotubule endsMicrotubulesMidbodyMidbody ringMitochondriaMitotic chromosomeMitotic spindleNuclear bodiesNuclear membraneNuclear specklesNucleoliNucleoli фибриллярный centerNucleoli rimNucleoplasmPeroxisomesPlasma membraneRods & RingsVesicles
Поиск
РасширенныйПоддерживаемый УтвержденоНеизвестноВариация интенсивностиПространственная вариацияКорреляция интенсивности клеточного цикла Пространственная корреляция клеточного цикла Биологический цикл клеток Пользовательские данные, зависящие от клеточного циклаМультилокализацияЛокализация 1Локация 2Локализация 3Локализация 4Локализация 5Локализация 6Главное расположениеДополнительное местоположение
Расположение
AnyActin filamentsAggresomeCell JunctionsCentriolar satelliteCentrosomeCleavage furrowCytokinetic bridgeCytoplasmic bodiesCytosolEndoplasmic reticulumEndosomesFocal адгезия sitesGolgi apparatusIntermediate filamentsKinetochoreLipid dropletsLysosomesMicrotubule endsMicrotubulesMidbodyMidbody ringMitochondriaMitotic chromosomeMitotic spindleNuclear bodiesNuclear membraneNuclear specklesNucleoliNucleoli фибриллярный centerNucleoli rimNucleoplasmPeroxisomesPlasma membraneRods & RingsVesicles
Клеточная линия
анйа-431A549AF22ASC TERT1BJCACO-2EFO-21FHDF / TERT166GAMGHaCaTHAP1HBEC3-KTHBF TERT88HDLM-2HEK 293HELHeLaHep G2HTCEpiHTEC / SVTERT24-BHTERT-HME1HTERT-RPE1HUVEC TERT2JURKATK-562LHCN-M2MCF7NB-4OE19PC-3REHRH-30RPTEC TERT1RT4SH-SY5YSiHaSK-MEL-30SuSaTHP-1U-2 ОСУ-251 МГ
Тип
ProteinRna
Фаза
G1SG2M
Ткань
AnyAdipose tissueAdrenal glandAppendixBone marrowBreastBronchusCartilageCaudateCerebellumCerebral cortexCervix, uterineChoroid plexusColonDorsal rapheDuodenumEndometriumEpididymisEsophagusEyeFallopian tubeGallbladderHairHeart muscleHippocampusHypothalamusKidneyLactating breastLiverLungLymph nodeNasopharynxOral mucosaOvaryPancreasParathyroid glandPituitary glandPlacentaProstateRectumRetinaSalivary glandSeminal vesicleSkeletal muscleSkinSmall intestineSmooth muscleSoft tissueSole из footSpleenStomachSubstantia nigraTestisThymusThyroid glandTonsilUrinary bladderVagina
Тип ячейки
Выражение
Не обнаружено Низкое Среднее Высокое
Ткань
AnyAdipose tissueAdrenal glandBloodBone marrowBrainBreastCervix, uterineDuctus deferensEndometriumEpididymisEsophagusFallopian tubeGallbladderHeart muscleIntestineKidneyLiverLungLymphoid tissueOvaryPancreasParathyroid glandPituitary glandPlacentaProstateRetinaSalivary glandSeminal vesicleSkeletal muscleSkinSmooth muscleStomachTestisThyroid glandTongueUrinary bladderVagina
Категория
Обогащенная ткань Обогащенная группа Улучшенная ткань Низкая тканевая специфичность Не обнаружено Обнаружено у всех Обнаружено во многих Обнаружено примерно в
ОРГАНИЗАЦИЯ СКЕЛЕТНЫХ МЫШЦ
ОРГАНИЗАЦИЯ СКЕЛЕТНЫХ МЫШЦ
Перед тем, как завершить чтение, вы должны заполнить следующее:
Шаг 01
Загрузите изображение размера страницы «Организация мышц.
Шаг 02
Прочтите руководство, которое поможет вам пометить изображение.
Шаг 03
Имея в руках изображение с новой меткой, прочтите следующие абзацы.
Для тех, кто не может участвовать в описанных выше шагах, было предоставлено альтернативное задание по организации скелетных мышц, дающее вам всю информацию, содержащуюся в интерактивном руководстве. ПРИМЕЧАНИЕ. Это альтернативное назначение временно отключено.
Клетки скелетных мышц — общая и микроскопическая структура
Каждая клетка скелетных мышц, также называемая мышечным волокном , развивает столько же эмбриональных миоцитов , слитых в одну длинную многоядерную клетку скелетных мышц. Эти мышечные волокна связаны в пучки, или пучки , и снабжены богатой сетью кровеносных сосудов и нервов. Затем пучки связываются вместе, образуя неповрежденную мышцу.Давайте рассечем скелетную мышцу, начиная с мышцы в целом снаружи и продолжая внутри до субмикроскопического уровня отдельной мышечной клетки.
В неповрежденной скелетной мышце богатая сеть нервов и кровеносных сосудов питает и контролирует каждую мышечную клетку. Эти мышечные волокна индивидуально обернуты, а затем связаны между собой несколькими различными слоями волокнистой соединительной ткани.
Эпимизий («эпимизий» — снаружи и «мизиум» — мышца) представляет собой слой плотной волокнистой соединительной ткани, которая окружает всю мышцу.Этот слой также часто называют лицевой панелью . Каждая скелетная мышца образована из нескольких связанных пучков волокон скелетных мышц, и каждый пучок окружен перимизием (перимизий) («перимизий»). Каждая мышечная клетка индивидуально обернута тонким слоем рыхлой (ареолярной) соединительной ткани, называемой endomysium («эндо» -внутри). Эти слои соединительной ткани непрерывны друг с другом, и все они выходят за пределы самих мышечных волокон, образуя сухожилия , которые прикрепляют мышцы к кости, перемещая кости при сокращении мышц.
Глубоко до эндомизия каждая клетка скелетных мышц окружена клеточной мембраной, известной как сарколемма (в этом обсуждении вы увидите префиксы sarc- и myo- , так что вы должны понимать, что эти префиксы, относящиеся к «мышце»). Цитоплазма или саркоплазма содержит большое количество гликогена, (форма хранения глюкозы) для получения энергии и миоглобина, — красного пигмента, похожего на гемоглобин, который может накапливать кислород.Однако большая часть внутриклеточного пространства занята стержнеобразными миофибриллами — цилиндрическими белковыми структурами. Каждое мышечное волокно содержит сотни или даже тысячи миофибрилл, которые проходят от одного конца каждого мышечного волокна до другого. Эти миофибриллы занимают около 80% внутриклеточного пространства и настолько плотно упакованы внутри этих клеток, что митохондрии и другие органеллы оказываются зажатыми между ними, в то время как ядра выталкиваются наружу и располагаются на периферии прямо под сарколеммой.
Каждая миофибрилла состоит из нескольких разновидностей белковых молекул, которые образуют миофиламенты , и каждая миофиламент содержит сократительные сегменты, обеспечивающие сокращение. Эти сократительные сегменты известны как саркомера (« sarc-» — мускул, « просто » — часть). Полоски, видимые микроскопически в волокнах скелетных мышц, образованы регулярным организованным расположением миофиламентов — во многом подобно тому, что мы увидели бы, если бы нарисовали полосы на палочках для еды и связали их вместе полиэтиленовой пленкой, при этом пластик представляет сарколемму.
Строчки, видимые под микроскопом в скелетных мышцах, образованы правильным расположением белков внутри клеток. Обратите внимание, что в каждой ячейке есть светлые и темные полосы. Темные области называются полосами A , что довольно легко запомнить, потому что «a» — вторая буква в слове «dark». Светлые области называются полосами I , и их также легко запомнить, потому что «i» — вторая буква в слове «свет». («А» означает анизотропный, а «I» — изотропный.Оба эти термина относятся к светопоглощающему характеру каждой полосы. Однако мы будем придерживаться полос A и I.) На изображении ниже показана микрофотография саркомера вместе с рисунком, представляющим различные части саркомера.
Название: Файл: Sarcomere.gif; Автор: Sameerb; Сайт: https://commons.wikimedia.org/wiki/File:Sarcomere.gif; Лицензия: Public Domain, без ограничений
Обратите внимание, что в середине каждой полосы I есть более темная линия, называемая линией z линией или z диском. Линии Z — это перегородки между соседними саркомерами. Саркомеры соединяются встык по всей длине миофибриллы. Кроме того, в середине каждой полосы A находится более светлая зона H (H для «helle» — «яркий»), и каждая зона H имеет более темную линию M (M для «середины»), идущую прямо вниз по середина полосы А.
Название: 1003_Thick_and_Thin_Filaments.jpg; Автор: OpenStax College; Сайт: http://cnx.org/contents/6df8aab3-1741-4016-b5a9-ac51b52fade0@3/Skeletal-Muscle; Лицензия: под лицензией Creative Commons Attribution 4.0 Лицензия.
Каждая миофибрилла, в свою очередь, содержит несколько разновидностей белковых молекул, называемых миофиламентами . Более крупные или толстые миофиламенты состоят из белка, миозина , и меньшие тонкие миофиламенты в основном состоят из белка актина.
Давайте обсудим каждую миофиламент по очереди. Каждая молекула актина состоит из двух цепей из молекул фиброзного актина (F-актин) и ряда из молекул тропонина и молекул тропомиозина .Каждый F-актин образован двумя цепочками из глобулярного актина (G-актин) , скрученных вместе в двойную спиральную структуру, очень похожую на скручивание двух нитей жемчуга друг с другом. Каждая молекула G-актина будет представлена жемчужиной на нашем гипотетическом ожерелье. Каждая субъединица G-актина имеет сайт связывания, с которым миозиновая головка прикрепляется к актину. Тропомиозин представляет собой длинную цепочку-подобный полипептид, который параллелен каждой цепи актина F и функционирует, чтобы либо скрыть, либо обнажить «активные центры» на каждой глобулярной молекуле актина.Каждая молекула тропомиозина достаточно длинна, чтобы покрывать активные сайты связывания на семи молекулах G-актина. Эти белки проходят от конца до конца по всей длине F-актина. С каждой молекулой тропомиозина связан третий полипептидный комплекс, известный как тропонин. Комплексы тропонина содержат три глобулярных полипептида (тропонин I, тропонин T и тропонин C), которые выполняют различные функции. Тропонин I связывается с актином, тропонин T связывается с тропомиозином и помогает расположить его на нитях актина F, а тропонин C связывает ионы кальция.На каждый тропомиозин приходится один тропониновый комплекс. Когда кальций связывается с тропонином C, он вызывает конформационные изменения во всем комплексе, что приводит к экспонированию сайтов связывания миозина на субъединицах G-актина. Подробнее об этом позже.
Толстые миофиламенты состоят в основном из белка миозина , , , и каждая толстая миофиламент состоит из примерно 300 молекул миозина, связанных вместе. Каждый миозин состоит из 6 белковых субъединиц, 2 тяжелых цепи, и легких цепей. Тяжелые цепи имеют форму, подобную клюшке для гольфа, имеют длинную стержневидную структуру, с которой соединена глобулярная миозиновая головка. Стержни или хвосты охватывают друг друга и взаимодействуют с хвостами других молекул миозина, образуя стержень толстой нити. Шаровидные головки выступают под прямым углом к валу. Голова половины молекул миозина ориентирована к одному концу толстого волокна, а другая половина — в противоположном направлении. Именно головки миозина связываются с активными участками актина.Соединение между головкой и стержнем молекул миозина функционирует как шарнир и, как таковое, называется шарнирной областью . Шарнирная область может сгибаться и, как мы увидим позже, создает силовой удар при сокращении мышцы. Центр толстых нитей состоит только из стержней тяжелых цепей. Кроме того, каждая миозиновая головка имеет АТФазу, которая связывается и гидролизует АТФ во время сокращения мышц. Именно АТФ обеспечивает энергию для сокращения мышц.Каждая из миозиновых головок связана с двумя легкими цепями миозина, которые играют роль в регулировании действий миозиновых головок, но точный механизм полностью не изучен. Трехмерное расположение головок миозина очень важно. Представьте, что вы смотрите на толстую нить с конца и что вверх торчит миозиновая головка. Двигаясь по окружности толстой нити, вы будете видеть миозиновые головки каждые 30 градусов. Это позволяет каждой толстой нити взаимодействовать с 6 тонкими нитями.Точно так же каждая тонкая нить может взаимодействовать с тремя толстыми нитями. Такое расположение требует наличия двух тонких волокон на каждую толстую нить в миофибрилле (см. Изображение ниже).
Адаптировано из следующего изображения: Название: 1022_Muscle_Fibers_ (small) .jpg; Автор: OpenStax College; Сайт: http://cnx.org/contents/6df8aab3-1741-4016-b5a9-ac51b52fade0@3/Skeletal-Muscle; Лицензия: под лицензией Creative Commons Attribution 4.0 License.
Во время сокращения мышцы головки миозина связывают вместе толстые и тонкие миофиламенты, образуя поперечных мостика , которые заставляют толстые и тонкие миофиламенты скользить друг по другу, что приводит к укорачиванию каждого саркомера, каждого волокна скелетной мышцы и мышцы как целиком, как две части приставной лестницы, скользящие друг по другу.Подводя итог, можно сказать, что для того, чтобы произошло укорачивание мышцы, головки миозина обладают тремя важными свойствами: 1. Головки могут связываться с активными участками молекул G-актина, образуя поперечных мостика. 2. Головки прикреплены к стержневидным частям молекул тяжелого миозина шарнирной областью , как уже обсуждалось, и 3. Головки содержат ферментов АТФаза , которые могут расщеплять АТФ, используя полученную энергию для изгиба шарнирной области и позволяет отделить миозиновые головки от актина.
Z-линия (или Z-диск) состоит из белков ( альфа-актинин, ), которые обеспечивают место прикрепления тонких филаментов. Точно так же М-линия состоит из белков (миомезина), которые удерживают молекулы миозина на месте. Полоса A образована молекулами миозина, а полоса I — это место, где тонкие волокна не перекрывают толстые волокна. Зона H — это та часть полосы A, где толстые и тонкие волокна не перекрываются.
Есть еще один важный структурный белок, который простирается от диска Z до линии M, проходя внутри толстого филамента.Из-за своего большого размера этот белок получил название тайтин (тайтин является крупнейшим известным белком в организме человека и содержит примерно 30 000 аминокислот). Он образует сердцевину толстых миофиламентов, удерживая их на месте и, таким образом, удерживая группу A. Кроме того, тайтин обладает способностью растягиваться и отскакивать, а также предотвращает чрезмерное растяжение и повреждение мышцы, а также возвращает ее к нормальной длине, когда мышца растягивается сверх своей нормальной длины в состоянии покоя. Напомним, что одним из свойств мышцы является ее эластичность, за это свойство отвечает титин.
Есть несколько других важных структурных белков, но мы остановимся только на одном: dystrophin . Дистрофин — это белок, расположенный между сарколеммой и крайними миофиламентами. Он связывает актин с интегральным мембранным белком, который, в свою очередь, связывает мышечную клетку с эндомизием всего мышечного волокна. Генетическая мутация гена, кодирующего дистрофин, является одной из коренных причин класса мышечных заболеваний, известных под общим названием мышечная дистрофия (MD) .Наиболее распространенной формой МД является мышечная дистрофия Дюшена (МДД), которая наследуется по типу «сцепленного с полом» и поражает мальчиков. Большинство пациентов с МДД привязываются к инвалидной коляске в раннем возрасте, обычно к 12 годам или около того. Затрудненное дыхание обычно становится проблемой к 20 годам и часто является причиной их преждевременной смерти.
Саркоплазматический ретикулум и Т-канальцы
© Encyclopædia Britannica, Inc., 2013. Загружена и помечена из графического квеста BYU-Idaho, декабрь 2013 г.
Заголовок: 1023_T-tubule.jpg; Автор: OpenStax College; Сайт: http://cnx.org/contents/6df8aab3-1741-4016-b5a9-ac51b52fade0@3/Skeletal-Muscle; Лицензия: под лицензией Creative Commons Attribution 4.0 License.
В волокнах скелетных мышц есть два набора канальцев, которые выполняют важные функции во время мышечных сокращений: саркоплазматический ретикулум и Т-канальцы.
Т-канальцы (поперечные канальцы) — это инвагинации или углубления сарколеммы.Они похожи на молодого придирчивого едока, продырявленного в картофельном пюре. Т-канальцы сообщаются с внеклеточным пространством и заполнены внеклеточной жидкостью. Они расположены на саркомере в точках перекрытия полосы A и полосы I. Т-канальцы с обеих сторон фланкированы расширенными участками эндоплазматического ретикулума клетки — саркоплазматической сетью. Саркоплазматический ретикулум (SR) представляет собой сложную сеть гладких эндоплазматических ретикулумов, которые окружают и покрывают каждую миофибриллу, подобно тому, как свободно связанный свитер покрывает ваши руки.Он накапливает кальций, который затем может высвобождаться в саркоплазму, когда потенциал действия проводится по сарколемме Т-канальца. Большая часть саркоплазматического ретикулума проходит параллельно миофибриллам, но есть прямоугольные увеличения SR на стыках полос A-I, которые фланкируют Т-канальцы. Эти увеличения известны как терминальные цистерны («концевые мешочки») (см. Изображение выше). Один Т-канальец вдоль двух терминальных цистерн, которые параллельны ему, образуют триаду .Триада имеет решающее значение для функции скелетных мышц. В каждой триаде мембрана Т-канальца содержит большое количество потенциал-зависимых белков, называемых дигидропиридиновыми (DHP) каналами или кальциевыми каналами L-типа. Хотя они называются каналами, они не позволяют кальцию перемещаться по ним; скорее, они физически связаны с каналами высвобождения кальция на терминальных цистернах, известных как каналы рецептора рианодина (RyR). Когда мембрана деполяризуется потенциалом действия, канал DHP обнаруживает деполяризацию и заставляет каналы RyR открываться, что приводит к высвобождению кальция из концевых цистерн SR.
