
ЖЕВАТЕЛЬНЫЕ МЫШЦЫ ЧЕЛОВЕКА их расположение, функции, кровоснабжение (Таблица)
Жевательные мышцы с каждой стороны связаны между собой генетически (по происхождению), морфологически (все пересекают один сустав и прикрепляются к нижней челюсти) и функционально (совершают жевательные движения – двигают нижнюю челюсть вверх, вперед-назад и в стороны)
|
Наименование мышц |
Начало |
Прикрепление |
Функция |
Кровоснабжение |
Иннервация |
|
1.Жевательная мышца (m.masseter) |
скуловая дуга |
угол и жевательная бугристость нижней челюсти |
поднимает опущенную нижнюю челюсть, выдвигает ее вперед |
челюстная артерия |
тройничный нерв |
|
2. |
поверхность чешуи височной кости |
венечный отросток нижней челюсти |
поднимает опущенную нижнюю челюсть, |
челюстная артерия |
—— |
|
3. Наружная рыловидная мышца (m.pterygoideuslateralis) |
подвисочный гребень, подвисочная область, крыловидные отростки |
нижнечелюстной сустав, шейка мыщелкового отростка нижней челюсти |
при двустороннем сокращении — выдвигает нижнюю челюсть вперед, при одностороннем – смещает ее в противоположную сторону |
челюстная артерия |
—— |
|
4. Внутренняя крыловидная мышца (m.pterygoideusmedialis) |
крыловидная ямка крыловидных отростков клиновидной кости |
крыловидная бугристость нижней челюсти |
—— |
—— |
—— |
 Височная мышца (m.temporalis)
Височная мышца (m.temporalis)Мышцы шеи и головы: анатомия, таблицы и функции
Мышцы шеи и головы человека классифицируются относительно выполняемого функционала. Функции мышц шеи очень разнообразны. Анатомия мышц лица и шеи, а также плечевого сплетения, представленная на презентации в энциклопедии и атласе с фото и таблицей, и дает представление о их структуре.
Функции мышц шеи очень разнообразны. Анатомия мышц лица и шеи, а также плечевого сплетения, представленная на презентации в энциклопедии и атласе с фото и таблицей, и дает представление о их структуре.
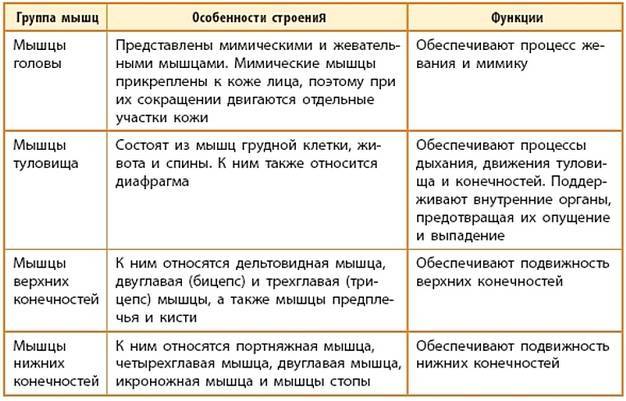
Данная мышца отличается сложной топографией и обусловлена разным строением, а также типом взаимоотношений с внутренними органами, кровеносными сосудами и нервами шеи. Анатомически мышцы головы и шеи и туловища подразделяются на обособленные группы.
Наклоны, подъемы и повороты головы выполняются за счет группы мышц шеи под названием поверхностные. Подобные функции выполняются при участии глубоких видов мышц. Эти группы мышц классифицируются как видимые.
Кроме них человек имеет средние, глотательные мышцы шеи, работающие при глотании, которые также участвуют в опускании нижней челюсти.
Разновидности мышц шеи
Анатомия мышц шеи обширна. Мышцы лица и шеи человека подразделяются на глубокие и поверхностные. Отдельно выделяются мышцы шеи крепящиеся к подъязычной кости и передней мышце шеи.
К поверхностным видам мышц также относятся подкожная шейная мышца и ключично-сосцевидная мышца. Она начинается от фасции груди в районе ключицы, и должна идти сверху боковой и передней поверхности шеи и закрепляться в нижней лицевой части. Сосцевидные шейные мышцы наиболее большие и сильные из все шейных мышц воротниковой зоны.
Мышца шеи и спины откидывающая назад голову называется грудино-ключично сосцевидная. Наклоны шеи и повороты головы в разные стороны происходят путем одностороннего сокращения этой мышцы. Двухстороннее сокращение держит голову вертикально, а при максимальном – запрокидывает назад. Эти мышцы туловища и мышца спины, позволяющая откидывать назад голову, выполняют так же несущую функцию. Мышца шеи и спины откидывающая назад голову дублирует функции широчайшей мышцы.
Среды мышц, непосредственно прикрепленных на подъязычной кости, есть мышцы, расположенные выше и ниже нее. Презентация первых и вторых насчитывает их по четыре вида.
К глубоким мышцам также относят переднюю, среднюю и заднюю лестничные мышцы, а также длинные и короткие шеи мышцы.
Этот крупный массив глубоких и поверхностных мышц шеи выполняет следующие основные функции:
• удерживает голову в состоянии равновесия;
• обеспечивает разностороннее движение шеи и головы;
• оказывает непосредственную помощь в процессах звукотворения и глотания.
Предпозвоночные мышцы шеи располагаются на боковой поверхности шейных позвонков. Мышцы шейного отдела позвоночника при двухстороннем сокращении выполняют функцию сгибания шейного отдела позвоночника. Иннервация мышц шеи проводится при помощи ветвей шейного сплетения и затылочными шейными нервами.
Нервы плечевого сплетения иннервируют кожу и мышцы верхних конечностей. Мышцы и нервы плечевого пояса начинаются в подключичной области плечевого сплетения, прилегающего к плечу.
Их функции в основном двигательные. Ветви плечевого сплетения снабжают мышцы плечевого пояса и шеи. Короткие ветви плечевого сплетения обеспечивают так же плечевые суставы.
Патологии мышц шеи
Мышцы головы, шеи и туловища работают во взаимосвязи и в ходе жизнедеятельности организма могут подвергаться разнообразным патологическим состояниям. Такое болезненное состояние, как ригидность мышц шеи характеризуется повышением тонуса мышц. Скованность и натяжение шейных мышц чаще всего вызываются сбоями в работе нервной системы, а также последствиями профессиональной деятельности.
К таким факторам можно отнести:
• длительные монотонные нагрузки;
• работа, требующая сохранения заданного положения долгое время;
• работа за рулем, конвейером и т.п.
Такие продолжительные ограничения подвижности ведут к спазму шейных и плечевых мышц, а также к постоянной ноющей боли в затылке. Чтобы восстановить нарушенное кровообращение рекомендуется разогревающая гимнастика для мускулатуры шейного отдела, затылка.
Явление гипертонуса
Гипертонус мышц шеи – это состояние напряженности, ригидности в мышцах сгибателях и разгибателях. Признаки гипертонуса – ноющая боль, отдающая в голову или область плеч, усиливающаяся при наклонах. Очень частым болевым синдромом есть скованность мышц шеи.
Причинами скованности шейных мышц могут быть патологические процессы воспалительного или травматического характера, а также повреждения нервных стволов с последующим нарушением процессов сокращения мышц. Если сводит мышцы шеи, то можно говорить о возникновении шейного спазма.
Причинами такого спазма может быть:
• гиперфункция щитовидной железы;
• нарушение мышечного баланса;
• реакция на медпрепараты;
• последствия физиологических изменений и т. п.
Дефанс мышц — напряжение мышечных волокон, главный диагностический критерий. Что делать при таком напряжении? Хорошие результаты дает гимнастика и массаж для всего тела.
Для мышц, которые могут называться вращательными, есть месторасположение в глубинных слоях мускулатуры.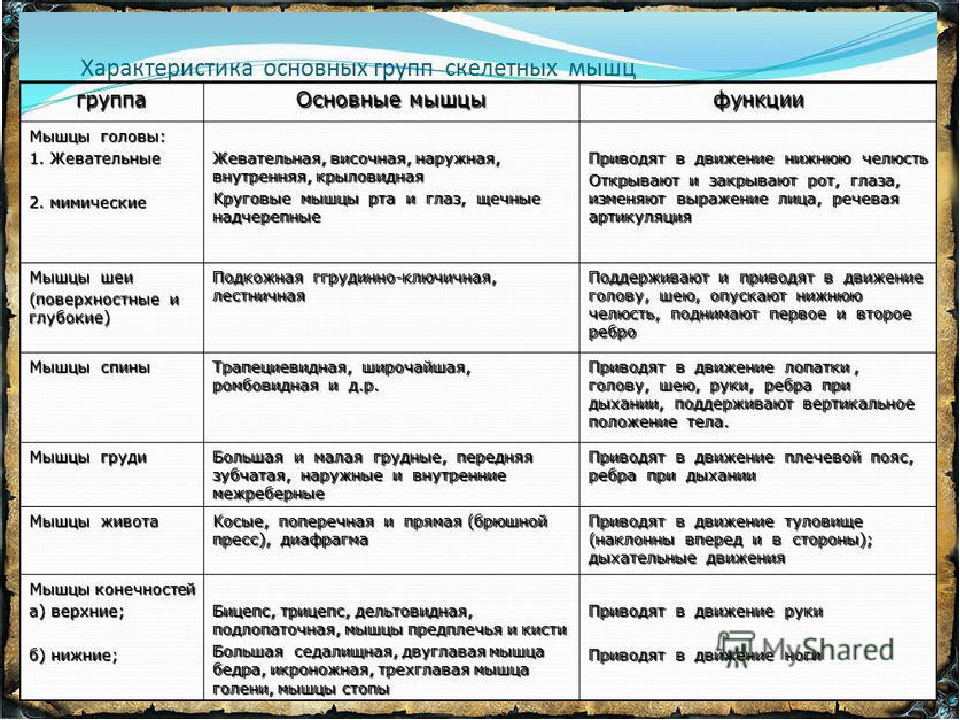 Мышца-вращатель шеи, груди или поясницы подразделяется на длинные и короткие и выполняет функцию поворота позвоночного столба и шейного отдела. Функциями широчайшей мышцы есть несущая и вспомогательная при движении людей. Наследственное заболевание Ландузи может свести к полной дистрофии мышцы плечевого сплетения и мышц туловища.
Мышца-вращатель шеи, груди или поясницы подразделяется на длинные и короткие и выполняет функцию поворота позвоночного столба и шейного отдела. Функциями широчайшей мышцы есть несущая и вспомогательная при движении людей. Наследственное заболевание Ландузи может свести к полной дистрофии мышцы плечевого сплетения и мышц туловища.
Методы лечения
Воспаление мышц шеи может возникать в результате переохлаждения или перенесенной большой физической нагрузки. Когда человек застудил мышцы шеи, то вследствие переохлаждения нарушается кровообращение и в мышечных тканях идут процессы воспаления. Что делать, если продуло и возникла ригидность шейных мышц? В таких случаях рекомендовано комплексное лечение.
К терапевтическим мероприятиям относят:
• прием болеутоляющих препаратов;
• жаропонижающие средства при необходимости;
• прогревающие процедуры;
• физиотерапевтические процедуры;
• полный покой пораженных участков.
При лечении мышца шеи требует полного покоя. До локализации воспалительных процессов старайтесь не напрягать простуженный шейный отдел, не поворачивать голову. Рекомендуется соблюдать постельный режим в течение приема медпрепаратов.
До локализации воспалительных процессов старайтесь не напрягать простуженный шейный отдел, не поворачивать голову. Рекомендуется соблюдать постельный режим в течение приема медпрепаратов.
Для снятия болевого синдрома можно принимать обычные анальгетики, а также нестероидные препараты с противовоспалительным действием, обязательно согласно рекомендации врача. На больное застуженное место на шее для уменьшения воспаления можно накладывать прогревающие компрессы и согревающие мази.
Самым простым является спиртовой компресс из х/б ткани, пропитанной спиртовым раствором, покрытый влагонепроницаемой пленкой и широким шерстяным шарфом. Также можно использовать прогревающие мази с пчелиным ядом. После снятия компресса тело пораженного участка должно находиться в сухом тепле. Хороший результат дает сухой компресс из разогретой пищевой соли в тканевом мешочке.
После снятия острого болевого синдрома шейные мышцы желательно разрабатывать для усиления кровообращения с помощью щадящего массажа шеи.
При наличии высокой температуры можно говорить о добавлении вирусной инфекции, требующей приема дополнительных антивирусных препаратов.
При лечении воспалений мышц шейного отдела наиболее подходят такие физиопроцедуры, как кварцевое прогревание, электрофорез, УВЧ. Для теста и достижения стойкого результата заболевшие люди должны пройти не менее 10 процедур. Укрепление мышц шеи с помощью гимнастики предотвращает боли и шейные остеохондрозы.
Наиболее часто встречающиеся простейшие, передающиеся с пищевыми продуктами
Наиболее часто встречающиеся простейшие, передающиеся с пищевыми продуктами
Согласно статистическим данным Всемирной организации здравоохранения, в мире более 4,5 млрд. человек заражены различными паразитами. В развивающихся странах, зараженность теми или иными паразитами может достигать 93-97%.
Существует огромное количество разнообразных паразитов, которые могут существовать во внутренних органах человека или населять его кожу и волосы. В том числе паразитарные заболевания могут вызывать и простейшие организмы, которые представляют группу одноклеточных или колониальных организмов. Большинство простейших — микроорганизмы, но некоторые могут достигают размеров в несколько миллиметров и хорошо видны невооружённым глазом. Настоящих многоклеточных форм среди простейших нет.
В том числе паразитарные заболевания могут вызывать и простейшие организмы, которые представляют группу одноклеточных или колониальных организмов. Большинство простейших — микроорганизмы, но некоторые могут достигают размеров в несколько миллиметров и хорошо видны невооружённым глазом. Настоящих многоклеточных форм среди простейших нет.
Способов попадания паразитов в организм человека довольно много, однако, одним из наиболее часто встречающихся путей заражения является заражение через пищевые продукты. Плохо промытые овощи, фрукты или зелень, недостаточно прожаренное или сваренное мясо, рыба, птица, неправильно обработанные продукты питания, могут привести к заражению простейшими.
Давайте же попробуем разобраться, какие простейшие, передаются с пищей и встречаются наиболее часто в нашей стране, а также как свести к минимуму риск заражения ими.
Простейшие:
Лямблиоз
Заболевание, вызывается лямблиями и поражающей кишечник человека. |
 Лямблиоз может протекать бессимптомно, в этом случае заболевание выявляется случайно, после проведения лабораторных исследований. Клинически заболевание проявляется умеренными болями в животе, вздутием живота, тошнотой, изжогой, расстройством стула. У человека, зараженного лямблиями, могут наблюдаться повышенная утомляемость, нарушение аппетита, снижение массы тела, бледность кожных покровов, мышечные подергивания.
Лямблиоз может протекать бессимптомно, в этом случае заболевание выявляется случайно, после проведения лабораторных исследований. Клинически заболевание проявляется умеренными болями в животе, вздутием живота, тошнотой, изжогой, расстройством стула. У человека, зараженного лямблиями, могут наблюдаться повышенная утомляемость, нарушение аппетита, снижение массы тела, бледность кожных покровов, мышечные подергивания. В случае гибели лямблий, продукты их распада всасываются из кишечника и способствуют возникновению различных форм аллергических реакций: кожного зуда, крапивницы, бронхиальной астмы.
Значительную роль в развитии заболевания играет состояние слизистой оболочки тонкой кишки и кислотность желудочного сока. У людей с пониженной кислотностью желудочного сока, нарушением слизистой оболочки кишечника, риск заражения лямблиозом, а также тяжесть клинических проявлений возрастают.
Балантидоз
Возбудителем балантидоза является балантидий кишечный — единственный представитель инфузорий, паразитирующий в толстом кишечнике человека. |
 Заболевание характеризуется изъязвлениями стенки толстой кишки с признаками общей интоксикации организма, но чаще протекает с незначительными клиническими проявлениями или вовсе бессимптомно. Источниками заболевания являются свиньи. Человек заболевает, как правило, при употреблении немытых овощей, ягод и зелени, где в качестве удобрений использовались свиные фекалии.
Заболевание характеризуется изъязвлениями стенки толстой кишки с признаками общей интоксикации организма, но чаще протекает с незначительными клиническими проявлениями или вовсе бессимптомно. Источниками заболевания являются свиньи. Человек заболевает, как правило, при употреблении немытых овощей, ягод и зелени, где в качестве удобрений использовались свиные фекалии. Возникновению признаков заболевания способствует ряд факторов: нарушение питания, снижение активности иммунной системы, переохлаждение организма, воспалительные заболевания кишечника и другие. В местах локализации паразитов возникает воспалительно-язвенный процесс, происходит отмирание клеток кишечника с образованием язв. У больных возникает общая интоксикация, появляются боли в животе, понос, в фекалиях – слизь и кровь. Наиболее опасными осложнениями балантидиаза являются прободение язв с развитием перитонита и кишечные кровотечения.
Амебиаз
Возбудителем амебиаза является дизентерийная амеба, обитающая в кишечнике человека. Заболеваемость амебиазом довольно высока — ежегодно регистрируются несколько сотен миллионов больных во всех странах. Для нескольких десятков тысяч больных заболевание заканчивается летальным исходом. В России эндемичными районами являются Закавказье, Калмыкия, Астраханская область и районы, примыкающие к среднеазиатским республикам. |
 Заболевание характеризуется частым водянистым стулом с примесью крови и слизи, болями в животе, лихорадкой и обезвоживанием организма.
Заболевание характеризуется частым водянистым стулом с примесью крови и слизи, болями в животе, лихорадкой и обезвоживанием организма. Если место обитания амебы в просвете толстой кишки, то паразитирование не приводит к возникновению клинических признаков заболевания. Если же дизентерийная амеба проникает в ткани слизистой оболочки толстой кишки, то образуются язвы. У больных появляются сильные боли в животе, развивается диарея (до 15 и более раз в сутки). Отличительный признак амебиаза – стул напоминает слизь и окрашен кровью («малиновое желе»). При разрушении сосудов стенки кишечника может возникнуть массивное кровотечение.
Больные амебной дизентерией подлежат обязательной госпитализации.
Токсоплазмоз
Возбудителем токсоплазмоза является токсоплазма. Заболевание характеризуется хроническим течением, поражением нервной системы, увеличением печени и селезенки, поражением скелетных мышц и миокарда. Цикл развития токсоплазмы довольно сложный: происходит не только со сменой хозяев, но и чередованием жизненных стадий паразита. Окончательные хозяева паразита – представители семейства кошачьих — американская рысь, бенгальский тигр и домашние кошки. |
Большинство случаев токсоплазмоза протекает бессимптомно. На характер проявлений заболевания существенное влияние оказывает общее состояние организма, а также активность иммунной системы.
У лиц с нарушениями иммунной системы заболевание приобретает особенно тяжелый характер. Увеличение количества очагов размножения паразита приводит к воспалительным изменениям в органах с последующим отмиранием тканей. Наиболее частое и грозное осложнение токсоплазмоза – энцефалит, летальность при котором достигает до 90%.
Наиболее частое и грозное осложнение токсоплазмоза – энцефалит, летальность при котором достигает до 90%.
Профилактика паразитозов
Чтобы не допустить заражения простейшими, нужно соблюдать несколько правил, а именно:
- Соблюдать простейшие правила личной гигиены, в том числе мытье рук после улицы, перед едой, после контакта с деньгами и животными.
- Стараться избегать контакта с бродячими животными, а при наличии у вас питомца регулярно посещать ветеринарную клинику. Обязательно проводить курсы дегельминтизации животных с периодичностью, рекомендуемой ветеринаром.
- Тщательно мыть фрукты, овощи, ягоды, даже если они с Вашего приусадебного участка.
- Не пить воду и не мыть посуду и продукты питания водой, в безопасности которой Вы не уверены.
- Употреблять в пищу только продукты, которые прошли необходимую термическую обработку.
- Обеспечить защиту от насекомых на Вашей кухне. Например, засетчивать оконные проемы, накрывать готовые блюда.

- Не есть вяленую или сушеную рыбу домашнего приготовления.
- Не употреблять домашнее соленое сало.
Проведение измерений при помощи медицинских динамометров: кистевого и станового.. Статьи. Поддержка. РАЗНОВЕС.РУ
Измерение силы и момента силы у людей проводят при помощи динамометров. Но не обычных, которые используются в промышленности, а специальных – медицинских. К медицинским динамометрам относят кистевой, также называемый ручным, динамометр и становой динамометр. В данной статье мы расскажем, как же проводят измерения при помощи данных приборов.
Итак, начнем с кистевого динамометра. Данный прибор предназначается для определения сжимающей силы мышц сгибающих пальцы обоих рук человека, а также для диагностики состояния и функции рук, как здоровых людей, так и восстанавливающихся после травм. Динамометр кистевой используется врачами, которые занимаются физиотерапией, кроме того, динамометр применяется в правоохранительных органах, вооруженных силах и МЧС. Приборы для измерения силы незаменимы для подготовки профессиональных спортсменов. В качестве примера кистевого динамометра можно привести такие приборы, как: механический ДК и электронный ДМЭР.
Приборы для измерения силы незаменимы для подготовки профессиональных спортсменов. В качестве примера кистевого динамометра можно привести такие приборы, как: механический ДК и электронный ДМЭР.
Динамометр кистевой ДК.
Для проведения измерений изометрической силы с использованием динамометра не требуется много времени, к тому же процесс замера не утомляет испытуемого. Для получения точных абсолютных результатов необходимо, чтобы пациент соблюдал определенное положение тела и угол отдельных суставов. Пусть обследуемый человек вытянет руку с кистевым динамометром и отведет её в сторону перпендикулярно туловищу. Свободная рука, при этом, должна быть расслаблена и опущена вниз. После чего, по команде, он должен будет сжать динамометр кистевой так сильно, как только сможет. Динамометрическое измерение может проходить поочередно обеими руками несколько раз, при этом, выбирается лучший результат для каждой руки.
Делать выводы на основании абсолютных результатов проведенных измерений можно только в динамике, когда предыдущие результаты были занесены в специальный дневник.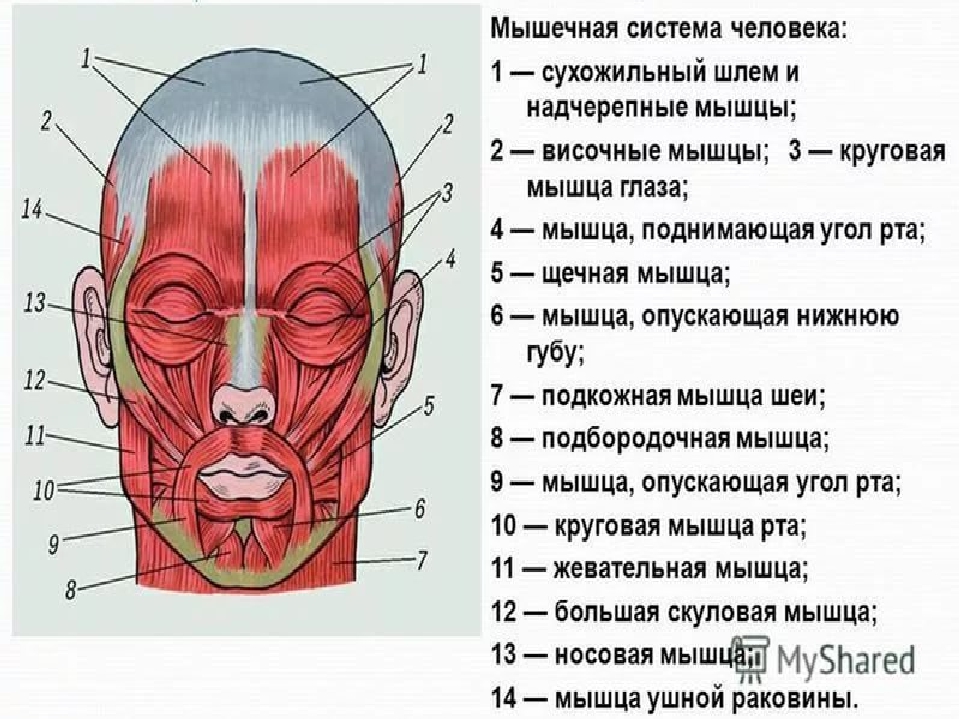 В противном случае, поскольку на результаты измерений, проведенных с использованием динамометра, оказывают влияние такие факторы, как возраст, пол испытуемого, а также рост и вес, следует использовать более объективные показатели. Самым объективным показателем силы будет являться так называемая, относительная величина мышечной силы. Это связано, помимо перечисленных факторов, с тем, что в ходе тренировок, рост абсолютных показателей силы тесно связан с ростом мышечной массы человека, и как следствие с его весом.
В противном случае, поскольку на результаты измерений, проведенных с использованием динамометра, оказывают влияние такие факторы, как возраст, пол испытуемого, а также рост и вес, следует использовать более объективные показатели. Самым объективным показателем силы будет являться так называемая, относительная величина мышечной силы. Это связано, помимо перечисленных факторов, с тем, что в ходе тренировок, рост абсолютных показателей силы тесно связан с ростом мышечной массы человека, и как следствие с его весом.
Чтобы определить величину относительной силы кисти, нужно абсолютные показания в килограммах, полученные измерением ручным динамометром, умножить на 100 и разделить на вес тела спортсмена. Для мужчин, не занимающихся спортом, этот показатель должен составлять 60-70, а для женщин 45-50.
Становая динамометрия, проводимая с использованием станового динамометра, это, можно сказать, комплексное измерение силовых качеств спортсмена, поскольку в таком исследовании участвуют практически все основные мышцы. Упражнение становой тяги с использованием динамометра должно применяться во всех учреждения диспансерного типа спортивно-оздоровительного профиля. В качестве примера станового динамометра можно привести ДС-200 и ДС-500.
Упражнение становой тяги с использованием динамометра должно применяться во всех учреждения диспансерного типа спортивно-оздоровительного профиля. В качестве примера станового динамометра можно привести ДС-200 и ДС-500.
Динамометр Становой ДС-200
Становая динамометрия подразумевает использование станового динамометра – прибора, который по виду напоминает обычный ножной эспандер, который состоит из рукояти, подножки, подкладываемой под ноги, троса и измерительного прибора с датчиком и отсчитывающим устройством. Испытуемый должен потянуть рукоять на себя и вверх так сильно, как только сможет, при этом, ноги должны быть прямыми в коленях.
Относительная величина становой силы рассчитывается точно так же, как и в ручной динамометрии, однако, здесь показатели индекса должны быть в разы больше. Например:
Если индекс менее 170 – то индекс относительной величины становой силы низкий.
- От 170 до 200 – ниже среднего.
- 200 — 230 – средний.
- 230 — 260 – выше среднего.
- Если же более 260 – то считается высоким.
Увеличение относительных показателей силы, как ручной, так и становой, как правило, говорит о повышении мышечной силы, а, следовательно, об увеличении мышечной массы в процентном соотношении.
Показания таких измерений используются в неврологии при обследовании заболеваний, которые могут сопровождаться мышечной слабостью, например, миастения, рассеянный склероз со слабостью конечностей, а также, различные последствия инсульта.
Отдельно следует выделить такой вид исследования, как динамография, при котором показатели силы и скорости сокращения мышц записываются на графике. Как видно из названия, суть этого метода состоит в том, что показания записываются в графическом виде в динамике (во времени). Часто, динамография связана с какими либо упражнениями или обстоятельствами, эффективность которых необходимо измерить.
У детей, также существуют усредненные показатели динамометрии, которые принято считать нормой. Усредненные величины различаются в зависимости от пола, роста, возрастной категории испытуемого. Измерения силы кисти правой руки и становой силы, обычно, проводят для детей в возрасте от восьми до 18 лет в два этапа, с небольшим перерывом для отдыха. Так, нормы показателей силы кисти правой руки для мальчиков составляют:
Усредненные величины различаются в зависимости от пола, роста, возрастной категории испытуемого. Измерения силы кисти правой руки и становой силы, обычно, проводят для детей в возрасте от восьми до 18 лет в два этапа, с небольшим перерывом для отдыха. Так, нормы показателей силы кисти правой руки для мальчиков составляют:
- От 13 до 18,5 кг – для возраста 8-11 лет.
- 21,6 — 37,6 кг – 12-15 лет.
- 45,9 — 51 кг – 16-19 лет.
Для девочек, норма колеблется в пределах:
- 9,8 — 17,1 кг – для возраста 8-11 лет.
- 19,9 — 28,3 кг – 12-15 лет.
- 31,3 — 33,8 кг – 16-19 лет.
Завершая статью, скажем только что динамометрия – это важный элемент антропометрии, который нашел свое применение в физиологии, спортивной медицине, гигиене спорта. Благодаря показателям абсолютной и относительной величины силы производится оценка степени физического развития человека.
что означают цифры на современных домашних весах?
Процент жировой ткани
Он состоит из суммы необходимого и запасенного жира. Средние значения таковы:
Средние значения таковы:
20 – 39 лет – 22–33% у женщин, 8–20% у мужчин;
40 – 59 лет: 24-34% и 11-22%;
60 – 79 лет: 25-36% и 13-25%.
Чем меньше (в рамках нормы), тем здоровее организм. У спортсменов вариант нормы ниже: примерно 14 – 20% у женщин и 6 – 14 у мужчин. Люди с избытком жировой ткани чаще страдают сердечно-сосудистыми заболеваниями, гипертонией и диабетом.
Костная масса
Это общий вес костей в организме, его среднее значение — 2 – 4% от общей массы человека. Если кости слишком легкие, чаще случаются переломы и вывихи. Нарастить костную массу можно с помощью физических упражнений и правильного питания.
Объем воды
Средние значения зависят от возраста, пола и насыщаемости организма водой. В среднем это 45 – 60% у женщин и 50 – 65 у мужчин. И недостаток, и избыток жидкости вредны для здоровья (но стоит учитывать и погрешности измерения).
Мышечная масса
Это полная масса скелетных мышц (есть еще гладкие и сердечные, которые никак не накачать, да и незачем). Средние значения для женщин – 30 – 40% для мужчин – 37 – 55%. Чем больше мышц, тем быстрее происходит метаболизм и лучше иммунитет.
И как это всё измеряется?
Бытовые весы используют технологию биоимпеданса, подавая слабые электрические разряды, когда человек на них стоит. Это не больно и совершенно безопасно! Сигнал встречает сопротивление жировой ткани и проходит без проблем через воду, содержащуюся в организме. На этих данных и строятся показания.
Конечно, полностью полагаться на домашние весы не стоит, поскольку погрешность измерения достигает 5%. Но кое-что новое о себе можно узнать, и это уже хорошо!
ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ. ПРАВИЛА ПОВЕДЕНИЯ НАСЕЛЕНИЯ
ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ. ПРАВИЛА ПОВЕДЕНИЯ НАСЕЛЕНИЯ
По данным Всемирной организации здравоохранения, ежегодно на земном шаре переносят инфекционные заболевания свыше 1 млрд. человек. В течение короткого срока могут заразиться большие массы людей. Так, холера Эль-Тор, начавшись в 1960 г. в Индонезии, к 1971 г. охватила все страны мира. Четвертая пандемия (эпидемия, охватывающая группу стран, континентов) гриппа за два года (1968-1970) поразила около 2 млрд. человек всех континентов и унесла около 1,5 млн. жизней. Нет-нет да и появляются больные чумой, холерой, бруцеллезом. Все еще высок уровень заболеваемости острой дизентерией, брюшным тифом, дифтерией, вирусным гепатитом, сальмонеллезом, гриппом. Особенно опасно их возникновение на предприятиях, в учебных заведениях, воинских коллективах, где один может заразить всех.
человек. В течение короткого срока могут заразиться большие массы людей. Так, холера Эль-Тор, начавшись в 1960 г. в Индонезии, к 1971 г. охватила все страны мира. Четвертая пандемия (эпидемия, охватывающая группу стран, континентов) гриппа за два года (1968-1970) поразила около 2 млрд. человек всех континентов и унесла около 1,5 млн. жизней. Нет-нет да и появляются больные чумой, холерой, бруцеллезом. Все еще высок уровень заболеваемости острой дизентерией, брюшным тифом, дифтерией, вирусным гепатитом, сальмонеллезом, гриппом. Особенно опасно их возникновение на предприятиях, в учебных заведениях, воинских коллективах, где один может заразить всех.
Вот почему очень важно знать признаки инфекционных заболеваний, пути их распространения, способы предупреждения и правила поведения.
ВОЗНИКНОВЕНИЕ ИНФЕКЦИОННЫХ ЗАБОЛЕВАНИЙ
Ноябрь 1990 г. Таежный город нефтедобытчиков Лангепас (Ханты-Мансийский автономный округ) превратился в огромный лазарет. В больницу с кишечной инфекцией обратилось свыше 2000 человек, более 100 были госпитализированы, из которых 13 находились в очень тяжелом состоянии. В чем же причина? В том, что водопроводные и канализационные трубы были проложены рядом, в одной траншее. В результате в водопроводную сеть стали проникать фекальные воды.
В больницу с кишечной инфекцией обратилось свыше 2000 человек, более 100 были госпитализированы, из которых 13 находились в очень тяжелом состоянии. В чем же причина? В том, что водопроводные и канализационные трубы были проложены рядом, в одной траншее. В результате в водопроводную сеть стали проникать фекальные воды.
Другой пример. В кемпинге «Родник», расположенном на окраине Ставрополя, 45 его поселенцев заболели холерой. Сложилась критическая ситуация, ведь в кемпинге за короткое время побывало 733 человека. Их надо было найти, изолировать и вылечить. Носителей холерного вибриона обнаруживали в Барнауле, Перми, Краснодаре и многих других городах. Только чрезвычайные меры предотвратили распространение инфекции. Виной всему оказался родник близ кемпинга. Оползневые явления повредили канализационную сеть, и нечистоты попали в ключевую воду.
Надо помнить, что возбудители инфекционных заболеваний, проникая в организм, находят там благоприятную среду для развития. Быстро размножаясь, они выделяют ядовитые продукты (токсины), которые разрушают ткани, что приводит к нарушению нормальных процессов жизнедеятельности организма. Болезнь возникает, как правило, через несколько часов или дней с момента заражения. В этот период, называемый инкубационным, идет размножение микробов и накопление токсических веществ без видимых признаков заболевания.
Болезнь возникает, как правило, через несколько часов или дней с момента заражения. В этот период, называемый инкубационным, идет размножение микробов и накопление токсических веществ без видимых признаков заболевания.
Носитель их заражает окружающих или обсеменяет возбудителями различные объекты внешней среды.
Различают несколько путей распространения: контактный, когда происходит прямое соприкосновение больного со здоровым человеком; контактно-бытовой
— передача инфекции через предметы домашнего обихода (белье, полотенце, посуда, игрушки), загрязненные выделениями больного; воздушно-капельный
— при разговоре, чихании; водный. Многие возбудители сохраняют жизнеспособность в воде, по крайней мере, несколько дней. В связи с этим передача острой дизентерии, холеры, брюшного тифа может происходить через нее весьма широко. Если не принимать необходимых санитарных мер, водные эпидемии могут привести к печальным последствиям.
А сколько инфекционных заболеваний передается с пищевыми продуктами? В Тульской области было выявлено пять случаев заболевания бруцеллезом.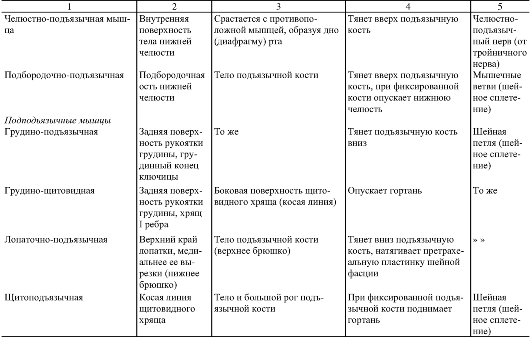 Причина? Пренебрежение ветеринарными требованиями и нормами: 65 голов крупного рогатого скота, больного бруцеллезом, совхоз направил на мясокомбинат, от продукции которого заразились люди,
Причина? Пренебрежение ветеринарными требованиями и нормами: 65 голов крупного рогатого скота, больного бруцеллезом, совхоз направил на мясокомбинат, от продукции которого заразились люди,
На сегодня ведущее значение приобрел сальмонеллез. Заболеваемость им увеличилась в 25 раз. Это одно из распространенных кишечно-желудочных заболеваний. Разносчиками могут являться различные животные: рогатый скот, свиньи, лошади, крысы, мыши и домашняя птица, особенно утки и гуси. Возможно такое заражение от больного человека или носителя сальмонелл.
Большую опасность для окружающих представляют больные, которые своевременно не обращаются к врачу, так как многие инфекционные болезни протекают легко. Но при этом происходит интенсивное выделение возбудителей во внешнюю среду.
Сроки выживания возбудителей различны. Так, на гладких поверхностях целлулоидных игрушек дифтерийная палочка сохраняется меньше, чем на мягких игрушках из шерсти или другой ткани. В готовых блюдах, в мясе, молоке возбудители могут жить долго.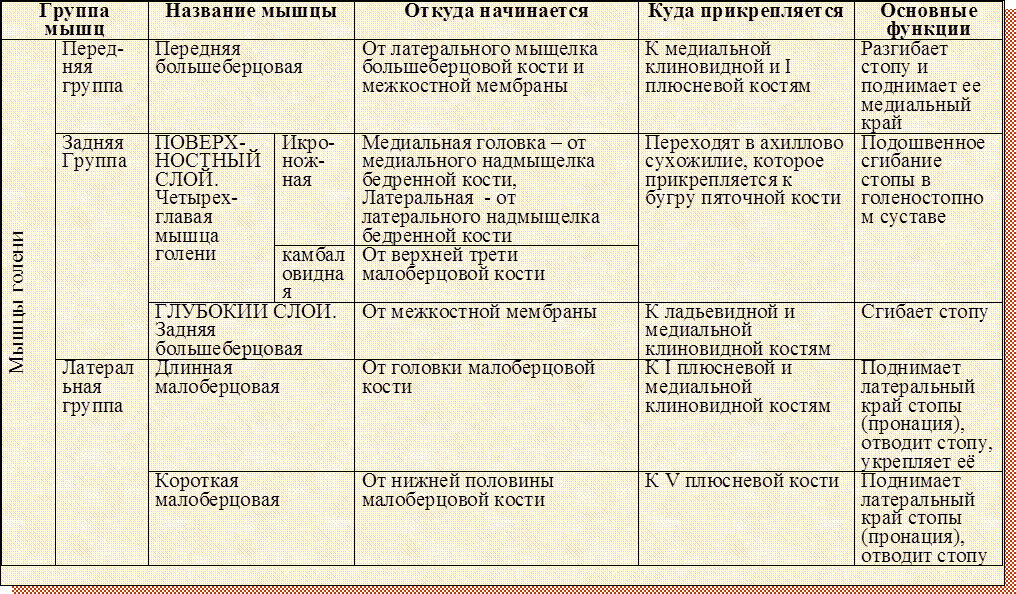 В частности, молоко является благоприятной питательной средой для брюшно-тифозной и дизентерийной палочек.
В частности, молоко является благоприятной питательной средой для брюшно-тифозной и дизентерийной палочек.
В организме человека на пути проникновения болезнетворных микробов стоят защитные барьеры — кожа, слизистая оболочка желудка, некоторые составные части крови. Сухая, здоровая и чистая кожа выделяет вещества, которые приводят к гибели микробов. Слизь и слюна содержат высокоактивный фермент—лизоцим, разрушающий многих возбудителей. Оболочка дыхательных путей также хороший защитник. Надежный барьер на пути микробов — желудок. Он выделяет соляную кислоту и ферменты, которые нейтрализуют большинство возбудителей заразных болезней. Однако если человек пьет много воды, то кислотность, разбавляясь, снижается. Микробы в таких случаях не гибнут и с пищей проникают в кишечник, а оттуда в кровь.
Необходимо отметить, что защитные силы более эффективны в здоровом, закаленном организме. Переохлаждение, несоблюдение личной гигиены, травма, курение, радиация, прием алкоголя резко снижают его сопротивляемость.
РАСПОЗНАВАНИЕ ИНФЕКЦИОННЫХ ЗАБОЛЕВАНИЙ
Наиболее типичными признаками инфекционного заболевания являются: озноб, жар, повышение температуры. При этом возникает головная боль, боль в мышцах и суставах, недомогание, общая слабость, разбитость, иногда тошнота, рвота, нарушается сон, ухудшается аппетит. При тифе, менингококковой инфекции появляется сыпь. При гриппе и других респираторных заболеваниях — чихание, кашель, першение в горле. Ангина и дифтерия вызывают боли в горле при глотании. При дизентерии — понос. Рвота и понос — признаки холеры и сальмонеллеза.
Рассмотрим кратко наиболее часто встречающиеся инфекции, пути их распространения и способы предупреждения.
Инфекции дыхательных путей — наиболее многочисленные и самые распространенные заболевания. Ежегодно ими перебаливает до 15-20% всего населения, а в период эпидемических вспышек гриппа — до 40%. Возбудители локализуются в верхних дыхательных путях и распространяются воздушно-капельным способом. Микробы попадают в воздух со слюной и слизью при разговоре, чихании, кашле больного (наибольшая концентрация на расстоянии 2-3 м от больного). Крупные капли, содержащие возбудителей, довольно быстро оседают, подсыхают, образуя микроскопические ядрышки. С пылью они вновь поднимаются в воздух и переносятся в другие помещения. При их вдыхании и происходит заражение. При высокой влажности воздуха в помещениях, недостаточном их проветривании и других нарушениях санитарно-гигиенических правил возбудители сохраняются во внешней среде дольше.
Микробы попадают в воздух со слюной и слизью при разговоре, чихании, кашле больного (наибольшая концентрация на расстоянии 2-3 м от больного). Крупные капли, содержащие возбудителей, довольно быстро оседают, подсыхают, образуя микроскопические ядрышки. С пылью они вновь поднимаются в воздух и переносятся в другие помещения. При их вдыхании и происходит заражение. При высокой влажности воздуха в помещениях, недостаточном их проветривании и других нарушениях санитарно-гигиенических правил возбудители сохраняются во внешней среде дольше.
При стихийном бедствии и крупных катастрофах обычно происходит скапливание людей, нарушаются нормы и правила общежития, что обусловливает массовость заболевания гриппом, дифтерией, ангиной, менингитом.
Грипп. Его вирус в течение короткого времени может поразить значительное количество людей. Он устойчив к замораживанию, но быстро погибает при нагревании, высушивании, под действием дезинфицирующих средств, при ультрафиолетовом облучении. Инкубационный период продолжается от 12 ч до 7 суток. Характерные признаки болезни — озноб, повышение температуры, слабость, сильная головная боль, кашель, першение в горле, насморк, саднение за грудиной, осипший голос. При тяжелом течении возможны осложнения — пневмония, воспаление головного мозга и его оболочек.
Инкубационный период продолжается от 12 ч до 7 суток. Характерные признаки болезни — озноб, повышение температуры, слабость, сильная головная боль, кашель, першение в горле, насморк, саднение за грудиной, осипший голос. При тяжелом течении возможны осложнения — пневмония, воспаление головного мозга и его оболочек.
Дифтерия характеризуется воспалительным процессом в глотке и токсическим поражением сердечно-сосудистой и нервной систем. Возбудитель болезни— дифтерийная палочка. Входными воротами инфекции чаще всего являются слизистые оболочки зева, гортани и носа. Передается воздушно-капельным путем.
Инкубационный период от 5 до 10 дней. Наиболее характерное проявление болезни—образование пленок в верхних дыхательных путях. Опасность для жизни представляет токсическое поражение ядами дифтерийных палочек организма больного. При их распространении может возникнуть нарушение дыхания.
Холера, дизентерия, брюшной тиф, сальмонеллез, инфекционный гепатит — все эти острые кишечные инфекции занимают второе место после воздушно-капельных. При этой группе заболеваний болезнетворные микроорганизмы проникают внутрь с проглатываемой пищей или водой.
При этой группе заболеваний болезнетворные микроорганизмы проникают внутрь с проглатываемой пищей или водой.
Разрушение водопроводных и канализационных сетей, низкая санитарная культура, беспечность и неосмотрительность в использовании открытых водоемов приводят к возникновению этих эпидемий.
Острая бактериальная дизентерия. Возбудители—дизентерийные бактерии, которые выделяются с испражнениями больного. Во внешней среде они сохраняются 30-45 дней. Инкубационный период — до 7 дней (чаще 2-3 дня). Заболевание сопровождается повышением температуры, ознобом, жаром, общей слабостью, головной болью. Начинается со схваткообразных болей в животе, с частого жидкого стула, в тяжелых случаях — с примесью слизи и крови. Иногда бывает рвота.
Брюшной тиф. Источник инфекции — больные или бактерионосители. Палочка брюшного тифа и паратифов выделяется с испражнениями и мочой. В почве и воде они могут сохраняться до четырех месяцев, в испражнениях до 25 дней, на влажном белье — до двух недель. Инкубационный период продолжается от одной до трех недель. Заболевание развивается постепенно: ухудшается самочувствие, нарушается сон, повышается температура. На 7-8-й день появляется сыпь на коже живота, грудной клетке. Заболевание длится 2-3 недели и может осложниться кишечным кровотечением или прободением кишечника на месте одной из многочисленных образовавшихся при этом язв.
Инкубационный период продолжается от одной до трех недель. Заболевание развивается постепенно: ухудшается самочувствие, нарушается сон, повышается температура. На 7-8-й день появляется сыпь на коже живота, грудной клетке. Заболевание длится 2-3 недели и может осложниться кишечным кровотечением или прободением кишечника на месте одной из многочисленных образовавшихся при этом язв.
ОСНОВЫ ЗАЩИТЫ И ПРАВИЛА ПОВЕДЕНИЯ НАСЕЛЕНИЯ
Инфекционные заболевания возникают при трех основных факторах: наличии источника инфекции, благоприятных условиях для распространения возбудителей и восприимчивого к заболеванию человека. Если исключить из этой цепи хотя бы одно звено, эпидемический процесс прекращается. Следовательно, целью предупреждающих мероприятий является воздействие на источник инфекции, чтобы уменьшить обсеменение внешней среды, локализовать распространение микробов, а также повысить устойчивость населения к заболеваниям.
Поскольку главным источником инфекции является больной человек или бактерионоситель, необходимо раннее выявление, немедленная их изоляция и госпитализация. При легком течении заболевания люди, как правило, поздно обращаются к врачу или совсем этого не делают. Помочь в скорейшем выявлении таких больных могут подворные обходы.
При легком течении заболевания люди, как правило, поздно обращаются к врачу или совсем этого не делают. Помочь в скорейшем выявлении таких больных могут подворные обходы.
Помещения, где находится больной, надо регулярно проветривать. Для него выделить отдельное помещение или отгородить ширмой. Обслуживающему персоналу обязательно носить защитные марлевые маски.
Важное значение для предупреждения развития инфекционных заболеваний имеет экстренная и специфическая профилактика.
Экстренная профилактика проводится при возникновении опасности массовых заболеваний, но когда вид возбудителя еще точно не определен. Она заключается в приеме населением антибиотиков, сульфаниламидных и других лекарственных препаратов. Средства экстренной профилактики при своевременном их использовании по предусмотренным заранее схемам позволяют в значительной степени предупредить инфекционные заболевания, а в случае их возникновения — облегчить их течение.
Специфическая профилактика — создание искусственного иммунитета (невосприимчивости) путем предохранительных прививок (вакцинации)— проводится против некоторых болезней (натуральная оспа, дифтерия, туберкулез, полиомиелит и др. ) постоянно, а против других — только при появлении опасности их возникновения и распространения.
) постоянно, а против других — только при появлении опасности их возникновения и распространения.
Повысить устойчивость населения к возбудителям инфекции возможно путем массовой иммунизации предохранительными вакцинами, введением специальных сывороток или гамма-глобулинов. Вакцины представляют собой убитых или специальными методами ослабленных болезнетворных микробов, при введении которых в организм здоровых людей у них вырабатывается состояние невосприимчивости к заболеванию. Вводятся они разными способами: подкожно, накожно, внутрикожно, внутримышечно, через рот (в пищеварительный тракт), путем вдыхания.
Для предупреждения и ослабления инфекционных заболеваний в порядке самопомощи и взаимопомощи рекомендуется использовать средства, содержащиеся в аптечке индивидуальной АИ-2.
При возникновении очага инфекционного заболевания в целях предотвращения распространения болезней объявляется карантин или обсервация.
Карантин вводится при возникновении особо опасных болезней (оспы, чумы, холеры и др.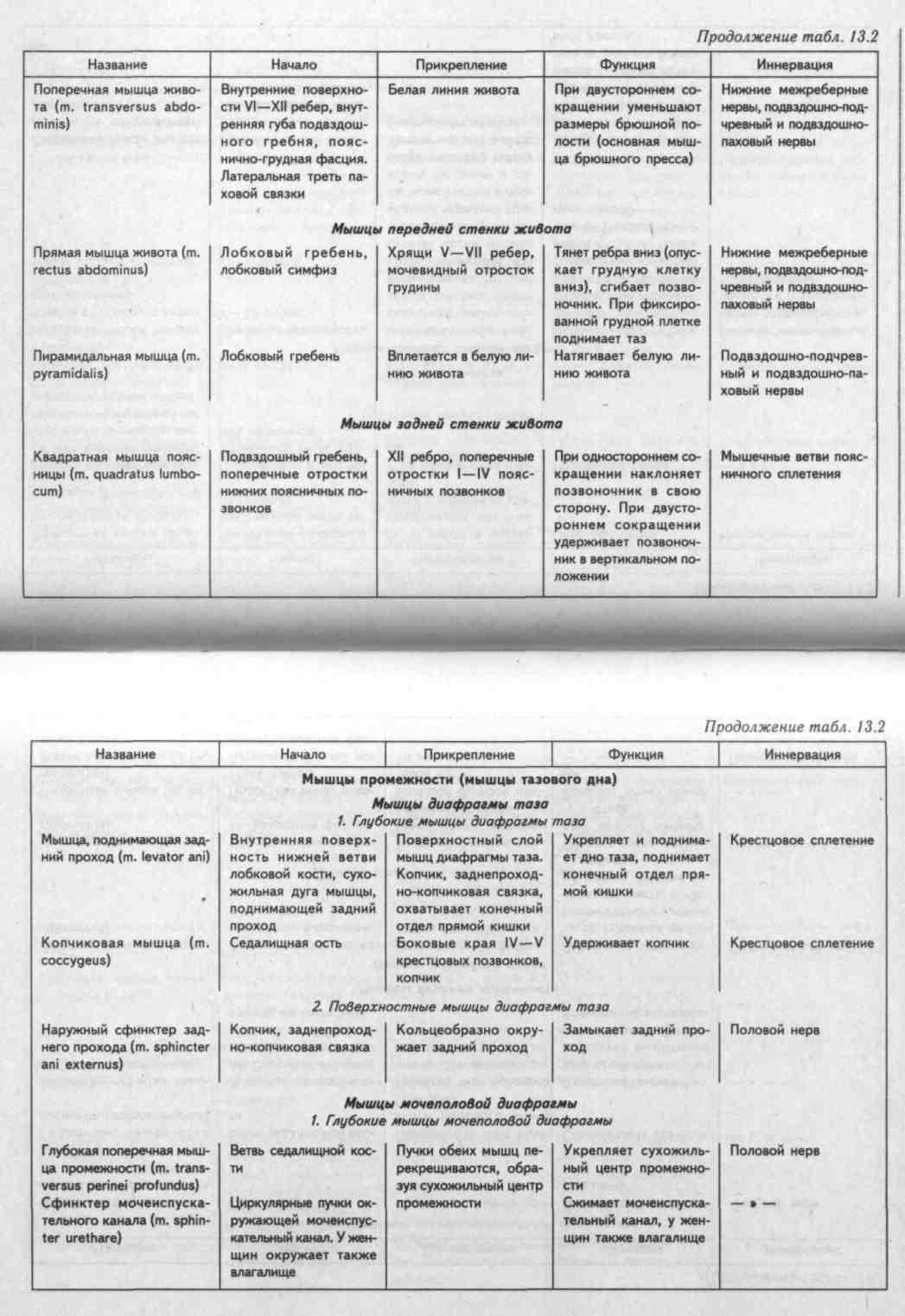 ). Он может охватывать территорию района, города, группы населенных пунктов.
). Он может охватывать территорию района, города, группы населенных пунктов.
Карантин представляет собой систему режимных, противоэпидемических и лечебно-профилактических мероприятий, направленных на полную изоляцию очага и ликвидацию болезней в нем.
Основными режимными мероприятиями при установлении карантина являются: охрана очага инфекционного заболевания, населенных пунктов в нем, инфекционных изоляторов и больниц, контрольно-передаточных пунктов. Запрещение входа и выхода людей, ввода и вывода животных, а также вывоза имущества. Запрещение транзитного проезда транспорта, за исключением железнодорожного и водного. Разобщение населения на мелкие группы и ограничение общения между ними. Организация доставки по квартирам (домам) населению продуктов питания, воды и предметов первой необходимости. Прекращение работы всех учебных заведений, зрелищных учреждений, рынков. Прекращение производственной деятельности предприятий или перевод их на особый режим работы.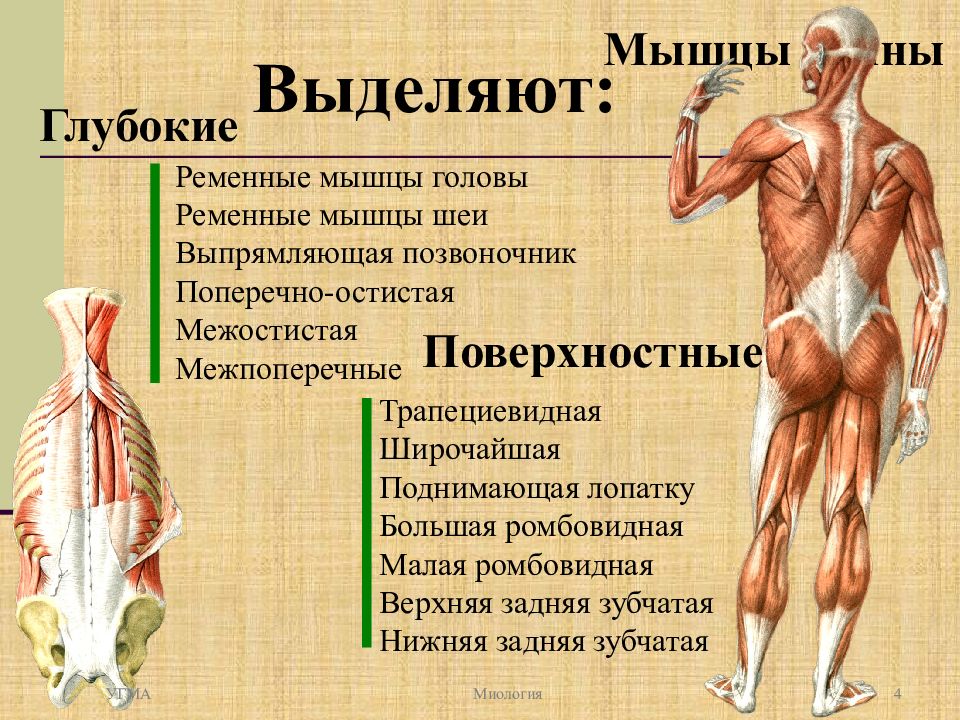
Противоэпидемические и лечебно-профилактические мероприятия в условиях карантина включают: использование населением медицинских препаратов, защиту продовольствия и воды, дезинфекцию, дезинсекцию, дератизацию, санитарную обработку, ужесточенное соблюдение правил личной гигиены, активное выявление и госпитализацию инфекционных больных.
Обсервация вводится в том случае, если вид возбудителя не является особо опасным. Цель обсервации — предупредить распространение инфекционных заболеваний и ликвидировать их. Для этого проводятся по существу те же лечебно-профилактические мероприятия, что и при карантине, но при обсервации менее строги изоляционно-ограничительные меры.
Срок карантина и обсервации определяется длительностью максимального инкубационного периода заболевания, исчисляемого с момента изоляции последнего больного и окончания дезинфекции в очаге.
Люди, находящиеся на территории очага инфекционного заболевания, должны для защиты органов дыхания пользоваться ватно-марлевыми повязками. Для кратковременной защиты рекомендуется использовать свернутый в несколько слоев платок или косынку, полотенце или шарф. Не помешают и защитные очки. Целесообразно пользоваться накидками и плащами из синтетических и прорезиненных тканей, пальто, ватниками, резиновой обувью, обувью из кожи или ее заменителей, кожаными или резиновыми перчатками (рукавицами).
Для кратковременной защиты рекомендуется использовать свернутый в несколько слоев платок или косынку, полотенце или шарф. Не помешают и защитные очки. Целесообразно пользоваться накидками и плащами из синтетических и прорезиненных тканей, пальто, ватниками, резиновой обувью, обувью из кожи или ее заменителей, кожаными или резиновыми перчатками (рукавицами).
Защита продовольствия и воды заключается главным образом в создании условий, исключающих возможность их контакта с зараженной атмосферой. Надежными средствами защиты могут быть все виды плотно закрывающейся тары.
Водой из водопровода и артезианских скважин разрешается пользоваться свободно, но кипятить ее обязательно.
В очаге инфекционною заболевания не обойтись без дезинфекции, дезинсекции и дератизации.
Дезинфекция проводится с целью уничтожения или удаления микробов и иных возбудителей с объектов внешней среды, с которыми может соприкасаться человек. Для дезинфекции применяют растворы хлорной извести и хлорамина, лизол, формалин и др. При отсутствии этих веществ используется горячая вода с мылом или содой.
При отсутствии этих веществ используется горячая вода с мылом или содой.
Дезинсекция проводится для уничтожения насекомых и клещей — переносчиков возбудителей инфекционных заболеваний. С этой целью используются различные способы: механический (выколачивание, встряхивание, стирка), физический (проглаживание утюгом, кипячение), химический (применение инсектицидов — хлорофоса, тиофоса, ДДТ и др.), комбинированный. Для защиты от укуса насекомых применяют отпугивающие средства (репелленты), которыми смазываются кожные покровы открытых частей тела.
Дератизация проводится для истребления грызунов — переносчиков возбудителей инфекционных заболеваний. Она проводится чаще всего с помощью механических приспособлений и химических препаратов.
Большую роль в предупреждении инфекционных заболеваний играет строгое соблюдение правил личной гигиены: мытье рук с мылом после работы и перед едой; регулярное обмывание тела в бане, ванне, под душем со сменой нательного и постельного белья; систематическая чистка и встряхивание верхней одежды и постельных принадлежностей; поддержание в чистоте жилых и рабочих помещений; очистка от грязи и пыли, обтирание обуви перед входом в помещение; употребление только проверенных продуктов, кипяченой воды и молока, промытых кипяченой водой фруктов и овощей, тщательно проваренных мяса и рыбы.
Успех ликвидации инфекционного очага во многом определяется активными действиями и разумным поведением всего населения. Каждый должен строго выполнять установленные режим и правила поведения на работе, на улице и дома, постоянно выполнять противоэпидемические и санитарно-гигиенические нормы.
как избежать дефицита жизненно важных микроэлементов — Российская газета
Наше тело нуждается практически во всех химических элементах. Но некоторые мы можем получать только с пищей. И потому неправильное питание приводит порой к серьезным заболеваниям. Ведь модные сегодня диеты, а то и просто привычка питаться на скорую руку консервированными продуктами — все это неизбежно ведет к дефициту многих жизненно важных минералов и веществ. Какие же микроэлементы особенно нужны нам, для чего и как определить их нехватку?
Вот лишь пять элементов из таблицы Менделеева. Нам их требуется совсем немного, но в обязательном порядке:
1 Железо (Fe) — входит в состав гемоглобина крови (60-75% железа в нашем организме содержится в эритроцитах — красных кровяных клетках). Гемоглобин переносит кислород, которым мы дышим, ко всем органам и тканям, поэтому жизнь без него невозможна так же, как и без кислорода. При дефиците железа в организме ухудшается клеточное дыхание, что ведет к дистрофии тканей и органов, потом развивается анемия, а в запущенных случаях — рак крови.
Человек должен получать 15-20 мг железа в день. Оно содержится в твороге и твердом сыре, бобовых, злаках, свекле, печени животных. Лучшему всасыванию железа из пищевых продуктов способствуют лимонная и аскорбиновая кислоты и фруктоза, которые содержатся во фруктах, ягодах, их соках.
Для примера: гречневая крупа, фасоль, горох, шоколад, черника содержат около 4 мг железа на 100 граммов продукта.
2 Медь(Сu) — химический элемент, продлевающий нам молодость, так как он отвечает за эластичность тканей. Преждевременная седина, морщины, обвисание кожи — верные признаки нехватки меди. При нехватке этого элемента в организме наблюдаются: задержка роста (у детей), анемия, дерматозы, депигментация волос, частичное облысение, потеря аппетита, сильное исхудание, понижение уровня гемоглобина, атрофия сердечной мышцы, варикозное расширение вен, причиной которой является все то же нарушение эластичности тканей.
Меди организму человека требуется 1-3 мг в сутки. Ею богаты печень, почки и мясо животных, морская и пресноводная рыба; морепродукты, крупы (перловая, пшеничная, гречневая, овсяная), картофель, укроп, некоторые фрукты и ягоды — черная смородина, малина, клюква, абрикосы, крыжовник, груши, клубника. Кстати, пиво способствует усвоению меди из пищи.
Для примера: 100 граммов печени трески содержит 12 мг меди, какао-порошок — 5 мг, печень говяжья — 4 мг.
3 Цинк (Zn) — элемент, особенно важный для мужчин, так как больше всего его содержится в сперме. Кроме того, он оказывает влияние на активность половых и гонадотропных гормонов гипофиза, участвует в жировом, белковом и витаминном обмене, в процессах кроветворения. Верный признак дефицита цинка — потеря обоняния и вкуса. Дети при недостатке цинка страдают гнойничковыми заболеваниями кожи и слизистых оболочек. У взрослых это проявляется в перевозбуждении нервной системы, быстром утомлении, ослаблением и выпадением волос, утолщением кожи, отеками слизистых оболочек рта и пищевода. Недостаточность цинка также приводит к бесплодию.
Цинк способствует заживлению ран, поэтому его необходимо принимать перед любой хирургической операцией, а также после нее. Кроме того, цинк помогает при лечении катаракты, замедляет разрушение сетчатки. Подобно витамину С, цинк полностью прекращает вирусную инфекцию, если захватить ее достаточно рано.
В среднем нам надо употреблять 10-20 мг цинка ежедневно. А беременным женщинам до 30 мг. Наиболее богаты этим микроэлементом отруби, проросшие зерна пшеницы, хлеб грубого помола.
Для примера: в 100 граммах дрожжей 10 мг цинка, в отварной говядине — 7 мг, в тыквенных семечках 7,4 мг, в какао-порошке — 6,3.
4 Кальций (Сa) — этот элемент не только отвечает за крепость нашего скелета, но и участвует во всех жизненных процессах организма. Нормальная свертываемость крови происходит только в присутствии солей кальция. Кальций играет важную роль в нервно-мышечной возбудимости тканей. При недостатке кальция наблюдаются: тахикардия, аритмия, боли в мышцах, беспричинные рвоты, запоры. Волосы делаются грубыми и выпадают. Ногти становятся ломкими. Кожа утолщается и грубеет. Глазной хрусталик теряет прозрачность. Любое падение может привести к серьезной травме, ведь кости делаются хрупкими.
В среднем человеку надо около 1000 мг кальция в сутки. Но эта величина разнится для людей разного возраста. Много кальция содержится в кунжуте, крапиве, твердом сыре, халве, зелени петрушки, сардинах, капусте.
Для примера: в ста граммах голландского сыра содержится около 1000 мг кальция, а в 100 граммах халвы — 824 мг.
5 Калий (K) — этот элемент отвечает за клеточные оболочки, делая их проницаемыми для прохождения солей. Поэтому он необходим для ясности ума, избавления от шлаков, лечения аллергии. Недостаток калия приводит к замедлению роста организма и нарушению половых функций, вызывает мышечные судороги, перебои в работе сердца.
Ежедневно нам надо до 2000 мг калия. Этот элемент можно пополнить, вписав в меню мясо и субпродукты, черную смородину, овсяную крупу, чернослив, арбуз, кукурузу.
Для примера: один банан среднего размера содержит 450 мг калия, чашка молока — 370 мг, один апельсин — 250 мг.
Кстати
Кроме микроэлементов нам нужны еще и ультрамикроэлементы, которые содержатся в организме человека и вовсе в очень малых количествах. К ним относятся хром, ванадий, селен, бор, никель, олово, серебро, золото и др. Вот как сказывается их недостаток:
Литий. Предполагают, что его недостаток вызывает состояние агрессии, депрессии и, как вторичное явление, пьянство.
Хром и ванадий — их дефицит приводит к заболеванию диабетом, к потере зрения.
Олово — без него человек страдает ранним облысением. А при длительной нехватке еще и развивается глухота.
Бор — его дефицит приводит к остеопорозу, так как этот элемент помогает сохранить в костях употребляемый кальций.
Селен — его нехватка бьет по сердцу. Он отвечает за нормальную работу сердечной мышцы, а также за противоопухолевую активность организма.
Кобальт — спасает от малокровия, так как без него не образуется витамин В12 (излечивающий анемию).
мышечной системы человека | Функции, схемы и факты
Следующие разделы предоставляют базовую основу для понимания общей мышечной анатомии человека с описанием крупных мышечных групп и их действий. Различные группы мышц работают согласованно, чтобы контролировать движения человеческого тела.
Шея
Движение шеи описывается в терминах вращения, сгибания, разгибания и бокового сгибания (т. Е. Движения, используемого для прикосновения уха к плечу).Направление действия может быть ипсилатеральным, что относится к движению в направлении сокращающейся мышцы, или контралатеральным, что относится к движению от стороны сокращающейся мышцы.
Получите подписку Britannica Premium и получите доступ к эксклюзивному контенту. Подпишитесь сейчасВращение — одно из важнейших действий шейного отдела позвоночника. Вращение в основном осуществляется грудинно-ключично-сосцевидной мышцей, которая сгибает шею в ипсилатеральную сторону и вращает шею в противоположную сторону.Вместе грудинно-ключично-сосцевидные мышцы по обеим сторонам шеи сгибают шею и поднимают грудину, чтобы способствовать принудительному вдоху. Передняя и средняя лестничные мышцы, которые также расположены по бокам шеи, действуют ипсилатерально, поворачивая шею, а также поднимая первое ребро. Сплениус головы и сплениус шеи, расположенные в задней части шеи, вращают голову.
Боковое сгибание также является важным действием шейного отдела позвоночника. В сгибание шейной стороны вовлекаются грудинно-ключично-сосцевидные мышцы.Задние лестничные мышцы, расположенные на нижних сторонах шеи, ипсилатерально сгибают шею в сторону и приподнимают второе ребро. Сплениус головы и шейный позвонок также помогают при сгибании шеи в стороны. Мышцы, выпрямляющие позвоночник (подвздошно-костная, длинная и спинальная) — это большие глубокие мышцы, которые увеличивают длину спины. Все три действуют для ипсилатерального изгиба шеи.
Под сгибанием шеи понимается движение, при котором подбородок касается груди. Это достигается в первую очередь грудинно-ключично-сосцевидными мышцами с помощью длинных коленных и длинных мышц головы, которые находятся в передней части шеи.Разгибание шеи противоположно сгибанию и выполняется многими из тех же мышц, которые используются для других движений шеи, включая шейную шейку шеи, звездочную мышцу головы, подвздошно-костную, длинную и спинную мышцы.
Спина
Послушайте, как врач объяснит причины и методы лечения боли в спине, называемой лордозом.
Узнайте о причинах и способах лечения боли в спине.
Contunico © ZDF Enterprises GmbH, Майнц См. Все видео по этой статьеСпина является источником многих мышц, которые участвуют в движении шеи и плеч.Кроме того, осевой скелет, проходящий через спину вертикально, защищает спинной мозг, который иннервирует почти все мышцы тела.
Множественные мышцы спины функционируют именно при движениях спины. Например, мышцы, выпрямляющие позвоночник, разгибают спину (сгибают ее назад) и сгибают спину в стороны. Мышцы semispinalis dorsi и semispinalis capitis также расширяют спину. Маленькие мышцы позвонков (мультифиди и вращатели) помогают вращать, разгибать и сгибать спину в стороны.Квадратная мышца поясницы в нижней части спины сгибает поясничный отдел позвоночника и помогает вдыхать воздух благодаря своим стабилизирующим воздействиям в месте прикрепления к 12-му ребру (последнему из плавающих ребер). Лопатка (лопатка) поднимается трапециевидной мышцей, которая проходит от задней части шеи до середины спины, большими ромбовидными и малыми ромбовидными мышцами в верхней части спины и мышцей, поднимающей лопатку, которая проходит по бокам и сзади на шее.
Набор данных об архитектуре мышц нижней конечности человека in vivo, полученный с помощью визуализации тензора диффузии
Abstract
«Золотой стандарт» эталонных наборов архитектуры мускулов человека основан на образцах трупов пожилых людей, которые вряд ли будут репрезентативными для значительной части человеческой популяции.Это важно для моделирования опорно-двигательного аппарата, где свойства общих моделей генерировать мышечную силу определяются этими данными, но могут быть неприменимы при применении к моделям молодых, здоровых людей. Получение данных об индивидуальной архитектуре мышц in vivo, затруднительно, однако недавно появилась возможность использовать диффузионную тензорную магнитно-резонансную томографию (DTI) как действенный метод достижения этого. DTI был использован здесь для предоставления набора данных об архитектуре 20 мышц нижних конечностей от 10 здоровых взрослых, включая длину мышечных волокон, которые являются важными входными данными для моделей мышц типа Хилла, обычно используемых при моделировании опорно-двигательного аппарата.Было обнаружено, что максимальная изометрическая сила и длина мышечных волокон не соотносятся с антропометрией объекта, что позволяет предположить, что эти факторы может быть трудно предсказать, используя алгоритмы масштабирования или оптимизации. Эти данные также подчеркивают высокий уровень анатомических различий, которые существуют между людьми с точки зрения архитектуры мышц нижних конечностей, что подтверждает необходимость включения свойств создания силы в модели опорно-двигательного аппарата для оптимизации их точности для клинической оценки.
Образец цитирования: Charles JP, Suntaxi F, Anderst WJ (2019) In vivo Набор данных об архитектуре мышц нижней конечности человека, полученный с использованием тензорной диффузионной визуализации. PLoS ONE 14 (10): e0223531. https://doi.org/10.1371/journal.pone.0223531
Редактор: Джулия Мольнар, Нью-Йоркский технологический институт, США
Поступила: 17 марта 2019 г .; Принята к печати: 23 сентября 2019 г .; Опубликовано: 15 октября 2019 г.
Авторские права: © 2019 Charles et al.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Доступность данных: Все соответствующие данные находятся в документе и его файлах с вспомогательной информацией. Скрипт MATLAB, использованный в этом исследовании, находится в свободном доступе по адресу DOI: 10.6084 / m9.figshare.96.
Финансирование: Эта работа финансировалась отделением ортопедической хирургии Питтсбургского университета.Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Введение
Скелетно-мышечная архитектура ( и . и . Макроскопическое расположение мышечных волокон [1]) нижней конечности человека хорошо определена, и опубликовано несколько обширных наборов данных [2, 3]. Однако эти наборы справочных данных «золотого стандарта» основаны на образцах трупов пожилых людей, которые по разным причинам, таким как возможные изменения в архитектуре мышц в результате старения [4], вряд ли будут репрезентативными для молодых, активных и здоровых взрослых [5 ].Эти различия были подчеркнуты в отношении объемов мышц [5, 6], хотя степень вариации свойств архитектуры мышц, таких как длина мышечных волокон, угол перистости и максимальная изометрическая сила, в значительной степени неизвестна. Это особенно важно в контексте моделирования опорно-двигательного аппарата с использованием безразмерных моделей мышц типа Хилла [7], которые определяются этими свойствами. Важно отметить, что различные анализы чувствительности показали, что эти модели особенно чувствительны даже к небольшим изменениям в мышечных волокнах и, в частности, длине провисания сухожилий [8–14].Более того, насколько хорошо эти параметры масштабируются по отношению к антропометрическим факторам тела, таким как масса тела или конечностей, также подробно не сообщалось, хотя было высказано предположение, что длина волокон может не особенно сильно зависеть от длины кости [15]. Хотя параметры архитектуры мышц можно оценить с помощью оптимизации [16–19], прямое измерение этих in vivo может повысить точность вычислительных моделей.
Используя ранее установленную основу диффузионно-тензорной визуализации (DTI) и волоконной трактографии, в сочетании с другими последовательностями магнитно-резонансной томографии [20], это исследование направлено на развитие предыдущей литературы и предоставление подробного описания архитектуры мышц нижней конечности in vivo человека. набор данных.Это также подчеркнет уровень межсубъектной изменчивости параметров мышечной архитектуры, которая существует у молодых здоровых взрослых, а также масштабные отношения между мышечной архитектурой и пропорциями тела.
Методы
Для настоящего исследования данные были собраны по 20 мышцам правых нижних конечностей 10 здоровых, непрофессионально атлетически подготовленных взрослых (5 мужчин, 5 женщин; возраст 27 ± 4 года. Масса тела 76 ± 12) кг; Таблица 1), которые подписали документы об информированном согласии до участия в этом исследовании, одобренном IRB.Анализируемые мышцы были разделены на 5 отдельных функциональных групп на основе основных функций (таблица 2), которые были основаны на предыдущих исследованиях архитектуры мышц человека [3]. Длина мышечных волокон, угол перистости волокон и объемы мышц оценивались для каждой из этих мышц с использованием проверенных методик магнитно-резонансной томографии (МРТ) и DTI [20]; см. ниже).
Таблица 1. Информация об участниках исследования.
Длина бедра — расстояние между самой проксимальной стороной большого вертела бедренной кости и самой дистальной стороной латерального мыщелка бедренной кости.Длина ноги — расстояние от большеберцового плато до центра голеностопного (тибиоталарного) сустава. L L — Полная длина нижней конечности. V LM — Общий объем мышц нижней конечности (сумма объемов исследуемых мышц). V L — Общий объем нижней конечности (сумма объемов мышц плюс жир, фасция и кожа).
https://doi.org/10.1371/journal.pone.0223531.t001
Получение МРТ и DTI
Все МРТ-изображения были получены от гребня подвздошной кости до голеностопного сустава с использованием сканера 3 Т (Biograph mMR, Siemens, Мюнхен, Германия), при этом каждый субъект находился в положении лежа на спине, а нижние конечности — в анатомическом положении.Визуализация состояла из двух последовательностей (рис. 1A и 1B): T1-взвешенное анатомическое турбо спин-эхо (TSE) (размер вокселя 0,47 × 0,47 × 6,5 мм 3 , время повторения [TR] — 650 мс, время эхо [TE] — 23 мс, количество срезов — 35 на сегмент, количество средних сигналов (NSA) — 1, коэффициент ускорения — 2), и планарное спин-эхо с однократным повторным фокусом и двойным перефокусированием (размер вокселя 2,96 × 2,96 × 6,5 мм). 3 , TR / TE 7900/65 мс, градиенты диффузии в 12 направлениях, значение b — 0 и 400 с / мм 2 , сильное подавление жира — восстановление спектральной ослабленной инверсии [SPAIR], количество срезов — 35 на сегмент, NSA — 2, коэффициент ускорения — 2, полоса пропускания — 2440 Гц / пиксель).Advanced B 0 регулировка шиммирования была выполнена для каждого сегмента, чтобы уменьшить пространственное искажение и минимизировать химический сдвиг остаточного жира в изображениях, взвешенных по диффузии, в направлении фазового кодирования (от переднего к заднему). Для каждого объекта изображения были получены в аксиальной ориентации срезов и повторены в общей сложности от пяти до шести сегментов, которые были объединены во время постобработки с использованием плагина Stitching для Fiji / ImageJ [21, 22]. Общее время получения изображения составляло ~ 37 минут на одного человека.
Рис 1.
Типичное T1-взвешенное МРТ анатомическое изображение (A) и изображение тензора диффузии (B) сегмента бедра одного пациента. Мышцы и кости были сегментированы в цифровом виде из изображений T1 для создания трехмерных представлений нижних конечностей (C) (определения сокращений мышц см. В таблице 2). Мышечные пучки (волокна) отслеживали по диффузионно-взвешенным МР-изображениям (D). С помощью этих трехмерных моделей на основе облака точек можно было измерить длину волокна (L f ) и угол перистости поверхностных волокон (θ, угол волокон относительно линии действия мышцы (синяя линия)).
https://doi.org/10.1371/journal.pone.0223531.g001
МРТ-изображения, взвешенные по T1, были сегментированы в цифровой форме в программе Mimics (Materialise, Левен, Бельгия) для создания трехмерных сеток каждой мышцы (рис. 1C). , что позволило определить объем отдельных мышц (живота) ( 3 мм).
Предварительная обработка DTI и волоконная трактография
Изображения тензора диффузии были предварительно обработаны для уменьшения артефактов изображения и улучшения отношения сигнал / шум.Чтобы уменьшить артефакты изображения, вызванные возможным движением объектов или пространственным искажением (вихревые токи и / или неоднородность магнитного поля), каждое взвешенное по диффузии изображение было зарегистрировано в недиффузионно-взвешенное изображение (со значением b 0) с использованием аффинного преобразования. в DTI-studio [23].
Чтобы уменьшить отношение сигнал / шум изображений, алгоритм подавления шума Rician был применен к изображениям DTI [24] в MedINRIA (www.med.inria.fr), где тензоры диффузии для каждого объекта были оценены и сглажены.Ручная установка пороговых значений удалила фоновые пиксели из оценки тензора. Мышечные пучки для каждой мышцы оценивались на основе этих тензоров с помощью трактографии в программном обеспечении Camino [25], создавая тракты волокон из областей интереса (ROI), построенных на основе анатомических изображений T1 MR (рис. 1D). Эти тракты отслеживались в двух направлениях (размер шага 1 мм) от исследуемых областей засева (ROI) и продолжались до тех пор, пока не прекращались на основании определенных критериев остановки кривизны волокна (изменение угла> 10 градусов на 5 мм).Эти параметры трактографии были одинаковыми для мышц и испытуемых.
Хотя мышечные волокна не обязательно могут проходить по всей длине мышечного пучка (пучки из ~ 5–10 волокон) и вместо этого могут быть соединены последовательно, было показано, что волокна в таком расположении могут активироваться одновременно, чтобы действовать как единое целое. волокно [26]. Поэтому здесь предполагалось, что длина волокна функционально эквивалентна длине пучка, и эти термины используются как синонимы. Исходя из этого предположения, пользовательский код MATLAB (доступен на www.figshare.com — DOI: 10.6084 / m9.figshare.96) был использован для измерения L f из этих волоконных трактов (эквивалентных мышечным пучкам), и приведенные здесь значения являются средними для всего диапазона длин волоконных трактов в каждой мышце. [20]. Это стандартная практика при измерении архитектуры мышц для входных данных в модели мышц типа Хилла [3, 27], и в этом контексте было показано, что оценка L f со средней точностью <1-7 мм [20].
Угол перистости этих волокон был измерен здесь как угол волокон относительно линии действия мышцы.Линия действия каждой мышцы оценивалась с использованием функции «подгонки центральной линии» на каждой трехмерной мышечной сетке в Mimics (из T1-взвешенных МРТ-изображений), которая оценивает линию, проходящую через осевые центроиды каждой сетки, и, следовательно, учитывает их часто -изогнутые формы. Предположение, что эта линия эквивалентна анатомической линии действия, было высказано ранее [6]. Пять поверхностных (2D) измерений угла перистости были записаны вручную в проксимальной, средней и дистальной областях каждой мышцы с помощью ImageJ [28] для получения репрезентативного среднего значения.Это также стандартная практика для оценки этого параметра для моделей опорно-двигательного аппарата, и было показано, что углы перистости поверхности оцениваются со средней точностью 4 ± 1 ° [20].
Все эти методы были выполнены одним и тем же исследователем для каждого субъекта, что обеспечило согласованность представленных данных по архитектуре мышц.
Прогнозирование оптимальной длины волокна
Ранее признанное ограничение измерения длины волокон по изображениям тензора диффузии заключается в том, что оценки оптимальной длины волокон (важный вход для моделей опорно-двигательного аппарата) не могут быть получены с использованием только этого метода.Это связано с тем, что длины саркомеров, которые нормированы на стандартизованную оптимальную длину саркомеров в состоянии покоя для оценки оптимальной длины волокон, не могут быть напрямую измерены по отслеживаемым волокнам. Поэтому оптимальные длины волокон были оценены с использованием длин саркомеров, указанных в [3], с использованием следующего уравнения [29]: где L f — оптимальная длина волокна, L f ’- длина сырого волокна (измеренная по DTI), L s — длина саркомера, а 2,7 мкм — общее значение для оптимальной длины саркомера [29].L s Значения были получены от Ward et al. [3], которые измерили L s в фиксированных мышцах, отсеченных от конечностей с большинством суставов (кроме голеностопного сустава) в анатомическом положении, как в настоящем исследовании.
Эти параметры затем использовались для расчета физиологической площади поперечного сечения (PCSA, мм 2 ), основного определяющего фактора выходной мышечной силы, с использованием уравнения (из [30]): где V m — объем мышцы (живота) (мм 3 ), L f — оптимальная длина мышечного волокна (мм), θ — угол перистости мышечного волокна.Чтобы оценить максимальную изометрическую силу (важный входной параметр для скелетно-мышечных моделей, F max ), индивидуальные значения PCSA умножались на изометрическое напряжение скелетных мышц (или удельное напряжение; 0,3 Н · мм -2 ; [7]). Использование этого общего значения для изометрического стресса хорошо установлено в исследованиях по моделированию опорно-двигательного аппарата [13, 31], и было показано, что оно не зависит от размера тела и сохраняется в филогении позвоночных [32]. Учитывая, что оценка этого значения для каждой отдельной мышцы нижней конечности выходила за рамки данного исследования, здесь предполагалось, что оно будет постоянным для всех мышц.Однако признается, что в действительности это может быть не так, поскольку в литературе сообщается о широком диапазоне значений (0,04–0,6 Нмм 2 ) для мышц нижних конечностей человека, в зависимости от функции или типа волокна [33, 34 ].
Специализация в параметрах мышечной архитектуры в функциональных группах (т.е. определенные функциональные группы мышц, такие как разгибатели колена, демонстрируют в целом схожую ориентацию мышечных волокон и функциональные возможности расширения) ранее была продемонстрирована в опорно-двигательной системе позвоночных [27, 30, 35–41 ].Таким образом, данные об архитектуре мышц, полученные здесь для каждой мышцы, были усреднены по каждой функциональной группе в пределах каждого индивидуума, а также в пределах сгруппированных средних значений (таблицы 1, 2 и таблицы S1 – S10). Это дает общее представление о степени специализации этих мышечных функциональных групп в нижних конечностях людей, участвовавших в этом исследовании, а также позволяет проводить сравнения со средними значениями аналогичных функциональных групп в предыдущих наборах данных по архитектуре (3).
Как эти переменные архитектуры мышц масштабируются в зависимости от массы тела, роста, общего объема конечности (V L ) и длины конечности (L L ) у разных людей в нашей исследуемой популяции, было протестировано с использованием линейной регрессии в GraphPad Prism (La Jolla). , Калифорния, США; www.graphpad.com). Длина конечности определялась как длина от самой проксимальной части большого вертела бедренной кости до самой дистальной части боковой лодыжки малоберцовой кости.
Результаты
Было определено среднее (± стандартное отклонение) in vivo архитектурные свойства для 20 мышц нижних конечностей, измеренные с помощью последовательностей DTI и T1 МРТ у 10 человек (таблица 3). Данные по архитектуре мышц для отдельных субъектов перечислены в таблицах S1 – S10.
Таблица 3.Средние (± стандартное отклонение) архитектурные характеристики 20 мышц нижних конечностей от 10 человек (5 мужчин, 5 женщин; возраст 27,3 ± 3,95 года.
Масса тела 76 ± 12,5 кг) плюс средние значения функциональной группы. L F : L m — Отношение длины мышечных волокон к длине мышц. PCSA — физиологическая площадь поперечного сечения. F max — расчетная максимальная изометрическая сила. Длины саркомеров, используемые для оценки оптимальной длины волокон, были получены от Ward et al., [3].
https: // doi.org / 10.1371 / journal.pone.0223531.t003
В среднем мышцами нижней конечности с наибольшим PCSA были латеральная широкая мышца бедра (3206 ± 1559 мм 2 ), средняя широкая мышца бедра (2938 ± 926 мм 2 ) , и камбаловидной мышцы (3226 ± 1042 мм 2 ). Мышцы с наименьшим PCSA у всех испытуемых: длинный разгибатель большого пальца стопы (196 ± 78 мм 2 ), подколенная мышца (202 ± 76 мм 2 ) и портняжная мышца (333 ± 84 мм 2 ).
Мышцы с самой длинной L f были в среднем; портняжная мышца (408 ± 30 мм), большая приводящая мышца (231 ± 61 мм) и двуглавая мышца бедра (длинная головка) (204 ± 38 мм).Самыми короткими мышцами L f были подколенная мышца (74 ± 14 мм), короткая приводящая мышца (76 ± 22 мм) и медиальная икроножная мышца (97 ± 22 мм). Когда длина волокна была приведена к длине мышцы (L f : L м ), мышцы с самыми большими отношениями L f : L м были; sartorius (0,85 ± <0,01), длинная головка двуглавой мышцы бедра (0,74 ± 0,15), подколенная мышца (0,78 ± 0,17), большая приводящая мышца (0,74 ± 0,17) и латеральная широкая мышца бедра (0,59 ± 0,14). Мышцы с наименьшими L f : L m были камбаловидной (0.32 ± 0,07), медиальная икроножная мышца (0,39 ± 0,08) и прямая мышца бедра (0,39 ± 0,12).
Степень, в которой объем мышцы, длина волокна и максимальная изометрическая сила масштабируются с учетом общего объема конечности (V LM ) и длины конечности (L L ), значительно варьировались между функциональными группами мышц (рисунки 2 и 3; таблица S11. ). Средний объем мышечных групп сильно масштабировался с помощью V LM (рис. 2A и 2B), хотя только сгибатели колена и разгибатели колена показали статистически значимое масштабное соотношение между F max и M L (R 2 > 0.5, р <0,05; Рис. 3C, таблица S11). Длина живота мышцы измерена с L L (рис. 3A и 3B), однако L f не особенно сильно масштабировалась с L L в какой-либо функциональной группе, при этом приводящие мышцы бедра показали самую сильную и только статистически значимую корреляцию (R 2 = 0,49, p = 0,02; рис. 3C, таблица S11).
Рис. 2. Масштабные соотношения между: индивидуальным общим объемом мышц конечности и общим объемом мышц живота в (А) приводящих, сгибателях и разгибателях колена бедра; (B) тыльные сгибатели голеностопного сустава и подошвенные сгибатели голеностопного сустава; и максимальная изометрическая сила в (C) приводящих мышцах бедра, сгибателях колена и разгибателях колена; (D) тыльные сгибатели голеностопного сустава и подошвенные сгибатели голеностопного сустава.
Пунктирные линии представляют собой среднее значение стандартной ошибки ± 1.
https://doi.org/10.1371/journal.pone.0223531.g002
Рис. 3. Масштабные соотношения между: индивидуальной длиной конечности и длиной живота мышцы в (A) приводящих, сгибателях и разгибателях колена бедра; (B) тыльные сгибатели голеностопного сустава и подошвенные сгибатели голеностопного сустава; и длину мышечных волокон в (C) приводящих, сгибателях и разгибателях коленного сустава бедра; (D) тыльные сгибатели голеностопного сустава и подошвенные сгибатели голеностопного сустава.
Пунктирные линии представляют собой среднее значение стандартной ошибки ± 1.
https://doi.org/10.1371/journal.pone.0223531.g003
Обсуждение
В этом исследовании использовалась проверенная методика для получения обширного набора данных in vivo и данных архитектуры мышц нижней конечности человека исключительно из МРТ-изображений 10 молодых здоровых людей. Эти данные определяют объем мышц, длину, оптимальную длину волокна, угол перистости волокна, PCSA и максимальную изометрическую силу. Техника использования DTI и трактографии мышечных волокон для сбора подробных данных по архитектуре мышц была описана ранее и показала свою достоверность и повторяемость [20, 42–56].В исследовании валидности метода сбора архитектуры мышц специально для моделей опорно-двигательного аппарата Чарльз и др. [20] обнаружили, что DTI может воспроизводить мышечную массу, длину волокон и PCSA в пределах 4%, 1% и 6% от соответствующих значений. переменные, измеренные при ручном вскрытии, соответственно. Точность этого метода повысила уверенность в нашей способности генерировать точный и надежный набор данных in vivo и мышечной архитектуры нижних конечностей из популяции молодых здоровых взрослых людей (таблица 2; таблицы S1 – S10).Этот набор данных основан на предыдущих попытках количественно оценить анатомию мышц нижних конечностей с помощью МРТ [6], сосредоточив внимание на сборе данных об архитектуре, необходимых для построения скелетно-мышечных моделей и моделирования.
Степень, в которой эти данные об архитектуре мышц масштабируются с антропометрическими параметрами, такими как длина конечности, объем конечности и масса тела, а также возраст, в функциональных группах мышц, может указывать на необходимость сбора такого обширного набора данных in vivo архитектуры мышц на молодых, здоровых лицах.Объем мышц живота и длина мышечного живота достаточно хорошо соотносились с массой конечностей и длиной конечностей, соответственно, в большинстве функциональных групп. Эти соотношения масштабирования особенно очевидны в мышцах подошвенного сгиба голеностопного сустава, где F max и объем мышц показывают сильную корреляцию с ростом пациента, массой тела, объемом конечности и длиной конечности. Эти результаты согласуются с результатами Хэндсфилда и др. [6], которые сообщили о столь же сильных масштабных взаимосвязях между объемом / длиной живота и длиной конечностей и массой тела.Однако эти данные показывают, что длина мышечных волокон плохо согласуется с длиной конечностей в любой функциональной группе мышц, что согласуется с предыдущими исследованиями [15].
В контексте моделирования опорно-двигательного аппарата это предполагает, что взаимосвязь между длиной мышечного волокна и длиной конечности не обязательно может быть точно предсказана с использованием алгоритмов масштабирования или оптимизации и может быть более сложной, чем другие переменные архитектуры мышц. Таким образом, в то время как антропометрическое масштабирование может использоваться для оценки общих анатомических свойств, таких как объем и длина мышц, визуализация анатомии нижних конечностей для конкретного субъекта, вероятно, необходима для точной оценки более сложных параметров архитектуры мышц, таких как длина мышечных волокон, особенно для использования в опорно-двигательном аппарате. моделирование.Отсутствие прямой корреляции между длиной волокон и длиной конечностей можно объяснить различиями у разных субъектов в длине внешней сухожильной части мышечно-сухожильного блока, которая, как было показано, связана с диапазоном движений сустава, особенно в дистальной части мышцы. группы нижней конечности [57].
Сбор тематических данных дополнительно оправдан различиями между этими данными и ранее опубликованными данными о трупной архитектуре, например, описанными Ward et al., [3]. В то время как общие тенденции в средних характеристиках архитектуры в наших данных близко следовали описанным ранее (многие из тех же мышц имеют большие значения PCSA и большие оптимальные длины волокон), можно увидеть много различий в абсолютных значениях (см. Дополнительную информацию для получения более подробной информации). . Учитывая анатомические вариации, наблюдаемые в нашем наборе данных, эти различия, скорее всего, связаны с разной степенью масштабирования мышечной архитектуры между функциональными группами мышц, а также с потенциальными эффектами старения.В исследовании кинематических и кинетических эффектов старения ДеВита и Хортобаджи [58] предположили, что старение приводит к перераспределению крутящего момента в суставах и мышцах по всей нижней конечности, при относительно более низком голеностопном суставе, но большем крутящем моменте в тазобедренном суставе и силе мышц у пожилых людей по сравнению с молодым людям. Хотя это, вероятно, связано с изменениями кинематики походки с возрастом, это изменение крутящего момента в суставах может также возникать в результате изменений в сложной архитектуре мышц (т.е. уменьшения L f : L m ), которые были показаны здесь посредством сравнения к архитектуре трупных мышц и были особенно очевидны в дистальных мышцах.
Это подтверждает точность наших данных, однако, поскольку объемы мышц и длина волокон, согласно прогнозам, уменьшатся только на 25% и 10% соответственно из-за эффектов старения [4], анатомических вариаций, в дополнение к эффектам фиксации формалина или возможным патологиям в образцы трупов в предыдущих исследованиях вскрытия [3], вероятно, является еще одной важной причиной различий между этими данными и данными о трупах. Этот высокий уровень анатомических вариаций также подтверждает потенциальную потребность в индивидуальном моделировании опорно-двигательного аппарата для клинической оценки.Индивидуализированные модели стали более распространенными [11, 12, 59–62], и с новыми методами сбора архитектуры мышц in vivo эти модели потенциально могут обеспечить более точные и надежные оценки мышечной функции по сравнению с универсальными или масштабированными универсальными моделями.
Ограничения
В то время как этот метод сбора in vivo архитектуры мышц становится все более распространенным [20, 53–56], все еще существуют некоторые ограничения, которые необходимо преодолеть перед широким использованием в сообществе моделирования и моделирования опорно-двигательного аппарата.Многие из этих ограничений, такие как предположения о том, как оценивались углы перистости и длина волокон, аналогичны тем, которые обсуждались ранее [20]. Одним из важных недостатков этого метода, который особенно важен для его приложений для моделирования мышц, является то, что невозможно оценить оптимальную длину волокон. Это часто делается в исследованиях рассечения с использованием лазерной дифракции для измерения длины саркомера [29], однако этот параметр не может быть измерен напрямую по последовательностям DTI. Хотя оптимальные длины волокон были рассчитаны здесь на основе ранее опубликованных длин саркомеров [3], сочетание DTI с дальнейшей медицинской визуализацией, такой как микроэндоскопия [63, 64], для получения in vivo длины саркомера из поверхностных мышц может обеспечить более точные оценки длины саркомера. оптимальная длина волокна в будущих исследованиях.Без специальной поправки на длину саркомера, представленные здесь данные о длине волокон требуют дальнейшего тестирования и оптимизации в рамках скелетно-мышечных моделей, чтобы гарантировать, что мышцы работают на правильной части кривой сила-длина [7].
Следует отметить, что ошибка измерения / наблюдателя могла способствовать отсутствию корреляции, наблюдаемой здесь между антропометрией объекта и архитектурой мышц. Скорее всего, это имело эффект во время ручной сегментации мышц для определения объемов мышц (и, соответственно, расчета F max ) и измерения углов перистости (которые мало влияют на результаты модели опорно-двигательного аппарата — см. Обсуждение ниже).Однако, поскольку определение длины in vivo мышечных волокон было в основном автоматизировано и для каждой мышцы каждого испытуемого использовались одни и те же критерии остановки волоконной трактографии, любые ошибки в этом параметре, вероятно, связаны с различиями в качестве изображения тензора диффузии. изображения, а не человеческая ошибка. Поскольку ручная сегментация мышц позвоночных является часто используемым методом для измерения объема мышц [6], и все изображения тензора диффузии были предварительно обработаны с использованием того же метода (см. Методы) перед анализом, влияние этих потенциальных ошибок на общую представленные здесь результаты, вероятно, были небольшими.
Точность и ограничения используемой здесь структуры волоконной трактографии обсуждались ранее [20]. Однако это оказало прямое влияние на представленные здесь данные. В то время как средняя точность оценки длины мышечных волокон составляла <1-7 мм, она варьировалась между испытуемыми и между группами мышц (2 мм в приводящих мышцах бедра, но 17 мм в разгибателях колена). Эта вариабельность может быть связана с качеством изображения тензора диффузии или артефактами частичного объема от кости или подкожно-жировой клетчатки, которые будут иметь различные эффекты в зависимости от местоположения или размера анализируемой мышцы (см. [45] для подробного обсуждения возможных источников вариации измерения в волоконной трактографии DTI).Несмотря на это изменение, даже большие средние расхождения в точности измерения длины волокон и углов перистости в основном оказываются ниже коэффициентов повторяемости, указанных Heemskerk et al. [45] (<50 мм для L f , <10,2 ° для θ), предполагая, что этот каркас является относительно точным и воспроизводимым. Однако следует отметить, что валидация этого каркаса была проведена на трупах [20], которые не были подвержены тем же физиологическим факторам, которые могли повлиять на повторяемость, описанную Heemskerk et al.[45] (например, движение, артефакты дыхания или температура тела), что может объяснить относительно высокую точность, о которой сообщалось ранее. Тем не менее, хотя этот метод демонстрирует несомненный потенциал для области биомеханического моделирования, для его широкого применения необходимо улучшение согласованности трактографии волокон между мышцами и людьми.
Что касается углов перистости, это исследование сообщает о более низких углах, чем ограниченные трехмерные углы, полученные из трактографии DTI, которые в настоящее время доступны в литературе.Ранее сообщалось о значениях ~ 30 ° для камбаловидной и медиальной икроножных мышц [53, 56], по сравнению со средними значениями 12 ° и 9 ° соответственно, указанными здесь, которые больше похожи на углы, измеренные с помощью ультразвуковых или трупных данных [3, 65–67]. Хотя эти отличия от других исследований DTI кажутся существенными, известно, что угол перистости в значительной степени зависит от положения сустава, и, по оценкам, с помощью DTI он изменяется от 9 ° [56] до 46 ° [46] с поворотами на 30 ° при тыльном / подошвенном сгибании голеностопного сустава. .Хотя субъектов в этом исследовании просили оставаться в анатомическом положении во время получения изображения (с тазобедренным, коленным и голеностопным суставами при 0 ° сгибания / разгибания), возможно, что голеностопный сустав не находился точно в этом положении для продолжительность каждого сканирования. Даже небольшие отклонения от нейтрального положения в голеностопном суставе могли вызвать большие изменения в углах перистости, особенно в тыльных или подошвенных мышцах голеностопного сустава, и, следовательно, могли объяснить эти различия.Однако, учитывая низкую чувствительность прогнозов мышечной функции в скелетно-мышечных моделях к вариациям входного параметра угла перистости [10, 13], двухмерные углы перистости поверхности, представленные здесь, могут быть достаточными для прогнозирования функций мышц, если эти данные будут использоваться в качестве входы в такие модели. Тем не менее, дальнейшее уточнение этой схемы для более точной оценки оптимальной длины волокна и углов перистости может оказаться полезным в будущих исследованиях.
Воздействие и будущее исследование
Это исследование представляет собой первый экземпляр обширного набора данных in vivo мышечной архитектуры нижних конечностей человека, созданного исключительно на основе медицинских изображений (DTI и MRI), с особым акцентом на последствиях для биомеханики и моделирования опорно-двигательного аппарата.Путем исследования масштабных соотношений между антропометрическими параметрами и важными свойствами генерирования мышечной силы, такими как длина мышечного волокна и максимальная изометрическая сила, эти данные показывают отсутствие корреляции между длиной мышечного волокна и антропометрией среди большинства функциональных групп мышц нижней конечности. Это означает, что алгоритмы оптимизации или масштабирования, часто используемые для оценки архитектуры мышц при моделировании опорно-двигательного аппарата, могут быть ненадежными, и что то, как длина мышечных волокон связана с пропорциями тела, может быть более сложным по сравнению с аналогичными отношениями с другими переменными архитектуры мышц.Тем не менее, учитывая различия между этими данными и ранее опубликованными данными об архитектуре трупа, возможно, что применение представленных здесь мышечных параметров к скелетно-мышечным моделям людей аналогичного возраста или антропометрии может дать более точные оценки мышечной функции, чем аналогичные данные из предыдущих исследований. . Кроме того, точные траектории мышечных волокон, реконструированные с помощью этого метода, также могут улучшить прогнозирование мышечной функции за счет более точного представления рычагов мышечного момента и могут быть включены в биомеханические модели с использованием таких методов, как описанные Chen et al., [68].
В то время как это исследование было сосредоточено на оценке архитектуры мышц у молодых, здоровых людей с использованием DTI, эта схема могла бы принести дополнительную пользу в области моделирования опорно-двигательного аппарата, измеряя аналогичные параметры в патологических популяциях (например, у людей с церебральным параличом [54], мышечной атрофией [69], мышечной дистрофия [70] и пожилые люди [71]), походка и функции мышц которых часто исследуются с помощью биомеханических моделей и симуляций [72, 73].
Кроме того, различия с предыдущими данными, а также различия в нашем наборе данных также подтверждают появление предметно-ориентированного моделирования опорно-двигательного аппарата.Хотя общие и масштабно-обобщенные модели обычно эффективны при тестировании общих прогнозов опорно-двигательного аппарата, более подробный анализ моделирования, например, прогнозирующий реабилитационные или послеоперационные результаты, может потребовать включения данных по архитектуре мышц для максимальной эффективности. Дальнейшая работа будет сосредоточена на проверке этих предположений и дальнейшей валидации этой структуры. Несмотря на то, что методы, используемые здесь для измерения архитектуры мышц in vivo , ранее были проверены специально для использования в моделировании опорно-двигательного аппарата [20], до сих пор неясно, насколько точно данные будут моделировать функции мышц в рамках этих моделей.Достоверность этих методов может быть дополнительно оценена путем сравнения мышечных сил, прогнозируемых с помощью моделей опорно-двигательного аппарата для конкретных субъектов, с экспериментально измеренными, например, с изокинетического динамометра. Точные прогнозы мышечных сил на основе моделей для конкретных субъектов еще больше повысили бы уверенность в достоверности этой схемы при измерении архитектуры мышц in vivo, и формировании основы индивидуализированных скелетно-мышечных моделей.
Вспомогательная информация
S1 Таблица.Архитектурные характеристики 20 мышц нижних конечностей субъекта 01 плюс средние значения функциональной группы (± стандартное отклонение).
Длины волокон и углы перистости выражены как средние значения (± стандартное отклонение) нескольких измерений, выполненных на разных участках каждой мышцы. L f : L м — отношение длины мышечных волокон к длине волокон. PCSA — физиологическая площадь поперечного сечения. F max — расчетная максимальная изометрическая сила. Длины саркомеров, используемые для оценки оптимальной длины волокон, были получены от Ward et al., [3].
https://doi.org/10.1371/journal.pone.0223531.s001
(DOCX)
S2 Стол. Архитектурные характеристики 20 мышц нижних конечностей Субъекта 02 плюс средние значения функциональной группы (± стандартное отклонение).
Длины волокон и углы перистости выражены как средние значения (± стандартное отклонение) нескольких измерений, выполненных на разных участках каждой мышцы. L f : L м — отношение длины мышечных волокон к длине волокон. PCSA — физиологическая площадь поперечного сечения.F max — расчетная максимальная изометрическая сила. Длины саркомеров, используемые для оценки оптимальной длины волокон, были получены от Ward et al., [3].
https://doi.org/10.1371/journal.pone.0223531.s002
(DOCX)
S3 Таблица. Архитектурные характеристики 20 мышц нижних конечностей Субъекта 03 плюс средние значения функциональной группы (± стандартное отклонение).
Длины волокон и углы перистости выражены как средние значения (± стандартное отклонение) нескольких измерений, выполненных на разных участках каждой мышцы.L f : L м — отношение длины мышечных волокон к длине волокон. PCSA — физиологическая площадь поперечного сечения. F max — расчетная максимальная изометрическая сила. Длины саркомеров, используемые для оценки оптимальной длины волокон, были получены от Ward et al., [3].
https://doi.org/10.1371/journal.pone.0223531.s003
(DOCX)
S4 Таблица. Архитектурные характеристики 20 мышц нижних конечностей субъекта 04 плюс средние значения функциональной группы (± стандартное отклонение).
Длины волокон и углы перистости выражены как средние значения (± стандартное отклонение) нескольких измерений, выполненных на разных участках каждой мышцы.L f : L м — отношение длины мышечных волокон к длине волокон. PCSA — физиологическая площадь поперечного сечения. F max — расчетная максимальная изометрическая сила. Длины саркомеров, используемые для оценки оптимальной длины волокон, были получены от Ward et al., [3].
https://doi.org/10.1371/journal.pone.0223531.s004
(DOCX)
S5 Таблица. Архитектурные характеристики 20 мышц нижних конечностей субъекта 05 плюс средние значения функциональной группы (± стандартное отклонение).
Длины волокон и углы перистости выражены как средние значения (± стандартное отклонение) нескольких измерений, выполненных на разных участках каждой мышцы.L f : L м — отношение длины мышечных волокон к длине волокон. PCSA — физиологическая площадь поперечного сечения. F max — расчетная максимальная изометрическая сила. Длины саркомеров, используемые для оценки оптимальной длины волокон, были получены от Ward et al., [3].
https://doi.org/10.1371/journal.pone.0223531.s005
(DOCX)
S6 Таблица. Архитектурные характеристики 20 мышц нижних конечностей Субъекта 06 плюс средние значения функциональной группы (± стандартное отклонение).
Длины волокон и углы перистости выражены как средние значения (± стандартное отклонение) нескольких измерений, выполненных на разных участках каждой мышцы.L f : L м — отношение длины мышечных волокон к длине волокон. PCSA — физиологическая площадь поперечного сечения. F max — расчетная максимальная изометрическая сила. Длины саркомеров, используемые для оценки оптимальной длины волокон, были получены от Ward et al., [3].
https://doi.org/10.1371/journal.pone.0223531.s006
(DOCX)
S7 Таблица. Архитектурные характеристики 20 мышц нижних конечностей субъекта 07 плюс средние значения функциональной группы (± стандартное отклонение).
Длины волокон и углы перистости выражены как средние значения (± стандартное отклонение) нескольких измерений, выполненных на разных участках каждой мышцы.L f : L м — отношение длины мышечных волокон к длине волокон. PCSA — физиологическая площадь поперечного сечения. F max — расчетная максимальная изометрическая сила. Длины саркомеров, используемые для оценки оптимальной длины волокон, были получены от Ward et al., [3].
https://doi.org/10.1371/journal.pone.0223531.s007
(DOCX)
S8 Таблица. Архитектурные характеристики 20 мышц нижних конечностей субъекта 08 плюс средние значения функциональной группы (± стандартное отклонение).
Длины волокон и углы перистости выражены как средние значения (± стандартное отклонение) нескольких измерений, выполненных на разных участках каждой мышцы.L f : L м — отношение длины мышечных волокон к длине волокон. PCSA — физиологическая площадь поперечного сечения. F max — расчетная максимальная изометрическая сила. Длины саркомеров, используемые для оценки оптимальной длины волокон, были получены от Ward et al., [3].
https://doi.org/10.1371/journal.pone.0223531.s008
(DOCX)
S9 Таблица. Архитектурные характеристики 20 мышц нижних конечностей Субъекта 09 плюс средние значения функциональной группы (± стандартное отклонение).
Длины волокон и углы перистости выражены как средние значения (± стандартное отклонение) нескольких измерений, выполненных на разных участках каждой мышцы.L f : L м — отношение длины мышечных волокон к длине волокон. PCSA — физиологическая площадь поперечного сечения. F max — расчетная максимальная изометрическая сила. Длины саркомеров, используемые для оценки оптимальной длины волокон, были получены от Ward et al., [3].
https://doi.org/10.1371/journal.pone.0223531.s009
(DOCX)
S10 Стол. Архитектурные характеристики 20 мышц нижних конечностей испытуемого 10 плюс средние значения функциональной группы (± стандартное отклонение).
Длины волокон и углы перистости выражены как средние значения (± стандартное отклонение) нескольких измерений, выполненных на разных участках каждой мышцы.L f : L м — отношение длины мышечных волокон к длине волокон. PCSA — физиологическая площадь поперечного сечения. F max — расчетная максимальная изометрическая сила. Длины саркомеров, используемые для оценки оптимальной длины волокон, были получены от Ward et al., [3].
https://doi.org/10.1371/journal.pone.0223531.s010
(DOCX)
S11 Стол. Результаты линейной регрессии (значения R
2 ) для проверки масштабных соотношений между параметрами архитектуры мышц (L f- длина мышечного волокна; L f : L м — отношение длины волокна к длине мышцы; F max- расчетная максимальная изометрическая сила; V м — объем мышц ; L м — длина мышцы) и возраст субъекта, рост, масса тела, общая масса конечностей и длина нижней конечности.Значения P указаны в скобках. Значения, выделенные курсивом и жирным шрифтом, указывают на статистическую значимость (p ≤ 0,05).
https://doi.org/10.1371/journal.pone.0223531.s011
(DOCX)
S12 Стол. Средние различия (% различий) между данными по архитектуре мышц нижних конечностей, полученными здесь из МРТ, и данными из предыдущего исследования трупов [3].
Мышечные массы по нашим данным оценивались по объемам мышц. L f : L m — Отношение длины мышечных волокон к длине мышц.PCSA — физиологическая площадь поперечного сечения. F max — расчетная максимальная изометрическая сила. Длины саркомеров, используемые для оценки оптимальной длины волокон, были получены от Ward et al., [3].
https://doi.org/10.1371/journal.pone.0223531.s012
(DOCX)
S2 Рис. Средние расчетные значения максимальной изометрической силы (F
max ; выражены в процентах массы тела) для каждой функциональной группы мышц для субъектов 1–10.Классификацию мышечных функциональных групп см. В таблице 1.Горизонтальная пунктирная линия представляет собой среднее значение из Ward et al., [3].
https://doi.org/10.1371/journal.pone.0223531.s014
(TIF)
Благодарности
Авторы хотели бы поблагодарить Chan Hong Moon за помощь в создании протоколов МРТ, используемых здесь, и Tom Gale за написание сценария MATLAB, используемого для измерения длины волокон мышечных волокон, отслеживаемых DTI. Мы также хотели бы поблагодарить трех рецензентов первоначальных версий этой рукописи, чьи конструктивные комментарии помогли значительно улучшить окончательную версию.Работа выполнена при финансовой поддержке отделения ортопедической хирургии Питтсбургского университета.
Ссылки
- 1. Либер Р.Л., Фриден Дж. Функциональное и клиническое значение архитектуры скелетных мышц. Мышечный нерв. 2000. 23 (11): 1647–66. pmid: 11054744.
- 2. Вицкевич Т.Л., Рой Р.Р., Пауэлл П.Л., Эджертон В.Р. Мышечная архитектура нижней конечности человека. Clin Orthop Relat Res. 1983; (179): 275–83. pmid: 6617027.
- 3.Ward SR, Eng CM, Smallwood LH, Lieber RL. Точны ли текущие измерения архитектуры мышц нижних конечностей? Clin Orthop Relat Res. 2009. 467 (4): 1074–82. pmid: 18972175.
- 4. Наричи М.В., Маганарис С.Н., Ривз Н.Д., Каподаглио П. Влияние старения на архитектуру мышц человека. J. Appl Physiol (1985). 2003. 95 (6): 2229–34. Epub 2003/07/08. pmid: 12844499.
- 5. Тейт CM, Уильямс GN, Барранс П.Дж., Бьюкенен Т.С. Морфология мышц нижних конечностей у юных спортсменов: анализ на основе МРТ.Медико-спортивные упражнения. 2006. 38 (1): 122–8. Epub 2006/01/06. pmid: 16394964.
- 6. Хэндсфилд Г.Г., Мейер СН, Харт Дж.М., Абель М.Ф., Блемкер СС. Связь 35 мышц нижних конечностей с ростом и массой тела количественно определена с помощью МРТ. J Biomech. 2014; 47 (3): 631–8. Epub 2013/12/26. pmid: 24368144.
- 7. Zajac FE. Мышцы и сухожилия: свойства, модели, масштабирование и применение в биомеханике и моторном управлении. Crit Rev Biomed Eng. 1989. 17 (4): 359–411. pmid: 2676342.
- 8.Сковил CY, Ронски JL. Чувствительность модели мышц Хилла к отклонениям в параметрах модели. J Biomech. 2006. 39 (11): 2055–63. Epub 2005/08/09. pmid: 16084520.
- 9. Акленд, округ Колумбия, Линь Ю.К., Панди М.Г. Чувствительность модельных прогнозов мышечной функции к изменениям в моментальных руках и свойствах мышц-сухожилий: анализ Монте-Карло. J Biomech. 2012. 45 (8): 1463–71. Epub 2012/04/14. pmid: 22507351.
- 10. О’Нил М.С., Ли Л.Ф., Ларсон С.Г., Демес Б., Стерн Дж. Т., Умбергер Б.Р.Трехмерная модель опорно-двигательного аппарата таза и задней конечности шимпанзе ( Pan troglodytes ). J Exp Biol. 2013; 216 (Pt 19): 3709–23. pmid: 24006347.
- 11. Валенте Дж., Питто Л., Тести Д., Сет А., Делп С. Л., Стагни Р. и др. Устойчивы ли индивидуальные модели опорно-двигательного аппарата к неопределенностям в идентификации параметров? PLoS One. 2014; 9 (11): e112625. Epub 2014/11/12. pmid: 253 .
- 12. Navacchia A, Myers CA, Rullkoetter PJ, Shelburne KB.Прогнозирование нагрузок на коленный сустав in vivo с помощью глобального вероятностного анализа. J Biomech Eng. 2016; 138 (3): 4032379. pmid: 26720096.
- 13. Чарльз Дж. П., Каппеллари О., Спенс А. Дж., Уэллс Д. Д., Хатчинсон-младший. Мышечный момент руки и анализ чувствительности модели опорно-двигательного аппарата задних конечностей мыши. J Anat. 2016; 229 (4): 514–35. pmid: 27173448.
- 14. Буяльски П., Мартинс Дж., Стирлинг Л. Анализ методом Монте-Карло оценки чувствительности оценки мышечной силы к свойствам мышцы-сухожилия с использованием модели мышц на основе Хилла.J Biomech. 2018. Epub 2018/08/28. pmid: 30146173.
- 15. Уорд С.Р., Смоллвуд Л.Х., Либер Р.Л. Масштабирование архитектуры мышц нижних конечностей человека до размеров скелета. В: XX Конгресс ISB, Кливленд, Огайо. 2005.
- 16. Modenese L, Ceseracciu E, Reggiani M, Lloyd DG. Оценка параметров опорно-двигательного аппарата для масштабированных и конкретных моделей опорно-двигательного аппарата с использованием метода оптимизации. J Biomech. 2016; 49 (2): 141–8. Epub 2015/11/18. pmid: 26776930.
- 17. Манал К., Бьюкенен Т. Оценка длины провисания сухожилия по конкретным предметам: численный метод. Журнал прикладной биомеханики. 2004. 20 (2): 195–203.
- 18. Винби ЧР, Ллойд Д.Г., Кирк ТБ. Оценка различных аналитических методов для индивидуального масштабирования параметров опорно-двигательного аппарата. J Biomech. 2008. 41 (8): 1682–8. Epub 2008/05/06. pmid: 18456272.
- 19. Ву В., Ли П.В., Брайант А.Л., Галеа М., Окленд, округ Колумбия. Индивидуальное моделирование опорно-двигательного аппарата при оценке функции плечевых мышц и суставов.J Biomech. 2016; 49 (15): 3626–34. Epub 2016/09/23. pmid: 28327299.
- 20. Чарльз Дж. П., Мун СН, Андерст В. Определение данных об архитектуре мышц нижних конечностей для моделей опорно-двигательного аппарата с использованием МРТ с тензором диффузии. J Biomech Eng. 2019; 141 (6): 060905–9. pmid: 30098157.
- 21. Прейбиш С., Заальфельд С., Томанчак П. Оптимальное сшивание мозаичных трехмерных изображений при получении микроскопических изображений. Биоинформатика. 2009. 25 (11): 1463–5. Epub 2009/04/07. pmid: 19346324.
- 22. Schindelin J, Arganda-Carreras I, Frize E, Kaynig V, Longair M, Pietzsch T. и др. Фиджи: платформа с открытым исходным кодом для анализа биологических изображений. Нат методы. 2012. 9 (7): 676–82. Epub 2012/06/30. pmid: 22743772.
- 23. Jiang H, van Zijl PC, Kim J, Pearlson GD, Mori S. DtiStudio: ресурсная программа для вычисления тензора диффузии и отслеживания пучков волокон. Вычислительные методы Программы Biomed. 2006. 81 (2): 106–16. Epub 2006/01/18. pmid: 16413083.
- 24.Аха-Фернандес С., Нитхаммер М., Кубики М., Шентон М.Э., Вестин С.Ф. Восстановление данных DWI с использованием оценщика Rician LMMSE. IEEE Trans Med Imaging. 2008. 27 (10): 1389–403. Epub 2008/09/26. pmid: 18815091.
- 25. Cook P, Bai Y, Nedjati-Gilani S, Seunarine K, Hall M, Parker G, et al. Camino: Реконструкция и обработка диффузионной МРТ с открытым исходным кодом. 14-е научное собрание Международного общества магнитного резонанса в медицине, Сиэтл, Вашингтон, США, 2006 г. п. 2759.
- 26.Бодин С.К., Рой Р.Р., Медоуз Д.А., Зернике Р.Ф., Сакс Р.Д., Фурнье М. и др. Архитектурные, гистохимические и сократительные характеристики уникальной двусуставной мышцы: полусухожильной мышцы кошки. J Neurophysiol. 1982; 48 (1): 192–201. pmid: 7119845.
- 27. Чарльз Дж. П., Каппеллари О., Спенс А. Дж., Хатчинсон-младший, Уэллс ди-джей. Скелетно-мышечная геометрия, архитектура мышц и функциональные особенности задней конечности мыши. PLoS One. 2016; 11 (4): e0147669. pmid: 27115354.
- 28.Шнайдер CA, Rasband WS, Eliceiri KW. NIH Image to ImageJ: 25 лет анализа изображений. Нат методы. 2012; 9 (7): 671–5. pmid: 22930834.
- 29. Фельдер А., Уорд С.Р., Либер Р.Л. Измерение длины саркомера позволяет с высоким разрешением нормализовать длину мышечных волокон в архитектурных исследованиях. J Exp Biol. 2005; 208 (Pt 17): 3275–9. Epub 2005/08/20. pmid: 16109889.
- 30. Мешки Р. Д., Рой Р. Р.. Архитектура мышц задних конечностей кошек: функциональное значение.J Morphol. 1982. 173 (2): 185–95. pmid: 7120421.
- 31. Хатчинсон-младший. Биомеханическое моделирование и анализ чувствительности двуногого бега. I. Сохранившиеся таксоны. J Morphol. 2004. 262 (1): 421–40. pmid: 15352201.
- 32. Медлер С. Сравнительные тенденции в сокращении скорости и производства силы в скелетных мышцах. Am J Physiol Regul Integr Comp Physiol. 2002; 283 (2): R368–78. pmid: 12121850.
- 33. Фукунага Т., Рой Р. Р., Шеллок Ф. Г., Ходжсон Дж. А., Эджертон В. Р..Удельное напряжение подошвенных сгибателей и дорсифлексоров человека. J. Appl Physiol (1985). 1996. 80 (1): 158–65. Epub 1996/01/01. pmid: 8847297.
- 34. Maganaris CN, Baltzopoulos V, Ball D, Sargeant AJ. Специфическое напряжение скелетных мышц человека in vivo. J. Appl Physiol (1985). 2001; 90 (3): 865–72. Epub 2001/02/22. pmid: 11181594.
- 35. Пауэлл П.Л., Рой Р.Р., Каним П., Белло М.А., Эджертон В.Р. Предсказуемость напряжения скелетных мышц на основе архитектурных определений задних конечностей морских свинок.J Appl Physiol Respir Environ Exerc Physiol. 1984. 57 (6): 1715–21. pmid: 6511546.
- 36. Либер Р.Л., Блевинс Ф.Т. Архитектура скелетных мышц задней конечности кролика: функциональные последствия дизайна мышц. J Morphol. 1989; 199 (1): 93–101. pmid: 2921772.
- 37. Пейн Р. К., Хатчинсон-младший, Робиллиард Дж. Дж., Смит Н. С., Уилсон А. М.. Функциональная специализация анатомии тазовых конечностей у лошадей ( Equus caballus ). J Anat. 2005. 206 (6): 557–74. pmid: 15960766.
- 38.Уильямс С.Б., Уилсон А.М., Родс Л., Эндрюс Дж., Пейн Р.С. Функциональная анатомия и мышечный момент рук тазовой конечности элитного спортсмена-спринтера: гоночной борзой ( Canis ownis ). J Anat. 2008. 213 (4): 361–72. pmid: 18657259.
- 39. Уильямс С.Б., Пейн Р.С., Уилсон А.М. Функциональная специализация тазовой конечности зайца ( Lepus europeus ). J Anat. 2007. 210 (4): 472–90. pmid: 17362487.
- 40. Аллен В., Элси Р.М., Джонс Н., Райт Дж., Хатчинсон-младший.Функциональная специализация и онтогенетическое масштабирование анатомии конечностей у животных Alligator mississippiensis . J Anat. 2010. 216 (4): 423–45. pmid: 20148991.
- 41. Пакстон Х., Энтони Н.Б., Корр С.А., Хатчинсон-младший. Влияние селективного разведения на архитектурные свойства тазовой конечности у цыплят-бройлеров: сравнительное исследование современных и исконных популяций. J Anat. 2010. 217 (2): 153–66. pmid: 20557402.
- 42. Хсу Э. У., Мори С. Аналитические выражения для коэффициентов кажущейся диффузии ЯМР в анизотропной системе и упрощенный метод определения ориентации волокна.Magn Reson Med. 1995. 34 (2): 194–200. pmid: 7476078.
- 43. Дэймон Б.М., Динг З., Андерсон А.В., Фрейер А.С., Гор Дж. Подтверждение отслеживания мышечных волокон на основе тензора диффузии с помощью МРТ. Magn Reson Med. 2002. 48 (1): 97–104. pmid: 12111936.
- 44. Deux JF, Malzy P, Paragios N, Bassez G, Luciani A, Zerbib P и др. Оценка сокращения икроножной мышцы с помощью тензорной диффузионной визуализации. Eur Radiol. 2008. 18 (10): 2303–10. Epub 2008/05/09. pmid: 18463875.
- 45. Heemskerk AM, Sinha TK, Wilson KJ, Ding Z, Damon BM.Повторяемость отслеживания волокон скелетных мышц на основе DTI. ЯМР Биомед. 2010. 23 (3): 294–303. pmid: 20099372.
- 46. Sinha U, Sinha S, Hodgson JA, Edgerton RV. Архитектура камбаловидной мышцы человека под разными углами голеностопного сустава по данным магнитно-резонансной диффузной тензорной визуализации. J. Appl Physiol (1985). 2011; 110 (3): 807–19. pmid: 21164150.
- 47. Froeling M, Nederveen AJ, Heijtel DF, Lataster A, Bos C, Nicolay K и др. Диффузионно-тензорная МРТ выявляет сложную мышечную архитектуру предплечья человека.J. Магнитно-резонансная томография. 2012. 36 (1): 237–48. pmid: 22334539.
- 48. Соарес Дж. М., Маркес П., Алвес В., Соуза Н. Автостопом по визуализации диффузионного тензора. Front Neurosci. 2013; 7: 31. pmid: 23486659.
- 49. Bolsterlee B, Veeger HE, van der Helm FC, Gandevia SC, Herbert RD. Сравнение измерений архитектурных параметров медиальной икроножной мышцы по ультразвуковым и диффузным тензорным изображениям. J Biomech. 2015; 48 (6): 1133–40. Epub 2015/02/16. pmid: 25682540.
- 50. Froeling M, Oudeman J, Strijkers GJ, Maas M, Drost MR, Nicolay K и др. Изменения мышц, обнаруженные с помощью диффузионно-тензорной визуализации после бега на длинные дистанции. Радиология. 2015. 274 (2): 548–62. Epub 2014/10/03. pmid: 25279435.
- 51. Дэймон Б.М., Фроилинг М., Бак А.К., Аудеман Дж., Динг З., Недервен А.Дж. и др. Отслеживание волокон с помощью тензорной диффузии скелетных мышц и МРТ: обоснование, методы сбора и анализа данных, приложения и будущие направления. ЯМР Биомед.2016. Epub 2016/06/03. pmid: 27257975.
- 52. Sieben JM, van Otten I., Lataster A, Froeling M, Nederveen AJ, Strijkers GJ, et al. Реконструкция in vivo архитектуры поясничного эректора позвоночника с использованием диффузно-тензорной МРТ. Clin Spine Surg. 2016; 29 (3): E139–45. pmid: 27007789.
- 53. Болстерли Б., Финни Т., Д’Суза А., Эгучи Дж., Кларк Е.К., Герберт Р.Д. Трехмерная архитектура всей камбаловидной мышцы человека in vivo. PeerJ. 2018; 6: e4610. Epub 2018/04/24.pmid: 29682414.
- 54. Сахрманн А.С., Стотт Н.С., Безье Т.Ф., Фернандес Дж.В., Хэндсфилд Г.Г. Слабость камбаловидной мышцы при церебральном параличе: архитектура мышц выявлена с помощью диффузно-тензорной визуализации. PLoS One. 2019; 14 (2): e0205944. Epub 2019/02/26. pmid: 30802250.
- 55. Bolsterlee B, D’Souza A, Herbert RD. Надежность и надежность измерений архитектуры мышц, полученных с помощью тензорной диффузионной визуализации с анатомически ограниченной трактографией. J Biomech. 2019; 86: 71–8.Epub 2019/02/12. pmid: 30739766.
- 56. Bolsterlee B, D’Souza A, Gandevia SC, Herbert RD. Как пассивное удлинение меняет архитектуру медиальной икроножной мышцы человека? J. Appl Physiol (1985). 2017; 122 (4): 727–38. Epub 2017/01/21. pmid: 28104754.
- 57. Мацукиё А., Гох А.С., Асагай Ю. Взаимосвязь между длиной мышечного сухожилия, диапазоном движений и сопротивлением пассивному движению у детей с нормальным и повышенным тонусом. J Phys Ther Sci.2017; 29 (2): 349–55. Epub 2017/03/08. pmid: 28265172.
- 58. ДеВита П., Хортобадьи Т. Возраст вызывает перераспределение суставных моментов и сил во время ходьбы. J. Appl Physiol (1985). 2000. 88 (5): 1804–11. Epub 2000/05/08. pmid: 10797145.
- 59. Scheys L, Spaepen A, Suetens P, Jonkers I. Расчетные длины момент-рука и мышца-сухожилие во время походки существенно различаются с использованием МРТ по сравнению с масштабированными стандартными скелетно-мышечными моделями нижних конечностей. Поза походки. 2008. 28 (4): 640–8.Epub 2008/06/04. pmid: 18534855.
- 60. Scheys L, Loeckx D, Spaepen A, Suetens P, Jonkers I. Нежесткая регистрация изображений на основе Атласа для автоматического определения моделей мышц линии действия: исследование для проверки. J Biomech. 2009. 42 (5): 565–72. Epub 2009/02/24. pmid: 19232618.
- 61. Scheys L, Desloovere K, Suetens P, Jonkers I. Уровень детализации предмета в скелетно-мышечных моделях влияет на расчет длины плеча тазобедренного сустава во время походки у детей с повышенной антеверсией бедренной кости.J Biomech. 2011. 44 (7): 1346–53. Epub 2011/02/03. pmid: 21295307.
- 62. Prinold JA, Mazzà C, Di Marco R, Hannah I, Malattia C, Magni-Manzoni S, et al. Модель стопы для конкретного пациента для оценки силы голеностопного сустава у пациентов с ювенильным идиопатическим артритом. Энн Биомед Eng. 2016; 44 (1): 247–57. Epub 2015/09/15. pmid: 26374518.
- 63. Чен Икс, Санчес Г.Н., Шнитцер М.Дж., Delp SL. Изменения длины саркомера латеральной широкой мышцы бедра человека при сгибании колена, измеренные с помощью микроэндоскопии in vivo.J Biomech. 2016; 49 (13): 2989–94. Epub 2016/08/03. pmid: 27481293.
- 64. Чен X, Delp SL. Длина саркомера камбаловидной мышцы человека измерена с помощью микроэндоскопии in vivo при двух углах сгибания голеностопного сустава. J Biomech. 2016; 49 (16): 4164–7. Epub 2016/11/22. pmid: 27866676.
- 65. Herbert RD, Heroux ME, Diong J, Bilston LE, Gandevia SC, Lichtwark GA. Изменения длины и трехмерной ориентации мышечных пучков и апоневрозов при пассивных изменениях длины икроножных мышц человека.J Physiol. 2015; 593 (2): 441–55. Epub 2015/01/30. pmid: 25630264.
- 66. Maganaris CN, Baltzopoulos V, Sargeant AJ. Измерения in vivo сложной архитектуры трицепса surae у человека: последствия для мышечной функции. J Physiol. 1998; 512 (Pt 2): 603–14. Epub 1998/10/09. pmid: 9763648.
- 67.
Наричи М.В., Бинзони Т., Хильтбранд Э., Фазель Дж., Терьер Ф., Черретелли П. Архитектура икроножной мышцы человека in vivo с изменяющимся углом сустава в состоянии покоя и во время постепенного изометрического сокращения.J Physiol. 1996; 496 (Pt 1): 287–97. Epub 1996/10/01. pmid: 8
6.
- 68. Чен Дж. С., Басава Р. Р., Чжан Ю., Чапо Р., Малис В., Синха Ю. и др. Пиксельное моделирование скелетных мышц без сетки. Comput Methods Biomech Biomed Eng Imaging Vis. 2016; 4 (2): 73–85. Epub 2016/01/01. pmid: 28748126.
- 69. Малис В., Синха У., Чапо Р., Наричи М., Смитаман Э, Синха С. Визуализация тензора диффузии и моделирование диффузии: применение для мониторинга изменений в медиальной икроножной мышце при атрофии неиспользования, вызванной односторонним подвешиванием конечностей.J. Магнитно-резонансная томография. 2019; 49 (6): 1655–64. Epub 2018/12/21. pmid: 30569482.
- 70. Ponrartana S, Ramos-Platt L, Wren TA, Hu HH, Perkins TG, Chia JM и др. Эффективность визуализации тензора диффузии в оценке тяжести заболевания при мышечной дистрофии Дюшенна: предварительное исследование. Pediatr Radiol. 2015; 45 (4): 582–9. Epub 2014/09/24. pmid: 25246097.
- 71. Sinha U, Csapo R, Malis V, Xue Y, Sinha S. Возрастные различия в индексах тензора диффузии и архитектуре волокон в медиальной и латеральной икроножной мышце.J. Магнитно-резонансная томография. 2015; 41 (4): 941–53. Epub 2014/04/29. pmid: 24771672.
- 72. Розенберг М, Стил К. Моделирование воздействия ортезов голеностопного сустава на потребность в мышцах и их набор у типично развивающихся детей и детей с церебральным параличом и походкой приседания. PLoS One. 2017; 12 (7): e0180219. Epub 2017/07/14. pmid: 28704464.
- 73. Стил К.М., Сет А., Хикс Дж. Л., Шварц М. С., Делп С. Л.. Вклад мышц в поддержку и прогресс во время ходьбы на одной конечности.J Biomech. 2010. 43 (11): 2099–105. Epub 2010/05/25. pmid: 20493489.
% PDF-1.6 % 97 0 объект > эндобдж 84 0 объект > поток 2020-08-04T16: 13: 21-04: 002021-05-21T08: 03: 05-07: 002021-05-21T08: 03: 05-07: 00itext-paulo-155 (itextpdf.sf.net-lowagie. com) application / pdfuuid: 240ae2aa-cf0c-435d-85c7-203b751af86auuid: 1ca56a2a-1dd2-11b2-0a00-b80018d1aeff конечный поток эндобдж 88 0 объект > эндобдж 2 0 obj > эндобдж 91 0 объект > / Font> / ProcSet [/ PDF / Text] >> / Rotate 0 / Type / Page >> эндобдж 1 0 obj > / Font> / ProcSet [/ PDF / Text] >> / Rotate 0 / Type / Page >> эндобдж 8 0 объект > / ExtGState> / Font> / ProcSet [/ PDF / Text] >> / Rotate 0 / Type / Page >> эндобдж 13 0 объект > / Font> / ProcSet [/ PDF / Text] >> / Rotate 0 / Type / Page >> эндобдж 15 0 объект > / Font> / ProcSet [/ PDF / Text] >> / Rotate 0 / Type / Page >> эндобдж 17 0 объект > / Font> / ProcSet [/ PDF / Text] >> / Rotate 0 / Type / Page >> эндобдж 19 0 объект > / Font> / ProcSet [/ PDF / Text] >> / Rotate 0 / Type / Page >> эндобдж 21 0 объект > / Font> / ProcSet [/ PDF / Text] >> / Rotate 0 / Type / Page >> эндобдж 24 0 объект > / Font> / ProcSet [/ PDF / Text] >> / Rotate 0 / Type / Page >> эндобдж 26 0 объект > / Font> / ProcSet [/ PDF / Text] >> / Rotate 0 / Type / Page >> эндобдж 28 0 объект > / Font> / ProcSet [/ PDF / Text] >> / Rotate 0 / Type / Page >> эндобдж 30 0 объект > / Font> / ProcSet [/ PDF / Text] >> / Rotate 0 / Type / Page >> эндобдж 32 0 объект > / Font> / ProcSet [/ PDF / Text] >> / Rotate 0 / Type / Page >> эндобдж 34 0 объект > / Font> / ProcSet [/ PDF / Text] >> / Rotate 0 / Type / Page >> эндобдж 36 0 объект > / Font> / ProcSet [/ PDF / Text] >> / Rotate 0 / Type / Page >> эндобдж 38 0 объект > / Font> / ProcSet [/ PDF / Text] >> / Rotate 0 / Type / Page >> эндобдж 40 0 объект > / ExtGState> / Font> / ProcSet [/ PDF / Text] >> / Rotate 0 / Type / Page >> эндобдж 42 0 объект > / Font> / ProcSet [/ PDF / Text] >> / Rotate 0 / Type / Page >> эндобдж 44 0 объект > / Font> / ProcSet [/ PDF / Text] >> / Rotate 0 / Type / Page >> эндобдж 46 0 объект > / Font> / ProcSet [/ PDF / Text] >> / Rotate 0 / Type / Page >> эндобдж 48 0 объект > / Font> / ProcSet [/ PDF / Text] >> / Rotate 0 / Type / Page >> эндобдж 50 0 объект > / Font> / ProcSet [/ PDF / Text] >> / Rotate 0 / Type / Page >> эндобдж 52 0 объект > / Font> / ProcSet [/ PDF / Text] >> / Rotate 0 / Type / Page >> эндобдж 54 0 объект > / Font> / ProcSet [/ PDF / Text] >> / Rotate 0 / Type / Page >> эндобдж 56 0 объект > / Font> / ProcSet [/ PDF / Text] >> / Rotate 0 / Type / Page >> эндобдж 58 0 объект > / ExtGState> / Font> / ProcSet [/ PDF / Text] >> / Rotate 0 / Type / Page >> эндобдж 61 0 объект > / ProcSet [/ PDF / Text / ImageC] / XObject >>> / Rotate 0 / Type / Page >> эндобдж 64 0 объект > / ProcSet [/ PDF / Text / ImageC] / XObject >>> / Rotate 0 / Type / Page >> эндобдж 67 0 объект > / ProcSet [/ PDF / Text / ImageC] / XObject >>> / Rotate 0 / Type / Page >> эндобдж 70 0 объект > / ProcSet [/ PDF / Text / ImageC] / XObject >>> / Rotate 0 / Type / Page >> эндобдж 73 0 объект > / ProcSet [/ PDF / Text / ImageC] / XObject >>> / Rotate 0 / Type / Page >> эндобдж 76 0 объект > / ProcSet [/ PDF / Text / ImageC] / XObject >>> / Rotate 0 / Type / Page >> эндобдж 183 0 объект [185 0 R] эндобдж 184 0 объект > поток Hw @) U; tc * MepN1qxlC0H, `JFK ’43 ^ $ zA4 $ * C_Q + $ A «Gi (z: V-GP * uUJY | Xv [ɷF4TRwXZ) {Vwa {י uFl 贽 vNº & 1Di» J3 ^ PzhM! Pј? Z5Z =
— — — — — — — — — — — — — — — — — — — — — — — — — — — — — — — — — — — —
|
Эксперимент по подергиванию мышц | Измерьте сокращение мышц
Проучите своих учеников урок, который они никогда не забудут, с помощью эксперимента Muscle Twitch Response.Вовлеките своих учеников и улучшите их понимание того, как нервная стимуляция контролирует сокращение мышц, в этом незабываемом эксперименте по активному обучению, в котором учащиеся проводят нервную стимуляцию на себе …
Протокол эксперимента
Студенты участвуют в практическом обучении, прикладывая электроды к себе. тела и электрически стимулировать нервы предплечья, чтобы продемонстрировать набор мышц.
Использование PowerLab 26T
×checkmark Элемент успешно добавлен в ваш запрос предложения.
Вот шаги для настройки и проведения этого классического эксперимента …
Шаг 1. Определите цели обучения
К концу занятия студенты должны уметь:
- Опишите и продемонстрируйте действие электрических раздражителей нервов. предплечья.
- Запишите и измерьте реакцию мышечных сокращений на нервную стимуляцию, и показать набор в ответ на подергивание как силу стимула. увеличивается.
Шаг 2. Настройте оборудование для записи сигналы
Используемое оборудование: PowerLab 26T
×checkmark Элемент успешно добавлен в ваш запрос предложения.
Используемое программное обеспечение: Lt: Программа обучения естествознанию / LabChart
Настройка PowerLab
- Подключите кабель Bio Amp к входу Bio Amp на PowerLab.
- Подключите кабели электродов к заземлению, Ch2 NEG и Ch2 POS на Bio. Кабель усилителя.
- Подключите заземляющий кабель к сухой заземляющей ленте. Обратите внимание, сухая земля При необходимости ремешок можно заменить одноразовым электродом.
- Подключите стимулирующий стержневой электрод к изолированному стимулятору.
вывод PowerLab:
- Подключите красный (положительный) разъем к красному выходу.
- Подключите черный (отрицательный) разъем к черному выходу.
- Убедитесь, что PowerLab подключен и включен.
- Подключите датчик импульсов ко входу 1 PowerLab.
- Поместите датчик импульсов диафрагмой вверх в верхней части лаборатории. скамейке и закрепите датчик липкой лентой.
Узнайте, как привлечь учащихся с помощью сбора реальных данных
Шаг 3: стимуляция нервов
В этой части деятельности записи не ведутся; это чисто для позволяют учащимся найти лучшую позицию для размещения стимулирующей планки электрод для предстоящего эксперимента.
Вопросы для студентов:
- Где мне нужно разместить стимулирующую планку электрод , чтобы получить лучшие дергания?
- Что происходит, когда нерв, снабжающий мышцу, стимулируется?
Для эффективной стимуляции две подушечки Стимулирующий стержневой электрод должен быть выровнен вдоль руки длина.Если цвет индикатора состояния стимула изменится с зеленого на до желтого цвета, вам нужно будет нанести больше электродной пасты на колодки.
Размещение электродов для стимуляции локтевого нерва в области локтя.
Процедура
- Нанесите небольшое количество электродной пасты на 2 металлические подушечки стимулирующий стержневой электрод.
- Поместите стимулирующий стержневой электрод на локтевой нерв добровольца на локоть. Примерное положение показано на диаграмме выше.
- Приложите давление к стимулирующему стержневому электроду, чтобы электрод не двигается.
- Установите ток на 8 миллиампер (мА) на панели изолированного стимулятора (при использовании PowerLab).
- На PowerLab переключатель изолированного стимулятора должен быть установлен в положение ВКЛ.В Изолированный стимулятор становится активным только во время записи. это все остальное время внутренне выключен.
- Выберите Пуск, и через кожа волонтера каждую секунду. Обратитесь к ним, если индикатор мигает. желтый.
- Если вы не можете добиться ответа, увеличьте ток стимулятора на 2 мА или переместите стимулирующий электрод.
- Обратите внимание на сокращения мизинца и другие пальцы.Изучите влияние небольших корректировок положения электродов и найдите положение, обеспечивающее наибольшую дергается.
- При достижении хорошего отклика выберите на устройстве «Стоп».
- Шариковой ручкой слегка наметьте на коже два маленьких крестика. с указанием положения электрода, обеспечивающего наибольшую эффективная стимуляция.
- Используйте эти отметки для изменения положения стимулирующего стержневого электрода на локоть для записи ЭМГ в следующих видах деятельности.
При подготовке к упражнению на скорость проводимости повторите выше, но на этот раз на запястье. При стимуляции запястья труднее вызвать подергивания. Студентам необходимо будет применять большие токи и большее давление. к стимулирующему стержневому электроду.
Шаг 4: размещение электродов
В этих упражнениях студенты будут стимулировать локтевой нерв на локоть или запястье и записывать мышечную активность (сложная мышца потенциал действия) и abductor digiti minimi.
Правильное размещение электродов
- Снимите все украшения с запястий добровольца.
- Оберните ремешок с сухой землей вокруг запястья добровольца. Обратите внимание, это может быть заменен одноразовым электродом. Убедитесь, что вы оставили достаточно место для размещения стимулирующего стержневого электрода на запястье.
- Отметьте 2 маленьких крестика на коже над отводящим пальцем. минимальная мышца, в которой будут размещены регистрирующие электроды, как показано на рисунке.Крестики должны находиться на расстоянии 3–4 см друг от друга.
- Слегка потрите отмеченную кожу, чтобы уменьшить ее электрическое сопротивление. Удалите все остатки.
- Получите 2 новых одноразовых электрода. Клейкие подушечки, возможно, потребуется слегка обрезаны, чтобы они подходили для рук меньшего размера.
- Присоедините кабели от канала 2 кабеля Bio Amp к электродам.
- Поместите электроды на кожу поверх крестовин, чтобы они прилегали. хорошо.Чтобы уменьшить перемещение электрода, используйте липкую ленту, чтобы прикрепить кабели к коже рядом с электродом.
- Нанесите небольшое количество электродной пасты на 2 металлические подушечки стимулирующий стержневой электрод.
- Поместите стимулирующий стержневой электрод на локтевой нерв добровольца на локоть (используя ранее сделанные отметки).
- Попросите добровольца положить руку в расслабленном положении на стол так, чтобы мизинец слегка касался пульса преобразователь.Предплечье должно опираться локтем на стол. свисает за край, чтобы обеспечить доступ к локтевому нерву.
Совет: Красная точка на обратной стороне Стимулирующий стержневой электрод указывает на положительный электрод. Расположите электрод так, чтобы положительный электрод был ближе к плечу.
Вид сверху на руку, опирающуюся на датчик импульсов.
Настройка должна выглядеть так:
Шаг 5: Выполните ответ на подергивание и прием на работу
В этой части деятельности вопрос для студентов: «Что происходит, когда электрический стимул к нерву, снабжающему мышцу прогрессивно увеличивается? «
Студенты исследуют, как нервная стимуляция управляет мышцами.
сокращение.Подергивание мышц — это однократное сокращение в ответ на
кратковременная пороговая стимуляция.
Используете PowerLab? Когда для начала записи выбрано Start (как показано ниже), PowerLab будет стимулировать, а затем записывать для
200 мс (0,2 с).
Процедура
- Попросите добровольца крепко удерживать стимулирующий стержневой электрод на месте. с его или ее свободной рукой в локте, чтобы стимулировать локтевой нерв.
- Убедитесь, что ток стимула установлен на 4 мА и Стимулятор включен.Выберите Start .
- Увеличьте стимулирующий ток на 1 мА и выберите Start .
- Продолжайте увеличивать стимулирующий ток с шагом 1 мА, выбирая Запускать каждый раз. Для большинства добровольцев порог стимул, при котором впервые наблюдается подергивание, находится в диапазоне 5–15 мА.
- Когда учащиеся впервые увидят ответ, добавьте комментарий в свою запись. отмечая используемый ток стимула.
- Продолжайте увеличивать ток стимула и выбирать Запустите , пока не достигнете максимума, который составляет 20 мА. При необходимости выберите Auto Scale в вашем программном обеспечении для анализа.
Наблюдения:
Теперь, когда ученики заметили подергивание мышц, могут ли они объяснить
что они видят с точки зрения EMG и Force?
Шаг 6: Анализ
Это возможность для студентов изучить и проанализировать записи набора мышечных сокращений.Проверьте понимание учащимися, задав такие вопросы, как:
Вопрос : Что произошло с реакцией сокращения мышц, когда сила тока на раздражитель увеличилась с 0 мА? Какой наименьший ток требовался для производства каждого из следующих элементов ?:
- сокращение (пороговый ток)
- максимальное сокращение (или «максимальный стимул» — ток, при котором реакция больше не увеличивается)
Ответ:
Ответ зависит от собственных данных учащихся.При 0 мА не может быть измеримого отклика. Обычно первое подергивание начинается примерно при 4–6 мА, а максимальная сила сокращения начинается примерно в диапазоне 12–15 мА.
Вопрос : Когда ток достигал следующих стадий, какая доля волокон в мышце сокращалась ?:
- на пороге
- при максимальном стимуле
- выше максимального стимула
Ответ:
Опять же, ответ зависит от собственных данных студентов:
При 0 мА мышечные волокна не сокращаются.
- На пороге задействованы всего несколько мышечных волокон.
- По определению, максимальный стимул наблюдается, когда сокращаются 100% задействованных волокон.
- Выше максимального стимула количество сокращающихся волокон не может увеличиваться.
Вопрос: Почему изменение силы стимула влияет на силу подергивания?
Ответ:
Более сильные стимулы стимулируют больше нервных волокон и, следовательно, задействуют больше двигательных единиц.
Этот эксперимент является частью более широкой лаборатории функции скелетных мышц, доступной на лейтенанте. Полная лаборатория продолжает изучать суммирование, столбняк и скорость проводимости в интерактивном и захватывающем виде.
Lt — это наша платформа для онлайн-обучения, которая экономит время преподавателей и привлекает студентов, объединяя практический сбор данных с помощью PowerLab с готовыми лабораторными работами и уроками для преподавания наук о жизни. Узнать больше…
Примечание: Эти действия включают применение
электрические токи к мышцам через электроды, расположенные на
кожа.Люди, у которых есть кардиостимуляторы, или которые страдают от
неврологические или сердечные расстройства не должны добровольно участвовать в этих
виды деятельности.
Сокращение мышц и ощущения (например, покалывание или кратковременный дискомфорт) могут
быть связаны со стимуляцией нервов. Если волонтер чувствует себя серьезным
дискомфорт во время занятий, прекратите упражнение
немедленно.
Преподавайте своим ученикам урок, который они никогда не забудут, с помощью совершенно новой PowerLab серии T.
PowerLab 26T
×checkmark Элемент успешно добавлен в ваш запрос предложения.
PowerLab серии T — это высококачественный и надежный регистратор данных, разработанный специально для обучения. Сделайте науку увлекательной с помощью нашего решения plug ‘n’ play, которое подключается к широкому спектру датчиков и удобному для пользователя программному обеспечению.У нас даже есть более 350 заранее написанных уроков, которые понравятся ученикам и которые сэкономят вам время и силы!
Вы заинтересованы в использовании Lt для проведения вашего курса? Попробуйте Lt сейчас — бесплатно!
Lt включает более 500 готовых к использованию, полностью настраиваемых уроков и лабораторий для преподавания физиологии, биологии, химии, медицины и курсов медсестер. Нажмите ниже, чтобы сразу просмотреть избранные уроки Lt, или подпишитесь на 90-дневную бесплатную пробную версию. Посмотреть все 11 коллекций контента »
Различные типы мышц в человеческом теле — Мышечная система — Edexcel — GCSE Physical Education Revision — Edexcel
В теле есть три типа мышц:
- гладкие мышцы — находятся во внутренних органах и кровеносных сосудах — это непроизвольно
- сердечная мышца — находится только в сердце — это непроизвольно
- скелетная мышца — прикреплена к скелету — это произвольно
Непроизвольные мышцы не находятся под нашим сознательным контролем, что означает, что мы не можем заставить их сжиматься, когда мы думаем об этом.
Произвольные мышцы находятся под нашим сознательным контролем, поэтому мы можем двигать этими мышцами, когда захотим.
- Вопрос
Какие типы мышечных волокон не находятся под сознательным контролем?
- Показать ответ
Сердечная мышца и гладкая мышца.
Типы мышечных волокон
Есть три различных типа скелетных мышц:
Каждый тип мышечного волокна имеет разные характеристики, которые показаны в таблице:
| Тип I | Тип IIa | Тип IIx | |
|---|---|---|---|
| Скорость сокращения | Медленно | Быстро | Очень быстро |
| Создаваемое усилие | Низкое | Среднее | Высокое | 2
| Сопротивление Низкий | |||
| Цвет | Красный | Розовый | Белый |
Велосипедистам на выносливость требуется высокий процент волокон типа I, чтобы их мышцы могли работать во время гонки, не уставая.Эти мышцы имеют красный цвет из-за большого количества капилляров, которые транспортируют насыщенную кислородом кровь к работающим мышцам.
Спринтеру потребуется высокий процент волокон типа IIx, чтобы его мышцы могли сокращаться очень быстро и создавать большую силу, чтобы они могли очень быстро бегать на дистанцию 100 м. Однако эти мышцы очень быстро устают, поэтому спринтер не сможет долго бегать с такой скоростью.
- Вопрос
Объясните, какой тип мышечных волокон требуется бегуну на длинные дистанции, чтобы добиться успеха в своем виде спорта.
- Показать ответ
Им потребуется высокий процент мышечных волокон типа I, поскольку они сокращаются медленно с небольшой силой, что обеспечит беговые движения в медленном темпе. Тем не менее, они могут продолжать сокращаться в течение длительного времени, не испытывая усталости, необходимой для бега на большие дистанции.
Анализ динамики сателлитных клеток человека на культивируемых миофибриллах скелетных мышц взрослых | Скелетная мышца
Бенцингер С.Ф., Ван YX, Дюмон Н.А., Рудницки М.А.Клеточная динамика в нише мышечных сателлитных клеток. EMBO Rep. 2013; 14 (12): 1062–72.
CAS PubMed PubMed Central Google Scholar
Фейге П., Брун С.Е., Ритсо М., Рудницки М.А. Ориентация мышечных стволовых клеток на регенерацию при гомеостазе, старении и болезнях. Стволовая клетка. 2018; 23 (5): 653–64.
CAS PubMed PubMed Central Google Scholar
Карлсон МЭ, Конбой ИМ. Потеря способности стволовых клеток к регенерации в старых нишах. Ячейка старения. 2007. 6 (3): 371–82.
CAS PubMed PubMed Central Google Scholar
Шефер Г., Ван де Марк Д. П., Ричардсон Дж. Б., Яблонка-Реувени З. Размер пула сателлитных клеток имеет значение: определение миогенной силы стареющих скелетных мышц. Dev Biol. 2006. 294 (1): 50–66.
CAS PubMed PubMed Central Google Scholar
Конбой И.М., Конбой М.Дж., Смайт GM, Рандо Т.А. Notch-опосредованное восстановление регенеративного потенциала старых мышц. Наука. 2003. 302 (5650): 1575–7.
CAS PubMed Google Scholar
Conboy IM, Rando TA. Регуляция передачи сигналов Notch контролирует активацию сателлитных клеток и детерминацию клеточной судьбы в постнатальном миогенезе. Dev Cell. 2002. 3 (3): 397–409.
CAS PubMed Google Scholar
Брак А.С., Конбой М.Дж., Рой С., Ли М., Куо С.Дж., Келлер С. и др. Повышенная передача сигналов Wnt во время старения изменяет судьбу мышечных стволовых клеток и увеличивает фиброз. Наука. 2007. 317 (5839): 807–10.
CAS PubMed Google Scholar
Brack AS, Rando TA. Внутренние изменения и внешние влияния на функцию миогенных стволовых клеток во время старения. Stem Cell Rev.2007; 3 (3): 226–37.
CAS PubMed Google Scholar
Бернет Дж. Д., Доулс Дж. Д., Холл Дж. К., Келли-Танака К., Картер Т. А., Олвин ББ. Передача сигналов P38 MAPK лежит в основе автономной клеточной потери самообновления стволовых клеток в старых скелетных мышцах. Nat Med. 2014; 20 (3): 265–71.
CAS PubMed PubMed Central Google Scholar
Price FD, von Maltzahn J, Bentzinger CF, Dumont NA, Yin H, Chang NC, et al. Ингибирование передачи сигналов JAK / STAT стимулирует функцию взрослых сателлитных клеток. Nat Med.2014. 20 (10): 1174–81.
CAS PubMed PubMed Central Google Scholar
Chakkalakal JV, Jones KM, Basson MA, Brack AS. Старая ниша нарушает покой мышечных стволовых клеток. Природа. 2012. 490 (7420): 355–60.
CAS PubMed PubMed Central Google Scholar
Розо М, Ли Л, Вентилятор С-М. Нацеливание на передачу сигналов β1-интегрина усиливает регенерацию в старых и дистрофических мышцах у мышей.Nat Med. 2016; 22 (8): 889–96.
CAS PubMed PubMed Central Google Scholar
Cosgrove BD, Gilbert PM, Porpiglia E., Mourkioti F, Lee SP, Corbel SY, et al. Омоложение стареющей популяции мышечных стволовых клеток восстанавливает силу травмированных старых мышц. Nat Med. 2014. 20 (3): 255–64.
CAS PubMed PubMed Central Google Scholar
Hay M, Thomas DW, Craighead JL, Economides C, Rosenthal J.Показатели успешности клинических разработок исследуемых препаратов. Nat Biotechnol. 2014; 32 (1): 40–51.
CAS PubMed Google Scholar
Van Norman GA. Фаза II испытаний по разработке лекарств и адаптивному дизайну испытаний. JACC Basic Transl Sci. 2019; 4 (3): 428–37.
PubMed PubMed Central Google Scholar
van der Worp HB, Howells DW, Sena ES, Porritt MJ, Rewell S, O’Collins V, et al.Могут ли животные модели болезней достоверно использоваться в исследованиях на людях? PLoS Med. 2010; 7 (3).
Hackam DG, Redelmeier DA. Перевод научных данных с животных на людей. ДЖАМА. 2006. 296 (14): 1727–32.
Google Scholar
Пасут А, Олейник П., Рудницкий М.А. Выделение мышечных стволовых клеток с помощью цитометрии сортировки клеток с активацией флуоресценции. В: DiMario JX, редактор. Миогенез: методы и протоколы. Тотова: Humana Press; 2012 г.п. 53–64.
Google Scholar
Коллинз С.А., Олсен И., Заммит П.С., Хеслоп Л., Петри А., Партридж Т.А. и др. Функция стволовых клеток, самообновление и поведенческая неоднородность клеток из ниши взрослых мышечных сателлитных клеток. Клетка. 2005. 122 (2): 289–301.
CAS PubMed Google Scholar
Файге П., Рудницкий М.А. Выделение сателлитных клеток и трансплантация мышам для отслеживания клонов в мышцах.Nat Protoc. 2020; 15 (3): 1082–97.
CAS PubMed Google Scholar
Шефер Г., Яблонька-Реувени З. Выделение и культивирование миофибрилл скелетных мышц как средство анализа сателлитных клеток. В: Helgason CD, Miller CL, редакторы. Основные протоколы клеточных культур. Тотова: Humana Press; 2005. с. 281–304.
Google Scholar
Kuang S, Le Grand F, Rudnicki MA.Асимметричное самообновление и обязательство сателлитных стволовых клеток в мышцах. Клетка. 2007. 129 (5): 999–1010.
CAS PubMed PubMed Central Google Scholar
Ван YX, Фейге П., Брун С.Е., Хекматнежад Б., Дюмон Н.А., Рено Дж.М. и др. Передача сигналов EGFR-Aurka устраняет дефекты полярности и регенерации в мышечных стволовых клетках с дефицитом дистрофина за счет увеличения асимметричных делений. Стволовая клетка. 2019; 24 (3): 419–432.e6.
PubMed PubMed Central Google Scholar
Инь Х, Пасут А., Сулеймани В.Д., Бенцингер С.Ф., Антоун Г., Торн С. и др. MicroRNA-133 контролирует определение коричневого жира в сателлитных клетках скелетных мышц, воздействуя на Prdm16. Cell Metab. 2013. 17 (2): 210–24.
CAS PubMed PubMed Central Google Scholar
Сил П., Бьорк Б., Ян В., Каджимура С., Чин С., Куанг С. и др. PRDM16 управляет переключением между коричневым жиром и скелетными мышцами. Природа. 2008. 454 (7207): 961–7.
CAS PubMed PubMed Central Google Scholar
Seale P, Sabourin LA, Girgis-Gabardo A, Mansouri A, Gruss P, Rudnicki MA. Pax7 требуется для спецификации миогенных спутниковых клеток. Клетка. 2000. 102 (6): 777–86.
CAS PubMed Google Scholar
Bentzinger CF, von Maltzahn J, Dumont NA, Stark DA, Wang YX, Nhan K, et al. Wnt7a стимулирует подвижность и приживление миогенных стволовых клеток, что приводит к увеличению мышечной силы. J Cell Biol. 2014; 205 (1): 97–111.
CAS PubMed PubMed Central Google Scholar
Le Grand F, Jones AE, Seale V, Scimè A, Rudnicki MA. Wnt7a активирует путь плоской клеточной полярности, чтобы управлять симметричной экспансией сателлитных стволовых клеток. Стволовая клетка. 2009. 4 (6): 535–47.
PubMed PubMed Central Google Scholar
Джадсон Р.Н., Кварта М., Аудхофф М.Дж., Солиман Х., Йи Л., Чанг С.К. и др.Ингибирование метилтрансферазы Setd7 позволяет увеличивать in vitro миогенные стволовые клетки с улучшенным терапевтическим потенциалом. Стволовая клетка. 2018; 22 (2): 177–190.e7.
CAS PubMed PubMed Central Google Scholar
фон Мальцан Дж., Рено Дж. М., Паризе Г., Рудницки М.А. Лечение Wnt7a улучшает мышечную дистрофию. Proc Natl Acad Sci. 2012; 109 (50): 20614–9.
Google Scholar
Каспер AM, Тернер округ Колумбия, Мартин NRW, Шарплс AP. Имитация упражнений в трехмерных биоинженерных скелетных мышцах для исследования клеточных и молекулярных механизмов физиологической адаптации. J. Cell Physiol. 2018. 233 (3): 1985–98.
CAS PubMed Google Scholar
Афшар Бакушли М., Липпманн Э.С., Малкахи Б., Айер Н., Нгуен С.Т., Тунг К. и др. 3D-модель культуры иннервируемых скелетных мышц человека позволяет исследовать нервно-мышечные соединения взрослых.eLife. 8.
Li EW, McKee-Muir OC, Gilbert PM. Глава пятая — клеточная биомеханика в регенерации скелетных мышц. В: Сассун Д., редактор. Актуальные темы биологии развития: Academic Press; 2018. с. 125–76.
Латиль М., Рошето П., Шатр Л., Санулли С., Мемет С., Риккетти М. и др. Стволовые клетки скелетных мышц после смерти принимают состояние покоящихся клеток и сохраняют регенеративную способность. Nat Commun. 2012; 3 (1): 903.
PubMed Google Scholar
Yucel N, Chang AC, Day JW, Rosenthal N, Blau HM. Очеловечивание модели МДД на мышах MDX: короткое и короткое. NPJ Regen Med. 2018; 3.
Шелтон М., Кочарян А., Лю Дж., Скерянц И.С., Стэнфорд В.Л. Устойчивое создание и распространение предшественников скелетных мышц и миоцитов из плюрипотентных стволовых клеток человека. Методы. 2016; 101: 73–84.
CAS PubMed Google Scholar
Пига Д., Салани С., Магри Ф., Бруса Р., Маури Е., Коми Г.П. и др.Модели индуцированных человеком плюрипотентных стволовых клеток для изучения и лечения мышечных дистрофий Дюшенна и Беккера. Ther Adv Neurol Disord. 2019; 12.
Гилберт П.М., Хавенстрайт К.Л., Магнуссон КЕГ, Сакко А., Леонарди Н.А., Крафт П. и др. Эластичность субстрата регулирует самообновление стволовых клеток скелетных мышц в культуре. Наука. 2010. 329 (5995): 1078–81.
CAS PubMed PubMed Central Google Scholar
Петроф Б.Дж., Шрагер Дж.Б., Стедман Х.Х., Келли А.М., Суини Х.Л. Дистрофин защищает сарколемму от стрессов, возникающих при сокращении мышц. Proc Natl Acad Sci. 1993. 90 (8): 3710–4.
CAS PubMed Google Scholar
Dumont NA, Wang YX, von Maltzahn J, Pasut A, Bentzinger CF, Brun CE, et al. Экспрессия дистрофина в мышечных стволовых клетках регулирует их полярность и асимметричное деление. Nat Med. 2015; 21 (12): 1455–63.
CAS PubMed PubMed Central Google Scholar
Александр М.С., Розкалне А., Коллетта А., Спинаццола Дж. М., Джонсон С., Рахимов Ф. и др. CD82 является маркером предполагаемого выделения сателлитных клеток мышц человека и связан с мышечными дистрофиями. Стволовая клетка. 2016; 19 (6): 800–7.
CAS PubMed PubMed Central Google Scholar
Гуревич Д.Б., Нгуен П.Д., Сигель А.Л., Эрлих О.В., Зоннтаг С., Фан Дж.М.Н. и др.Асимметричное деление клональных мышечных стволовых клеток координирует регенерацию мышц in vivo. Наука. 2016; 353 (6295).
Gissel H. Роль Ca2 + в повреждении мышечных клеток. Ann N Y Acad Sci. 2006; 1066 (1): 166–80.
Google Scholar
Тедеско Ф.С., Мойл Л.А., Пердигеро Э. Мышечные интерстициальные клетки: краткое практическое руководство по популяциям несателлитных клеток в скелетных мышцах. В: Perdiguero E, Cornelison D, редакторы.Стволовые клетки мышц: методы и протоколы. Нью-Йорк: Спрингер; 2017. с. 129–47.
Google Scholar
Giordani L, He GJ, Negroni E, Sakai H, Law JYC, Siu MM, et al. Высокомерная одноклеточная картография выявляет новые популяции клеток, резидентных в скелетных мышцах. Mol Cell. 2019; 74 (3): 609–621.e6.
CAS PubMed Google Scholar
Smith LR, Barton ER.SMASH — полуавтоматический анализ мышц с использованием гистологической сегментации: приложение MATLAB. Скелетная мышца. 2014; 4:21.
PubMed PubMed Central Google Scholar
Аагард П., Андерсен Дж. Л., Дайре-Поульсен П., Лефферс А. М., Вагнер А., Магнуссон С. П. и др. Механизм увеличения сократительной силы перистых мышц человека в ответ на силовую тренировку: изменения в архитектуре мышц. J Physiol. 2001; 534 (Pt 2): 613–23.
CAS PubMed PubMed Central Google Scholar
Драгиева П., Захариева М., Кожухаров Ю., Марков К., Стоянов Г.С. Малая поясничная мышца: морфометрическое исследование трупа. Cureus. 2018; 10 (4).
Hanson P, Magnusson SP, Sorensen H, Simonsen EB. Анатомические различия поясничных мышц у молодых черных и белых мужчин. J Anat. 1999. 194 (Pt 2): 303–7.
PubMed PubMed Central Google Scholar
Харрисон, Британская Колумбия, Аллен Д.Л., Лейнванд, Лос-Анджелес. IIb или не IIb? Регуляция экспрессии гена тяжелой цепи миозина у мышей и мужчин. Скелетная мышца. 2011; 1: 5.
CAS PubMed PubMed Central Google Scholar
Фрай С.С., Ноерен Б., Мула Дж., Убеле М.Ф., Вестгейт П.М., Керн П.А. и др. Специфический для типа волокна ответ сателлитных клеток на аэробную тренировку у взрослых, ведущих малоподвижный образ жизни. J Physiol. 2014; 592 (Pt 12): 2625–35.
CAS PubMed PubMed Central Google Scholar
Machado L, Esteves de Lima J, Fabre O, Proux C, Legendre R, Szegedi A, et al. Фиксация in situ переопределяет покой и раннюю активацию стволовых клеток скелетных мышц. Cell Rep., 2017; 21 (7): 1982–93.
CAS PubMed Google Scholar
van Velthoven CTJ, de Morree A, Egner IM, Brett JO, Rando TA. Транскрипционное профилирование покоящихся мышечных стволовых клеток in vivo.
