
Сайт учителей биологии МБОУ Лицей № 2 города Воронежа
Строение и функции скелетных мышц
Мышцы обеспечивают передвижение человека, работу отдельных частей его тела и многих внутренних органов (сердца, легких, желудка и др.). Мышцы состоят из мышечной ткани. Различают гладкие и скелетные мышцы.
Гладкие мышцы образуют стенки кровеносных сосудов, дыхательных путей, желудка, кишечника. Сокращаются гладкие мышцы медленно и могут долго находиться в таком состоянии. Они принимают участие в работе внутренних органов, и независимо от нашей воли управляются вегетативным отделом нервной системы и гуморально.
К скелетным мышцам относятся поперечно-полосатые мышцы головы, туловища и конечностей. Скелетные мышцы сокращаются быстро. Их работа обеспечивает произвольные движения.
Строение мышц. Каждая мышца является отдельным органом, т. е. целостным образованием, имеющим свою определенную, присущую только ему форму, строение, функцию, развитие и положение в организме.
В мышце различают активно сокращающуюся часть — брюшко — и пассивную часть, при помощи которой она прикрепляется к костям, — сухожилие. Снаружи мышца покрыта соединительнотканной оболочкой.
Сухожилие — это часть мышцы; состоит из толстых соединительнотканных волокон. Сухожилия мало растяжимы и прочны; с помощью сухожилий мышцы прикрепляются к кости и при сокращении приводит её в движение. На одном конце сухожильные волокна тесно сплетаются с мышечными, на другом — вплетены в надкостницу. Форма сухожилий разнообразна: у длинных мышц — цилиндрическая, у широких — в виде пластин.
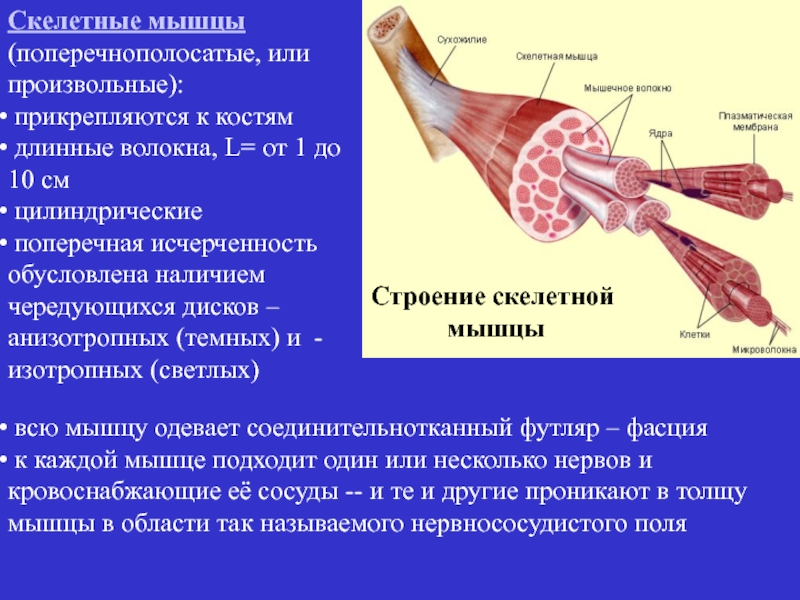
Скелетные мышцы состоят из пучков поперечнополосатых волокон. Мышечные пучки объединяются соединительнотканной оболочкой, которая на концах мышцы переходит в сухожильные части мышцы.
Каждая мышца связана нервами с центральной нервной системой; кроме того, в мышцу проникают кровеносные сосуды. Нервы и сосуды разветвляются в толще мышцы соответственно мышечным пучкам.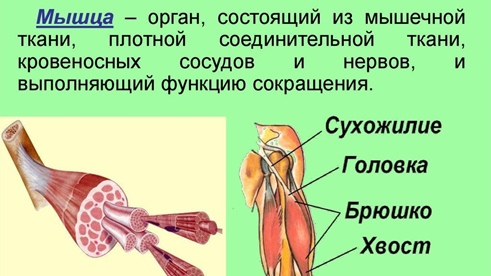 Мышечные пучки состоят из мышечных волокон.
Мышечные пучки состоят из мышечных волокон.Мышечное волокно — минимальный структурный элемент мышцы. Снаружи оно окружено тонкой оболочкой из соединительной ткани, а внутри содержит многочисленные тонкие сократительные нити — миофибриллы и множество ядер. Каждая миофибрилла состоит из тончайших белковых нитей и способна сокращаться.
Миофибриллы — цилиндрические нити толщиной 1 — 2 мкм, идущие вдоль от одного конца мышечного волокна до другого.
Миофибрилла — это сократимый элемент мышечной клетки, она способна сокращаться в присутствии АТФ. Миофибриллы состоят из сократительных белков
актина и миозина.
Мышца может сократиться только тогда, когда к ней придет сигнал от центральной нервной системы. Если нерв поврежден, мышца сокращаться не будет.
Для нормальной работы мышц необходимы питательные вещества и кислород, поступающие с кровью, так как энергия мышечного сокращения связана с биологическим окислением органических веществ мышечного волокна.
Образующиеся при работе мышц продукты распада уносятся кровью. Вот почему ухудшение кровоснабжения нарушает деятельность мышц и нередко вызывает болевые ощущения.
Основное свойство мышечной ткани – сократимость. На этом свойстве основана работа мышц. В возбужденном состоянии мышца укорачивается и утолщается – сокращается, затем расслабляется и принимает прежние размеры. При сокращении мышцы производят работу по передвижению тела, конечностей или удерживанию груза.
Функции и строение скелетных мышц
Биология. 8 класс. Мищук
Вспомните строение мышечной ткани. Какие виды мышечных тканей вам известны? Какое строение имеют мышечные клетки и какие функции они выполняют?
Функции скелетных мышц. Вам известно, что в организме человека, как и у всех других позвоночных, для осуществления различных движений имеется два вида мышечной ткани: гладкая и поперечнополосатая (ил. 12). Среди поперечнополосатой мышечной ткани различают скелетную и сердечную. В соответствии с разновидностями мышечной ткани различают гладкие, скелетные и сердечную мышцы. Именно скелетные мышцы являются активной частью опорно-двигательной системы организма. Они обеспечивают равновесие и движение тела в пространстве, глотательные и дыхательные движения, перемещение одних частей тела относительно других.
Среди поперечнополосатой мышечной ткани различают скелетную и сердечную. В соответствии с разновидностями мышечной ткани различают гладкие, скелетные и сердечную мышцы. Именно скелетные мышцы являются активной частью опорно-двигательной системы организма. Они обеспечивают равновесие и движение тела в пространстве, глотательные и дыхательные движения, перемещение одних частей тела относительно других.
В скелетных мышцах интенсивно происходит обмен веществ с преобразованием химической энергии органических веществ в механическую и тепловую. Около 30 % образованной энергии используется для механической работы мышцы, остальная энергия преобразуется в тепло. Тепло, образующееся в скелетных мышцах, участвует в процессах терморегуляции.
Строение скелетной мышцы. В мышце (ил. 87 а) различается активная часть — брюшко, или тело, и пассивная часть — сухожилия. Брюшко мышцы образовано пучками поперечнополосатых мышечных волокон. Пучки связаны между собой рыхлой соединительной тканью. В ней проходят кровеносные сосуды и нервы.
Пучки связаны между собой рыхлой соединительной тканью. В ней проходят кровеносные сосуды и нервы.
Снаружи и пучок, и мышца полностью покрыты тонкой оболочкой. Количество пучков в мышце зависит от ее функциональных особенностей. В мышце проходят нервы, кровеносные и лимфатические сосуды (ил. 87 б).
Ил. 87. Строение скелетной мышцы: а — внешнее строение; б — пучки мышечных волокон; в — мышечное волокно
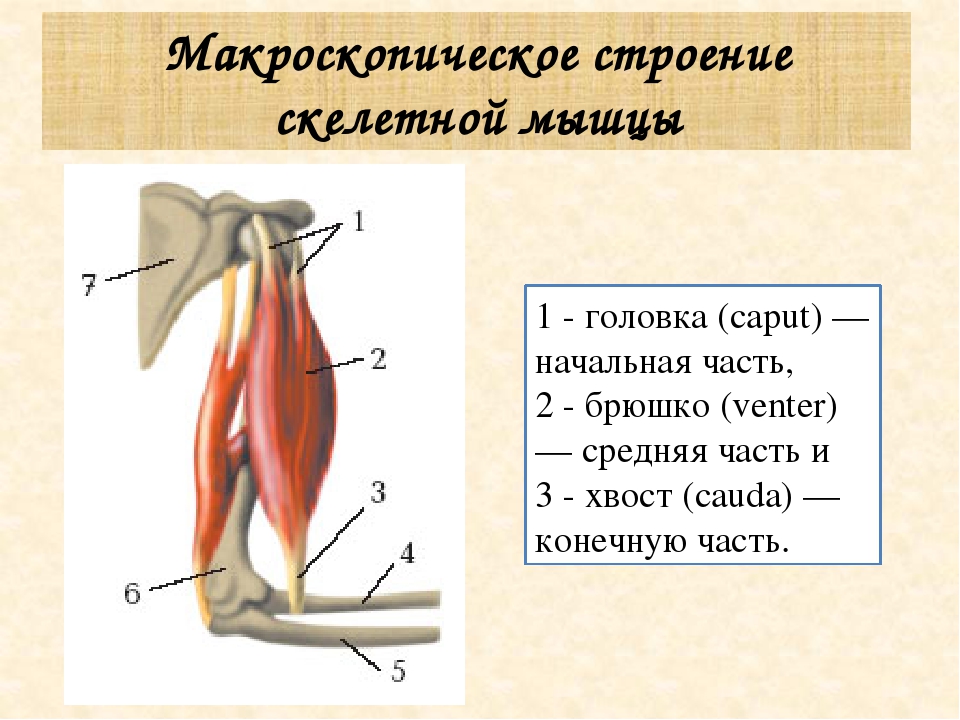
Часть мышцы, остающаяся неподвижной при ее сокращении, называется головкой, а подвижный конец — хвостом. Мышцы, имеющие одну головку, относятся к простым. Существуют мышцы с двумя (двуглавая мышца), тремя (трехглавая мышца) и даже четырьмя (четырехглавая мышца) головками. От головки и хвоста отходят сухожилия, с помощью которых мышца крепится к костям (иногда к коже, суставным сумкам, хрящам и т. п.) (ил. 87 а). Сухожилия почти не растягиваются, но они очень прочные и выдерживают большие нагрузки. Прочность сухожилиям придает плотная соединительная ткань.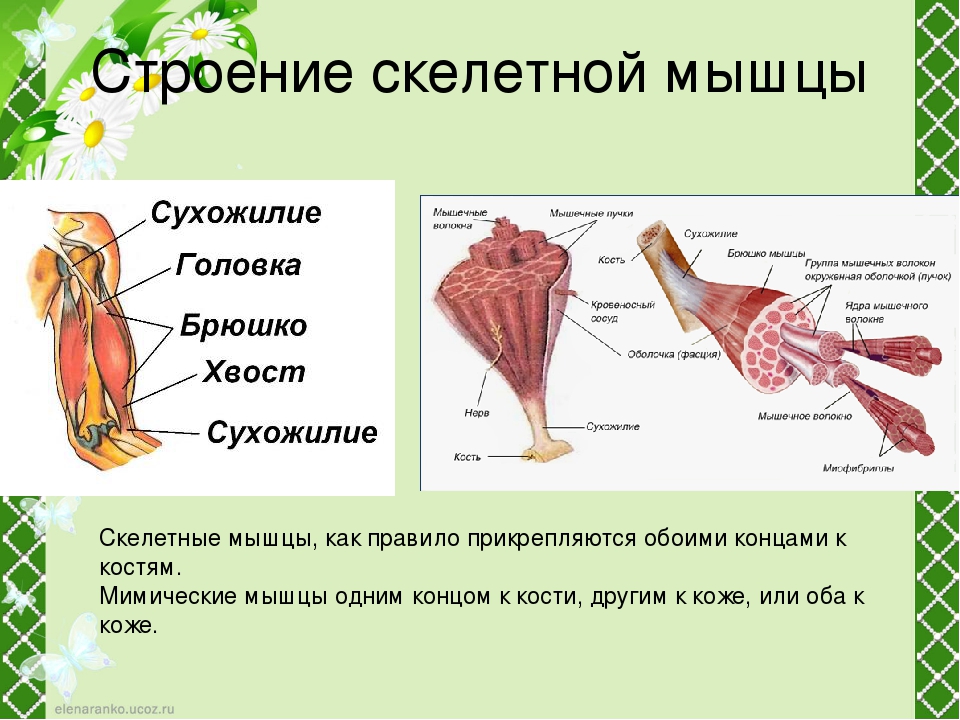 Вспомните особенности ее строения (ил. 9 б, с. 12).
Вспомните особенности ее строения (ил. 9 б, с. 12).
Мышцы в основном имеют красно-бурый цвет, а сухожилия — белый. Сухожилия снабжаются кровью несколько меньше, чем мышечное брюшко, они тоньше, чем мышцы, и очень прочные. Например, пяточное сухожилие выдерживает нагрузку около 400 кг, а сухожилие четырехглавой мышцы бедра — около 600 кг.
Структурной и функциональной единицей скелетной мышцы является мышечное волокно (ил. 87 в). Снаружи оно покрыто оболочкой. В цитоплазме мышечного волокна содержится много ядер и органелл, обеспечивающих процессы жизнедеятельности мышцы. Здесь содержатся специальные органеллы — миофибриллы, обеспечивающие мышцам выполнение сократительной функции. Миофибрилла — это тонкое волоконце, в состав которого входят белки — актин и миозин. Актин образует тонкое сократительное волоконце, а миозин — толстое сократительное волоконце. На ил. 88 схематически представлен сократительный элемент миофибриллы.
Актин и миозин имеют различные физико-химические характеристики, что обусловливает различие в их цвете.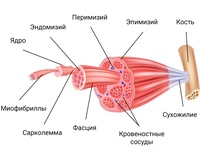
Ил. 88. Сократительной элемент миофибриллы
Лабораторное исследование
Тема: Микроскопическое строение скелетной мышечной ткани.
Цель: исследовать строение скелетной мышечной ткани, установить связь между ее строением и функциями, научиться распознавать скелетную мышечную ткань.
Оборудование: микроскопы, микропрепарат скелетной мышечной ткани.
Ход исследования
1. Подготовьте микроскоп к работе.
2. Рассмотрите поочередно при малом, а затем при большом увеличении микроскопа постоянный микропрепарат скелетной мышечной ткани. Найдите мышечное волокно — структурную единицу скелетной мышечной ткани. Исследуйте количество ядер в нем и исчерченность волокна.
3. Сопоставьте увиденное с ил.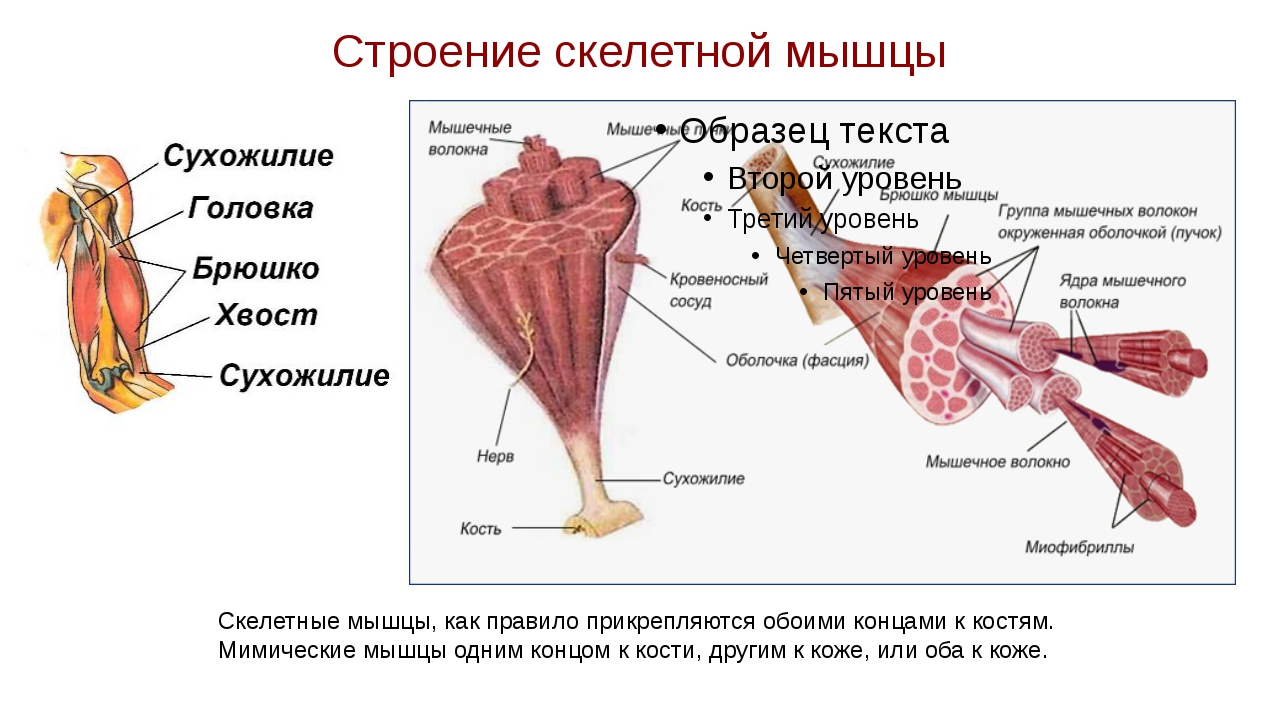 12 б (§ 2) учебника.
12 б (§ 2) учебника.
4. Схематически изобразите увиденную под микроскопом мышечную ткань.
5. Сделайте вывод: чем обусловлены особенности строения скелетной мышечной ткани?
Скелетная мышца. Скелетная мышечная ткань. Мышечное волокно. Миофибриллы. Актин. Миозин. Сухожилие
Цвет мышц зависит от количества мышечного пигмента миоглобина, содержащегося в цитоплазме мышечных волокон. Миоглобин — белок, по химическому составу и свойствам близкий к гемоглобину крови. Он легко связывает кислород, образуя соединение оксимиоглобин. Поэтому миоглобин является источником кислорода для мышц. В зависимости от содержания миоглобина различаются белые и красные мышечные волокна. Белые мышечные волокна быстро сокращаются и обеспечивают активное движение, а красные сокращаются медленно и служат для поддержания равновесия.
1. Из какой ткани образована скелетная мышца? 2. Какие функции выполняют скелетные мышцы? 3. Опишите строение скелетной мышцы. 4. Что такое сухожилие? Каково его строение? 5. Опишите строение мышечного волокна. 6. Что такое миофибрилла? Каковы особенности ее строения? 7. Почему скелетные мышцы называют поперечнополосатыми? 8. Обоснуйте значение скелетных мышц в организме человека. 9. В чем проявляется взаимосвязь строения и функций скелетной мышцы? 10. Сравните поперечнополосатую скелетную мышечную ткань с поперечнополосатой сердечной. 11. В чем проявляется взаимосвязь строения и функций мышечного волокна? 12. Проведите исследование, которое докажет или опровергнет утверждение о том, что к работающим мышцам поступает больше крови, чем к неработающим. В одну руку возьмите эспандер или теннисный мячик и сжимайте его в течение 1-3 мин. Другую руку опустите вниз. После окончания работы сравните руки по цвету. Сформулируйте вывод.
Что такое сухожилие? Каково его строение? 5. Опишите строение мышечного волокна. 6. Что такое миофибрилла? Каковы особенности ее строения? 7. Почему скелетные мышцы называют поперечнополосатыми? 8. Обоснуйте значение скелетных мышц в организме человека. 9. В чем проявляется взаимосвязь строения и функций скелетной мышцы? 10. Сравните поперечнополосатую скелетную мышечную ткань с поперечнополосатой сердечной. 11. В чем проявляется взаимосвязь строения и функций мышечного волокна? 12. Проведите исследование, которое докажет или опровергнет утверждение о том, что к работающим мышцам поступает больше крови, чем к неработающим. В одну руку возьмите эспандер или теннисный мячик и сжимайте его в течение 1-3 мин. Другую руку опустите вниз. После окончания работы сравните руки по цвету. Сформулируйте вывод.
Скелетные мышцы, их строение и функции
Скелетные мышцы, их строение и функции
Функции мышц. Мышцы — это органы тела, состоящие
из мышечной ткани, способной сокращаться под влиянием нервных
импульсов. Они являются активным элементом опорно-двигательной
системы, так как обеспечивают разнообразные движения при
перемещении человека в пространстве, сохранение равновесия,
дыхательные движения, сокращения стенок внутренних органов,
голосообразование и др.
Мышцы — это органы тела, состоящие
из мышечной ткани, способной сокращаться под влиянием нервных
импульсов. Они являются активным элементом опорно-двигательной
системы, так как обеспечивают разнообразные движения при
перемещении человека в пространстве, сохранение равновесия,
дыхательные движения, сокращения стенок внутренних органов,
голосообразование и др.
Рис. 12.8. Мышечная система человека:
1 — мышцы лица; 2 — мышцышеи; 3 —
дельтовидная мышца; 4 — большая грудная мышца; 5
— двуглавая мышца плеча; 6 —- наружная косая мышца
живота; 7 — прямая мышца живота; 8 — мышцы предплечья;
9 — мышцы кисти; 10 — четырехглавая мышца бедра; Ц —
мышцы голени; 12 — икроножная мышца; 13 —
двуглавая мышца бедра; 14 — большая ягодич-ная мышца;
15 — широчайшая мышца спины; 16 — трехглавая
мышца плеча; 17 — трапециевидная мышца.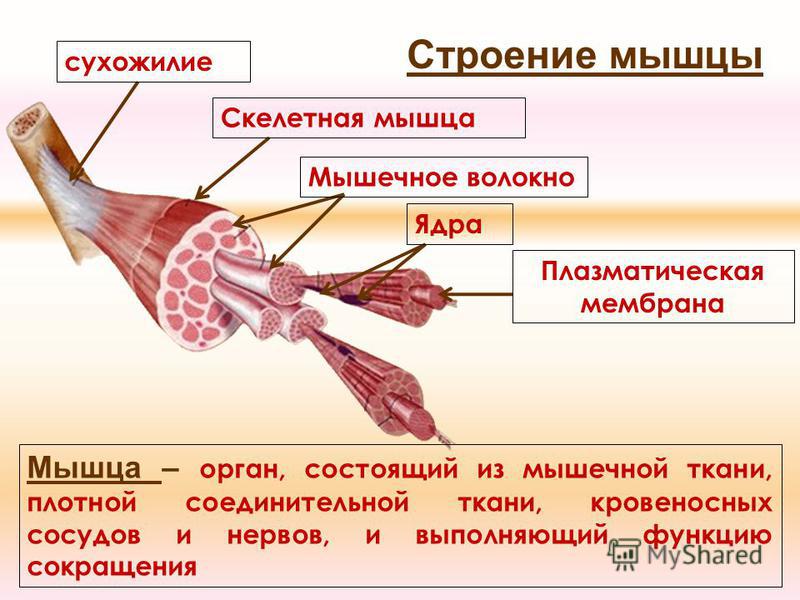
Соединение со скелетом дало основание называть их скелетной мускулатурой (рис. 12.8). Общее число мышц около 600, а доля их от массы тела человека оставляет в среднем около 30%.
Строение мышцы. Мышца состоит из пучков поперечнополосатых мышечных волокон, соединенных рыхлой соединительной тканью в пучки первого порядка. Они, в свою очередь, объединяются в пучки второго порядка и т. д. В итоге мышечные пучки всех порядков объединяются соединительной оболочкой, образуя мышечное брюшко. Соединительнотканные прослойки, имеющиеся между мышечными пучками по концам брюшка, переходят в сухожильную часть мышцы, крепящейся к кости. Во время сокращения происходит укорочение мышечного брюшка и сближение ее концов. При этом сократившаяся мышца с помощью сухожилия тянет за собой кость, которая выполняет роль рычага. Так совершаются разнообразные движения.
Так совершаются разнообразные движения.
Каждая мышцз является целостным (отдельным) органом, имеющим
определенную форму, строение и функцию, развитие и положение в
организме. Мышцы обильно снабжены кровеносными сосудами и
нервами. В каждом движении принимают участие несколько мышц.
Мышцы, действующие совместно в одном направлении и вызывающие
сходный эффект, называются синергистами, а совершающие
противоположно направленные движения —антагонистами.
Например, сгибателем локтевого сустава является двуглавая
мышца плеча (бицепс), а разгибателем — трехглавая (трицепс)-
Сокращение мышц-сгибателей локтевого сустава сопровождается
расслаблением мышц-разгибателей. Однако при постоянной
нагрузке на сустав (например, при удержании гири в
горизонтально вытянутой руке) мышцы-сгибатели и разгибатели
локтевого сустава действуют уже не как антагонисты, а как
синергисты.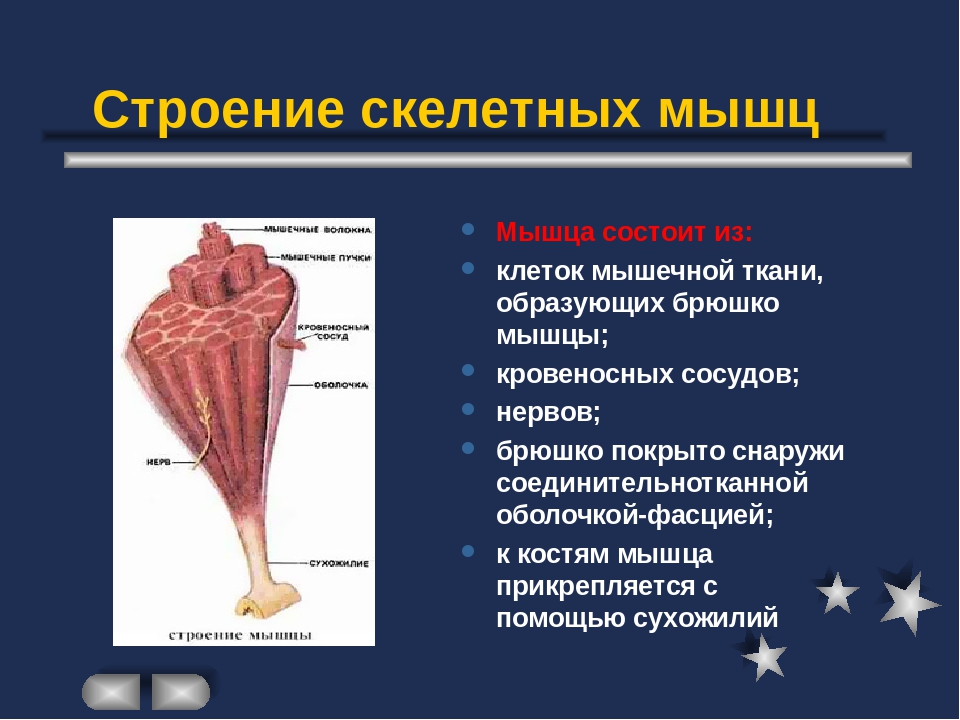 Таким образом, действия мышц нельзя сводить к
выполнению только одной функции, так как они
многофункциональны. Поскольку в каждом движении участвуют
мышцы как одной, так и другой группы, наши движения точны и
плавны.
Таким образом, действия мышц нельзя сводить к
выполнению только одной функции, так как они
многофункциональны. Поскольку в каждом движении участвуют
мышцы как одной, так и другой группы, наши движения точны и
плавны.
По характеру выполняемых основных движений и по действию на сустав различают следующие виды мышц: сгибатели и разгибатели, приводящие и отводящие, вращающие, приподнимающие и опускающие и др. Выделяют также мимические, жевательные и дыхательные мышцы.
Нервная регуляция деятельности мышц. В большинстве
движений участвует множество мышц, причем сокращение и
расслабление различных групп мышц происходит в определенном
порядке и с определенной силой. Такая согласованность движений
называется координацией движений.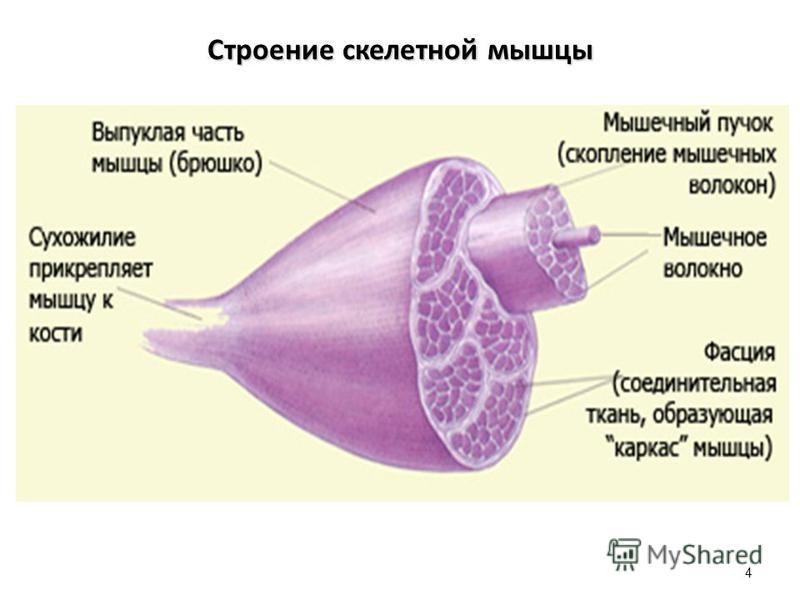 Она осуществляется нервной
системой. Скелетные мышцы иннервируются соматическим отделом
нервной системы. К каждой мышце подходит один или несколько
нервов, проникающих в ее толщу и разветвляющихся на множество
мелких отростков, которые достигают мышечных волокон.
Посредством нервов осуществляется связь мышц с ЦНС, которая
регулирует любые двигательные акты (ходьба, бег, пищевые
движения и т. д.) и длительное напряжение мышц — тонус,
поддерживающий определенное положение тела в пространстве.
Деятельность мышц носит рефлекторный характер. Мышечный
рефлекс может запускаться с раздражения рецепторов,
находящихся в самой мышце или в сухожилиях, либо с раздражения
зрительных, слуховых, обонятельных, осязательных рецепторов.
Она осуществляется нервной
системой. Скелетные мышцы иннервируются соматическим отделом
нервной системы. К каждой мышце подходит один или несколько
нервов, проникающих в ее толщу и разветвляющихся на множество
мелких отростков, которые достигают мышечных волокон.
Посредством нервов осуществляется связь мышц с ЦНС, которая
регулирует любые двигательные акты (ходьба, бег, пищевые
движения и т. д.) и длительное напряжение мышц — тонус,
поддерживающий определенное положение тела в пространстве.
Деятельность мышц носит рефлекторный характер. Мышечный
рефлекс может запускаться с раздражения рецепторов,
находящихся в самой мышце или в сухожилиях, либо с раздражения
зрительных, слуховых, обонятельных, осязательных рецепторов.
В регуляции безусловно-рефлекторных движений принимает участие
мозжечок. Он осуществляет координацию движения, регуляцию
мышечного тонуса, способствует поддержанию равновесия и позы
тела. При поражении мозжечка его регуляторные двигательные
функции нарушаются.
Он осуществляет координацию движения, регуляцию
мышечного тонуса, способствует поддержанию равновесия и позы
тела. При поражении мозжечка его регуляторные двигательные
функции нарушаются.
Сокращаясь, мышца действует на кость как на рычаг и производит
механическую работу. На осуществление работы мышцы
затрачивается энергия, которая образуется в результате распада
и окисления органических веществ, поступивших в мышечную
клетку. Основным источником энергии является АТФ. Кровь
доставляет мышцам питательные вещества и кислород и уносит
образующиеся продукты диссимиляции (углекислый газ и др.). При
длительной работе наступает утомление и снижение
работоспособности мышцы, возникающее из-за несоответствия
между ее кровоснабжением и возросшими потребностями в
питательных веществах и кислороде. Кроме того, утомление
возникает и вследствие процессов, происходящих в нервных
центрах.
Кроме того, утомление
возникает и вследствие процессов, происходящих в нервных
центрах.
Русский физиолог И. М. Сеченов первым пришел к выводу, что работоспособность мышц зависит от величины нагрузки и ритма работы. Подобрав их оптимальные соотношения, можно добиться высокой производительности работы мышц. И. М. Сеченов установил также, что мышечное утомление проходит и работоспособность восстанавливается гораздо быстрее в результате смены видов деятельности, а не полного бездействия. Тренировка мышц увеличивает их массу, силу и работоспособность. Чрезмерная же работа приводит к утомлению, а бездеятельность — к атрофии.
Систематическая мышечная работа усиливает кровоснабжение мыши
и костей, к которым они прикрепляются. Это приводит
кувеличению мышечной массы и усиленному росту костей. Сильные
мышцы легко справляются с поддержанием туловища в нужном
положении, противостоят развитию сутулости, искривлению
позвоночника.
Это приводит
кувеличению мышечной массы и усиленному росту костей. Сильные
мышцы легко справляются с поддержанием туловища в нужном
положении, противостоят развитию сутулости, искривлению
позвоночника.
Гигиена опорно-двигательной систем ы. Человек рождается с
очень гибким скелетом. Поэтому в детском возрасте особенно
внимательно нужно следить за осанкой ребенка, позой ученика за
партой. Слабо развитые мышцы и неправильная осанка ребенка
могут привести к развитию искривления позвоночника, сутулости,
которые нарушают нормальную деятельность органов грудной
полости и пищеварения. Для предупреждения плоскостопия
(уплощение свода стопы) не следует в период активного роста
человека носить тесную обувь, а также длительно носить обувь
на высоком каблуке. На формировании опорно-двигательного
аппарата организма положительно сказываются активный образ
жизни, подвижные игры, регулярные занятия физкультурой и
спортом.
На формировании опорно-двигательного
аппарата организма положительно сказываются активный образ
жизни, подвижные игры, регулярные занятия физкультурой и
спортом.
Строение и функции скелетных мышц, Биология
Вспомните
1. Каковы особенности строения мышечных тканей?
2. Какое строение имеют мышцы для животных, обитающих в наземно-воздушной среде?
Мышцы, соединенные с костями скелета, называют поперечно-полосатыми или скелетной мускулатурой. Они выполняют в организме целый ряд функций: передвижение человека и частей его тела в пространстве, поддержание позы, дыхательные движения, жевание и глотание, артикуляция и мимика, защита внутренних органов.
В теле человека различные исследователи насчитывают от 400 до 600 мышц. По массе они составляют от 35 до 50% у тренированных спортсменов.
Строение скелетных мышц. Структурной основой скелетных мышц является поперечно-полосатая мышечная ткань, которая состоит из многоядерных клеток , имеющих вид поперечно исчерченных волокон, способных к изменению своей длины, то есть к сокращению.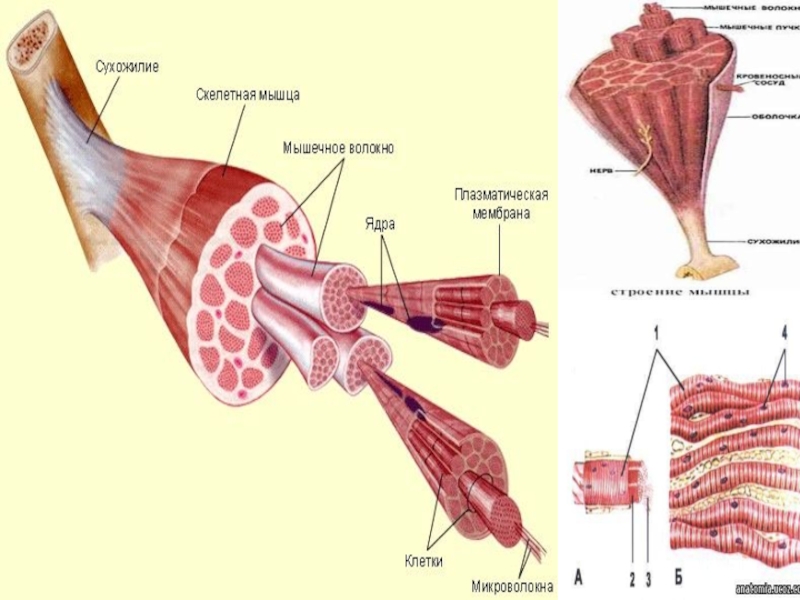 Именно эта ткань образует часть мышцы , называемую брюшко (рис. 22). Волокна собраны в пучки, каждый пучок покрыт оболочкой из соединительной ткани. Пучки, в свою очередь, собраны в скелетную мышцу и тоже покрыты общей соединительно-тканной оболочкой фасцией. На концах мышц эта оболочка утолщается и превращается в сухожилия, которые прикрепляют мышцу к специальным шероховатостям, бугоркам и выростам на костях (см. рис. 22).
Именно эта ткань образует часть мышцы , называемую брюшко (рис. 22). Волокна собраны в пучки, каждый пучок покрыт оболочкой из соединительной ткани. Пучки, в свою очередь, собраны в скелетную мышцу и тоже покрыты общей соединительно-тканной оболочкой фасцией. На концах мышц эта оболочка утолщается и превращается в сухожилия, которые прикрепляют мышцу к специальным шероховатостям, бугоркам и выростам на костях (см. рис. 22).
Рис. 22. Группы скелетных мышц. Мышцы тела человека подразделяют в соответствии с их расположением в организме (рис. 23).
Рис. 23. Мышцы тела человека Мышцы головы и шеи. Мышцы головы по функциям делят на жевательные и мимические. Жевательные мышцы одним концом прикреплены к костям черепа, а другим — к нижней челюсти. Из их названия ясно, что они необходимы для механического измельчения и перемешивания пищи, то есть для её пережёвывания. Мимические мышцы одним концом прикреплены к лицевой части черепа, а другим — к внутренней поверхности кожи лица.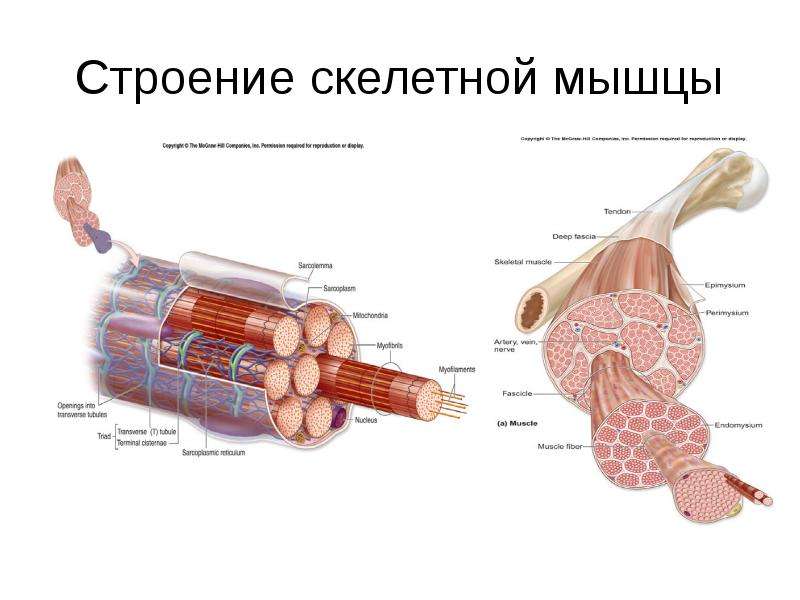 Круговые мышцы рта и глаз вообще не прикреплены к костям. Таким образом, они являются исключением среди мышц скелета, обычно прикреплённых к костям с обеих сторон. Мимические мышцы осуществляют открывание и закрывание глаз, придают лицу определённое выражение, а также служат для произнесения некоторых звуков. Для человека, который постоянно общается с другими людьми, мимические мышцы очень важны. Все мы знаем, что иногда выражение лица говорит больше, чем любые слова. Мышцы шеи нужны для движения не только самой шеи, но и головы, а также нижней челюсти.
Круговые мышцы рта и глаз вообще не прикреплены к костям. Таким образом, они являются исключением среди мышц скелета, обычно прикреплённых к костям с обеих сторон. Мимические мышцы осуществляют открывание и закрывание глаз, придают лицу определённое выражение, а также служат для произнесения некоторых звуков. Для человека, который постоянно общается с другими людьми, мимические мышцы очень важны. Все мы знаем, что иногда выражение лица говорит больше, чем любые слова. Мышцы шеи нужны для движения не только самой шеи, но и головы, а также нижней челюсти.
Мышцы спины осуществляют движения головы, шеи, лопаток. Они могут приподнимать и опускать руки. Кроме того , мышцы спины необходимы для поддержания вертикального положения тела.
Одна группа мышц груд и присоединена к костям плечевого пояса и рук и участвует в их движении. Другая группа называется межрёберпыми мышцами. Именно эти мышцы поднимают и опускают рёбра при внешнем дыхании.
Мышцы живота. Передние и боковые стенки живота образованы мышцам и, которые называются брюшным прессом. Их так назвали потому, что при совместном сокращении они надавливают на внутренние органы, располагающиеся в брюшной полости. Мышцы живота необходимы для поворотов туловища в стороны и наклонов. Они участвуют в дыхательных движениях, а также во многих других процессах жизнедеятельности. При этом брюшной пресс выполняет не только двигательную, но и защитную функцию. К мышцам живота относят также диафрагму, которая герметично разделяет полость тела человека на грудную и брюшную полости. Основная функция диафрагмы — участие в дыхательных движениях.
Их так назвали потому, что при совместном сокращении они надавливают на внутренние органы, располагающиеся в брюшной полости. Мышцы живота необходимы для поворотов туловища в стороны и наклонов. Они участвуют в дыхательных движениях, а также во многих других процессах жизнедеятельности. При этом брюшной пресс выполняет не только двигательную, но и защитную функцию. К мышцам живота относят также диафрагму, которая герметично разделяет полость тела человека на грудную и брюшную полости. Основная функция диафрагмы — участие в дыхательных движениях.
Мышцы плечевого пояса и руки обеспечивают сложнейшие перемещения руки и её отделов.
Мышцы тазового пояса и ноги. Тазовые мышцы обеспечивают движение бедра, мышцы бедра участвуют в движении бедра и голени. Мышцы голени необходимы для движения стопы, и, наконец, мышцы стопы сгибают и разгибают пальцы ног.
Моя лаборатория
Мышечные ткани образованы клетками, обладающими свойствами возбудимости и сократимости.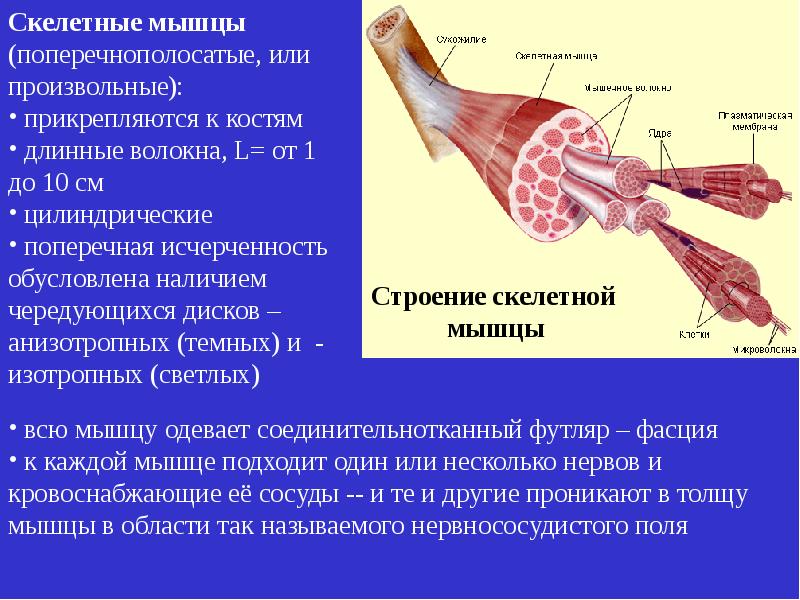 Возбудимость это способность клеток отвечать на внешние раздражители, а сократимость — способность клеток этих тканей менять свои размеры под действием этих самых раздражителей. Дело в том, что в состав мышечных тканей входят особые сократительные белки — актин и миозин, которые, взаимодействуя между собой, уменьшают длину мышечных клеток, и вся мышца сокращается (рис. 24).
Возбудимость это способность клеток отвечать на внешние раздражители, а сократимость — способность клеток этих тканей менять свои размеры под действием этих самых раздражителей. Дело в том, что в состав мышечных тканей входят особые сократительные белки — актин и миозин, которые, взаимодействуя между собой, уменьшают длину мышечных клеток, и вся мышца сокращается (рис. 24).
Рис. 24. Схема мышечного волокна
Поперечно-полосатые мышечные клетки (волокна) очень тонкие, но длинные. Мышечные сократительные белки расположены в этих клетках в строгом порядке и образуют регулярно чередующиеся светлые и тёмные полоски поперёк волокна мышцы, хорошо различимые под микроскопом. Поэтому скелетные мышцы и получили название поперечно-полосатых. Сокращение клеток гладкой мышечной ткани обеспечивается теми же сократительными белками, что и клеток поперечно-полосатых мышц, но эти белки расположены не так упорядоченно, поэтому поперечная исчерченность клеток не видна.
- Какова роль скелетных мышц в работе опорно-двигательной системы и всего организма?
- Каково строение скелетной мышцы?
- Охарактеризуйте особенности основных групп скелетных мышц в связи с их расположением в организме.
 Назовите наиболее развитые мышцы в организме человека и опишите их функции.
Назовите наиболее развитые мышцы в организме человека и опишите их функции.
Подумайте!
Какое значение для мышцы имеют многочисленные кровеносные сосуды и нервные окончания, пронизывающие её?
Общая анатомия мышц, строение мышцы как органа. Развитие скелетных мышц, их классификация (по форме, строению, расположению и т.Д.). Анатомический и физиологический поперечник мышц.
3 0(II) Общая анатомия мышц
В теле и внутренних органах человека находятся скелетные мышцы из исчерченной мышечной ткани, гладкие – из неисчерченной мышечной ткани, сердечная – из мышечной ткани особенного строения и функции.
Скелетные
мышцы связаны с костями и действуют
вместе с ними и суставами в единой
биомеханической системе рычагов,
обеспечивая статику и динамику тела.
Часть из них входит во внутренние органы:
глаз, ухо, полость рта, глотку, пищевод,
гортань, задний проход, влагалище,
мочеиспускательный канал, обеспечивая
движение, формируя стенку и сфинктеры
органов.
Общая масса скелетной мускулатуры составляет у новорожденных – 20-22 %, взрослых – 40 %, стариков – 25-30 % от массы всего тел.
Гладкие мышцы располагаются в коже, сосудах, стенках полых внутренних органов, выделительных протоках желез. Сердечная мышца состоит из проводящих и сократительных мышечных клеток – кардиомиоцитов.
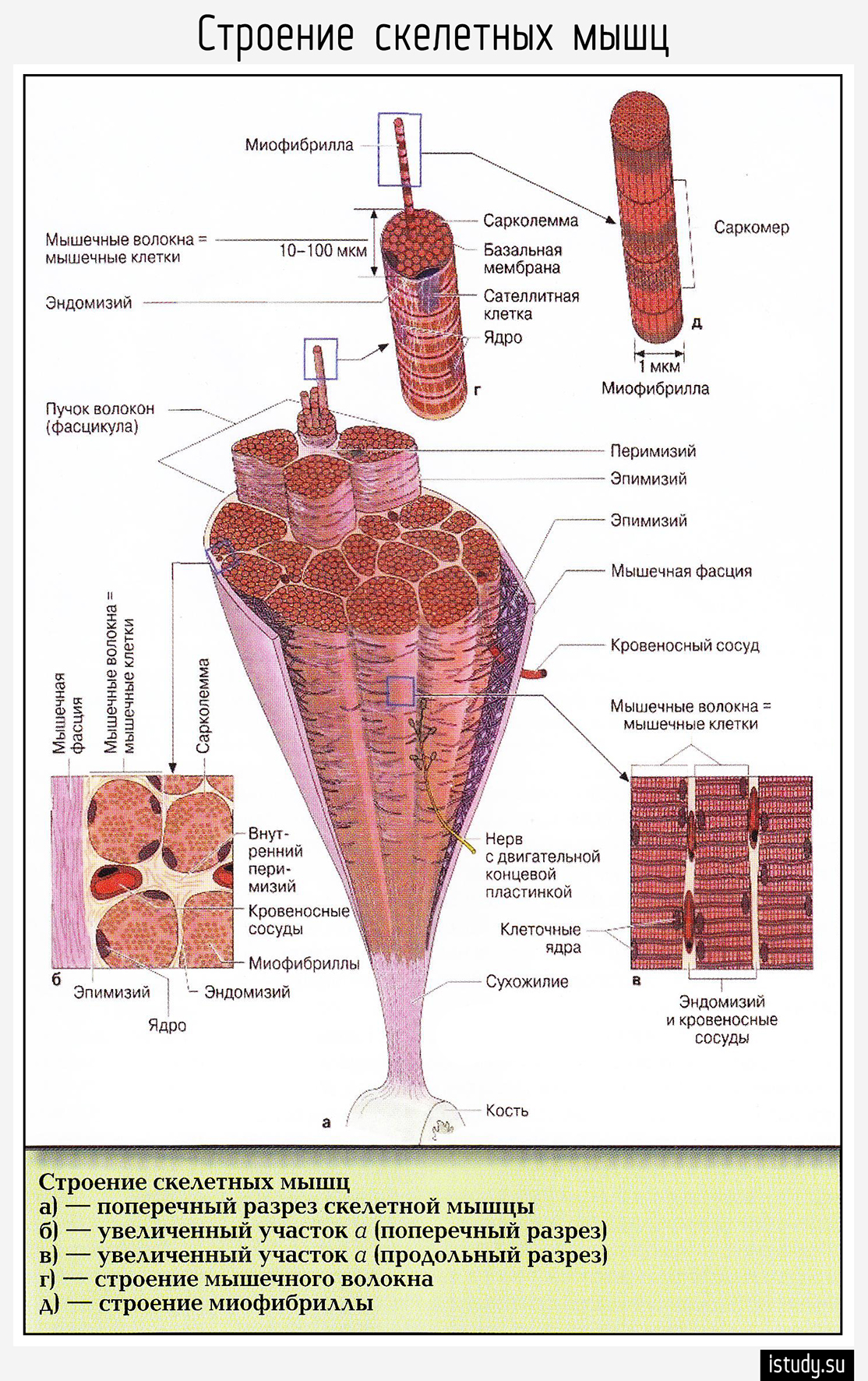
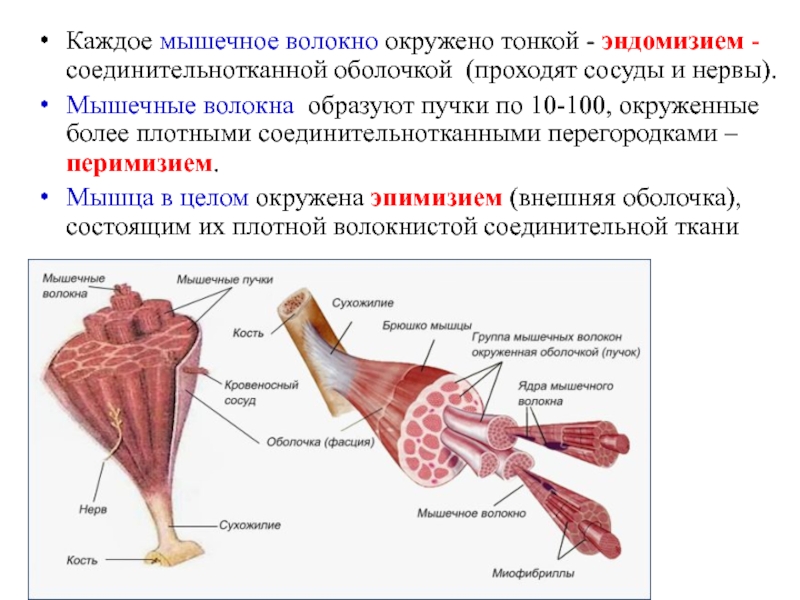
Мускул (мышца) – орган, построенный из мышечных волокон (клеток), каждое из них обладает соединительно-тканной оболочкой – эндомизием. В пучки мышечные волокна объединяет другая фиброзная оболочка – перимизий, а весь мускул заключается в общую фиброзную оболочку, образованную фасцией – эпимизий. Между пучками проходят сосуды и нервы, снабжающие мышечные волокна.
На макроуровне скелетная мышца имеет:
брюшко (вентер) – мясистая часть органа, занимающая его средину;
сухожилие (тендо), относящееся к дистальному концу, оно может быть в виде апоневроза, сухожильных перемычек, длинных пучков продольных фиброзных волокон;
головку, составляющую проксимальную часть;
сухожилие и головка крепятся на противоположных концах костей.
Сила мышцы на 1 см 2 ее поперечного сечения называется абсолютной и составляет от 50 до 100 Н, что зависит от длины мышечных волокон и площади поперечного сечения. Начало мышцы на кости находится ближе к срединной оси тела – это фиксированная точка (пунктум фиксум), конец мышцы лежит на кости дистальнее и, являясь местом прикрепления, называется подвижной точкой (пунктум мобиле). При сокращении мышцы точки сближаются, а при изменении положения тела могут меняться местами.
Мышцы развиваются из мезодермы. На туловище они возникают из первичной сегментированной мезодермы – сомитов: 3-5 затылочных, 8 шейных, 12 грудных, 5 поясничных, 5 крестцовых, 4-5 копчиковых.
Каждый сомит подразделяется на склеротом, дерматом и миотом – из него и развиваются мышцы туловища.
Сомиты появляются рано, когда длина зародыша составляет 10-15 мм.
Из
дорсальных частей миотомов возникают
глубокие, собственные (аутохтонные)
мышцы спины, из вентральных – глубокие
мышцы груди и живота. Они закладываются,
развиваются и остаются в пределах
туловища – поэтому называются аутохтонными
(местными, туземными). Очень рано миотомы
связываются с нервной системой и каждому
мышечному сегменту соответствует
нервный сегмент. Каждый
нерв следует за развивающейся мышцей,
врастает в нее и, пока она не
дифференцировалась, подчиняет своему
влиянию.
Они закладываются,
развиваются и остаются в пределах
туловища – поэтому называются аутохтонными
(местными, туземными). Очень рано миотомы
связываются с нервной системой и каждому
мышечному сегменту соответствует
нервный сегмент. Каждый
нерв следует за развивающейся мышцей,
врастает в нее и, пока она не
дифференцировалась, подчиняет своему
влиянию.
В процессе развития часть скелетных мышц перемещается с туловища и шеи на конечности – трункофугальные мышцы: трапециевидная, грудино-ключично-сосцевидная, ромбовидные, поднимающая лопатку и др.
Часть мышц, наоборот, направляется с конечностей на туловище – трункопетальные мышцы: широчайшая мышца спины, большая и малая грудные, большая поясничная.
Мышцы
головы мимические и жевательные, над-
и подъязычные мышцы шеи развиваются из
несегментированной вентральной
мезодермы, входящей в состав висцеральных
(жаберных) дуг. Они называются висцеральными
и, например, жевательные мышцы развиваются
на основе первой висцеральной дуги, а
мимические – второй. Однако мышцы
глазного яблока и языка развиваются из
затылочных миотомов сегментированной
мезодермы. Глубокие передние и задние
мышцы шеи также возникают из затылочных
шейных миотомов, а поверхностная и
средняя группа мышц в передней области
шеи развиваются на основе несегментированной
мезодермы висцеральных дуг.
Однако мышцы
глазного яблока и языка развиваются из
затылочных миотомов сегментированной
мезодермы. Глубокие передние и задние
мышцы шеи также возникают из затылочных
шейных миотомов, а поверхностная и
средняя группа мышц в передней области
шеи развиваются на основе несегментированной
мезодермы висцеральных дуг.
Страница не найдена |
Страница не найдена |404. Страница не найдена
Архив за месяц
ПнВтСрЧтПтСбВс
12
12
1
3031
12
15161718192021
25262728293031
123
45678910
12
17181920212223
31
2728293031
1
1234
567891011
12
891011121314
11121314151617
28293031
1234
12
12345
6789101112
567891011
12131415161718
19202122232425
3456789
17181920212223
24252627282930
12345
13141516171819
20212223242526
2728293031
15161718192021
22232425262728
2930
Архивы
Мар
Апр
Май
Июн
Июл
Авг
Сен
Окт
Ноя
Дек
Метки
Настройки
для слабовидящих
Механико-математическая модель поперечнополосатой мышцы Текст научной статьи по специальности «Нанотехнологии»
электронное научно-техническое издание
НАУКА и ОБРАЗОВАНИЕ
Эл МЭ ФС 77 — 30569. Государственная регистрация N30420900025. ISSN 1994-0408
Государственная регистрация N30420900025. ISSN 1994-0408
Механико-математическая модель поперечнополосатой мышцы # 08, август 2010
авторы: Галямова Е. В., Гуськов А. М., Сюзев В. В.
УДК: 612.741, 51-7, 57.087
МГТУ им. Н.Э. Баумана
1. Введение
Моделирование мышечного сокращения является важным компонентом при изучении физиологических особенностей движения человека. Знание информативных параметров механических (упруго-вязких) свойств мышцы используется в медицине [1], в изобразительном искусстве при рассмотрении влияния подкожной мускулатуры на форму тела человека [2-5]. В спорте моделирование движения мышц человека помогает тренерам повышать эффективность спортивных тренировок [3, 4, 6], причем, возможности современных компьютеров позволяют проводить исследования и вводить коррекцию в методику тренировки непосредственно во время ее проведения [7].
В статье описываются упруго-вязкие свойства мышцы спортсмена, как основные информативные параметры для разработки принципов организации обратной связи и
контроля состояния спортсмена (степени усталости) до, во время и после тренировки. Основная задача новой модели состояла в нахождении такой зависимости между физическими параметрами мышцы, которая позволила бы сравнивать различные мышцы, имеющие сходное состояние, в относительных единицах.
В начале статьи подробно рассмотрено строение мышцы и механизмы сокращения в мышце. Сделан краткий обзор существующих моделей сокращения скелетных мышц и математически обоснована предлагаемая новая модель сокращения мышечной ткани. Отметим также, что именно на основании литературных данных найдены ключевые численные показатели, как опорные для моделирования. Обоснована необходимость введения в модель дополнительного элемента — упругой нити, с помощью которой могут быть получены искомые зависимости.
2. Строение скелетной мышцы
Основным элементом скелетной мышцы является мышечная клетка. В связи с тем, что мышечная клетка по отношению к своему поперечному сечению (0,05-0,11 мм) относительно длинна (волокна бицепса, например, имеют длину до 15 см), ее называют также мышечным волокном. Скелетная мышца состоит из большого количества этих структурных элементов, составляющих 85-90% от ее общей массы. Так, например, в состав бицепса входит более одного миллиона волокон. Между мышечными волокнами расположена тонкая сеть мелких кровеносных сосудов (капилляров) и нервов (приблизительно 10% от общей массы мышцы). От 10 до 50 мышечных волокон соединяются в пучок. Пучки мышечных волокон и образуют скелетную мышцу.
Мышечные волокна, пучки мышечных волокон и мышцы окутаны соединительной тканью. Общее строение скелетной мышцы показано на рисунке 1. ниечных яПппкЛы
ниечных яПппкЛы
Сократительные нити бвпк& миозина
Мышечные волокна на своих концах переходят в сухожилия. Через сухожилия, прикрепленные к костям, мышечная сила воздействует на кости скелета. Сухожилия и другие эластичные элементы мышцы обладают, кроме того, и упругими свойствами. При высокой и резкой внутренней нагрузке — сильной мышечной тяге, или при сильном и внезапном внешнем силовом воздействии эластичные элементы мышцы растягиваются и тем самым смягчают силовые воздействия, распределяя их в течение более продолжительного промежутка времени. Поэтому после хорошей разминки в мускулатуре редко происходят разрывы мышечных волокон и отрывы от костей. Сухожилия обладают значительно большим пределом прочности на растяжение (около 7 000 К/кв см), чем мышечная ткань (около 6ОК/кв см), поэтому, они гораздо тоньше, чем брюшко мышцы.
В мышечном волокне содержится основное вещество, называемое саркоплазмой. В саркоплазме находятся митохондрии (30-35% от массы волокна), в которых протекают процессы обмена веществ и накапливаются вещества, богатые энергией, например, фосфаты, гликоген и жиры. В саркоплазму погружены тонкие мышечные нити (миофибриллы), лежащие параллельно длинной оси мышечного волокна. Миофибриллы составляют в совокупности приблизительно 50% массы волокна, их длина равна длине мышечных волокон, и они являются, собственно говоря, сократительными элементами мышцы. Они состоят из небольших, последовательно включаемых элементарных блоков, именуемых также саркомерами. Особенность строения саркомеров позволяет им укорачиваться при соответствующем нервном импульсе. Процесс сократительного акта в саркомере можно упрощенно сравнить с движениями гребцов в академической лодке (рис.2). Саркомеры состоят из двух видов белковых филаментов: более тонких -актиновых и более толстых — миозиновых. Из филаментов миозина с обеих сторон, подобно веслам в лодке, выступают отростки (миозиновые мостики).
В саркоплазме находятся митохондрии (30-35% от массы волокна), в которых протекают процессы обмена веществ и накапливаются вещества, богатые энергией, например, фосфаты, гликоген и жиры. В саркоплазму погружены тонкие мышечные нити (миофибриллы), лежащие параллельно длинной оси мышечного волокна. Миофибриллы составляют в совокупности приблизительно 50% массы волокна, их длина равна длине мышечных волокон, и они являются, собственно говоря, сократительными элементами мышцы. Они состоят из небольших, последовательно включаемых элементарных блоков, именуемых также саркомерами. Особенность строения саркомеров позволяет им укорачиваться при соответствующем нервном импульсе. Процесс сократительного акта в саркомере можно упрощенно сравнить с движениями гребцов в академической лодке (рис.2). Саркомеры состоят из двух видов белковых филаментов: более тонких -актиновых и более толстых — миозиновых. Из филаментов миозина с обеих сторон, подобно веслам в лодке, выступают отростки (миозиновые мостики).
Так как длина саркомера в состоянии покоя равна приблизительно лишь 0,0002 мм, то, для того, чтобы, к примеру, образовать цепочки из звеньев миофибрилл бицепса длиной 10-15 см, необходимо „соединить» огромное количество саркомеров. Толщина мышечных волокон зависит, главным образом, от количества и поперечного сечения миофибрилл.
Рис. 2. Самый малый строительный элемент скелетной мышцы (саркомер) в состоянии
покоя
Реагируя на нервный сигнал и последующую химическую реакцию, отростки миозина временно пристыковываются к филаментам актина (в виде мостиков соединения, а затем отводятся в „позицию под углом 45″» (рис. 3).
За счет этих движений, которые можно сравнить с опусканием в воду весел (захват воды) и последущим гребком, филаменты актина перемещаются между филаментами миозина. р
р
ч
После выполнения „гребка» отростки миозина примерно также, как весла, поднимаются из воды, отрываются от актина и возвращаются в исходное положение. За один такой „гребок» саркомер укорачивается всего лишь примерно на 1 % своей длины. Следовательно, для достижения телескопического соединения филаментов, вызывающего эффективное напряжение, требуется большое число „гребцов». Нервная система, выступая в роли „рулевого», может, в зависимости от структуры мышечного волокна и требуемой величины напряжения, подавать сигналы с частотой от 7 до более чем 50-ти „гребков» в секунду. В связи с тем, что огромное количество саркомеров, расположенных по ходу миофибриллы, включается последовательно, их единичные минимальные сокращения суммируются, и миофибрилла сокращается на 25-30%. Так как большое количество миофибрилл расположено рядом, их относительно небольшие сократительные силы складываются в суммарную силу мышечного волокна и в итоге в мышцы. ——1——-н
——1——-н
Рис. 4. Самый малый строительный элемент скелетной мышцы (саркомер) в
расслабленном состоянии
При сильном и предельном укорачивании мышцы (длина саркомера 0,0016-0,0013 мм) концы филаментов актина все глубже проникают между филаментами миозина и их тяга постоянно уменьшается, образовывать новые мостики становится все сложней. Напряжение мышцы постоянно спадает (рис. 5).
л
—1 1 1 г4\
1
1—Ч*—
Рис. 5. Самый малый строительный элемент скелетной мышцы (саркомер) в состоянии
сокращения
3. Механические модели мышцы
Механические модели мышцы
Многие существующие механические модели, описывающие сократительные и упругие элементы мышцы, имеют в своей основе классическую модель мышцы Хилла, в частности, механическая модель мышечной активности, предложенная исследователями в области спортивной медицины [8] , приведена на рисунке 6.
Как видно из рисунка 6, такая модель позволяет описать несколько состояний мышцы: а -в покое, Ь — в статическом режиме работы, с — в динамическом режиме работы, ё — в растянутом состоянии. Сократительный компонент СК, который состоит из мышечных волокон или миофибрилл, а также два эластичных компонента: параллельный эластичный компонент Пар, в состав которого входят трубчатые соединительно-тканные оболочки мышечных волокон (эндомизий) и пучки мышечных волокн (перемизий), и последовательный эластичный компонент Пос, образованный сухожилиями, позволяют моделировать состояния мышцы.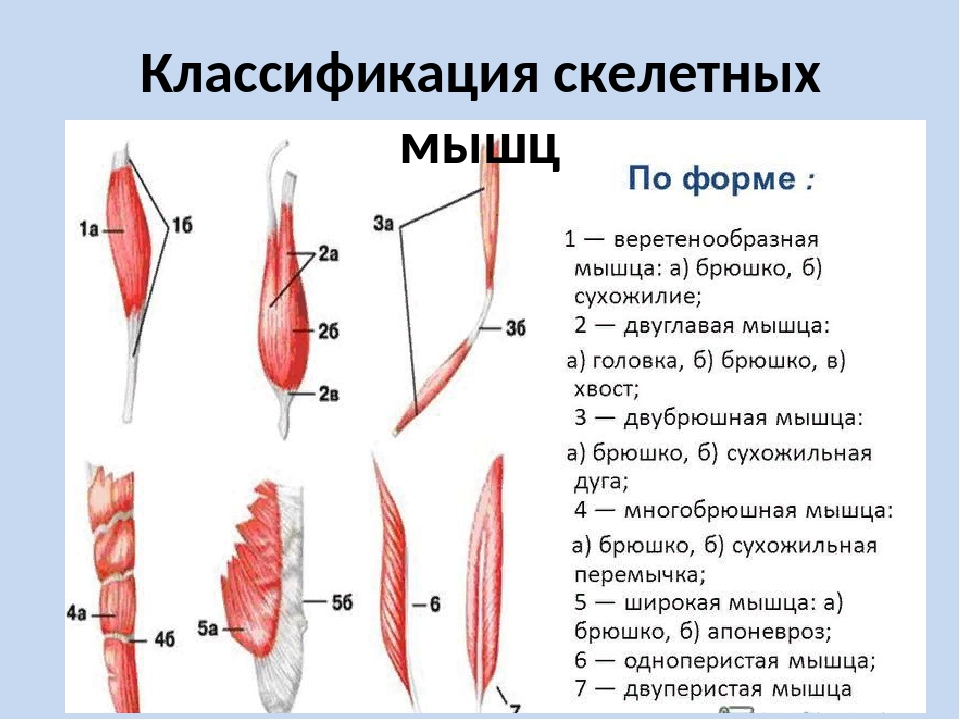 При этом внутренняя энергия мышцы складывается из энергии сокращения (СК) и энергии предварительного растягивания (Пар + Пос), а внешняя прикладываемая к мышце сила рассматривается как внешнее сопротивление.
При этом внутренняя энергия мышцы складывается из энергии сокращения (СК) и энергии предварительного растягивания (Пар + Пос), а внешняя прикладываемая к мышце сила рассматривается как внешнее сопротивление.
Процесс сокращения элементарного блока миофибриллы представляет собой энергетический процесс, в котором химическая энергия превращается в механическую работу. Взаимодействие сократительных и эластичных компонентов мышцы наглядно изображено на механической модели мышцы (рис. 3). Сократительный компонент мышцы (СК) состоит из миофибрилл. Эластичный компонент подразделяется на последовательно включаемый эластичный компонент (Пос) и параллельно-эластичный компонент (Пар). В состав первого входят сухожилия и другие элементы соединительной ткани мышцы, второй образуется, в частности, из соединительно-тканных оболочек мышечных волокон и их пучков. Если укорачивается сократительный компонент, то сначала растягивается Пос (рисунок 3 Ь). Лишь после того, как развиваемая в Пос сила напряжения превысит величину внешней силы (например, сопротивление соперника или поднимаемого с земли отягощения), сократится вся мышца.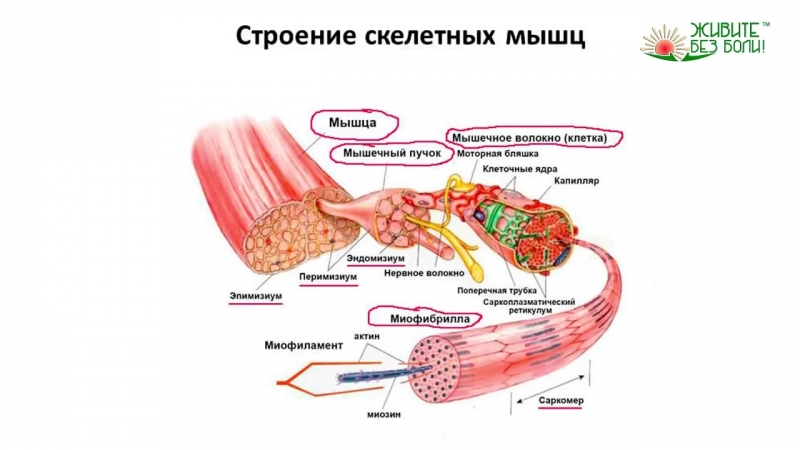 Напряжение Пос во время укорачивания мышцы остается постоянным (рисунок 3 с). Пар помогает сначала укоротить сократительный компонент, а затем вернуть его к длине покоя. Если мышца растягивается, то внешняя сила настолько сильно удлиняет Пос, что в конце концов за ним приходится следовать и сократительному компоненту (СК) (рисунок 3 ё). При длине покоя мышца может развить очень высокое напряжение. Во-первых, потому что оптимальная степень контакта филаментов актина и миозина позволяет создать максимальное количество мостиковых соединений и тем самым активно и сильно развить напряжение сократительного комнонента. Во-вторых, потому что эластичный компонент мышцы уже как пружина предварительно растянут, уже создано дополнительное напряжение. Активно развитое напряжение сократительного компонента суммируется с упругим напряжением, накопленным в эластичном компоненте, и реализуется в одно высокое, результирующее напряжение мышцы. Последующее предварительное растяжение мышцы, которое значительно превосходит состояние при длине покоя, приводит к недостаточному контакту филаментов актина и миозина.
Напряжение Пос во время укорачивания мышцы остается постоянным (рисунок 3 с). Пар помогает сначала укоротить сократительный компонент, а затем вернуть его к длине покоя. Если мышца растягивается, то внешняя сила настолько сильно удлиняет Пос, что в конце концов за ним приходится следовать и сократительному компоненту (СК) (рисунок 3 ё). При длине покоя мышца может развить очень высокое напряжение. Во-первых, потому что оптимальная степень контакта филаментов актина и миозина позволяет создать максимальное количество мостиковых соединений и тем самым активно и сильно развить напряжение сократительного комнонента. Во-вторых, потому что эластичный компонент мышцы уже как пружина предварительно растянут, уже создано дополнительное напряжение. Активно развитое напряжение сократительного компонента суммируется с упругим напряжением, накопленным в эластичном компоненте, и реализуется в одно высокое, результирующее напряжение мышцы. Последующее предварительное растяжение мышцы, которое значительно превосходит состояние при длине покоя, приводит к недостаточному контакту филаментов актина и миозина. При этом заметно ухудшаются условия для
При этом заметно ухудшаются условия для
развития значительного и активного напряжения саркомеров. Тем не менее при большом предварительном растягивании задействованных мышц, например, при широком замахе в метании копья, спортсмены достигают более высоких результатов, чем без замаха. Этот феномен объясняется тем, что увеличение предварительного напряжения эластичного компонента превосходит снижение активного развития напряжения сократительного компонента.
Добавим в описанную модель еще один элемент — упругую нить, считая ее аналогом мышечного волокна, оставив модель Хилла для описания упруго-вязких свойств остальных компонентов мышцы и объединив все остальные компоненты мышцы, кроме мышечного волокна, одним словом — подложка. В следующем разделе опишем предлагаемую модель математически.
3. Математическая модель мышцы
Представим мышцу в виде двумерной распределенной системы «упругая нить -упруго-вязкая подложка».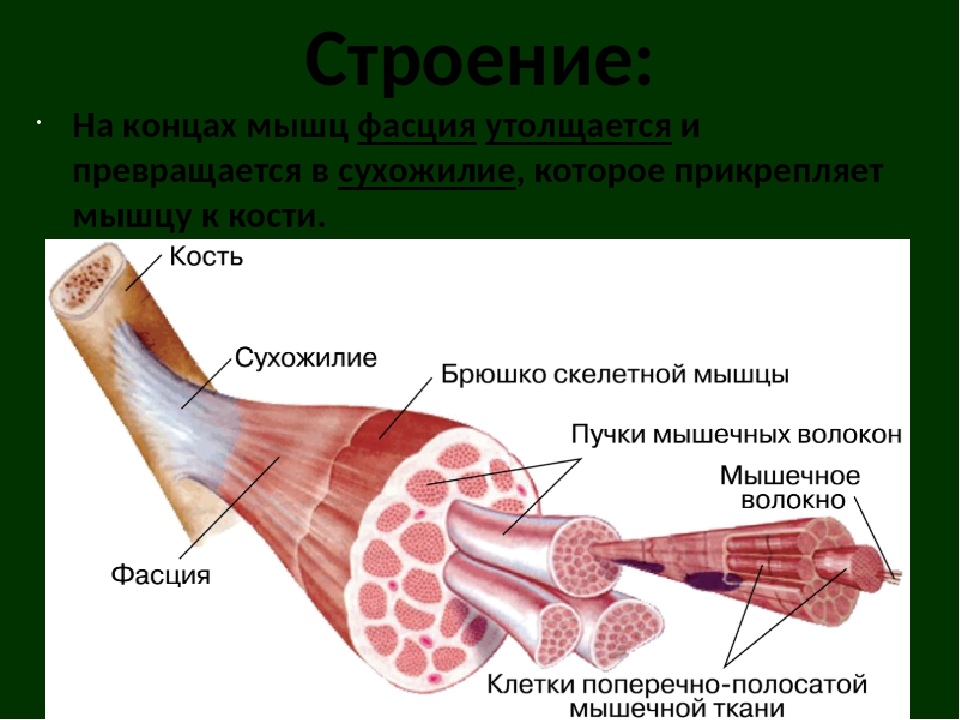 Нить закреплена с двух сторон и имеет предварительное натяжение. Упругая нить является аналогом мышечных волокон. Обозначим активную сократительную силу, растягивающую нить, через Т. Причем, естественно принять, что в расслабленной мышце сократительная сила близка к нулю, а в максимально напряженной (в состоянии тетануса) — максимальна. Таким образом, способность мышцы сокращаться описывается наличием сократительной силы Т. Упруго-вязкая подложка является аналогом пассивных компонентов мышцы и подмышечных тканей. В этой модели мы пренебрегаем массой мышечных волокон по сравнению с массой инерционных, пассивных компонентов. В дальнейшем изучаются колебания мышцы в направлении, перпендикулярном свободной поверхности. Уравнения колебаний указанной системы в величинах идеализированной механической модели мышцы позволяют связать
Нить закреплена с двух сторон и имеет предварительное натяжение. Упругая нить является аналогом мышечных волокон. Обозначим активную сократительную силу, растягивающую нить, через Т. Причем, естественно принять, что в расслабленной мышце сократительная сила близка к нулю, а в максимально напряженной (в состоянии тетануса) — максимальна. Таким образом, способность мышцы сокращаться описывается наличием сократительной силы Т. Упруго-вязкая подложка является аналогом пассивных компонентов мышцы и подмышечных тканей. В этой модели мы пренебрегаем массой мышечных волокон по сравнению с массой инерционных, пассивных компонентов. В дальнейшем изучаются колебания мышцы в направлении, перпендикулярном свободной поверхности. Уравнения колебаний указанной системы в величинах идеализированной механической модели мышцы позволяют связать
соотношения основного параметра модели — натяжения мышцы , — с показателями колебательного процесса системы, как условный период затухающих колебаний, декремент и частота свободных колебаний системы. Колебательный процесс в мышце может быть вызван нанесением ей ударного импульса, перпендикулярного свободной поверхности. Ударный импульс наносится в центр системы, а регистрация колебаний осуществляется в точке, расстояние от которой до места удара известно. Декремент колебаний определяется из логарифмического отношения двух последовательных амплитуд колебания, разделенного на соответствующий временной интервал по формуле:
Колебательный процесс в мышце может быть вызван нанесением ей ударного импульса, перпендикулярного свободной поверхности. Ударный импульс наносится в центр системы, а регистрация колебаний осуществляется в точке, расстояние от которой до места удара известно. Декремент колебаний определяется из логарифмического отношения двух последовательных амплитуд колебания, разделенного на соответствующий временной интервал по формуле:
4
4+1′ ‘ (1)
где Р — логарифмический декремент колебаний; А> А+1 — амплитуды двух последовательных колебаний, отделенных друг от друга периодом свободных колебаний
связанных с угловой частотой — .
Частота собственных колебаний Р мышцы определяется по формуле:
Р = (2)
3. 1. Математическое определение модели
1. Математическое определение модели
В данном приближении мышца локально рассматривается как плоский объект, имеющий сложную структуру. Двумерная система «упругая нить — упруго-вязкая подложка» представляется набором параллельных нитей, соединенных с упруго-вязкой подложкой. Предположим, что в движение вовлекается некоторый эффективный слой подложки глубиной И . Принимается линейный закон распределения по координате У компоненты поля перемещения, нормальной к поверхности мышцы:
где
нормальная к поверхности мышцы составляющая поля вектора
перемещения,
перемещение точки волокна, отстоящей от левого края на расстоянии ж
вдоль оси
Оу
— Н — глубина эффективного слоя подложки,
-У — координата, направленная вовне от свободной поверхности мышцы,
— г — координата оси волокна.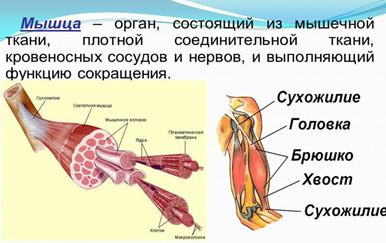 2
2
д£
-Н
п / \ т
где
& — ширина рассматриваемой полосы подложки,
Р — плотность подложки,
(г, г)
— скорость осин струны в точке х .
Произведя вычисления
в
(4)
получаем
(5)
Силы, действующие на малый эффект нити, показаны на рисунке 7.
А / 5 • Р44
т 7 4 1 } гь
\ чф
± Г
Рис. — д
— д
д£’
(6)
где
Т — сила натяжения нити, принимаемая неизменной вдоль всей нити,
И=н.
распределенная поперечная сила со стороны подложки, направленная против
оси
у
.
Уравнение
(6)
дополняется условиями закрепления нити и начальными условиями. Вводится понятие эффективной длины нити Ь . По концам нить закреплена
Распределенная сила $ определяется через напряжение в мышце — подложке с, умноженное на эффективную ширину Ъ
(8)
Реологические характеристики подложки представим трехкомпонентной моделью упруго —
вязкого тела:
Рис. — коэффициент вязкости вязкого элемента.
— коэффициент вязкости вязкого элемента.
Введем следующую нормировку коэффициента вязкости
т —
Величина В играет роль постоянной времени для процессов затухания свободных колебаний мышцы. Тогда уравнения закона состояния упруго-вязкого элемента, представленного на рис. 2, принимает вид
(10)
В соответствии с законом распределения
перемещений
уравнение
представляется, как
(3)
(10)
\ ‘ Нд1 * н
(11)
Считая, что вязкая составляющая достаточно мала, представим решение
уравнения
в виде трехчленного регулярного разложения по параметру В
(11)
Н
(12)
После подстановки
выражений
в уравнения колебаний
получим следующее уравнение колебаний нити
(12)
(6)
в?
д£
(13)
где обозначены:
М = т
ЬВ2Е„
Н
ЬА
И
а
,_га
н
Таким образом, имеем следующий список из восьми физических параметров,
характеризующих динамику мышцы
{т. ЬЛН.Т.Ег.Ея
ЬЛН.Т.Ег.Ея
(14)
3.2. Приведение к безразмерному виду, выбор масштабов
Используем три масштаба: масштаб перемещений И ~ ш в , масштаб длины , [2*] = т и масштаб времени Т* — I7* 1= 3
(15)
Подставим
(15)
(13)
и разделим на
Ж
блок :
Зт М дт М
т,с
М2? дС?
(16)
Масштаб длины назначим равным эффективной длине нити
(17)
Для выбора масштаба времени, предположим, что в случае отсутствия диссипации в подложке собственные колебания по первой форме
(т>0 = 17(т)г:1п М
(18)
имеют круговую частоту равную единице
в
2тг1 т2
Предположим, что в случае с диссипацией приближенно выполняется закон
движения
(18)
тогда для амплитудной функции ^ уравнение колебаний в безразмерной форме
можно записать в виде
-Ц^ + Ъг-^ + дЬ =0
(20)
где нормированный коэффициент демпфирования определяется как
й
2%1м{к + ъТ/1?)
(21)
Важно отметить, что коэффициент демпфирования я зависит от натяжения волокон мышцы 7.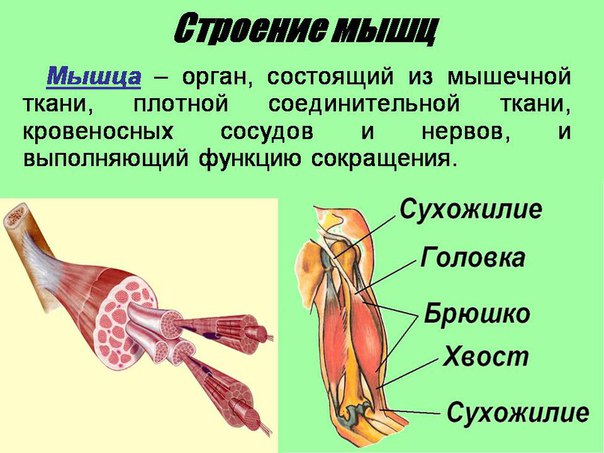 ) , где 2о — координата фиксации акселерометра.
) , где 2о — координата фиксации акселерометра.
Благодарность
Авторы статьи благодарны Министерству науки и образования РФ за финансовую поддержку в виде Государственного контракта №П1274 от 27.08.2009 на проведение поисковой научно-исследовательской работы по теме «Разработка методологии совершенствования учебно-тренировочного процесса учащихся ВУЗов и спортсменов высшей квалификации на основе изучения закономерностей физиологии опорно-двигательного аппарата», в рамках которой получены представленные материалы.
Литература
1. Бэгшоу К. Мышечное сокращение. — М.: Мир, 1985. 159 с.
2. Новоселов В.С., Королев В.С. Модель возбуждения мышцы // Труды 4-й Международной конференции «Идентификация систем и проблемы управления» -М. , 2005. С. 367-374.
, 2005. С. 367-374.
3. Волынкина Г.Ю. Электромиографическое исследование состояний длительного эмоционального напряжения / Г.Ю. Волынкина, Ш.М. Замаховер, Д.Р. Зельдович, Ю.А. Тевеленок // Актуальные вопросы клинической и судебной психиатрии:
Сборник трудов Ленинградского научного общества невропатологов и психиатров. 1970. Вып. 14. С. 250-256.
4. Таркова К.Р. Соотношения между потенциалами действия скелетной мышцы и её напряжением при различных функциональных состояниях // Материалы конференции молодых ученых Воронежского медицинского института. Воронеж, 1996. С. 157-158.
5. Marque C., a.a. Uterine EHG processing for obstetrical monitoring // IEEE Trans. Biomed. Eng. 1986. V. 33, № 12. P. 1182-1187.
6. Верхошанский Ю.В. Горизонты научной теории и методологии спортивной тренировки // Теория и практика физической культуры, №7, 1998. С. 41-54.
С. 41-54.
7. Мирский М.Л. Сравнительная достоверность параметров интерференционной ЭМГ человека // Материалы конференции молодых научных работников. М., 1963. С. 8486.
8. Пахомова Т.Г. О взаимосвязи между твердостью, вязкостью, силой и биоэлектрической активностью мышц человека: Автореф. диссертация кандидата биологических наук. Тарту, 1973. С. 16.
9. Зайцев А. А. Электромиографическая характеристика активности «быстрой» и «медленной» мышц при динамической работе // Физиология человека и животных (Тула). 1973. Вып. 7. С. 40-45.
10. Зайцев А.А. Электромиографическая характеристика активности «быстрой» и «медленной» мышц в связи с утомлением // Материалы 10-й Всесоюзной научной конференции по физиологии и биомеханике мышечной деятельности. М., 1968. Т.1. С.176-178.
11. сЬоо1-co11ection.edu.ru/d1rstore/000005a5-1000-4ddd-bf72-570046bc5020/b08_042.jpg
сЬоо1-co11ection.edu.ru/d1rstore/000005a5-1000-4ddd-bf72-570046bc5020/b08_042.jpg
скелетных мышц | Определение и функции
Скелетная мышца , также называемая произвольной мышцей , у позвоночных, наиболее распространенная из трех типов мышц тела. Скелетные мышцы прикреплены к костям с помощью сухожилий, и они производят все движения частей тела по отношению друг к другу. В отличие от гладких мышц и сердечной мышцы, скелетные мышцы находятся под произвольным контролем. Однако, как и сердечная мышца, скелетная мышца имеет поперечно-полосатую форму; его длинные, тонкие, многоядерные волокна пересекаются правильным рисунком из тонких красных и белых линий, что придает мышце характерный вид.Волокна скелетных мышц связаны между собой соединительной тканью и сообщаются с нервами и кровеносными сосудами. Для получения дополнительной информации о структуре и функции скелетных мышц, см. мышечная и мышечная система человека.
Строение поперечно-полосатой или скелетной мышцы. Поперечно-полосатая мышечная ткань, такая как ткань двуглавой мышцы человека, состоит из длинных тонких волокон, каждое из которых, по сути, представляет собой пучок более тонких миофибрилл. Внутри каждой миофибриллы находятся филаменты белков миозина и актина; эти нити скользят друг мимо друга по мере того, как мышца сокращается и расширяется.На каждой миофибрилле можно увидеть регулярно встречающиеся темные полосы, называемые Z-линиями, в местах наложения актиновых и миозиновых филаментов. Область между двумя линиями Z называется саркомером; саркомеры можно рассматривать как первичную структурную и функциональную единицу мышечной ткани.
Encyclopædia Britannica, Inc. скелетная мышцаМикрофотография, показывающая расположение волокон скелетных мышц в поперечном сечении.
© Эд Решке / Питер Арнольд, Inc.
Британская викторина
Человеческое тело
Возможно, вы знаете, что человеческий мозг состоит из двух половин, но какая часть человеческого тела состоит из крови? Проверьте обе половины своего разума в этой викторине по анатомии человека.
Ткани скелетных мышц: гистология | Kenhub
Гистология скелетных мышц: хотите узнать об этом больше?
Наши увлекательные видео, интерактивные викторины, подробные статьи и HD-атлас помогут вам быстрее достичь лучших результатов.
С чем вы предпочитаете учиться?
«Я бы честно сказал, что Kenhub сократил мое учебное время вдвое». — Прочитайте больше. Ким Бенгочеа, Университет Реджиса, Денвер
Автор:
Джули Долл Бакалавр, Магистр наук
• Рецензент:
Урудж Зехра MBBS, MPhil, PhD  » data-popover-cta=»{"href":"/en/quality","title":"Read more."}»/>
» data-popover-cta=»{"href":"/en/quality","title":"Read more."}»/>
Последняя редакция: 23 февраля 2021 г.
Время чтения: 15 минут
Скелетная мышца — это возбудимая сократительная ткань, отвечающая за поддержание осанки и движение орбит, вместе с аппендикулярным и осевым скелетами.Он прикрепляется к костям и глазницам через сухожилия. Возбудимая ткань реагирует на раздражители электрическими сигналами. Сократительная ткань способна создавать силовое напряжение.
Ткани скелетных мышц также растяжимы и эластичны. Растяжимая ткань может растягиваться, а эластичная ткань может возвращаться к своей первоначальной форме после деформации.
| Функция | Поддерживает осанку и позволяет двигаться |
| Ячеистая структура | Сарколемма — клеточная мембрана Терминальная цистерна — расширение сарколеммы, в которой хранится кальций Т-канальцы — инвагинации сарколеммы, передающие потенциал действия внутрь мышечной клетки 9000mic3 Саркоплазма — цитоплазма — модифицированный эндоплазматический ретикулум Актин, миозин — сократительные элементы Саркомер — функциональная единица (состоящая из актина и миозина) Вспомогательные белки — тайтин, тропомодулин, альфа-актинин, десмин, небулин, дистрофин53, миозин53, |
| Типы волокон (ячеек) | Тип I — используйте аэробный метаболизм для работы; они кажутся красными из-за большого количества миоглобина; они медленно сокращаются и устойчивы к утомлению. Тип IIa — получают энергию от окислительного гликолиза; имеют высокое количество гликогена, более интенсивны, чем тип I, они быстро сокращаются и устойчивы к утомлению Тип IIb — получают энергию от анаэробного гликолиза, выглядят розовыми, быстро сокращаются и склонны к утомлению |
| Оболочка ткани | Endomysium — вокруг одного мышечного волокна Perimysium — вокруг нескольких мышечных волокон -> объединяет их в пучков Эпимизий — вокруг всей мышцы |
| Нервно-мышечное соединение | Место на сарколемме, где моторные волокна соединяются с мышцами, чтобы передать команду сокращения; Нейротрансмиттер ацетилхолин |
| Клинические отношения | Мышечная дистрофия, миопатия совокупного актина, миотубулярные миопатии |
В этой статье речь пойдет о гистологии скелетных мышц.
Терминология
Специальные термины используются для описания структур, связанных со скелетной мышечной тканью. Термины «мышечная ткань» часто начинаются с мио-, мис- или сарко-. Цитоплазма мышечных клеток обозначается как , саркоплазма . Плазматическая мембрана называется сарколеммой , а эндоплазматический ретикулум называется саркоплазматической сетью . Мышечное волокно также может называться миофибром .
По отдельности клетки скелетных мышц обозначаются как мышечных волокон . Длина волокна скелетных мышц зависит от его местоположения. В передней части бедра мышечное волокно может быть длиной в метр. Напротив, мышечные волокна, составляющие стремечко, небольшую мышцу внутреннего уха, имеют длину всего несколько миллиметров. Миофибриллы представляют собой палочковидные субъединицы мышечных клеток.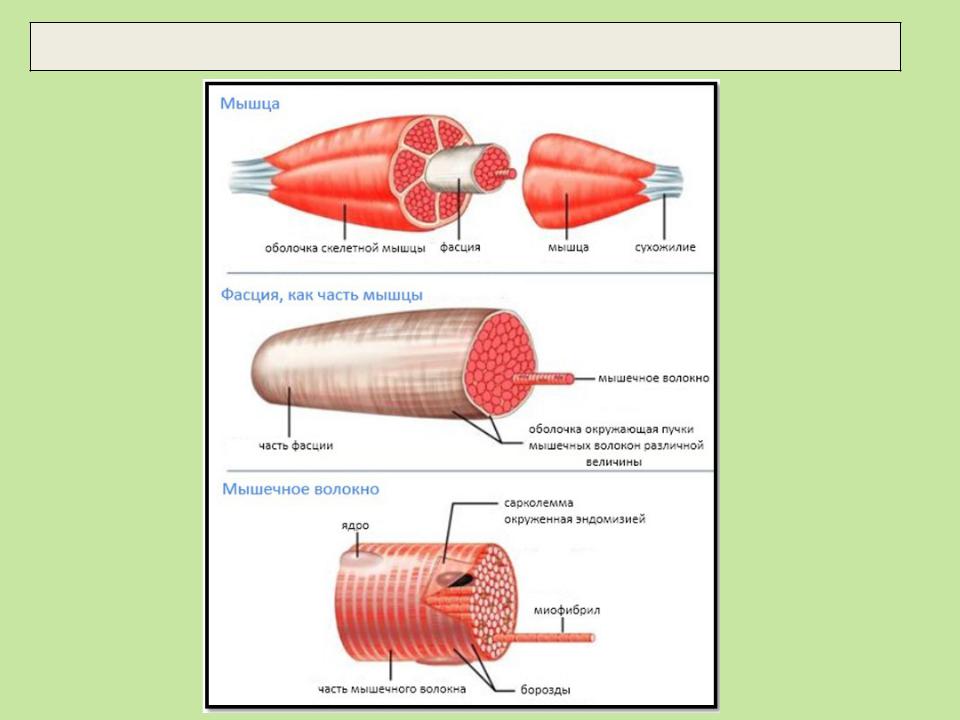 Нити актина и миозина, составляющие миофибриллы, организованы в саркомеров .Под микроскопом саркомеры придают скелетным мышцам полосатый вид.
Нити актина и миозина, составляющие миофибриллы, организованы в саркомеров .Под микроскопом саркомеры придают скелетным мышцам полосатый вид.
Ткань скелетных мышц развивается за счет слияния отдельных миобластов или ранних мышечных клеток. Это слияние приводит к характерной многоядерной структуре. Поскольку клетки слиты и многоядерны, они образуют структурный синцитий . Ядра скелетно-мышечной ткани имеют овальную форму и расположены на периферии клетки. Они сопровождаются сателлитными клетками между внешней пластинкой и сарколеммой. Клетки-сателлиты являются предшественниками клеток скелетных мышц и отвечают за способность мышечной ткани к регенерации. У них небольшое количество цитоплазмы, и из-за их расположения иногда их можно принять за ядра клеток скелетных мышц.
Волокна скелетных мышц
Скелетная мышца в разрезе. Пятно: H&E. Среднее увеличение.Структура
Ткань скелетных мышц состоит из совокупности мышечных волокон, заключенных в соединительнотканные оболочки.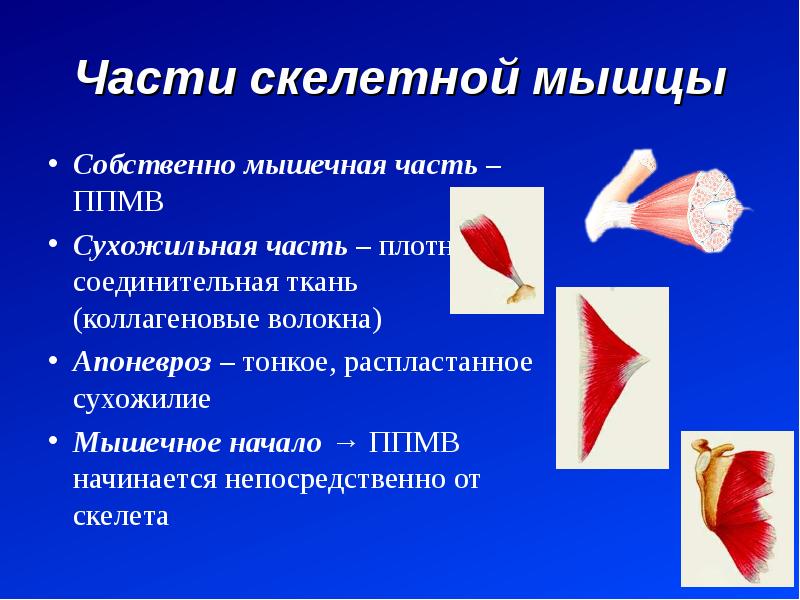 Существует три типа соединительнотканных оболочек, названных в честь их расположения. Endomysium окружает отдельные мышечные волокна. Он состоит из тонкого слоя ретикулярных волокон и пропускает только нервные волокна и капилляры небольшого диаметра, таким образом действуя как место метаболического обмена.
Существует три типа соединительнотканных оболочек, названных в честь их расположения. Endomysium окружает отдельные мышечные волокна. Он состоит из тонкого слоя ретикулярных волокон и пропускает только нервные волокна и капилляры небольшого диаметра, таким образом действуя как место метаболического обмена.
Перимизий — это немного более толстый слой соединительной ткани, состоящий в основном из коллагена I и III типов, и окружающий группу волокон. Эта группа волокон называется пучком или пучком. Fascicles — функциональные единицы ткани скелетных мышц.
Перимизий содержит кровеносные сосуды и нервные волокна немного большего размера, чем те, которые проходят через эндомизий.
Эпимизий окружает всю совокупность пучков, составляющих отдельную мышцу. Эта плотная соединительная ткань, состоящая в основном из коллагена I типа, обеспечивает сосудисто-нервное кровоснабжение мышцы.
Стартовый набор по тканям скелетных мышц ждет вас здесь.
Типы
Существует три типа волокон скелетных мышц: Тип I, Тип IIa и Тип IIb.
- Мышечные волокна типа I , также называемые медленными окислительными волокнами, специализируются на аэробной активности. Они маленькие, содержат большое количество миоглобина и кажутся красными в свежей ткани. Подергивание мышцы — это однократное сокращение мышцы. Волокна типа I составляют медленных, устойчивых к усталости двигательных единиц.Мышцы глубокой спины, отвечающие за поддержание осанки, в основном состоят из медленных окислительных волокон типа I.
- Мышечные волокна типа IIa также известны как быстрые окислительные гликолитические волокна. В свежих тканях эти волокна кажутся немного светлее, чем волокна типа I. Они содержат много митохондрий и имеют более высокое содержание миоглобина, чем волокна типа IIb.
 В отличие от волокон типа I, волокна типа IIa содержат большое количество гликогена. Благодаря этому они способны к анаэробному гликолизу и составляют быстро сокращающихся моторных единиц, устойчивых к утомлению.Волокна типа IIa более устойчивы к усталости, чем волокна типа IIb, и используются в движениях, требующих высокой постоянной мощности. Многие спортсмены имеют большое количество этих волокон, особенно пловцы.
В отличие от волокон типа I, волокна типа IIa содержат большое количество гликогена. Благодаря этому они способны к анаэробному гликолизу и составляют быстро сокращающихся моторных единиц, устойчивых к утомлению.Волокна типа IIa более устойчивы к усталости, чем волокна типа IIb, и используются в движениях, требующих высокой постоянной мощности. Многие спортсмены имеют большое количество этих волокон, особенно пловцы. - Мышечные волокна типа IIb также называют быстрыми гликолитическими волокнами. Это крупные волокна, которые в свежих тканях кажутся светло-розовыми. Волокна типа IIb содержат меньше митохондрий и меньшее количество миоглобина. Хотя они содержат низкий уровень окислительных ферментов, они демонстрируют высокую анаэробную ферментативную активность и содержат большое количество гликогена.Волокна типа IIb более склонны к утомлению, чем волокна типа I и типа IIa, и образуют быстро сокращающиеся, склонные к утомлению двигательные единицы.
Волокна типа IIb обладают самым быстрым уровнем активности АТФазы и обнаруживаются в мышцах, используемых для коротких и быстрых сокращений, таких как икроножная мышца, мышца ноги, которая используется при прыжках.
Хотите проверить свои знания о мышечной ткани? Попробуйте наши тесты по тканям и бесплатные рабочие листы по маркировке!
Саркомеры
Саркомер — функциональная единица клетки скелетных мышц.Каждый саркомер имеет длину около 2,5 микрометра. Он состоит из нескольких параллельных нитей миозина и актина . Нити актина и миозина перекрываются в определенных местах, образуя несколько полос и зон. Диск Z образует границу саркомера с обеих сторон. Тонкие актиновые нити выступают в любом направлении от Z-диска, но не пересекают всю длину саркомера. Они имеют диаметр почти 8 нм и имеют прочно связанные регуляторные белки, называемые тропонин и тропомиозин .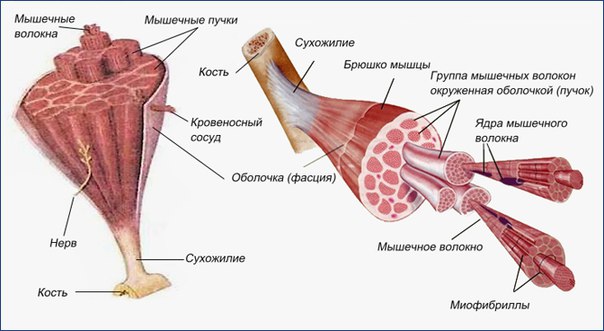
В центре саркомера отсутствуют актиновые филаменты, и он обозначен как зона H . Линия M проходит по середине зоны H перпендикулярно нитям. Между актиновыми нитями находятся толстые миозиновые нити. Их диаметр составляет приблизительно 15 нм с областью глобулярной головки, состоящей из тяжелой и легкой цепи. Эта головка обладает активностью АТФазы и способностью связываться и перемещаться по актиновой нити. Они не связаны с дисками Z, но пересекают зону H.
Саркомер разделен на три полосы. Полоса A находится посередине и соответствует филаментам миозина вместе с тонкими филаментами, перекрывающимися на обоих концах. Есть две полосы I по обе стороны от полосы А и представляют область, в которой присутствуют только актиновые филаменты.
Саркоплазматические структуры
- Sarcolemma — клеточная плазматическая мембрана мышечной клетки
- Саркоплазматический ретикулум — модифицированный эндоплазматический ретикулум.
 Саркоплазматическая сеть напоминает кусок кружева, окружающий миофибриллы. Он состоит из сети канальцев с резервуарами на обоих концах. Этот резервуар называется терминальной цистерной.
Саркоплазматическая сеть напоминает кусок кружева, окружающий миофибриллы. Он состоит из сети канальцев с резервуарами на обоих концах. Этот резервуар называется терминальной цистерной. - Терминальная цистерна — увеличенные области на обоих концах саркоплазматической сети. Концевые цистерны изолируют кальций, который используется в цикле сокращения. По обе стороны от t-канальца находятся терминальные цистерны. Все вместе они называются триадой.
- Трубочки — также называемые поперечными канальцами. Т-канальцы — это углубления на поверхности мышечных клеток. Их стенки продолжаются сарколеммой, то есть внутренняя поверхность t-канальца открыта для внеклеточного матрикса. Т-канальцы ответственны за перемещение потенциалов действия во внутреннюю область мышечной клетки.
Дополнительные белки
Толстые и тонкие филаменты в миофибриллах поддерживаются дополнительными белками. Эти белки поддерживают скорость и выравнивание нитей во время цикла сокращения. Некоторые из них описаны ниже.
Эти белки поддерживают скорость и выравнивание нитей во время цикла сокращения. Некоторые из них описаны ниже.
- Титин — это большой эластичный белок, который прикрепляет толстые волокна к Z-линиям, предотвращая чрезмерное растяжение миофибриллы
- Тропомодулин действует как актиновый колпачок. Он прикрепляется к свободному концу актиновой нити, чтобы сохранить ее длину.
- α-Актинин представляет собой короткий палочковидный белок, который собирает тонкие волокна в параллельные пучки и прикрепляет их к линии Z
- Desmin — это промежуточная нить, которая образует решетку, окружающую саркомер около Z-линий, чтобы прикрепить их друг к другу и к плазматической мембране.
- Небулин представляет собой тонкий вытянутый белок, идущий параллельно тонким филаментам. Он помогает альфа-актинину связывать тонкие волокна с Z-линией и считается важным при развитии мышечной ткани.
- Считается, что дистрофин связывает актиновые филаменты с внешней пластиной мышечной клетки
- Myomesin прикрепляет толстые волокна к М-линии.
Нервно-мышечный переход
Нервно-мышечное соединение, сокращенно НМС, — это место, в котором окончание двигательного нерва образует синапс с мышечной клеткой.Моторные нейроны высвобождают ацетилхолин в синаптическую щель нервно-мышечного соединения. Молекулы ацетилхолина проходят через синаптическую щель и связываются с рецепторами на мембране мышечных волокон. Это приводит к деполяризации и последующему сокращению мышечной клетки.
Сокращение
Механизм сокращения мышц упоминается как теория скользящих волокон . В этом процессе нити актина и миозина скользят друг мимо друга, укорачивая саркомер.Механизм скользящего филамента требует гидролиза молекул аденозинтрифосфата (АТФ) и связывания и высвобождения миозиновых головок на актиновых филаментах.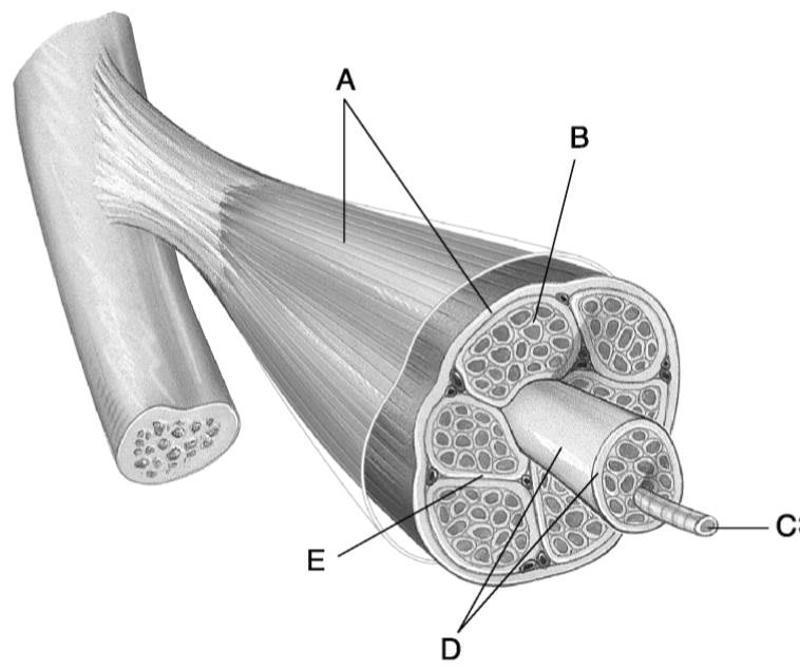
- После возбуждения двигательным нервным окончанием АТФ связывается с миозином, уменьшая сродство актина к миозину. Миозин высвобождает актин.
- Миозин гидролизует АТФ до аденозиндифосфата (АДФ) и молекулы фосфата. Это обеспечивает энергию, необходимую для поворота миозиновой головки на угол 90 ° и присоединения к новому актину.
- Силовой удар — Когда мышца не сокращается, сайт связывания миозина частично блокируется молекулой тропонина. Во время сокращения кальций связывается с тропонином, уменьшая его сродство к сайту связывания миозина и раскрывая его, так что миозин связывается более плотно и высвобождает фосфат. Высвобождение фосфата заставляет миозиновую головку качаться к линии М, и прикрепленная актиновая нить скользит вместе с ней.
- В конце силового удара миозин высвобождает молекулу АДФ.Головка возвращается в исходное положение и снова готова начать цикл.
Контроль сокращения скелетных мышц
Два основных проприорецептора влияют на силу и продолжительность мышечного сокращения: органы сухожилия Гольджи и волокна мышечного веретена.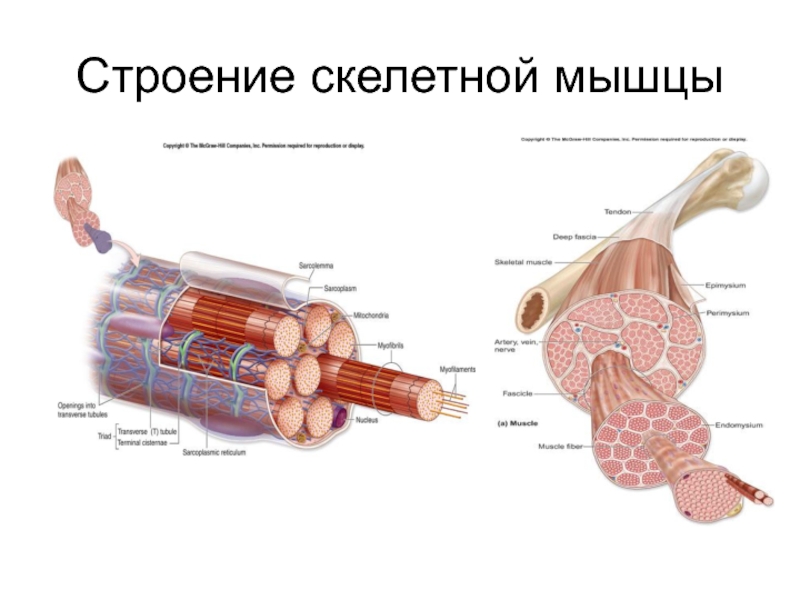 Органы сухожилий Гольджи передают информацию о силе сокращения. Мышечные веретена обнаруживают изменения длины мышцы.
Органы сухожилий Гольджи передают информацию о силе сокращения. Мышечные веретена обнаруживают изменения длины мышцы.
Органы сухожилия Гольджи
Органы сухожилия Гольджи — это рецепторы, обнаруженные на стыке между миофибрилл и сухожилий.Рецепторные окончания органа сухожилия Гольджи переплетаются с коллагеновыми волокнами сухожилия. Когда мышца сокращается, сухожилие растягивается, а рецепторы сжимаются в паутине соединительной ткани. Давление инициирует потенциал действия в нервном окончании и посылает сигнал через центральную нервную систему. Роль органа сухожилия Гольджи заключается в предотвращении чрезмерного сокращения мышц.
Волокна мышечного веретена
Мышечные веретена обнаруживают изменение длины, положения и скорости мышц.Волокна мышечного веретена представляют собой длинные тонкие инкапсулированные волокна, расположенные параллельно миофибриллам. Их также называют интрафузионных мышечных волокон, , в то время как экстрафузальные волокна присутствуют вне капсулы веретена. Концы интрафузальных мышечных волокон прикрепляются к миофибриллам, поэтому при растяжении мышцы растягиваются и волокна мышечного веретена. Растяжение веретена заставляет его деполяризоваться и передавать эту информацию через спинной мозг.
Концы интрафузальных мышечных волокон прикрепляются к миофибриллам, поэтому при растяжении мышцы растягиваются и волокна мышечного веретена. Растяжение веретена заставляет его деполяризоваться и передавать эту информацию через спинной мозг.
Клинические корреляции
Мышечная дистрофия
Мышечная дистрофия — это общий термин для ряда заболеваний, включающих мутации в ДНК, кодирующей белки дистрофина.Поврежденный дистрофин приводит к разрушению и ослаблению мышечной ткани. Некоторые примеры включают мышечную дистрофию Дюшенна и Беккера и фациально-плечевую мышечную дистрофию. Несмотря на наличие генетической связи, расстройства имеют широкий спектр фенотипических проявлений и требуют множества анализов для постановки правильного диагноза.
Миопатия с совокупным актином
Миопатия с совокупным актином, также называемая миопатией с накоплением актина, представляет собой заболевание, при котором актиновых филаментов накапливаются в волокнах скелетных мышц. Это вызывает сильную мышечную слабость и снижение мышечного тонуса. Люди с миопатией актиновых агрегатов часто не выживают в младенчестве, потому что диафрагма, основная мышца, используемая при дыхании, не может функционировать. Выжившие люди, как правило, имеют плохую осанку, мелкую моторику и трудности при ходьбе.
Это вызывает сильную мышечную слабость и снижение мышечного тонуса. Люди с миопатией актиновых агрегатов часто не выживают в младенчестве, потому что диафрагма, основная мышца, используемая при дыхании, не может функционировать. Выжившие люди, как правило, имеют плохую осанку, мелкую моторику и трудности при ходьбе.
Миотубулярные (центроядерные) миопатии
Миотубулярная миопатия, генетическое заболевание, вызванное мутацией в протеине динамин , характеризуется большими центральными ядрами.Ядра окружены областью, плотно заполненной митохондриями, но без миофибрилл. Образующиеся мышечные волокна обычно имеют небольшие размеры и округлую форму. У пациентов с миотубулярной миопатией часто наблюдается паралич экстраокулярных мышц.
Гистология скелетных мышц: хотите узнать об этом больше?
Наши увлекательные видео, интерактивные викторины, подробные статьи и HD-атлас помогут вам быстрее достичь лучших результатов.
С чем вы предпочитаете учиться?
«Я бы честно сказал, что Kenhub сократил мое учебное время вдвое.” — Прочитайте больше. Ким Бенгочеа, Университет Реджиса, Денвер
Скелетные мышцы — ветеринарная гистология
Функция
Одна из важнейших функций скелетных мышц — сокращаться и вызывать движение. Скелетные мышцы действуют не только для движения, но и для остановки движения. Постоянные небольшие сокращения скелетных мышц необходимы, чтобы удерживать тело в вертикальном положении в любом положении, даже в состоянии покоя. Мышцы также предотвращают чрезмерное движение костей и суставов, поддерживая стабильность скелета и предотвращая повреждение или деформацию скелетных структур.Скелетные мышцы расположены по всему телу у отверстий внутренних путей, чтобы контролировать движение различных веществ. Эта мышца позволяет произвольно контролировать определенные функции, такие как глотание, мочеиспускание и дефекация. Скелетные мышцы также защищают внутренние органы (особенно органы брюшной полости), выступая в качестве внешнего барьера или щита от внешних травм и поддерживая вес органов.
Скелетные мышцы способствуют выделению тепла, в основном за счет дрожи или интенсивных движений.Для сокращения мышц требуется энергия, а при расщеплении АТФ выделяется тепло. Это тепло очень заметно во время и после интенсивных активных движений, таких как бег. В этом случае длительное движение мышц вызывает повышение температуры тела. Точно так же, когда температура окружающей среды низкая, случайные скелетные мышцы вокруг аппендикулярного скелета вызывают случайные сокращения скелетных мышц, генерируя тепло (дрожь).
Структура
Каждая скелетная мышца — это орган, состоящий из различных интегрированных тканей.Эти ткани включают волокна скелетных мышц, кровеносные сосуды, нервные волокна и соединительную ткань. Каждая скелетная мышца имеет три уровня соединительной ткани, которые ее окружают. Эти уровни соединительной ткани обеспечивают структуру мышцы в целом, а также разделяют мышечные волокна внутри мышцы на группы. Каждая большая мышца обернута оболочкой из плотной соединительной ткани неправильной формы, называемой эпимизием , которая удерживает мышечные пучки в группе, а также позволяет мышце сокращаться, сохраняя при этом ее структурную целостность.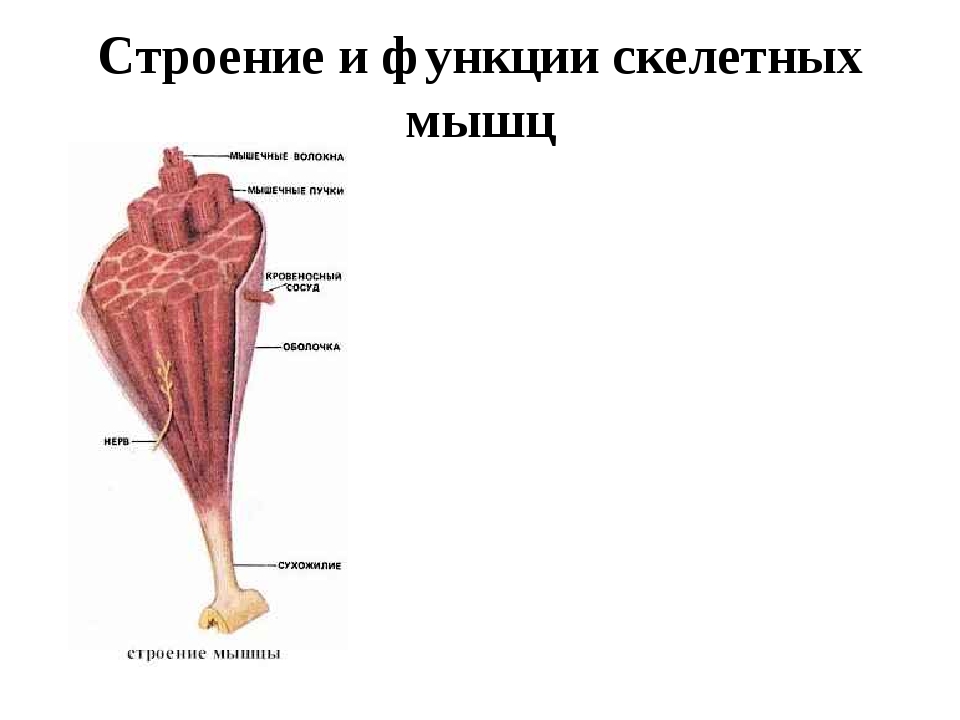 Это самый внешний слой. Эпимизий также позволяет мышце двигаться независимо от других групп мышц и других типов тканей. Большинство отдельных мышц, которые вы изучаете при макроанатомии, окружены эпимизием.
Это самый внешний слой. Эпимизий также позволяет мышце двигаться независимо от других групп мышц и других типов тканей. Большинство отдельных мышц, которые вы изучаете при макроанатомии, окружены эпимизием.
Мышечное волокно — термин, синоним мышечной клетки. Мышечное волокно — это клетка с множеством периферически расположенных ядер, которая проходит на ВСЕЙ длине реальной мышцы. Внутри каждой скелетной мышцы мышечные волокна организованы в отдельные пучки (называемые пучками ) с промежуточным слоем соединительной ткани, называемым перимизием .Внутри каждого пучка каждое мышечное волокно заключено в чрезвычайно тонкий слой соединительной ткани, называемый эндомизием .
Некоторая терминология, связанная с мышечными волокнами, восходит к греческому sarco , что означает «плоть». Плазматическая мембрана мышечных волокон называется сарколеммой , цитоплазма называется саркоплазмой , а специальный гладкий эндоплазматический ретикулум, который хранит и контролирует высвобождение ионов кальция (Ca ++), называется саркоплазматическим ретикулумом (SR). .Функциональной единицей волокна скелетных мышц является саркомер, высокоорганизованная структура сократительных миофиламентов , актина (тонкая нить) и миозина (толстая нить), а также других поддерживающих белков. Отдельные саркомеры соединяются в длинную цепочку, называемую миофибриллой , и прикрепляются к сарколемме на своем конце. Миофибриллы связаны вместе в саркоплазме. Кроме того, есть инвагинации сарколеммы в саркоплазму, окружающую миофибриллы.Они называются Т-канальцами. Система t-канальцев передает потенциал действия от сарколеммы в клетку, чтобы стимулировать миофибриллы.
.Функциональной единицей волокна скелетных мышц является саркомер, высокоорганизованная структура сократительных миофиламентов , актина (тонкая нить) и миозина (толстая нить), а также других поддерживающих белков. Отдельные саркомеры соединяются в длинную цепочку, называемую миофибриллой , и прикрепляются к сарколемме на своем конце. Миофибриллы связаны вместе в саркоплазме. Кроме того, есть инвагинации сарколеммы в саркоплазму, окружающую миофибриллы.Они называются Т-канальцами. Система t-канальцев передает потенциал действия от сарколеммы в клетку, чтобы стимулировать миофибриллы.
Отчетливые линии или полосы на саркомере видны там, где миозиновые и актиновые нити перекрываются, а также там, где саркомеры последовательно соединяются друг с другом. Это то, что придает скелетным мышцам полосатый вид при гистологии. Фактически, скелетная мышца и сердечная мышца (поскольку сердечная мышца также имеет саркомеры) также известны как поперечно-полосатая мышца. Когда саркомеры и, следовательно, миофибриллы сокращаются, сокращается вся мышечная клетка. Поскольку актин и его комплекс тропонин-тропомиозин (выступающий от Z-дисков к центру саркомера) образуют нити, которые тоньше миозина, вместе они называются тонкой нитью саркомера. Поскольку нити миозина и их многочисленные головки (выступающие от центра саркомера к Z-дискам, но не полностью к ним) имеют большую массу и толще, их называют нитью толщиной s .Как упоминалось ранее, для сокращения саркомера кальций высвобождается из саркоплазматической сети после того, как генерируется потенциал стимулирующего действия из Т-канальцев. Кальций взаимодействует с молекулами актиновых нитей, тропонина и тропомиозина, которые обычно блокируют сайты связывания миозина на актине. Связывание АТФ с миозином вызывает высвобождение миозиновой головки из актина.
Когда саркомеры и, следовательно, миофибриллы сокращаются, сокращается вся мышечная клетка. Поскольку актин и его комплекс тропонин-тропомиозин (выступающий от Z-дисков к центру саркомера) образуют нити, которые тоньше миозина, вместе они называются тонкой нитью саркомера. Поскольку нити миозина и их многочисленные головки (выступающие от центра саркомера к Z-дискам, но не полностью к ним) имеют большую массу и толще, их называют нитью толщиной s .Как упоминалось ранее, для сокращения саркомера кальций высвобождается из саркоплазматической сети после того, как генерируется потенциал стимулирующего действия из Т-канальцев. Кальций взаимодействует с молекулами актиновых нитей, тропонина и тропомиозина, которые обычно блокируют сайты связывания миозина на актине. Связывание АТФ с миозином вызывает высвобождение миозиновой головки из актина.
Сокращение
Сокращение саркомера включает несколько этапов. Чтобы понять этот шаг, вы должны понимать, что миозиновая головка имеет несколько важных характеристик.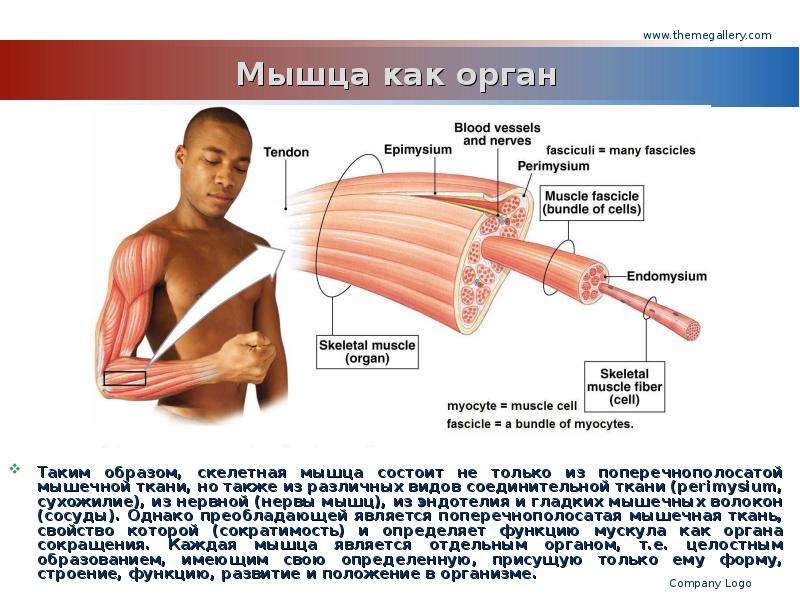
- Он имеет АТФ-связывающие сайты, в которые помещаются молекулы АТФ. АТФ представляет собой потенциальную энергию.
- Он имеет сайты связывания актина, в которые помещаются молекулы актина.
- У него есть «петля» в месте выхода из ядра толстой миофиламента. Это позволяет голове раскачиваться вперед и назад, и это движение вызывает сокращение мышц.
Во время сжатия происходят следующие шаги:
- Активированная миозиновая головка прикрепляется к сайту связывания актина; сокращение относится к активации поперечных мостиков миозина — сайтов, которые генерируют силу
- Фосфатная группа P на миозиновой головке отваливается
- Затем миозиновая головка производит силовой удар, который приводит к скольжению нитей, заставляя тонкие нити двигаться навстречу друг другу.АДФ на миозине тоже отваливается.
- Как только силовой удар завершен, АТФ снова прикрепляется к миозиновой головке, заставляя головку отделяться от актинового участка и возвращаться в исходное положение.
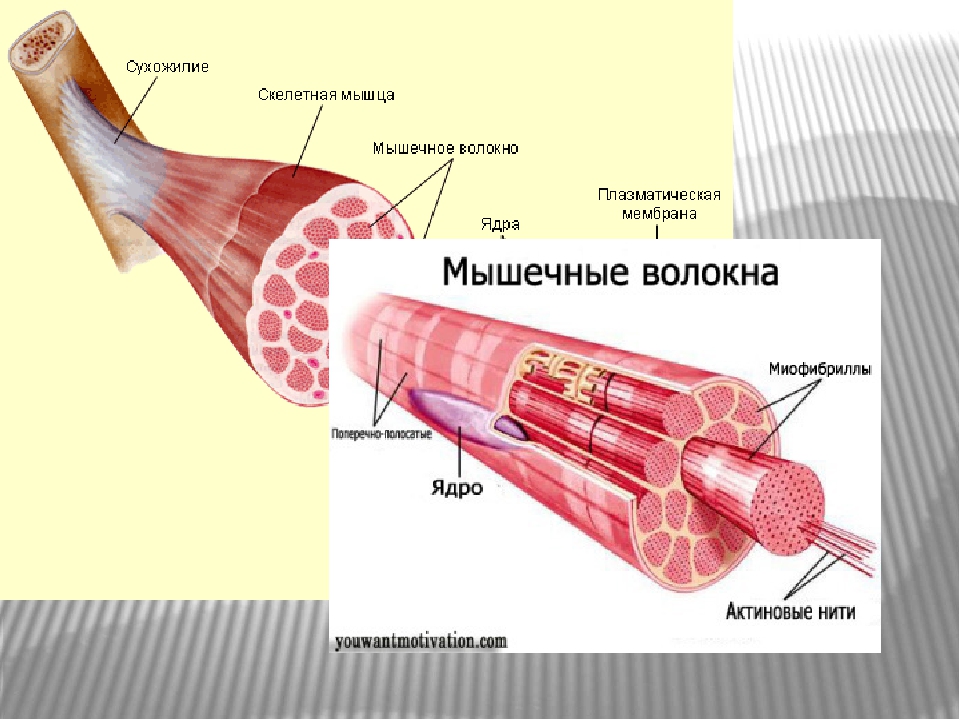
- АТФ, прикрепленный к головке миозина, расщепляется АТФазой, заставляя головки миозина снова активироваться; цикл может повторяться снова и снова, пока присутствуют кальций и АТФ. Для полного сокращения мышц часто требуется несколько циклов этого процесса.
РИСУНОК : Скелетная мышца
Эпимизий — обзор | Темы ScienceDirect
Микроскопический вид
Мышечные волокна или клетки обычно проходят от сухожилия к сухожилию в мышце и не разветвляются и не образуют синцитии.В поперечном сечении волокна в мышцах взрослых людей имеют многоугольную или многогранную форму. На диаметр мышечных волокон влияют многие факторы, такие как вид, порода, возраст, вес, пол, уровень питания, положение и функция мышцы, а также упражнения. Измерения волокон в отдельных мышцах покажут различия в размере волокон, которые будут отражены в виде колоколообразной кривой на гистограмме. Различия в размере волокон у разных видов не связаны напрямую с массой тела (свинья> лошадь, корова, кролик> овца). Размер клетчатки у самцов больше, чем у самок, и имеет тенденцию увеличиваться с возрастом к зрелости.
Размер клетчатки у самцов больше, чем у самок, и имеет тенденцию увеличиваться с возрастом к зрелости.
Клеточные особенности волокон скелетных мышц лучше всего оцениваются на продольных срезах. Волокна ограничены плазматической мембраной или сарколеммой, которая покрыта внешней пластинкой (окрашенной периодической кислотной реакцией Шиффа). Тонкие удлиненные ядра обычно располагаются под сарколеммой в виде спирали на расстоянии 10-50 мкм друг от друга. В мышечно-сухожильных соединениях мышечные волокна имеют множество центрально расположенных ядер.Ядра сателлитных клеток расположены между сарколеммой и внешней пластинкой. Волокна содержат сотни продольно выровненных миофибрилл, состоящих из повторяющихся саркомеров. Характерная поперечная исчерченность волокон скелетных мышц является результатом параллельного расположения полос в соседних миофибриллах. Самые большие полосы, называемые в соответствии с их внешним видом в поляризованном свете, — это полосы A (анизотропные или двулучепреломляющие, кажутся яркими) и полосы I (иостропные, кажутся темными).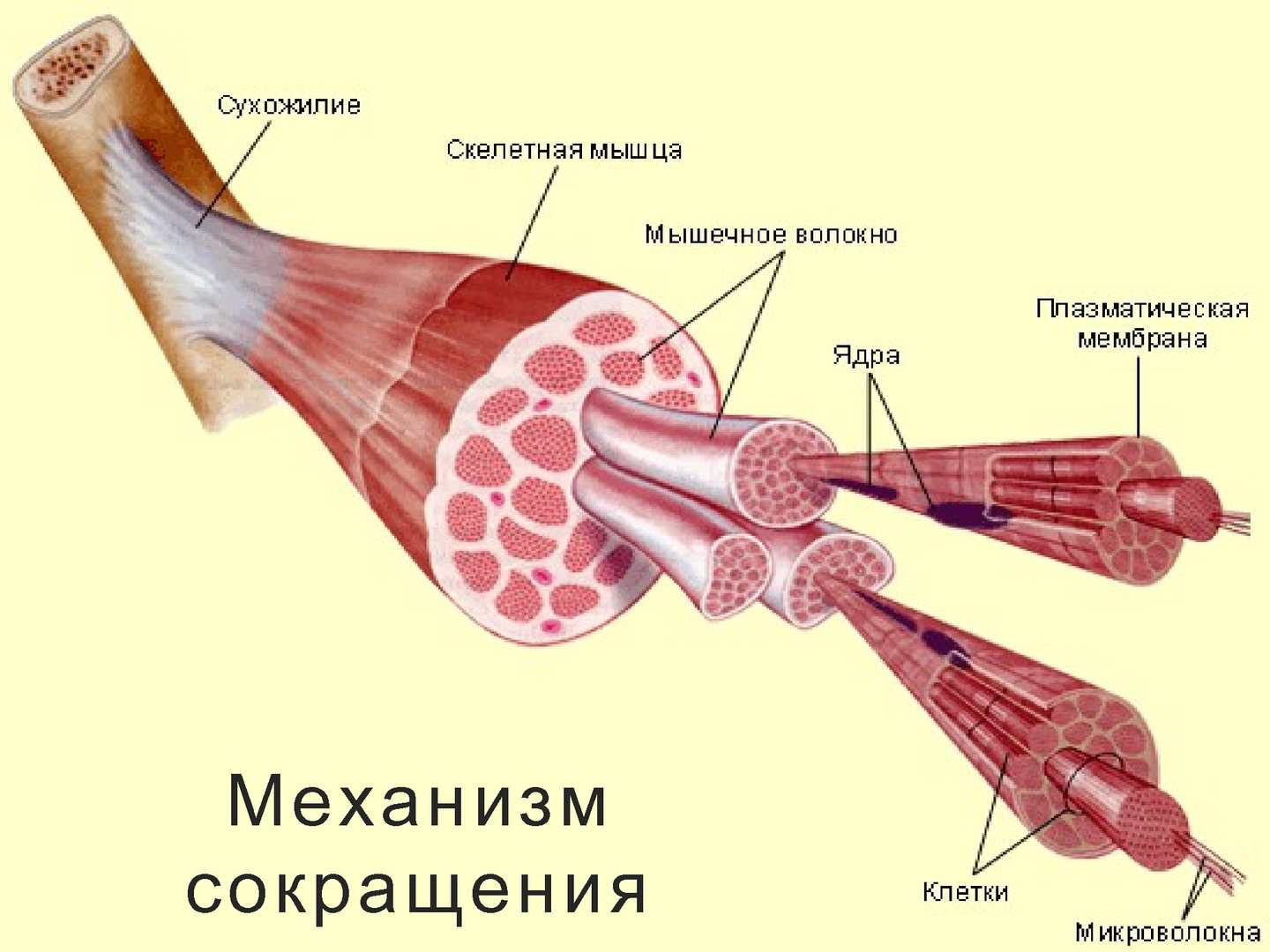 I-полосы, состоящие из тонких миофиламентов, разделены пополам Z-линиями (дисками, полосами), которые образуют конец каждого саркомера; A-полосы, состоящие из толстых волокон, разделены пополам H-полосами с меньшим двулучепреломлением.Картина полос, названная в честь появления в поляризованном свете, меняется на противоположную при исследовании световой микроскопией с фазово-контрастной оптикой, световой микроскопией с обычной оптикой на срезах, окрашенных обычными катионными красителями, или просвечивающей электронной микроскопией.
I-полосы, состоящие из тонких миофиламентов, разделены пополам Z-линиями (дисками, полосами), которые образуют конец каждого саркомера; A-полосы, состоящие из толстых волокон, разделены пополам H-полосами с меньшим двулучепреломлением.Картина полос, названная в честь появления в поляризованном свете, меняется на противоположную при исследовании световой микроскопией с фазово-контрастной оптикой, световой микроскопией с обычной оптикой на срезах, окрашенных обычными катионными красителями, или просвечивающей электронной микроскопией.
Нанесение гистохимических красителей, таких как АТФаза или NADH-TR, на замороженные срезы скелетных мышц позволит продемонстрировать различные популяции волокон, которые нельзя различить на залитых парафином срезах, окрашенных обычными красителями, такими как гематоксилин и эозин; однако типы волокон распознаются ультраструктурными исследованиями.Гистохимическая уникальность этих типов волокон коррелирует с различиями в их физиологических характеристиках, таких как скорость сокращения и утомляемость; их биохимическая и метаболическая активность; их грубый цвет; и их структура, выявленная ультраструктурно. Таблица 46.5 суммирует эти функции.
Таблица 46.5 суммирует эти функции.
ТАБЛИЦА 46.5. Характеристики основных типов волокон скелетных мышц млекопитающих
| Тип I | Тип IIA | Тип IIB | |
|---|---|---|---|
| Морфологические характеристики: | Темный цвет | Бледный | |
| Содержание гликогена | Низкое | Высокое | Высокое |
| Содержание миоглобина | Высокое | Высокое | Низкое |
| Липидные глобулы | Многочисленные | Многочисленные | Мало |
| Митохрондриальное содержание | Высокий | Промежуточный | Низкий |
| Физиологические характеристики: | |||
| Скорость подергивания | Медленно | Быстро | Быстро |
| Утомляемость | устойчивыйУстойчивый | Восприимчивый |
Таблица воспроизведена из Основы токсикологической патологии , 2-е изд. В. М. Хашек, К. Г. Руссо и М. А. Валлиг, ред. (2010) Academic Press, Таблица 12.9, стр. 366, с разрешения
В. М. Хашек, К. Г. Руссо и М. А. Валлиг, ред. (2010) Academic Press, Таблица 12.9, стр. 366, с разрешения
Большинство мышц будет иметь смесь всех типов волокон, чтобы создать так называемый шахматный узор дифференциального гистохимического окрашивания. Волокна, иннервируемые одними и теми же нервами, будут иметь один и тот же тип волокон, а реиннервируемые волокна могут показывать обратные типы волокон. В некоторых мышцах будет преобладать один тип волокон, например, камбаловидная мышца (красная мышца с высоким содержанием волокон типа I и способная к длительному действию или переносу веса) и икроножная мышца (белая мышца с высоким содержанием волокон типа II и способная к внезапному действию и целенаправленное движение).Пропорции различных типов волокон могут различаться в зависимости от вида, породы, возраста, физических упражнений и некоторых мышечных заболеваний.
Ультраструктурный вид
Поверхность волокна покрыта плазматической мембраной (сарколеммой) и внешней пластинкой.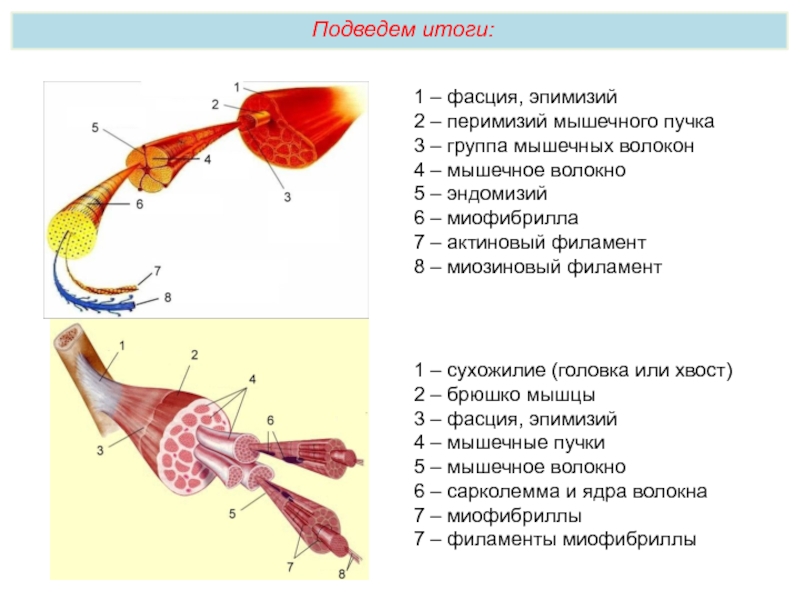 Удлиненные субсарколеммальные ядра окружены скоплением митохондрий, липидных капель, гранул гликогена, элементов саркоплазматической сети и аппарата Гольджи. Волокно содержит обильный сократительный материал, который состоит из миофибрилл из 0.Диаметр 5–1,0 мкм. Миофибриллы состоят из повторяющихся единиц, называемых саркомерами, длиной 2–3 мкм. Саркомеры заканчиваются плотными Z-линиями, которые содержат αα-актинин, актин и тропомиозин. Тонкие миофиламенты диаметром 60 Å, содержащие актин, тропонин и тропомиозин, проходят по обе стороны от Z-линии, образуя I-полосы. Средняя половина саркомера содержит толстые (диаметром 160 Å) миофиламенты, которые состоят из миозина и пересекаются с соседними тонкими волокнами. Центр саркомера, состоящий только из толстых нитей, представляет собой полосу H и делится пополам относительно плотной линией M.Сокращение волокон приводит к укорачиванию саркомеров из-за скольжения толстых и тонких волокон друг по другу, что приводит к сужению полос I и H. Поперечные сечения миофибрилл имеют различный вид в зависимости от расположения в саркомере, но края полосы А будут иметь толстые нити, окруженные гексагональной сеткой тонких нитей.
Удлиненные субсарколеммальные ядра окружены скоплением митохондрий, липидных капель, гранул гликогена, элементов саркоплазматической сети и аппарата Гольджи. Волокно содержит обильный сократительный материал, который состоит из миофибрилл из 0.Диаметр 5–1,0 мкм. Миофибриллы состоят из повторяющихся единиц, называемых саркомерами, длиной 2–3 мкм. Саркомеры заканчиваются плотными Z-линиями, которые содержат αα-актинин, актин и тропомиозин. Тонкие миофиламенты диаметром 60 Å, содержащие актин, тропонин и тропомиозин, проходят по обе стороны от Z-линии, образуя I-полосы. Средняя половина саркомера содержит толстые (диаметром 160 Å) миофиламенты, которые состоят из миозина и пересекаются с соседними тонкими волокнами. Центр саркомера, состоящий только из толстых нитей, представляет собой полосу H и делится пополам относительно плотной линией M.Сокращение волокон приводит к укорачиванию саркомеров из-за скольжения толстых и тонких волокон друг по другу, что приводит к сужению полос I и H. Поперечные сечения миофибрилл имеют различный вид в зависимости от расположения в саркомере, но края полосы А будут иметь толстые нити, окруженные гексагональной сеткой тонких нитей. Саркоплазма, окружающая миофибриллы, содержит элементы системы поперечных (Т) канальцев и саркоплазматического ретикулума (SR), митохондрии, липидные капли, гранулы гликогена и цитозоль.Т-канальцы представляют собой инвагинации сарколеммы, которые часто видны на краю I-образной полосы с двумя соседними элементами SR, образующими «триаду». Трубчатая система функционирует как канал, позволяющий быстро распространять электрический импульс от моторной концевой пластинки к остальной части волокна, чтобы вызвать высвобождение кальция, хранящегося в SR, с последующим связыванием с регуляторными белками и взаимодействием актина и миозина. чтобы начать сокращение.
Саркоплазма, окружающая миофибриллы, содержит элементы системы поперечных (Т) канальцев и саркоплазматического ретикулума (SR), митохондрии, липидные капли, гранулы гликогена и цитозоль.Т-канальцы представляют собой инвагинации сарколеммы, которые часто видны на краю I-образной полосы с двумя соседними элементами SR, образующими «триаду». Трубчатая система функционирует как канал, позволяющий быстро распространять электрический импульс от моторной концевой пластинки к остальной части волокна, чтобы вызвать высвобождение кальция, хранящегося в SR, с последующим связыванием с регуляторными белками и взаимодействием актина и миозина. чтобы начать сокращение.
Физиология человека — мышцы
Физиология человека — мышцыНервная система «общается» с мышцами через нервно-мышечные (также называемые мионевральными) соединениями.Эти соединения (Рисунок 1) работают очень похоже на синапс между нейронами (видео нервно-мышечного соединения). Другими словами:
- импульс поступает на наконечник колбы,
- химический медиатор высвобождается из пузырьков (каждая из которых содержит 5000-10 000 молекул ацетилхолина) и распространяется через нервно-мышечные расщелина,
- молекулы передатчика заполняют рецепторные участки в мембране мышцы и увеличивают проницаемость мембраны для натрия,
- натрий диффундирует внутрь и мембранный потенциал становится менее отрицательным,
- и, если пороговый потенциал достигнут, возникает потенциал действия,
импульс проходит по мембране мышечной клетки, и мышца сокращается.
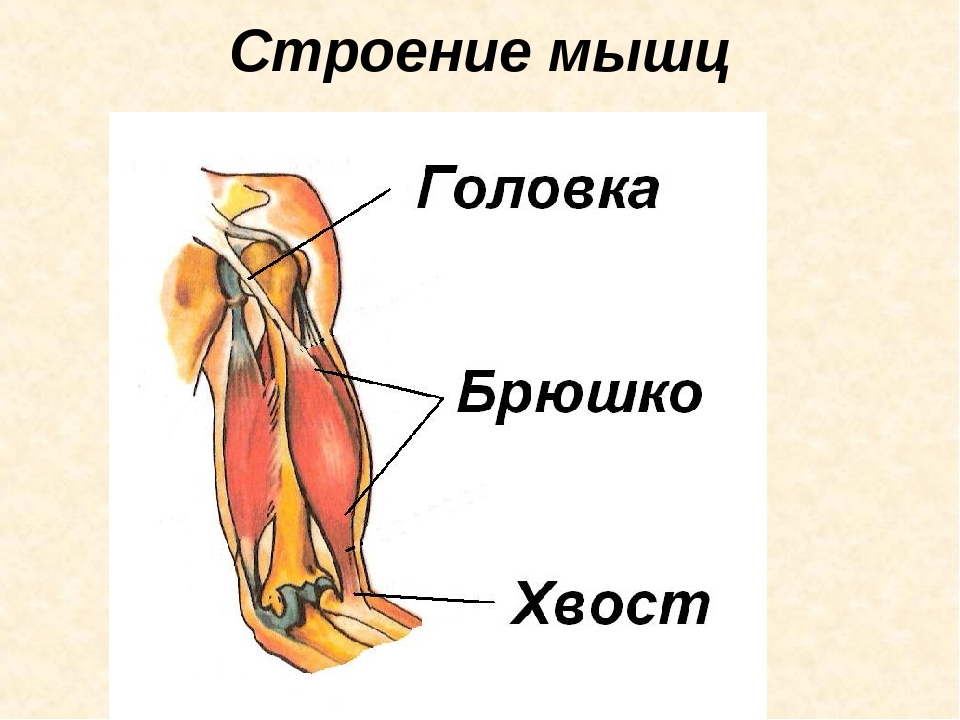
Некоторые мышцы (скелетные) не будут сокращаться без стимуляции.
нейронами; другие мышцы (некоторые гладкие и сердечные мышцы) сокращаются без
нервное возбуждение, но на их сокращения также могут влиять нервные
система. Таким образом, нервная и мышечная системы тесно взаимосвязаны.
Давайте теперь сосредоточимся на мышцах — какова их структура и как она работает.
Изображение нервно-мышечного соединения в большом увеличении (Hirsch 2007).
Характеристики мышцы:
- возбудимость — реагирует на раздражители (например, нервные импульсы)
- сократимость — возможность укорочения
- растяжимость — растягивается при вытягивании
Функции мышцы:
- движение
- поддержание осанки
- выработка тепла (например, дрожь при терморегуляции)
Типы мышц:
- скелет:
- прикреплен к костям и перемещает скелет
- также называется поперечно-полосатой мышцей (из-за его внешнего вида под микроскопом, как показано на фотографии слева)
- произвольная мышца
- гладкая (фото справа)
- Непроизвольная мышца
- мышца внутренних органов (e.
 g., в стенках кровеносных сосудов, кишечника и других «полых» структур и органов тела)
g., в стенках кровеносных сосудов, кишечника и других «полых» структур и органов тела) - сердечный:
- мышца сердца
- принудительное
Структура скелетных мышц
Структура скелетных мышц:
Скелетные мышцы обычно прикрепляются к кости сухожилиями, состоящими из соединительной ткани. Эта соединительная ткань также покрывает всю мышцу и называется эпимизием. Скелетные мышцы состоят из множества субъединиц или пучков, называемых фасиклами (или пучками). Фасцикулы также окружены соединительной тканью (называемой перимизием), и каждый пучок состоит из многочисленных мышечных волокон (или мышечных клеток; мышечная часть 1). Мышечные клетки, заключенные в оболочку эндомизия, состоят из множества фибрилл (или миофибрилл), и эти миофибриллы состоят из длинных белковых молекул, называемых миофиламентами. В миофибриллах есть два типа миофиламентов: толстые миофиламенты и тонкие миофиламенты.
Эта соединительная ткань также покрывает всю мышцу и называется эпимизием. Скелетные мышцы состоят из множества субъединиц или пучков, называемых фасиклами (или пучками). Фасцикулы также окружены соединительной тканью (называемой перимизием), и каждый пучок состоит из многочисленных мышечных волокон (или мышечных клеток; мышечная часть 1). Мышечные клетки, заключенные в оболочку эндомизия, состоят из множества фибрилл (или миофибрилл), и эти миофибриллы состоят из длинных белковых молекул, называемых миофиламентами. В миофибриллах есть два типа миофиламентов: толстые миофиламенты и тонкие миофиламенты.
Источник: Википедия.
Скелетные мышцы значительно различаются по размеру, форме и расположению волокон. Они варьируются от очень крошечных нитей, таких как стремечковая мышца среднего уха, до больших масс, таких как мышцы бедра. Скелетные мышцы могут состоять из сотен или даже тысяч мышечных волокон, связанных вместе и завернутых в соединительнотканный покров. |
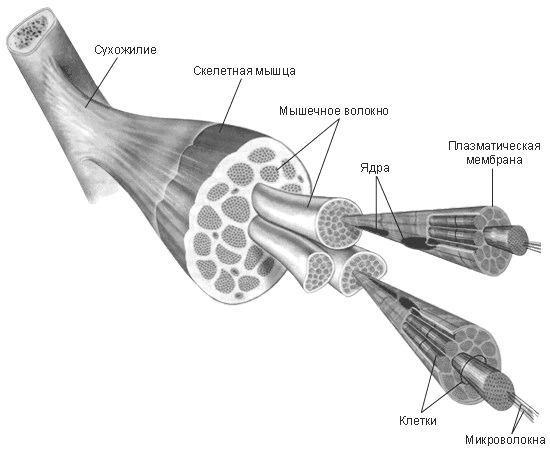 Каждая мышца окружена соединительнотканной оболочкой, называемой эпимизием.Фасция, соединительная ткань вне эпимизия, окружает и разделяет мышцы. Части эпимизия выступают внутрь, чтобы разделить мышцу на части. Каждый отсек содержит пучок мышечных волокон. Каждый пучок мышечных волокон называется пучком и окружен слоем соединительной ткани, называемым перимизием. Внутри пучка каждая отдельная мышечная клетка, называемая мышечным волокном, окружена соединительной тканью, называемой эндомизием. Скелетные мышцы имеют обильное снабжение кровеносных сосудов и нервов.Прежде чем скелетное мышечное волокно сможет сократиться, оно должно получить импульс от нейрона. Обычно артерия и по крайней мере одна вена сопровождают каждый нерв, который проникает в эпимизий скелетной мышцы. Ветви нерва и кровеносные сосуды следуют за соединительнотканными компонентами мышцы нервной клетки и с одним или несколькими мельчайшими кровеносными сосудами, называемыми капиллярами (Источник: training.seer.cancer.gov).
Каждая мышца окружена соединительнотканной оболочкой, называемой эпимизием.Фасция, соединительная ткань вне эпимизия, окружает и разделяет мышцы. Части эпимизия выступают внутрь, чтобы разделить мышцу на части. Каждый отсек содержит пучок мышечных волокон. Каждый пучок мышечных волокон называется пучком и окружен слоем соединительной ткани, называемым перимизием. Внутри пучка каждая отдельная мышечная клетка, называемая мышечным волокном, окружена соединительной тканью, называемой эндомизием. Скелетные мышцы имеют обильное снабжение кровеносных сосудов и нервов.Прежде чем скелетное мышечное волокно сможет сократиться, оно должно получить импульс от нейрона. Обычно артерия и по крайней мере одна вена сопровождают каждый нерв, который проникает в эпимизий скелетной мышцы. Ветви нерва и кровеносные сосуды следуют за соединительнотканными компонентами мышцы нервной клетки и с одним или несколькими мельчайшими кровеносными сосудами, называемыми капиллярами (Источник: training.seer.cancer.gov).
Клеточная мембрана мышечной клетки называется сарколеммой, и эта мембрана, как и мембрана нейронов, поддерживает мембранный потенциал.Таким образом, импульсы проходят по мембранам мышечных клеток точно так же, как и по мембранам нервных клеток. Однако «функция» импульсов в мышечных клетках — вызывать сокращение. Чтобы понять, как сокращается мышца, вам нужно немного узнать о структуре мышечных клеток.
| Скелетная мышца — это мышца, прикрепленная к скелету. Сотни или тысячи мышечных волокон (клеток) связываются вместе, образуя отдельные скелетные мышцы.Мышечные клетки представляют собой длинные цилиндрические структуры, которые связаны плазматической мембраной (сарколеммой) и лежащей выше базальной пластинкой и, когда сгруппированы в пучки (пучки), образуют мышцы. Сарколемма образует физический барьер против внешней среды, а также передает сигналы между внешней средой и мышечной клеткой. Саркоплазма — это специализированная цитоплазма мышечной клетки, которая содержит обычные субклеточные элементы наряду с аппаратом Гольджи, обилием миофибрилл, модифицированным эндоплазматическим ретикулумом, известным как саркоплазматический ретикулум (SR), миоглобином и митохондриями.Поперечные (Т) -рубочки инвагинируют сарколемму, позволяя импульсам проникать в клетку и активировать SR. Как показано на рисунке, SR образует сеть вокруг миофибрилл, накапливая и обеспечивая Ca 2+ , который необходим для сокращения мышц. Миофибриллы — это сократительные единицы, которые состоят из упорядоченного расположения продольных миофиламентов. Миофиламенты могут быть толстыми (состоящими из миозина) или тонкими (состоящими в основном из актина).Характерные «полосы» скелетных и сердечных мышц легко наблюдаются при световой микроскопии в виде чередующихся светлых и темных полос на продольных срезах. Светлая полоса (известная как I-полоса) состоит из тонких волокон, тогда как темная полоса (известная как A-полоса) состоит из толстых нитей. |
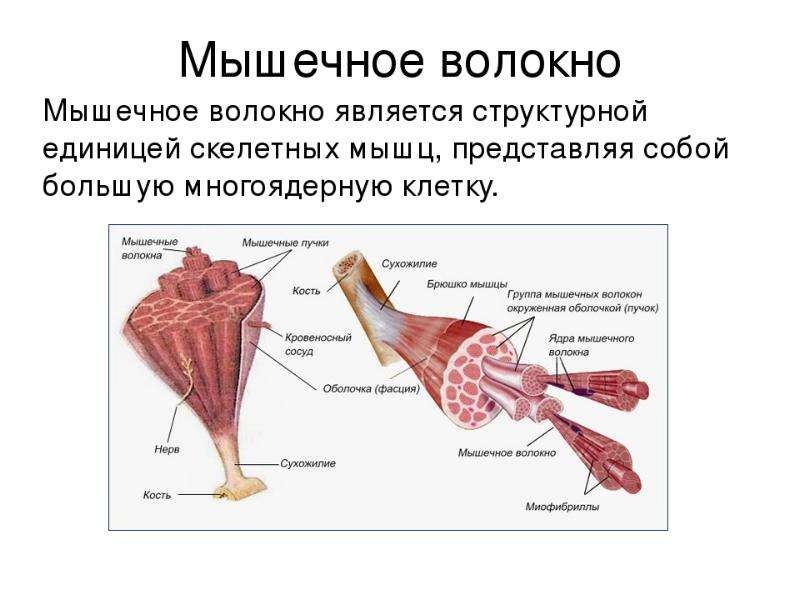
 Z-линия (также известная как Z-диск или Z-полоса) определяет боковую границу каждой саркомерной единицы. Сокращение саркомера происходит, когда Z-линии сближаются, заставляя миофибриллы сокращаться, и, следовательно, сокращается вся мышечная клетка, а затем и вся мышца (Источник: Davies and Nowak 2006).
Z-линия (также известная как Z-диск или Z-полоса) определяет боковую границу каждой саркомерной единицы. Сокращение саркомера происходит, когда Z-линии сближаются, заставляя миофибриллы сокращаться, и, следовательно, сокращается вся мышечная клетка, а затем и вся мышца (Источник: Davies and Nowak 2006).SARCOLEMMA имеет уникальную особенность: в ней есть дыры. Эти «отверстия» ведут в трубки, называемые ПОПЕРЕЧНЫМИ ТРУБКАМИ (или сокращенно Т-ТРУБКАМИ). Эти канальцы проходят вниз в мышечную клетку и огибают МИОФИБРИЛЫ. Однако эти канальцы НЕ открываются внутрь мышечной клетки; они полностью проходят и открываются где-то еще на сарколемме (т. е. эти канальцы не используются для транспортировки вещей в мышечные клетки и из них).Функция T-TUBULES заключается в проведении импульсов с поверхности клетки (SARCOLEMMA) вниз в клетку и, в частности, в другую структуру клетки, называемую SARCOPLASMIC RETICULUM.
Мышечное волокно возбуждается через двигательный нерв, который генерирует потенциал действия, который распространяется по поверхностной мембране (сарколемме) и поперечной трубчатой системе в более глубокие части мышечного волокна. Белок рецептора (DHP) воспринимает деполяризацию мембраны, изменяет ее конформацию и активирует рецептор рианодина (RyR), который высвобождает Ca 2+ из SR.Затем Ca 2+ связывается с тропонином и активирует процесс сокращения (Jurkat-Rott and Lehmann-Horn 2005).
Мембраны саркоплазматического ретикулума (SR) в непосредственной близости от Т-канальца. «RyR» — это белки, способствующие высвобождению кальция из SR, «SERCA2» — это белки, которые помогают транспортировать кальций в SR (Brette and Orchard 2007).
SARCOPLASMIC RETICULUM (SR) немного похож на эндоплазматический ретикулум других клеток, например. г., он полый. Но основная функция SARCOPLASMIC RETICULUM — ХРАНЕНИЕ ИОНОВ КАЛЬЦИЯ. Саркоплазматический ретикулум очень богат клетками скелетных мышц и тесно связан с МИОФИБРИЛАМИ (и, следовательно, МИОФИЛАМЕНТАМИ). Мембрана SR хорошо приспособлена для обработки кальция: существуют «насосы» (активный транспорт) для кальция, так что кальций постоянно «закачивается» в SR из цитоплазмы мышечной клетки (так называемая SARCOPLASM). В результате в расслабленной мышце очень высокая концентрация кальция в SR и очень низкая концентрация в саркоплазме (и, следовательно, среди миофибрилл и миофиламентов).Кроме того, мембрана имеет специальные отверстия или «ворота» для кальция. В расслабленной мышце эти ворота закрыты, и кальций не может пройти через мембрану. Итак, кальций остается в SR. Однако, если импульс проходит по мембране SR, кальциевые «ворота» открываются, и, следовательно, кальций быстро диффундирует из SR в саркоплазму, где расположены миофибриллы и миофиламенты. Как вы увидите, это ключевой шаг в сокращении мышц.
г., он полый. Но основная функция SARCOPLASMIC RETICULUM — ХРАНЕНИЕ ИОНОВ КАЛЬЦИЯ. Саркоплазматический ретикулум очень богат клетками скелетных мышц и тесно связан с МИОФИБРИЛАМИ (и, следовательно, МИОФИЛАМЕНТАМИ). Мембрана SR хорошо приспособлена для обработки кальция: существуют «насосы» (активный транспорт) для кальция, так что кальций постоянно «закачивается» в SR из цитоплазмы мышечной клетки (так называемая SARCOPLASM). В результате в расслабленной мышце очень высокая концентрация кальция в SR и очень низкая концентрация в саркоплазме (и, следовательно, среди миофибрилл и миофиламентов).Кроме того, мембрана имеет специальные отверстия или «ворота» для кальция. В расслабленной мышце эти ворота закрыты, и кальций не может пройти через мембрану. Итак, кальций остается в SR. Однако, если импульс проходит по мембране SR, кальциевые «ворота» открываются, и, следовательно, кальций быстро диффундирует из SR в саркоплазму, где расположены миофибриллы и миофиламенты. Как вы увидите, это ключевой шаг в сокращении мышц.
Миофибриллы состоят из миофиламентов двух типов: толстых и тонких.В скелетных мышцах эти миофиламенты расположены по очень правильной и точной схеме: толстые миофиламенты обычно окружены 6 тонкими миофиламентами (вид с торца). На виде сбоку тонкие миофиламенты можно увидеть над и под каждым толстым миофиламентом.
Поперечный разрез миофибрилл, показывающий расположение толстых и тонких миофиламентов.
Бар = 100 нм. Изображение Widrick et al. (2001)
Источник: Цховребова и Триник (2003).
Строение мышц
Каждая миофибрилла состоит из множества субъединиц, выстроенных встык. Эти субъединицы, конечно, состоят из миофиламентов и называются SARCOMERES. Рисунки выше и ниже показывают только очень маленькую часть всего длина миофибриллы, поэтому вы можете увидеть только один полный SARCOMERE.
В каждом саркомере тонкие миофиламенты проходят с каждого конца.Толстый миофиламенты находятся в середине саркомера и не расширяются до конца. Из-за такого расположения при просмотре скелетных мышц под микроскопом концы саркомера (там, где только тонкие миофиламенты найдены) кажутся светлее, чем центральная часть (которая темная, потому что наличия толстых миофиламентов). Таким образом, миофибрилла имеет чередующиеся светлые и темные области, потому что каждая состоит из множества выстроенных в линию саркомеров концы с концами. Вот почему скелетная мышца называется ПОЛОСКОЙ МЫШЦЫ (т.е., чередующиеся светлые и темные участки выглядят как полосы или полосы). Светлые области называются I-BAND, а более темные — A-BAND. Рядом с центром каждого I-BAND находится тонкая темная линия, называемая Z-LINE (или Z-мембрана на рисунке ниже). Z-LINE — это место, где соседние саркомеры сходятся вместе и тонкие миофиламенты соседних саркомеров перекрываются немного. Таким образом, саркомер можно определить как область между Z-линиями.
Используется с разрешения Джона
W.Kimball
Толстые миофиламенты состоят из белка, называемого МИОЗИН. Каждый МИОЗИН молекула имеет хвост, который образует ядро толстой миофиламента плюс головка, выступающая из сердцевины нити. Эти головы MYOSIN также обычно называют ПЕРЕКРЕСТНЫМИ МОСТАМИ.
МИОЗИН HEAD имеет несколько важных характеристик:
- он имеет АТФ-связывающие сайты, в которые помещаются молекулы АТФ.АТФ представляет собой потенциальная энергия.
- у него есть сайты связывания ACTIN, в которые помещаются молекулы ACTIN. Актин является частью тонкой миофиламента и будет рассмотрено более подробно в ближайшее время.
- он имеет «шарнир» в месте выхода из ядра толстой миофиламента. Это позволяет голове поворачиваться вперед и назад, а «поворот» как будет описано вкратце, что на самом деле вызывает сокращение мышц.
Молекулы актина (или G-актин, как указано выше) имеют сферическую форму и образуют длинные
цепи. Каждая тонкая миофиламент содержит две такие цепи, которые наматываются на
друг друга. Молекулы ТРОПОМИОЗИНА — это одиночные тонкие молекулы, которые обвивают
цепочка ACTIN. В конце каждого тропомиозина находится молекула ТРОПОНИНА.
Молекулы ТРОПОМИОЗИНА и ТРОПОНИНА связаны друг с другом.Каждый
из этих 3 белков играет ключевую роль в сокращении мышц:
- АКТИН — когда актин соединяется с ГОЛОВКОЙ МИОЗИНА, АТФ, связанный с голова распадается на АДФ. Эта реакция высвобождает энергию, которая вызывает МИОСИН ГОЛОВА ПОВОРОТА.
- ТРОПОМИОЗИН — В расслабленной мышце МИОЗИНОВЫЕ ГОЛОВКИ толстой миофиламента лежат против молекул ТРОПОМИОЗИНА тонкой миофиламента.Пока ГОЛОВКИ МИОЗИНА остаются в контакте с ТРОПОМИОЗИНОМ, ничего не происходит (т. Е. мышца остается расслабленной).
- ТРОПОНИН — молекулы тропонина имеют участки связывания для ионов кальция. Когда ион кальция заполняет это место, он вызывает изменение формы и положения ТРОПОНИНА. И когда ТРОПОНИН смещается, он притягивает ТРОПОМИОЗИН, к которому он прилагается. Когда ТРОПОМИОЗИН перемещается, ГОЛОВА МИОЗИНА, которая касалась теперь тропомиозин вступает в контакт с лежащей в основе молекулой ACTIN.
Нити раздвижные
Сокращение мышц
1 — Поскольку скелетная мышца является произвольной мышцей, для ее сокращения требуется нервный импульс. Итак, шаг 1 в сокращении — это когда импульс передается. от нейрона к САРКОЛЕММЕ мышечной клетки.
2 — Импульс проходит по САРКОЛЕММЕ и вниз ТРУБКИ.От Т-ТРУБОК импульс переходит в САРКОПЛАЗМИЧЕСКУЮ РЕТИКУЛУМ.
3 — Когда импульс движется по саркоплазматической сети (SR), кальциевые ворота в мембране СР открываются. В результате КАЛЬЦИЙ рассеивается вне SR и среди миофиламентов.
4 — Кальций заполняет сайты связывания в молекулах ТРОПОНИНА. Как отмечалось ранее, это изменяет форму и положение ТРОПОНИНА, что, в свою очередь, вызывает движение присоединенной молекулы ТРОПОМИОЗИНА.
5 — Движение ТРОПОМИОЗИНА позволяет ГОЛОВЕ МИОЗИНА контактировать с АКТИНОМ (анимация: сокращение и разрушение миофиламентов
АТП и мостовое движение).
6 — Контакт с ACTIN приводит к повороту ГОЛОВКИ МИОЗИНА.
7 — Во время поворота ГОЛОВКА МИОЗИНА надежно прикреплена к ACTIN. Итак, когда ГОЛОВКА поворачивается, она тянет ACTIN (и, следовательно, весь тонкая миофиламент) вперед.(Очевидно, одна ГОЛОВА МИОЗИНА не может тянуть вся тонкая миофиламентная нить. Многие МИОЗИНОВЫЕ ГОЛОВКИ вращаются одновременно, или почти так, и их коллективных усилий достаточно, чтобы вытащить все тонкая миофиламент).
8 — В конце вертлюга АТФ входит в сайт связывания на поперечный мост, и это разрывает связь между поперечным мостом (миозин) и актин. ГОЛОВА МИОЗИНА затем поворачивается назад. Когда он поворачивается назад, ATP распадается на ADP & P, и поперечный мостик снова связывается с актином. молекула.
9 — В результате HEAD снова прочно привязан к ACTIN. Однако, поскольку ГОЛОВА не была прикреплена к актину при повороте назад, HEAD будет связываться с другой молекулой ACTIN (т.е. на тонком миофиламенте). Как только HEAD присоединен к ACTIN, поперечный мост снова вертится, ТАК ШАГ 7 ПОВТОРЯЕТСЯ.
Пока присутствует кальций (присоединенный к ТРОПОНИНУ), шаги с 7 по 9 продолжу. И при этом тонкая миофиламент «тянется» МИОЗИНОВЫМИ ГОЛОВКАМИ толстой миофиламента.Таким образом, THICK & THIN миофиламенты действительно скользят мимо друг друга. При этом расстояние между Z-линиями саркомер уменьшается. По мере того как саркомеры становятся короче, миофибриллы, конечно, становится короче. И, очевидно, мышечные волокна (и вся мышца) становятся короче.
Скелетные мышцы расслабляются, когда прекращается нервный импульс. Нет импульса значит что мембрана SARCOPLASMIC RETICULUM больше не проницаема к кальцию (т.д., отсутствие импульса означает, что КАЛЬЦИЕВЫЕ ВОРОТА закрываются). Итак, кальций больше не распространяется. КАЛЬЦИЕВЫЙ НАСОС в мембране теперь будет транспортировать кальций обратно в SR. При этом ионы кальция покидают связывающую сайты, посвященные МОЛЕКУЛАМ ТОПОНИНА. Без кальция ТРОПОНИН возвращается к своему исходная форма и положение, как и прилагаемый ТРОПОМИОЗИН. Это означает что ТРОПОМИОЗИН теперь снова на месте, в контакте с ГОЛОВКОЙ МИОЗИНА. Итак, головка MYOSIN больше не контактирует с ACTIN и, следовательно, мышца перестает сокращаться (т.э., расслабляет).
Сокращение
Итак, в большинстве случаев кальций является «переключателем», который заставляет мышцы «включение и выключение» (сокращение и расслабление). Когда мышца используется для расширенного Период, запасы АТФ могут уменьшиться. Когда концентрация АТФ в мышцах снижается, ГОЛОВКИ МИОЗИНА остаются связанными с актином и больше не могут поворачиваться. Это снижение в уровне АТФ в мышце вызывает МЫШЕЧНУЮ УСТАЛОСТЬ. Хотя кальций все еще присутствует (и нервный импульс передается мышце), сокращение (или, по крайней мере, сильное сокращение) невозможно.
Анимация, иллюстрирующая сокращение мышц:
Crash Course: мышцы
Сокращение мышечных клеток
Сокращение скелетных мышц
Механизм скольжения нити при сокращении миофибриллы (Wiley)
Потенциалы действия и сокращение мышц
Виды сокращений:
- 1 — изотонический — напряжение или сила, создаваемая мышцей, больше
чем нагрузка и мышца укорачиваются
2 — изометрический — нагрузка больше, чем создаваемое натяжение или сила мышцами и мышца не укорачивается
ГЛАДКИЕ МЫШЦЫ:
- непроизвольная мышца; иннервируется вегетативной нервной системой (висцеральной эфферентные волокна)
- в основном обнаруживается в стенках полых органов и трубок
- Веретенообразные ячейки, как правило, размещены в листах
- клетки не имеют Т-канальцев и очень мало саркоплазматического ретикулума Клетки
- не содержат саркомеров (поэтому не имеют бороздок), но состоят из толстые и тонкие миофиламенты.Тонкие нити в гладкой мускулатуре не содержат тропонин. Кальций
- не связывается с тропонином, а скорее с белком, называемым кальмодулином. Комплекс кальций-кальмодулин «активирует» миозин, который затем связывается с актином, и начинается сокращение (поворот поперечных мостиков).
Два типа гладкой мускулатуры:
1 — висцеральный, или унитарные, гладкие мышцы
- найдены в стенках полых органов (напр.г., мелкие кровеносные сосуды, пищеварительные тракта, мочевыделительной системы и репродуктивной системы)
- несколько волокон сжимаются как единое целое (поскольку импульсы легко проходят через щелевые переходы от клетки к клетке) и в некоторых случаях самовозбудимы (генерируют спонтанные потенциалы действия и сокращения)
- 2 — многокомпонентная гладкая мышца
- состоит из двигательных единиц, которые активируются нервной стимуляцией
- обнаружен в стенках крупных кровеносных сосудов, в глазу (изменение формы линзы, чтобы разрешить размещение и размер зрачка для регулировки количество света, попадающего в глаз) и у основания волосяного фолликула (мышцы «гусиная шишка»)
Полезные ссылки:
Актин Миозиновая анимация
Введение к физиологии мышц и дизайну
Макгроу-Хилл: Костно-мышечной системы
Цитированная литература:
Бретт, Ф., и К. Орчард. 2007. Возрождение исследования сердечных Т-канальцев. Физиология 22: 167-173.
Дэвис, К. Э., и К. Дж. Новак. 2006. Молекулярные механизмы мышечных дистрофий: старые и новые игроки. Nature Reviews Molecular Cell Biology 7: 762-773.
Хирш, Н. П. 2007. Нервно-мышечный переход в здоровье и болезни. Британский журнал анестезии 99: 132-138.
Хопкинс, П. М. 2005. Основы анестезии, второе изд. Мосби, Лондон.
Юркат-Ротт, К., и Ф. Леманн-Хорн. 2005. Мышечные каналопатии и критические точки в функциональных и геномных исследованиях. Журнал клинических исследований 115: 2000-2009.
Цховребова Л. и Дж. Триник. 2003. Титин: собственность и семейные отношения. Nature Reviews Molecular Cell Biology 4: 679-689.
Видрик, Дж. Дж., Дж. Дж. Роматовски, К. М. Норенберг, С. Т. Кнут, Дж. Л. В. Бейн, Д. А. Райли, С. В. Траппе, Т. А. Траппе, Д. Л. Костилл и Р. Х. Фиттс. 2001. Функциональные свойства медленных и быстрых волокон икроножной мышцы после 17-дневного космического полета.Журнал прикладной физиологии 90: 2203-2211.
Назад
к программе BIO 301
Лекция Примечания 4 — Защита крови и тела
Документ без названия
Документ без названия БИО 378
Скелетная мышца
Характеристики мышцы:
- возбудимость — реагирует на раздражители (например, нервные импульсы)
- сократимость — возможность укорочения
- растяжимость — растягивается при вытягивании
Типы мышц:
- скелет:
- прикреплен к костям и перемещает скелет
- также называется поперечно-полосатой мышцей (из-за его внешнего вида под микроскопом, как показано на фотографии слева)
- произвольная мышца
- гладкая (фото справа)
- Непроизвольная мышца
- мышца внутренних органов (e.g., в стенках кровеносных сосудов, кишечника и других «полых» структур и органов тела)
- сердечный:
- мышца сердца
- принудительное
Строение скелетных мышц
Структура скелетных мышц:
Скелетные мышцы обычно прикрепляются к кости сухожилиями, состоящими из соединительной ткани.Эта соединительная ткань также покрывает всю мышцу и называется эпимизием. Скелетные мышцы состоят из множества субъединиц или пучков, называемых фасиклами (или пучками). Фасцикулы также окружены соединительной тканью (называемой перимизием), и каждый пучок состоит из многочисленных мышечных волокон (или мышечных клеток). Мышечные клетки, заключенные в оболочку эндомизия, состоят из множества фибрилл (или миофибрилл), и эти миофибриллы состоят из длинных белковых молекул, называемых миофиламентами. В миофибриллах есть два типа миофиламентов: толстые миофиламенты и тонкие миофиламенты.
Скелетные мышцы значительно различаются по размеру, форме и расположению волокон. Они варьируются от очень крошечных нитей, таких как стремечковая мышца среднего уха, до больших масс, таких как мышцы бедра. Скелетные мышцы могут состоять из сотен или даже тысяч мышечных волокон, связанных вместе и завернутых в соединительнотканный покров. Каждая мышца окружена соединительнотканной оболочкой, называемой эпимизием. Фасция, соединительная ткань вне эпимизия, окружает и разделяет мышцы.Части эпимизия выступают внутрь, чтобы разделить мышцу на части. Каждый отсек содержит пучок мышечных волокон. Каждый пучок мышечных волокон называется пучком и окружен слоем соединительной ткани, называемым перимизием. Внутри пучка каждая отдельная мышечная клетка, называемая мышечным волокном, окружена соединительной тканью, называемой эндомизием. Скелетные мышцы имеют обильное снабжение кровеносных сосудов и нервов. Прежде чем скелетное мышечное волокно сможет сократиться, оно должно получить импульс от нейрона.Обычно артерия и по крайней мере одна вена сопровождают каждый нерв, который проникает в эпимизий скелетной мышцы. Ветви нерва и кровеносные сосуды следуют за соединительнотканными компонентами мышцы нервной клетки и с одним или несколькими мельчайшими кровеносными сосудами, называемыми капиллярами (Источник: training.seer.cancer.gov). |
Клеточная мембрана мышечной клетки называется сарколеммой, и эта мембрана, как и мембрана нейронов, поддерживает мембранный потенциал.Таким образом, импульсы проходят по мембранам мышечных клеток точно так же, как и по мембранам нервных клеток. Однако «функция» импульсов в мышечных клетках — вызывать сокращение. Чтобы понять, как сокращается мышца, вам нужно немного узнать о структуре мышечных клеток.
| Скелетная мышца — это мышца, прикрепленная к скелету. Сотни или тысячи мышечных волокон (клеток) связываются вместе, образуя отдельные скелетные мышцы.Мышечные клетки — это длинные цилиндрические структуры, которые связаны плазматической мембраной (сарколемма). Саркоплазма — это специализированная цитоплазма мышечной клетки, которая содержит обычные субклеточные элементы наряду с аппаратом Гольджи, обилием миофибрилл, модифицированным эндоплазматическим ретикулумом, известным как саркоплазматический ретикулум (SR), миоглобином и митохондриями. Поперечные (Т) -рубочки инвагинируют сарколемму, позволяя импульсам проникать в клетку и активировать SR. Как показано на рисунке, SR образует сеть вокруг миофибрилл, накапливая и обеспечивая Ca 2+ , который необходим для сокращения мышц. Миофибриллы — это сократительные единицы, которые состоят из упорядоченного расположения продольных миофиламентов. Миофиламенты могут быть толстыми (состоящими из миозина) или тонкими (состоящими в основном из актина). Характерные «полосы» скелетных и сердечных мышц легко наблюдаются при световой микроскопии в виде чередующихся светлых и темных полос на продольных срезах. Светлая полоса (известная как I-полоса) состоит из тонких волокон, тогда как темная полоса (известная как A-полоса) состоит из толстых нитей.Z-линия (также известная как Z-диск или Z-полоса) определяет боковую границу каждой саркомерной единицы. Сокращение саркомера происходит, когда Z-линии сближаются, заставляя миофибриллы сокращаться, и, следовательно, сокращается вся мышечная клетка, а затем и вся мышца (Источник: Davies and Nowak 2006). |
SARCOLEMMA имеет уникальную особенность: в нем есть отверстия. Эти «отверстия» ведут в трубки, называемые ПОПЕРЕЧНЫМИ ТРУБКАМИ (или сокращенно Т-ТРУБКАМИ).Эти канальцы проходят вниз в мышечную клетку и огибают МИОФИБРИЛЫ. Однако эти канальцы НЕ открываются внутрь мышечной клетки; они полностью проходят и открываются где-то еще на сарколемме (т. е. эти канальцы не используются, чтобы вводить и выводить предметы в мышечную клетку). Функция T-TUBULES заключается в проведении импульсов с поверхности клетки (SARCOLEMMA) вниз в клетку и, в частности, в другую структуру клетки, называемую SARCOPLASMIC RETICULUM.
SARCOPLASMIC RETICULUM (SR) немного похож на эндоплазматический ретикулум других клеток, т.е.г., он полый. Но основная функция SARCOPLASMIC RETICULUM — ХРАНЕНИЕ ИОНОВ КАЛЬЦИЯ. Саркоплазматический ретикулум очень богат клетками скелетных мышц и тесно связан с МИОФИБРИЛАМИ (и, следовательно, МИОФИЛАМЕНТАМИ). Мембрана SR хорошо приспособлена для обработки кальция: существуют «насосы» (активный транспорт) для кальция, так что кальций постоянно «закачивается» в SR из цитоплазмы мышечной клетки (так называемая SARCOPLASM). В результате в расслабленной мышце очень высокая концентрация кальция в SR и очень низкая концентрация в саркоплазме (и, следовательно, среди миофибрилл и миофиламентов).Кроме того, мембрана имеет специальные отверстия или «ворота» для кальция. В расслабленной мышце эти ворота закрыты, и кальций не может пройти через мембрану. Итак, кальций остается в SR. Однако, если импульс проходит по мембране SR, кальциевые «ворота» открываются, и, следовательно, кальций быстро диффундирует из SR в саркоплазму, где расположены миофибриллы и миофиламенты. Как вы увидите, это ключевой шаг в сокращении мышц.
Миофибриллы состоят из миофиламентов двух типов: толстых и тонких.В скелетных мышцах эти миофиламенты расположены по очень правильной и точной схеме: толстые миофиламенты обычно окружены 6 тонкими миофиламентами (вид с торца). На виде сбоку тонкие миофиламенты можно увидеть над и под каждым толстым миофиламентом.
Поперечный разрез миофибрилл, показывающий расположение толстых и тонких миофиламентов.
Бар = 100 нм. Изображение Widrick et al. (2001)
Источник: Цховребова и Триник (2003).
Строение мышц
Каждая миофибрилла состоит из множества субъединиц, выстроенных встык. Эти субъединицы, конечно же, состоят из миофиламентов и называются SARCOMERES. На рисунках выше и ниже показан только очень небольшой участок всей длины миофибриллы, поэтому вы можете увидеть только один полный SARCOMERE.
В каждом саркомере тонкие миофиламенты проходят с каждого конца.Толстые миофиламенты находятся в середине саркомера и не доходят до концов. Из-за такого расположения, когда скелетные мышцы просматриваются под микроскопом, концы саркомера (где обнаруживаются только тонкие миофиламенты) кажутся светлее, чем центральный участок (который темный из-за наличия толстых миофиламентов). Таким образом, миофибрилла имеет чередующиеся светлые и темные области, потому что каждая состоит из множества саркомеров, выстроенных встык. Вот почему скелетная мышца называется ПОЛОСКОЙ МЫШЦЫ (т.е. чередующиеся светлые и темные участки выглядят как полосы или полосы). Рядом с центром каждой I-BAND есть тонкая темная линия, называемая Z-LINE. Z-LINE — это место, где соседние саркомеры сходятся вместе, а тонкие миофиламенты соседних саркомеров слегка перекрываются. Таким образом, саркомер можно определить как область между Z-линиями.
Используется с разрешения Джона В. Кимбалла
Толстые миофиламенты состоят из белка, называемого МИОЗИН. Каждая молекула МИОЗИНА имеет хвост, который образует сердцевину толстой миофиламента, и головку, выступающую из сердцевины волокна.Эти МИОЗИНОВЫЕ головы также обычно называют ПЕРЕКРЕСТНЫМИ МОСТАМИ.
ГОЛОВКА МИОЗИН имеет несколько важных характеристик:
- он имеет АТФ-связывающие сайты, в которые подходят молекулы АТФ. АТФ представляет собой потенциальную энергию.
- он имеет сайты связывания ACTIN, в которые подходят молекулы ACTIN. Актин входит в состав тонкой миофиламента.
- он имеет «шарнир» в месте выхода из ядра толстой миофиламента.Это позволяет голове поворачиваться вперед и назад, и, как будет вскоре описано, именно «вращение» на самом деле вызывает сокращение мышц.
Молекулы актина (или G-актин, как указано выше) имеют сферическую форму и образуют длинные цепи. Каждая тонкая миофиламент содержит две такие цепи, которые наматываются друг на друга. Молекулы ТРОПОМИОЗИНА — это одиночные тонкие молекулы, которые обвивают цепь АКТИНА.В конце каждого тропомиозина находится молекула ТРОПОНИНА. Молекулы ТРОПОМИОЗИНА и ТРОПОНИНА связаны друг с другом. Каждый из этих 3 белков играет ключевую роль в сокращении мышц:
- АКТИН — когда актин объединяется с ГОЛОВКОЙ МИОЗИНА, АТФ, связанный с головой, распадается на АДФ. Эта реакция высвобождает энергию, которая заставляет МИОЗИНОВУЮ ГОЛОВУ ПОВОРАЧАТЬ.
- ТРОПОМИОЗИН — В расслабленной мышце МИОЗИНОВЫЕ ГОЛОВКИ толстой миофиламента лежат напротив молекул ТРОПОМИОЗИНА тонкой миофиламента.Пока МИОЗИНОВЫЕ ГОЛОВКИ остаются в контакте с ТРОПОМИОЗИНОМ, ничего не происходит (т. Е. Мышца остается расслабленной).
- ТРОПОНИН — Молекулы тропонина имеют участки связывания для ионов кальция. Когда ион кальция заполняет это место, он вызывает изменение формы и положения ТРОПОНИНА. И когда ТРОПОНИН смещается, он тянет ТРОПОМИОЗИН, к которому он прикреплен. Когда ТРОПОМИОЗИН перемещается, ГОЛОВКА МИОЗИНА, которая касалась тропомиозина, теперь вступает в контакт с лежащей в основе молекулой АКТИНА.
Нити раздвижные
Сокращение мышц
1 — Поскольку скелетная мышца является произвольной мышцей, сокращение требует нервного импульса. Итак, шаг 1 в сокращении — это когда импульс передается от нейрона к SARCOLEMMA мышечной клетки.
2 — Импульс проходит по САРКОЛЕММЕ и вниз по Т-ТРУБКАМ. От Т-ТРУБОК импульс переходит к САРКОПЛАЗМИЧЕСКОЙ СЕТЧИКУ.
3 — Когда импульс проходит по саркоплазматической сети (SR), кальциевые ворота в мембране SR открываются. В результате КАЛЬЦИЙ диффундирует из SR и между миофиламентами.
4 — Кальций заполняет сайты связывания в молекулах ТРОПОНИНА. Как отмечалось ранее, это изменяет форму и положение ТРОПОНИНА, что, в свою очередь, вызывает движение присоединенной молекулы ТРОПОМИОЗИНА.
5 — Движение ТРОПОМИОЗИНА позволяет ГОЛОВЕ МИОЗИНА связываться с АКТИНОМ.
6 — Контакт с ACTIN приводит к повороту ГОЛОВКИ МИОЗИНА.
7 — Во время поворота ГОЛОВКА МИОЗИНА надежно прикреплена к ACTIN. Таким образом, когда ГОЛОВКА поворачивается, она тянет АКТИН (и, следовательно, всю тонкую миофиламент) вперед. (Очевидно, одна МИОЗИНОВАЯ ГОЛОВКА не может тянуть всю тонкую миофиламент. Многие МИОЗИНОВЫЕ ГОЛОВКИ вращаются одновременно или почти так, и их коллективных усилий достаточно, чтобы потянуть всю тонкую миофиламент).
8 — В конце вертлюга АТФ входит в сайт связывания на поперечном мостике, и это разрывает связь между поперечным мостиком (миозином) и актином. ГОЛОВА МИОЗИНА затем поворачивается назад. Когда он поворачивается назад, АТФ распадается на АДФ и Ф, и поперечный мостик снова связывается с молекулой актина.
9 — В результате HEAD снова прочно привязан к ACTIN. Однако, поскольку ГОЛОВА не была прикреплена к актину при повороте назад, ГОЛОВА будет связываться с другой молекулой АКТИНА (т.е., еще один назад на тонкой миофиламенте). После того, как ГОЛОВКА присоединена к ACTIN, поперечный мост снова поворачивается, ТАК, ШАГ 7 ПОВТОРЯЕТСЯ.
Пока присутствует кальций (связан с ТРОПОНИН), шаги с 7 по 9 будут продолжаться. И, при этом, тонкая миофиламент «тянется» МИОЗИНОВЫМИ ГОЛОВКАМИ толстой миофиламента. Таким образом, ТОЛСТЫЕ и ТОНКИЕ миофиламенты фактически скользят мимо друг друга. При этом расстояние между Z-линиями саркомера уменьшается.Чем короче саркомеры, тем короче миофибриллы. И, естественно, мышечные волокна (и вся мышца) становятся короче.
Скелетные мышцы расслабляются, когда прекращается нервный импульс. Отсутствие импульса означает, что мембрана SARCOPLASMIC RETICULUM больше не проницаема для кальция (т.е. отсутствие импульса означает, что КАЛЬЦИЕВЫЕ ВОРОТА закрываются). Итак, кальций больше не диффундирует. НАСОС КАЛЬЦИЯ в мембране теперь будет транспортировать кальций обратно в SR. Когда это происходит, ионы кальция покидают сайты связывания на МОЛЕКУЛАХ ТОПОНИНА.Без кальция ТРОПОНИН возвращается к своей исходной форме и положению, как и прикрепленный ТРОПОМИОЗИН. Это означает, что ТРОПОМИОЗИН теперь снова на своем месте и находится в контакте с ГОЛОВКОЙ МИОЗИНА. Таким образом, головка MYOSIN больше не контактирует с ACTIN, и, следовательно, мышца перестает сокращаться (т.е. расслабляется).
Сокращение
Итак, в большинстве случаев кальций является «переключателем», который включает и выключает мышцы (сокращение и расслабление).Когда мышца используется в течение длительного периода, запасы АТФ могут уменьшиться. Когда концентрация АТФ в мышцах снижается, МИОЗИНОВЫЕ ГОЛОВКИ остаются связанными с актином и больше не могут поворачиваться. Это снижение уровня АТФ в мышцах вызывает УСТАЛОСТЬ МЫШЦ. Даже если кальций все еще присутствует (и нервный импульс передается мышцам), сокращение (или, по крайней мере, сильное сокращение) невозможно.
Контрольные вопросы:
Что такое пучок (или фасция), мышечное волокно, миофибрилла, миофиламент? Что такое сарколемма? Что такое тубула? Что такое саркоплазматический ретикулум? Какие бывают 2 типа миофиламентов? Почему скелетные мышцы выглядят поперечно-полосатыми? Что такое саркомер? Какие 3 белка составляют тонкие миофиламенты и как эти белки расположены в тонких миофиламентах? Какой тип белка состоит из толстых миофиламентов? Что такое поперечный мост? Что происходит с расстоянием между Z-линиями во время сокращения? Какова роль каждого из следующих элементов в сокращении скелетных мышц: сарколемма, t-трубочка, саркоплазматический ретикулум, кальций, тропонин, тропомиозин, актин, поперечные мостики, АТФ? Что происходит с кальцием, когда мышцы расслабляются?
Анимация, иллюстрирующая сокращение мышц:
Механизм скольжения нити при сокращении миофибриллы (Wiley)
Как структура и состав мышц влияют на качество мяса и мяса
Скелетные мышцы состоят из нескольких тканей, таких как мышечные волокна, соединительная и жировая ткани.Этот обзор направлен на описание особенностей этих различных мышечных компонентов и их взаимосвязи с технологическими, пищевыми и сенсорными свойствами мяса / мяса различных видов домашнего скота и рыб. Таким образом, сократительный и метаболический типы, размер и количество мышечных волокон, содержание, состав и распределение соединительной ткани, а также содержание и липидный состав внутримышечного жира играют роль в определении внешнего вида, цвета, нежности мяса / мяса. , сочность, вкус и технологическая ценность.Интересно, что биохимические и структурные характеристики мышечных волокон, внутримышечной соединительной ткани и внутримышечного жира, по-видимому, играют независимую роль, что позволяет предположить, что свойства этих различных компонентов мышц могут независимо модулироваться генетикой или факторами окружающей среды для достижения эффективности производства и улучшения качества мяса. / качество мяса.
1. Введение
Мышечная масса животных и видов рыб, используемых для производства продуктов питания для людей, составляет от 35 до 60% их массы тела.Поперечно-полосатые скелетные мышцы, прикрепленные к позвоночнику, участвуют в произвольных движениях и облегчают передвижение и осанку. Скелетные мышцы обладают широким разнообразием форм, размеров, анатомического расположения и физиологических функций. Для них характерен сложный вид, так как помимо мышечных волокон они содержат соединительную, жировую, сосудистую и нервную ткани. Мышечные волокна, внутримышечная соединительная ткань и внутримышечный жир играют ключевую роль в определении качества мяса и рыбы.Что касается мяса и водных продуктов, различные заинтересованные стороны, то есть производители, забойщики, переработчики, дистрибьюторы и потребители, предъявляют различные и конкретные требования к качеству, которые зависят от использования ими продуктов. Качество обычно описывается четырьмя терминами: безопасность (гигиеническое качество), полезность (питательная ценность), удовлетворение (органолептическое качество) и удобство обслуживания (простота использования, способность к переработке и цены). Удовлетворенность определяется качествами, воспринимаемыми потребителями.Они включают цвет, текстуру и сочность, а также вкус, который связан с ароматами, выделяемыми во рту при употреблении продукта. Удовлетворенность также обусловлена технологическими качествами, которые отражают способность продукта к переработке. Чаще всего они связаны со снижением технологического выхода из-за снижения водоудерживающей способности во время хранения в холодильнике (экссудации) и варки или из-за повреждения, которое происходит после нарезки. Лучшие технологические качества связаны с низкими потерями.Питательные качества зависят в первую очередь от питательной ценности жиров, углеводов и белков, входящих в состав пищи. Считается, что мясо, богатое белками с высоким содержанием незаменимых аминокислот и полиненасыщенных жирных кислот, обладает хорошими питательными качествами. Наконец, гигиенические качества отражают способность продукта безопасно потребляться. В первую очередь они связаны с бактериальной нагрузкой продукта и наличием в продукте химических остатков, таких как гербициды или пестициды, а также других загрязнителей окружающей среды.Среди перечисленных качеств критически важными для потребителей являются нежность, цвет и полезность говядины. Однако основной причиной отказа потребителя от покупки говядины является непостоянство его нежности [1]. Лучшее качество у рыбы — плотное, вязкое мясо с хорошей водоудерживающей способностью [2]. В мясе и рыбном мясе на эти качества влияют многие факторы in vivo, и посмертные (pm), такие как виды, генотипы, факторы питания и окружающей среды, условия убоя и переработка pm.Поскольку эти факторы также влияют на структуру и состав скелетных мышц, их влияние на качество мяса может в значительной степени включать прямую связь между внутримышечными биологическими свойствами и характеристиками качества мяса. Однако между видами такие отношения не всегда ясны. Таким образом, цель данной статьи — дать обзор структуры и состава (мышечные волокна, внутримышечная соединительная ткань и внутримышечный жир) мышц домашнего скота и рыбы и их взаимосвязи с различными качествами.Недавние геномные исследования различных видов выращиваемых животных с целью выявления новых биомаркеров качества мяса были ранее рассмотрены [3] и, при необходимости, будут кратко рассмотрены в этой статье.
2. Структура мышц
2.1. Макроскопическая шкала
Скелетная мышца состоит примерно на 90% из мышечных волокон и на 10% из соединительной и жировой тканей. Соединительная ткань в скелетных мышцах делится на эндомизий, окружающий каждое мышечное волокно, перимизий, окружающий пучки мышечных волокон, и эпимизий, окружающий мышцу в целом [4, 5].
Когда куски мяса состоят из уникальной мышцы, эпимизий удаляется. Однако, когда кусок мяса включает несколько мышц, отсутствует только внешний эпимизий (рис. 1). Скелетная мышца также содержит жировую ткань и, в меньшей степени, сосудистую и нервную ткани. У рыбы съедобная часть, филе, состоит из нескольких мышц (миомеров), которые соединены друг с другом и разделены соединительнотканными оболочками толщиной в несколько миллиметров, известными как миосепты. Миосепты демонстрируют структурную непрерывность от позвоночной оси до кожи.Их роль заключается в передаче сил сокращения волокон одного миомера к другому, а также к скелету и коже. Эта конкретная структура с чередующимися мышечными и соединительными оболочками называется метамерной организацией. У «круглой» рыбы промышленного размера форма миомеров филе напоминает букву W (рис. 2). Однако эта организация более сложна в поперечном сечении (т. Е. Котлета) (рис. 3). Миосепта может рассматриваться как эпимизия мышц наземных видов сельскохозяйственных животных.Другие внутримышечные соединительные ткани рыб демонстрируют организацию, аналогичную структуре, обнаруженной у наземных животных. Уникальной характеристикой рыбьих мышц является анатомическое разделение на макроскопическом уровне трех основных типов мышц: большая белая мышца, поверхностная красная мышца (вдоль кожи) и промежуточная розовая мышца. Эти мышцы присутствуют в каждом миомере (рис. 3). Рыбное филе также содержит внутримышечную жировую ткань, расположенную в миомере между миофибриллами и в перимизии, но в основном в миосептах, разделяющих миомеры.
2.2. Микроскопические весы
Мышечные волокна представляют собой удлиненные, многоядерные и веретенообразные клетки диаметром примерно от 10 до 100 микрометров и длиной от нескольких миллиметров у рыб до нескольких сантиметров у наземных животных. У всех видов размер волокон увеличивается с возрастом животного и является важным параметром послеродового роста мышц. Плазматическая мембрана мышечных волокон известна как сарколемма. Площадь поперечного сечения (ППС) волокон зависит от их метаболического и сократительного типов (см. Раздел 3.1 для типов мышечных волокон). У рыб распределение волокон по размеру варьируется в зависимости от важности гипертрофической (увеличение размера клеток из-за увеличения объема) и гиперплазической стадии роста (увеличение объема мышц из-за увеличения количества клеток). Одновременное присутствие мелких и крупных волокон приводит к так называемой «мозаичной» структуре, обычно встречающейся у рыб (рис. 4).
Независимо от вида миофибриллы, выстроенные в пучки, занимают почти весь внутриклеточный объем мышечных волокон.Миофибриллы имеют диаметр примерно 1 мкм и м и состоят из небольших субъединиц: миофиламентов (рис. 1). Продольные поперечные сечения миофибрилл, наблюдаемые с помощью электронной микроскопии, демонстрируют чередование темных (полосы A) и светлых областей (полосы I). Каждая полоса I разделена на две части линией Z. Повторяющаяся единица, находящаяся между двумя линиями Z, — это саркомер, который является сократительной функциональной единицей миофибриллы (рис. 5). Тонкие миофиламенты в основном состоят из актина, тропонинов T, I и C (которые регулируют сокращение мышц) и тропомиозина, расположенного встык вдоль актиновой нити.Толстые миофиламенты в основном состоят из набора молекул миозина, активность АТФазы которых катализирует расщепление аденозинтрифосфата (АТФ) до аденозиндифосфата (АДФ) и обеспечивает химическую энергию, необходимую для сокращения мышц. Саркоплазма, то есть цитоплазма мышечных волокон, содержит множество растворимых белков, включая ферменты гликолитического пути и миоглобин, который переносит кислород в митохондрии и окрашивает клетки в красный цвет. Он также содержит гранулы гликогена, которые представляют собой первичный локальный запас энергии мышечных клеток, помимо липидных капель.
3. Биохимический состав мышц
Скелетные мышцы содержат примерно 75% воды, 20% белка, 1–10% жира и 1% гликогена. Биохимические свойства основных мышечных компонентов (то есть миофибрилл, соединительной и жировой ткани) описаны ниже.
3.1. Мышечные волокна
Мышечные волокна обычно характеризуются сократительными и метаболическими свойствами [6, 7]. Сократительные свойства в первую очередь зависят от изоформ тяжелых цепей миозина (MyHC), присутствующих в толстых филаментах.В большинстве зрелых скелетно-поперечно-полосатых мышц млекопитающих экспрессируются четыре типа MyHC: I, IIa, IIx и IIb. Активность АТФазы этих MyHC связана со скоростью сокращения: медленным (тип I) и быстрым (типы IIa, IIx и IIb). Волокна типа I демонстрируют низкую интенсивность сокращения, но устойчивы к утомлению. Они преобладают в постуральных и дыхательных мышцах. Для сокращения мышц требуется энергия АТФ, потребности в котором сильно различаются для разных типов мышечных волокон [8].
В мышцах используются два основных пути регенерации АТФ: окислительный (аэробный) путь, посредством которого пируват окисляется митохондриями, и гликолитический (анаэробный) путь, при котором пируват превращается в молочную кислоту в саркоплазме.Относительная важность этих двух путей определяет тип метаболических волокон: окислительные (красные; богатые миоглобином, который является переносчиком кислорода и пигментом, отвечающим за красный цвет) или гликолитические (белые; почти не содержат миоглобина, поскольку потребности в кислороде сильно ограничены). . Обычно окислительные красные волокна имеют меньшую CSA, чем гликолитические белые волокна. Однако разница в размере между типами волокон может варьироваться в зависимости от мышцы и внутри одной и той же мышцы. Например, CSA окислительного волокна больше, чем CSA гликолитического волокна в красной части мышцы semitendinosus у свиней [10].Точно так же в мышце Rectus abdominis крупного рогатого скота окислительное красное волокно CSA больше, чем CSA белого гликолитического волокна [11]. Наконец, мышечные волокна — это динамические структуры, которые могут переключаться с одного типа на другой по следующему пути: IIIAIIXIIB [12]. Сводка свойств различных типов волокон в зрелых скелетных мышцах млекопитающих представлена в таблице 1. Несмотря на очевидное присутствие их генов, ни одна из трех изоформ взрослых Fast MyHC не присутствует в зрелых мышцах всех видов млекопитающих.Фактически, IIb MyHC не экспрессируется у овец и лошадей и был обнаружен только в определенных мышцах крупного рогатого скота с сильными различиями между породами [13]. Напротив, сильная экспрессия IIb MyHC наблюдается в скелетных мышцах свиней обычных пород, отобранных по худобе и высоким показателям роста [14]. Независимо от вида, наиболее важным фактором, определяющим состав мышечных волокон, является тип мышц, вероятно, связанный с их конкретной физиологической функцией. Для данной мышцы состав волокон варьируется в зависимости от вида.Так, мышца свиньи Longissimus содержит примерно 10% волокон типа I, 10% IIA, 25% IIX и 55% IIB, тогда как коровий Longissimus содержит в среднем 30% волокон типа I, 18% IIA и 52% IIX. . На состав мышечных волокон также влияют порода, пол, возраст, физическая активность, температура окружающей среды и методы кормления. Как и у млекопитающих, мышечные волокна птиц можно классифицировать по их сократительной и метаболической активности. Однако описаны дополнительные классы, например, многотонные иннервируемые медленные волокна типов IIIa и IIIb, специфичные для мускулов птиц [15].У птиц трудно сопоставить изоформу MyHC с типом волокна из-за одновременного присутствия в зрелых волокнах взрослых и развивающихся типов MyHC. Рыбы также обладают различными типами мышечных волокон, которые характеризуются своими сократительными и метаболическими свойствами. Однако, в отличие от млекопитающих или птиц, у рыб можно наблюдать анатомическое разделение между двумя основными типами волокон. Например, у форели быстрые волокна (похожие на волокна IIB млекопитающих) находятся в центре в области поперечного сечения тела, а медленные волокна (аналогичные типу I млекопитающих) находятся на периферии вдоль продольной линии под кожа [16].Помимо этих двух основных типов волокон, у определенных видов или на определенных стадиях развития могут быть обнаружены второстепенные типы, такие как промежуточный тип (например, тип розового волокна, сопоставимый с типом IIA). Два основных типа белого и красного волокна были связаны с экспрессией быстрого и медленного MyHC, соответственно [17]. Однако может быть сложно систематически сопоставить изоформу MyHC с типом волокна из-за одновременного присутствия нескольких MyHC в одном и том же волокне у рыб, особенно в мелких мышечных волокнах.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| : очень низкий; ++: низкий; +++: средний; ++++: высокий; +++++: очень высокий. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
3.2. Внутримышечная соединительная ткань
Соединительная ткань, которая окружает мышечные волокна и пучки волокон, представляет собой рыхлую соединительную ткань. Он состоит из клеток и внеклеточного матрикса (ECM), который в основном состоит из сложной сети коллагеновых волокон, обернутых в матрикс протеогликанов (PG) [4, 18, 19]. В этой статье основное внимание уделяется молекулам, которые, как было продемонстрировано или предположительно, играют роль в определении сенсорного качества мяса. Коллагены — это семейство волокнистых белков.Независимо от типа коллагена, основная структурная единица коллагена (тропоколлаген) представляет собой спиральную структуру, состоящую из трех полипептидных цепей, намотанных друг на друга, образуя спираль. Молекулы тропоколлагена стабилизируются межцепочечными связями с образованием фибрилл диаметром 50 нм. Эти фибриллы стабилизированы внутримолекулярными связями (дисульфидные или водородные мостики) или межмолекулярными связями (включая пиридинолин и дезоксипиридинолин), известными как поперечные связи (КС). В скелетных мышцах обнаружены различные типы коллагена.Фибриллярные коллагены I и III являются основными, которые появляются у млекопитающих [19]. У рыб преобладает коллаген I и V типов [20]. Другими основными компонентами соединительной ткани являются PG [21]. PG представляют собой сложные многофункциональные молекулы, которые состоят из основного белка с молекулярной массой от 40 до 350 кДа, связанных ковалентными связями с несколькими десятками гликозаминогликановых цепей (GAG). PG образуют большие комплексы, связываясь с другими PG и волокнистыми белками (такими как коллаген). Они связывают катионы (например,г., натрий, калий и кальций) и воду [22]. Пропорция и степень внутримышечного сшивания коллагена зависят от типа мышц, вида, генотипа, возраста, пола и уровня физических упражнений [23]. Общее содержание коллагена колеблется от 1 до 15% сухого веса мышц у взрослого крупного рогатого скота [19], тогда как оно колеблется от 1,3 ( Psoas major ) до 3,3% ( Latissimus dorsi ) сухого веса мышц у крупных белых свиней. на стадии товарного убоя [24]. У домашней птицы коллаген равен 0.От 75 до 2% сухого веса мышц [25]. Сообщалось о различном содержании в рыбе в зависимости от вида (количество колеблется от 1 до 10% между сардинами и морями [26]), внутри вида и между передней и задней частями (более богатыми) филе [27]. PG составляют небольшую часть сухой массы мышц (от 0,05% до 0,5% у крупного рогатого скота в зависимости от мышц) [28].
3.3. Внутримышечный жир
У млекопитающих резервный жир расположен в нескольких внешних и внутренних анатомических местах, например, вокруг и внутри мышц для межмышечных и внутримышечных жиров (МВФ).В этой статье мы уделяем основное внимание МВФ, поскольку межмышечный жир обрезается во время разделки и, таким образом, оказывает меньшее влияние на свинину и говядину. У рыбы жир располагается подкожно, в перимизии и миосепте, и в основном последние влияют на качество мяса и рассматриваются в этой статье. МВФ в основном состоит из структурных липидов, фосфолипидов и запасных липидов (триглицеридов). Последние в основном (примерно 80%) хранятся в мышечных адипоцитах, находящихся между волокнами и пучками волокон, и небольшая часть (5–20%) хранится в виде липидных капель внутри миофибрилл в цитоплазме (внутриклеточные липиды) [29].Между типами мышц содержание фосфолипидов относительно постоянно (т.е. колеблется от 0,5 до 1% свежих мышц у свиней), тогда как содержание триглицеридов в мышцах сильно варьируется независимо от вида [30, 31]. Содержание IMF сильно зависит от размера и количества внутримышечных адипоцитов. У свиней [32, 33] и крупного рогатого скота [30, 34] индивидуальные вариации содержания IMF в данной мышце между животными с аналогичным генетическим фоном были связаны с вариациями в количестве внутримышечных адипоцитов.Напротив, было продемонстрировано, что различия в содержании IMF в данной мышце между животными одного и того же генетического происхождения и подвергнутыми разному потреблению энергии с пищей связаны с вариациями в размере адипоцитов [33]. У рыб увеличение ширины миосепты, вероятно, связано с увеличением количества и размера адипоцитов [35]. Содержание IMF варьируется в зависимости от анатомического происхождения мышц, возраста, породы, генотипа, рациона и условий выращивания домашнего скота [30, 36–39]. Например, китайские и американские свиньи (напр.g., Meishan и Duroc, соответственно) или европейских местных пород свиней (например, иберийских и баскских) имеют более высокие уровни IMF, чем у европейских обычных генотипов, таких как крупная белая, ландрас или пьетрен [40]. Содержание IMF варьируется от 1 до примерно 6% от массы мускулов свежего Longissimus у обычных генотипов свиней на стадии коммерческого убоя, со значениями до 10% у некоторых пород [38]. У крупного рогатого скота содержание IMF в мышцах Longissimus варьируется от 0,6% у бельгийских голубых до 23.3% у чернокожих японцев при убое в возрасте 24 месяцев [41]. У французских пород крупного рогатого скота было продемонстрировано, что отбор по мышечной массе был связан со снижением содержания IMF и коллагена. Например, основные мясные породы шаролез, лимузин и блонд д’Аквитан имеют меньше МВФ, чем выносливые породы, такие как Обрак и Салер, причем все они демонстрируют более низкие уровни МВФ, чем молочные породы [42] или американские или азиатские породы, выращиваемые в тех же условиях. [36, 43]. В рыбе содержание IMF также варьируется между видами от менее 3% в «постных» видах, таких как треска, до более 10% в «жирных» видах, таких как атлантический лосось [37], но также и внутри видов.Например, в мясе лосося содержание жира может колебаться от 8 до 24% [44].
4. Отношения между различными мышечными компонентами
Исследования, основанные на сравнении типов мышц, показывают, что содержание IMF обычно положительно коррелирует с процентным содержанием окислительных волокон и отрицательно — с гликолитическими волокнами [45]. Хотя окислительные волокна, особенно медленные волокна, демонстрируют более высокое содержание внутримиоцеллюлярных липидов, чем быстрые гликолитические волокна [46], и хотя содержание IMF часто было выше в окислительных, чем в гликолитических мышцах свиней (т.е.e., Semispinalis против Longissimus мускулов) [47], многие исследования также указывают на отсутствие строгой связи между общим содержанием IMF и составом мышечных волокон [6]. В крайних случаях содержание IMF может быть в три раза выше в белом гликолитике, чем в красной окислительной части мышцы Semitendinosus у свиньи [34] (рис. 6). Отрицательная корреляция между содержанием IMF и окислительным метаболизмом была также обнаружена в мышце свиньи Longissimus при функциональном геномном подходе [48].Однако положительные генетические и фенотипические корреляции наблюдались между содержанием IMF и CSA мышечных волокон в мышце свиньи Longissimus [49]. У рыб, у которых белые и красные мышцы анатомически разделены, предполагается, что красные мышцы демонстрируют более высокое содержание жира, чем белые, из-за большего количества жировых клеток в перимизии и большего количества липидных капель в мышечных волокнах. У атлантического лосося отмечена отрицательная генетическая корреляция (rg = -0,85) между общим количеством волокон и содержанием IMF, что предполагает, что при аналогичном весе отбор на низкий IMF приведет к увеличению количества волокна [50].Кроме того, наблюдалась отрицательная корреляция между содержанием коллагена и IMF (rg = -0,8), что указывает на то, что увеличение IMF вызовет относительное снижение содержания коллагена в мышцах, вероятно, из-за его «растворения» в мышечной ткани [51]. Систематической взаимосвязи между биохимическими характеристиками соединительной ткани и типом мышечных волокон у мясных животных не обнаружено. Напротив, у рыб содержание коллагена в красных мышцах выше, чем в белых [52].
5.Механизмы изменения мускулов и качества мяса и плоти: модуляция мышечными свойствами
После убоя мясо обычно хранят в холодном помещении при 4 ° C в течение от 2 до 30 дней в зависимости от вида, последующих методов обработки и упаковки. Самые длительные сроки хранения используются для говядины (от одной до двух недель для тушек до одного месяца для мясных кусков, хранящихся под вакуумом), чтобы облегчить естественный процесс размягчения (старения). Уменьшение CSA мышечных волокон, наблюдаемое во время охлаждения, является результатом латерального сжатия миофибрилл, амплитуда которого зависит от убойного стресса животных и технологии оглушения (Рисунок 7) [53].Фаза старения характеризуется различными ультраструктурными изменениями и приводит к фрагментации мышечных волокон. Действие различных протеолитических систем приводит к характерным разрывам миофибрилл по Z-линиям (рис. 7). Митохондрии деформируются, а их мембраны изменяются [18, 54]. В результате деградации костамеров, то есть соединения цитоскелетных белков с сарколеммой, сарколемма отделяется от периферических миофибрилл [55]. Согласно Ouali et al.[54], ферментативный процесс начинается сразу после кровотечения с активацией каспаз, которые ответственны за повреждение клеточных компонентов во время апоптоза. Другие протеолитические системы (например, кальпаин, протеасома и катепсины) вступают во владение, чтобы продолжить деградацию белка в клетках и мышечной ткани [56].
Соединительная ткань также претерпевает морфологические изменения при старении мяса [19, 21], которые выявляются уже в 12 часов вечера у кур [25], но только после 2 недель вечера у крупного рогатого скота [57].Это разложение способствует солюбилизации коллагена во время приготовления, улучшая нежность приготовленного мяса. Также было высказано предположение о косвенном влиянии PG на нежность приготовленного мяса. Фактически, во время старения снижение устойчивости перимизия связано с уменьшением количества PG вместе с увеличением растворимости коллагена из-за повышенной активности некоторых ферментов. Одна из гипотез состоит в том, что PG могут разрушаться (спонтанно или ферментативно) во время созревания и больше не защищать коллаген от ферментативных атак [21].У рыб тендеризация мяса связана с постепенным разрушением эндомизия [58] и отслоением волокон друг от друга из-за разрыва связей с эндомизием и миосептой [59]. Рыбы с мягким мясом демонстрируют большее расщепление эндомизия (коллагена, PG) [60]. В миофибриллах рыб наблюдаются слабые ультраструктурные изменения актомиозинового комплекса, в отличие от мышц крупного рогатого скота [61]. Так, у морского леща ( Sparus aurata ) полосы I и Z разрушаются лишь частично после 12 дней хранения в холодильнике [62].
6. Связь между свойствами мышц и качеством мяса
Среди различных компонентов качества мяса рассматриваются технологические, пищевые и сенсорные параметры. Компонент качества питания в первую очередь определяется химическим составом мышечной ткани при убое, в то время как технологические и сенсорные компоненты являются результатом сложных взаимодействий между химическим составом и метаболическими свойствами мышцы при убое и биохимическими изменениями после полудня, которые приводят к ее превращению в мясо. [56, 63].Структура и состав мышц, кинетика изменения ТМ и применяемые дополнительные методы использования и обработки мяса (например, измельчение, приготовление) различаются в зависимости от вида и отруба, что приводит к существенным внутренним различиям в качестве мяса между видами животных и порезы. Следовательно, иерархия между наиболее желательными качественными компонентами варьируется между видами. Яркие примеры включают нежность у крупного рогатого скота, твердость рыбного мяса и способность удерживать воду у свиней и кур.
6.1. Технологическое качество
После убоя, в зависимости от вида и рынка, туши хранятся в холодном помещении, а затем разрезаются на куски или мускулы. Во время хранения меняется внутренняя структура мышц. Мышечные волокна сжимаются в поперечном направлении, вытесняя внутриклеточную воду во внеклеточные пространства, размер которых увеличивается. Впоследствии эта вода выводится через отрезанные концы мышц [53]. Что касается переработки в готовые продукты, то технологическое качество связано с водоудерживающей способностью мяса, то есть его способностью удерживать внутреннюю воду.Водоудерживающая способность сильно зависит от скорости и степени снижения pm pH. Высокая скорость в сочетании с высокой температурой мышц (например, из-за стресса или интенсивной физической активности непосредственно перед убоем) вызывает денатурацию мышечных белков, снижение водоудерживающей способности и усиление экссудации, а также потерю мяса у свиней и птицы при варке. Значительное снижение pH (например, кислое мясо) снижает чистый электрический заряд белков, что также снижает водоудерживающую способность [64, 65].Измерение pH в течение одного часа после убоя, а затем на следующий день для оценки скорости и степени снижения pH, определение цвета и потери воды при хранении в холодильнике являются основными показателями технологического качества мяса. Состав мышечных волокон влияет на технологическое качество мяса, такое как водоудерживающая способность, которая зависит от эволюции кинетики pH и температуры мышц. Снижение pH PM обычно происходит быстрее в гликолитических мышцах, чем в окислительных [66], хотя эта взаимосвязь не носит систематического характера.Фактически, рН в 45 мин после полудня намного ниже в мышце Psoas major (27% волокна I), чем в мышце Longissimus (10% волокон I) [6], что можно объяснить более низкой буферной способностью. волокна типа I (таблица 1) или различия в кинетике снижения температуры ПМ в зависимости от анатомического расположения мышц. Кроме того, стимуляция метаболизма гликолита в мышцах в течение часа после убоя увеличивает скорость снижения pH, что в сочетании с высокой мышечной температурой может привести к денатурации белка и бледному, мягкому и экссудативному (PSE) синдрому в белых мышцах, особенно в свиньи и куры.Напротив, степень падения pH после пм (предельное значение pH; обычно определяется через 24 часа после полудня) постоянно больше в белых гликолитических мышцах, чем в красных окислительных мышцах, из-за более высокого содержания гликогена в мышцах in vivo и во время убоя у быстро сокращающихся белых гликолитические волокна. В мышцах крупной белой свиньи Longissimus увеличение скорости и степени снижения pH PM связано с более бледным цветом, большей яркостью и экссудацией [49, 67]. У свиней были идентифицированы два основных гена, которые существенно влияют на кинетику снижения pH PM и водоудерживающую способность.Мутация в гене RYR1 (также известном как ген галотана), который кодирует рецептор рианодина, который является частью канала высвобождения кальция саркоплазматического ретикулума, ответственна за быстрое снижение pH pm и развитие мяса PSE [68] . Другой дефект качества свинины связан с мутацией в гене PRKAG3, который кодирует субъединицу AMP-активируемой протеинкиназы (AMPK) [69]. Эта мутация приводит к очень высокому уровню гликогена в мышцах при убое (+ 70%), особенно в гликолитических мышцах, что в значительной степени отвечает за снижение pH после полудня и «кислое мясо» с низкой водоудерживающей способностью.Интересно, что длинная мышца мутантных свиней PRKAG3 содержит больше окислительных волокон [47] и более низкую буферную способность [70], что способствует низкому предельному pH в дополнение к большему производству лактата из гликогена. Недавнее протеомное исследование крупного рогатого скота выявило корреляцию между метаболическими, антиоксидантными и протеолитическими ферментами со снижением pH. Эти данные позволяют лучше понять биологические механизмы раннего вечера, участвующие в снижении pH [71].
6.2. Пищевая ценность
Мясо и плоть являются важным источником белков, незаменимых аминокислот (АК), незаменимых жирных кислот (ЖК), минералов и витаминов (А, Е и В), которые определяют качество питания.Профиль AA относительно постоянен между мышцами или между видами [72]. Однако мышцы, богатые коллагеном, имеют более низкую питательную ценность из-за высокого содержания в них глицина, несущественного АА [19]. По сравнению с белыми мышцами красные мышцы имеют большее содержание миоглобина и, следовательно, обеспечивают большее количество гемового железа, которое легко усваивается организмом. Хотя IMF составляет небольшую часть мышечной массы, он участвует в потреблении ЖК человеком, потому что содержание и природа (т.е. профиль) мясных ЖК варьируются в зависимости от вида, анатомического происхождения данной мышцы и диеты животных [30, 73].Диетические стратегии интенсивно изучались и оптимизировались для снижения потребления насыщенных жирных кислот и увеличения цис-мононенасыщенных и полиненасыщенных жирных кислот или других биоактивных липидов в продуктах животного происхождения для потребления человеком [30, 73]. Кроме того, поскольку n-3 жирные кислоты с более чем 20 атомами углерода в основном включаются в фосфолипиды, а не в триглицериды, можно обогатить содержание мяса этими полиненасыщенными жирными кислотами без увеличения IMF. Например, что касается биоактивных липидов, особенность мяса жвачных животных заключается в наличии жирных кислот, которые прямо или косвенно являются результатом биогидрирования рубца и которые, как предполагается, являются биоактивными жирными кислотами, такими как руменовая кислота, которая является основным природным изомером конъюгированные линолевые кислоты [30] и, как известно, предотвращают определенные формы рака на животных моделях.Однако во время выдержки после полудня и хранения мяса липиды претерпевают изменения (например, перекисное окисление), важность которого зависит от состава ЖК в мясе. Эти изменения могут ухудшить сенсорные (например, цвет, вкус) и питательные качества мяса [63, 74].
6.3. Сенсорное качество
6.3.1. Цвет и внешний вид
Состав мышечных волокон влияет на цвет мяса через количество и химическое состояние миоглобина. Высокое содержание миоглобина в волокнах типа I и типа IIA приводит к положительной взаимосвязи между долей этих волокон и интенсивностью красного цвета.В глубоких мышцах и мясе, хранящемся под вакуумом, миоглобин находится в восстановленном состоянии и имеет пурпурно-красный цвет. Под воздействием кислорода миоглобин превращается в оксимиоглобин, что придает мясу привлекательный ярко-красный цвет. Во время хранения мяса миоглобин может окисляться до метмиоглобина, что приводит к появлению коричневого непривлекательного цвета, который негативно воспринимается потребителями [75, 76]. Многие факторы до и pm, такие как вид животных, пол, возраст, анатомическое расположение и физиологическая функция мышц, физическая активность, кинетика снижения pH после полудня, скорость охлаждения туши и упаковка мяса, влияют на концентрацию и химическое состояние пигментов и, как следствие, цвет мяса [77].Таким образом, мышцы крупного рогатого скота, овец, лошадей и перелетных птиц (например, гусей, уток), которые содержат большое количество волокон типа I, богатых миоглобином, склонны к образованию метмиоглобина и снижению стабильности цвета. Напротив, высокая доля гликолитических волокон приводит к производству белого мяса, как у кур и свиней. КРС с двумя мускулами (мутация в гене миостатина ) имеет мышцы с высокой долей быстрых гликолитических волокон и, как следствие, бледное мясо [3].
Цвет мяса также зависит от диеты. Например, кормление телят коровьим молоком, не содержащим железа, ограничивает биосинтез миоглобина, в результате чего мясо становится бледным в результате дефицита железа.
У рыб только поверхностная латеральная красная мышца, богатая миоглобином, имеет интенсивный (обычно коричневый) цвет, тогда как белая мышца довольно полупрозрачна. В случае лососевых оранжево-красный цвет мякоти обусловлен присутствием в мышечных волокнах пищевых каротиноидных пигментов, таких как астаксантин.Различия в уровнях липидов могут приводить к вариациям толщины миосепты (то есть признаку «белых полос»), что может быть обнаружено обученной сенсорной панелью у рыб, которые демонстрируют контрастирующую мышечную массу, связанную с различным содержанием липидов [78]. На данном срезе рыбы (поперечном сечении) красные мышцы также можно наблюдать на краю белой мышцы, что составляет примерно 90% мышцы. Восприятие потребителями красной мышцы, которая быстро окисляется после полудня до коричневой, а затем до черной, обычно отрицательно, и эту красную мышцу иногда удаляют для продуктов премиум-класса (например,г., копченое филе). Помимо цвета, количество и распределение мраморности внутри мышечного среза влияют на внешний вид и, таким образом, могут повлиять на принятие мяса и мясных продуктов потребителями (см. Раздел 6.3.3). У рыб другим серьезным дефектом внешнего вида мяса (филе) является так называемый дефект «зияющего», который возникает в результате частичного разрушения миосепты или границы раздела волокон / миосепт. Биологическое и / или технологическое происхождение этого дефекта качества остается неясным.
6.3.2. Нежность
Нежность и ее изменчивость являются важнейшими сенсорными характеристиками для потребителей говядины. Мясо говядины имеет гораздо более высокую базовую жесткость (определяемую пропорцией, распределением и природой внутримышечной соединительной ткани) и более низкий процесс размягчения PM, чем у свинины или птицы [63]. Таким образом, продолжительность старения после полудня важна для нежности говядины [79]. У свиней и домашней птицы кинетика подкисления мускулов PM, которая быстрее, чем у крупного рогатого скота [79], сильно влияет на текстуру (т.е.е. сочность, нежность) и технологические свойства мяса (например, водоудерживающая способность) [63]. У крупного рогатого скота взаимосвязь между характеристиками волокон и нежностью является сложной и варьируется в зависимости от мускулатуры, пола, возраста и породы [80]. Например, у быков болезненность Longissimus thoracis часто связана с уменьшением CSA волокон и повышением окислительного метаболизма, тогда как в мышцах Vastus lateralis и semitendinosus чем выше гликолитическая активность, тем мягче мясо [81].Однако отрицательная корреляция между интенсивностью окислительного метаболизма и болезненностью также наблюдалась в мышцах крупного рогатого скота Longissimus [82]. Используя биомаркеры нежности говядины, Picard et al. [83] продемонстрировали, что у пород, характеризующихся более быстрым гликолитическим метаболизмом, таких как французские мясные породы, самые нежные Longissimus thoracis являются наиболее окислительными. Напротив, у пород, у которых метаболизм мышц более окислительный, таких как абердин-ангусский, наиболее гликолитический Longissimus thoracis является самым нежным.Это согласуется с тем фактом, что у пород с окислительной мускулатурой, таких как ангус или молочные породы, ребрышки с низкой интенсивностью красного цвета более нежные. Напротив, среди основных французских пород говядины, которые демонстрируют больше гликолитических мускулов, чем краснее мускулы, тем нежнее мясо [83]. Более высокая доля гликолитических волокон может улучшить нежность определенных мышц за счет ускорения старения после полудня из-за наличия более высокого отношения кальпаин / кальпастатин (два белка, участвующих в протеолизе) [84] в мясе видов животных с медленным старением мяса, такие как крупный рогатый скот и овцы [82].Однако по мнению других авторов, улучшение нежности мяса, связанное с увеличением доли волокон типа I, объясняется более высоким белковым обменом и связанной с ним протеолитической активностью в окислительных волокнах [85]. У быков, за исключением стейков из ребер, нежность мяса, по-видимому, связана не с CSA волокон, а с метаболическими свойствами мышечных волокон.
Функциональное геномное исследование свиней показало отрицательное влияние обилия быстрых волокон и высокого гликолитического метаболизма на нежность мяса [48].Это исследование также демонстрирует, что снижение экспрессии генов синтеза белка (например, генов антиапоптотических белков теплового шока и гена кальпастатина) и повышение уровня экспрессии генов, участвующих в деградации белка (особенно протеасом), связаны с более низкой силой сдвига ( т.е. улучшенная нежность) в 13:00. Отрицательная связь между средним значением CSA быстрых гликолитических волокон и болезненностью была продемонстрирована у свиней [86]. Таким образом, стратегия, направленная на увеличение общего количества волокон в сочетании с умеренным CSA волокон и увеличение процента медленно сокращающихся окислительных волокон, может быть многообещающим средством увеличения количества мышц при сохранении сенсорных качеств свинины [6].Напротив, у кур увеличение CSA волокон в мышце Pectoralis связано со снижением содержания гликогена в мышцах, более высоким предельным pH и водоудерживающей способностью, а также улучшенной болезненностью [87]. Однако противоречивые данные по цыплятам также сообщают о негативном влиянии клетчатки CSA на водоудерживающую способность и нежность мяса [88]. У рыб при сравнении видов была обнаружена отрицательная корреляция между средним диаметром мышечных волокон и плотностью мяса. Однако эта взаимосвязь кажется более противоречивой внутри видов: аналогичные результаты были получены для копченого атлантического лосося и сырого мяса коричневой и радужной форели, тогда как другие исследования не продемонстрировали взаимосвязи между размером волокон и текстурой мяса лосося или трески.В целом, как и у свиней, похоже, что для качества рыбных продуктов лучше гиперплазический, чем гипертрофический рост мышц.
Соединительная ткань влияет на нежность мяса своим составом и структурой [4], особенно у крупного рогатого скота, при этом коллаген обычно считается основным определяющим фактором силы сдвига. Однако есть существенные различия между сырым и вареным мясом. Сила сдвига сырого мяса тесно связана с содержанием в нем коллагена [21, 89].В приготовленном мясе уровень корреляции между содержанием, термической растворимостью или уровнем сшивки коллагена и силой сдвига мяса неясен и варьируется в зависимости от типа мышц и условий приготовления [90, 91]. Во время нагревания волокна коллагена сжимаются и давят на мышечные волокна, величина которых зависит от степени сшивания коллагена и организации эндомизия и перимизия. Уровень взаимодействия между коллагеном и мышечными волокнами модулирует тепловую денатурацию коллагена (т.е.е., его желатинизация) и, следовательно, развитие нежности мяса во время приготовления [89]. Считается, что у свиней и кур коллаген ограниченно влияет на сенсорное качество мяса. Причина в том, что животных забивают на относительно ранней физиологической стадии, на которой внутримышечный коллаген не имеет значительных поперечных сшивок [19].
Помимо состава, структура соединительной ткани, в частности ее организация и размер перимизиевых пучков (которые определяют зернистость мяса, особенно говядины), также играют роль в формировании текстуры мяса [ 92].Согласно Purslow [23], взаимосвязь между зернистостью мяса и консистенцией указывает на то, что нежность положительно коррелирует с долей пучков небольшого диаметра (называемых первичными пучками), но этот параметр не позволяет точно предсказать нежность. Эллис-Ури и др. [80] не продемонстрировали значимой взаимосвязи между зернистостью мяса и нежностью, оцененной опытной сенсорной группой, усилием сдвига или содержанием коллагена и растворимостью. Кроме того, поперечная сила мышцы увеличивается с увеличением толщины вторичных пучков перимизия у крупного рогатого скота [93] и свиней [94].Встречаются более крупные связки (например, третичные, четвертичные), но они редко рассматриваются в исследованиях, посвященных нежности мяса. Таким образом, их влияние на структуру соединительной ткани мышц и нежность мяса остается неясным.
Что касается рыб, сравнения между видами продемонстрировали положительную взаимосвязь между плотностью сырого мяса и содержанием в нем коллагена. Однако внутри видов такой связи не наблюдалось. Что касается влияния сшивки коллагена на твердость сырого мяса, только низкая зависимость (= 0.25) между содержанием гидроксилизилпиридинолина (КЛ) и механической прочностью филе наблюдалась у лосося [95]. Из-за своей низкой термостойкости по сравнению с таковой у млекопитающих коллаген мышечной рыбы не сохраняет свои структурные свойства во время приготовления. Таким образом, текстура приготовленной мякоти во многом зависит от миофибриллярных белков. Сравнения между видами выявили положительную корреляцию между содержанием мышечного коллагена и нежностью и эластичностью приготовленного мяса [26].Однако ни один из этих результатов не был обнаружен среди видов рыб. Виды рыб с плотной плотью демонстрируют очень плотную сеть коллагеновых волокон в эндомизии, тогда как эта сеть гораздо более рыхлая у видов с менее плотной плотью [96].
6.3.3. Сочность и вкус
У крупного рогатого скота и ягнят повышенная доля волокон типа I связана с улучшением сочности и вкуса мяса [85, 97]. Такое благоприятное влияние на вкус, вероятно, объясняется высоким содержанием фосфолипидов в волокнах типа I, причем фосфолипиды являются основным фактором, определяющим вкус приготовленного мяса [98].Однако высокое содержание полиненасыщенных ЖК в фосфолипидах увеличивает риск появления прогорклого вкуса. У свиней высокий процент быстрых оксидогликолитических волокон ухудшает водоудерживающую способность и сочность мяса [85, 99]. Часто считается, что МВФ играет ключевую роль в определении сенсорных качеств мяса или плоти у разных видов животных, положительно влияя на сочность, вкус и нежность, хотя его влияние на сенсорные характеристики варьируется у разных видов [37]. Принято считать, что очень низкие уровни IMF приводят к получению сухого мяса с плохим вкусом.Однако высокая корреляция между IMF и оценками сенсорного качества, присвоенными обученной группой, может наблюдаться только тогда, когда происходят важные изменения и высокие максимальные уровни IMF (например, у свиней) [100]. Фактически, другие факторы могут модулировать эту взаимосвязь, например, предельный pH мяса свиней или содержание и тип внутримолекулярных CL коллагена у крупного рогатого скота [37]. Например, говядина с аналогичным уровнем IMF (примерно 3,2%), полученная из четырех разных пород (ангус, симменталь, шароле и лимузин), показала похожий вкус, но более сочную у лимузина и более низкую сочность у пород ангус [101].Что касается оценки свежего мяса и мясных продуктов потребителями, влияние МВФ кажется противоречивым. Перед потреблением потребители предпочитают меньше мраморной свинины, тогда как во время потребления мясо с самой мраморной отделкой считается сочнее, нежнее и вкуснее [100, 102, 103]. Хотя жиры являются ключевым фактором в развитии вкуса во время приготовления мяса и сочности мяса, потребители часто сопротивляются мясу с видимым IMF. Таким образом, несколько исследований продемонстрировали, что уровень общей приемлемости свинины увеличивается с содержанием IMF до 2.5–3,5% [102, 104]. Однако другие исследования показывают, что значительное число потребителей предпочитают менее мраморную свинину (от 1 до 1,5% IMF) [100, 105]. Также было отмечено различие между группами потребителей, основанное на предпочтении умеренно или слегка мраморной говядины, связанное со вкусом или питательными ожиданиями, соответственно [106]. Таким образом, оценка взаимосвязи между содержанием IMF и сенсорными характеристиками мяса зависит от диетических привычек и культур потребителей, а также от рассматриваемых продуктов.Например, было продемонстрировано, что нежность, сочность и приемлемость сухого окорока увеличивается с увеличением содержания IMF [107]. Однако обратное наблюдается для вареной ветчины, приемлемость которой снижается с увеличением IMF с 2 до 4% в мышце Semimembranosus [108]. Аналогичным образом, изменение IMF от 2,9 до 10,7% по-разному влияет на приемлемость филе лосося в зависимости от конкретного продукта. Пониженное содержание IMF более благоприятно для запеченного филе, тогда как для копченого филе — наоборот [109].
7. Заключение
Три основных компонента мышц (т.е. мышечные волокна, соединительная ткань и жировая ткань) участвуют в определении различных показателей качества мяса, но в разной степени в зависимости от вида, типа мышц и мяса после убоя. -техники обработки. Относительная независимость характеристик этих трех основных компонентов мышц предполагает, что можно независимо управлять этими характеристиками с помощью генетических, пищевых и экологических факторов, чтобы контролировать качество продуктов и, таким образом, лучше соответствовать ожиданиям производителей, переработчиков мяса и потребители.Следовательно, точное знание структурных и биохимических характеристик каждого мышечного компонента и их взаимосвязи с показателями роста и параметрами качества мяса является предпосылкой для понимания и контроля биологической основы количества и качества продуктов животного происхождения. Дальнейшие исследования должны быть сосредоточены на изменении свойств мышц, которые определяют основные компоненты качества мяса у различных видов: нежность у крупного рогатого скота, водоудерживающая способность и нежность у свиней и домашней птицы, а также текстура мяса у рыбы.
