
Настольная книга хирурга: анатомический атлас нацистов помогает спасать жизни
Автор фото, Erich Lepier
Подпись к фото,Пернкопф добивался от иллюстраторов того, чтобы все выглядело максимально реалистично
Когда у хирурга и преподавателя Университета Вашингтона в Сент-Луисе Сьюзан Маккиннон во время сложной операции появляется необходимость уточнить расположение какого-либо нерва, она обращается анатомическому атласу, выпущенному еще в середине прошлого века.
Благодаря нарисованным от руки подробнейшим иллюстрациям Маккиннон быстро находит ответ на свой вопрос, и операция проходит успешно.
Этот атлас хорошо известен хирургам разных стран и, по признанию многих из них, по сей день остается самым подробным и точным. Но каждый раз, открывая этот атлас, хирургу приходится решать не менее сложный, чем сама операция, этический вопрос: можно ли пользоваться этой книгой, созданной нацистами в нацистской Германии.
Автор атласа — Эдуард Пернкопф, профессор анатомии, занимавший в свое время пост ректора Венского университета, был нацистом, так же как и четыре его помощника-иллюстратора — Эрих Лепиер, Людвиг Шротт, Карл Эндтрессер и Франц Батке. В анатомический театр, где работали авторы атласа, привозили тела жертв нацистского режима. Поэтому как минимум половина из 800 иллюстраций атласа написаны при вскрытии тел узников, погибших от рук нацистов.
Кожа, мышцы, связки, нервы, органы и кости изображены чрезвычайно подробно и правдоподобно. Картинки не для слабонервных. Пернкопф добивался от иллюстраторов того, чтобы все выглядело максимально реалистично. Единственное небольшое отступление от правды жизни — более яркие, контрастные, чем в жизни цвета — имело важный практический смысл: так читателю книги было проще ориентироваться.
Первый том атласа Пернкопфа вышел в 1937 году, второй — в 1941-м. Работу над атласом прервала война: три иллюстратора из четырех ушли на фронт.
Сейчас атлас больше не переиздается. Разрозненные тома можно найти в букинистических магазинах; в зависимости от года выпуска цена сильно разнится — от нескольких десятков до нескольких тысяч долларов.
Несмотря на то, что многие врачи и специалисты готовы заплатить немалые деньги за этот атлас, мало кто из них может держать эту книгу открыто у себя на столе — дома или в клинике.
Потому что на иллюстрациях — вскрытые и разобранные по кусочкам тела жертв нацистов.
Все дело в «темном происхождении» книги Пернкопфа. Связанный с ее использованием сложный этический вопрос по сей день не решен, что создает сложности в работе ученых и врачей.
Сьюзан Маккиннон говорит, что ее, конечно, не может не смущать история создания книги, однако отказ от использования этого атласа подчас противоречит врачебной этике: ведь от обращения к книге может зависеть исход операции.
Переживший Холокост Джозеф Полак, профессор, специалист по медицинской этике и законодательству в области медицины, говорит, что книга сама по себе представляет нравственную дилемму: ее создание было связано с настоящим злом, но теперь она может послужить на благо многим людям.
Автор фото, Keiligh Baker
Подпись к фото,Несколько экземпляров атласа Пернкопфа хранятся в Британской библиотеке
Эдуард Пернкопф работал над книгой в течение 20 лет. Известно, что Пернкопфу, талантливому выпускнику Венского университета, в карьерном росте помогло членство в Национал-социалистической немецкой рабочей партии Адольфа Гитлера.
Коллеги отзывались о Пернкопфе как о ревностном национал-социалисте, который с 1938 года каждый день приходил на работу в нацистской форме.
Когда Пернкопфа назначили деканом медицинского факультета Венского университета, он уволил с факультета всех евреев, в том числе и трех Нобелевских лауреатов.
В 1939 году в Германии был принят закон, согласно которому тела всех казненных заключенных немедленно доставлялись в анатомические театры для проведения исследований и для обучения студентов.
По словам Сабин Хильдебрандт из Медицинской школы Гарварда, по меньшей мере половина из всех 800 иллюстраций атласа сделаны во время вскрытия тел жертв нацизма. В их числе геи, лесбиянки, цыгане, политические диссиденты, евреи.
Автор фото, None
Подпись к фото,Авторы атласа — Эдуард Пернкопф (в центре) и четыре иллюстратора
В первом издании первого тома, вышедшем в 1937 году, подписи двух иллюстраторов — Эриха Липиера и Карла Эндрессера включали нацистскую символику — свастику и эмблему СС.
Даже в двухтомнике 1964 года на английском языке были сохранены оригинальные подписи иллюстраторов с нацисткой символикой, в более поздних изданиях они были заретушированы.
Тысячи экземпляров атласа были проданы по всему миру, книга была переведена на пять языков. В предисловии и введении книги говорится о чрезвычайно реалистичных, ярких иллюстрациях, которые можно назвать произведениями искусства, однако нет ни слова о том, как эти иллюстрации создавались.
В предисловии и введении книги говорится о чрезвычайно реалистичных, ярких иллюстрациях, которые можно назвать произведениями искусства, однако нет ни слова о том, как эти иллюстрации создавались.
Только в 1990-х ученые и студенты медицинских вузов начали задаваться вопросом о людях, чьи тела были использованы для создания этих иллюстраций. После того как в 1994 году стала известна жестокая правда, атлас перестал переиздаваться.
Королевская коллегия хирургов Англии заявляет, что в вузах и медицинских учреждениях Британии атлас больше не используется, хотя несколько экземпляров многотомника хранятся в библиотеках.
Однако опрос, который журнал Neurosurgery провел среди хирургов, показал, что 59% врачей, специалистов по хирургии периферических нервов, знают о существовании атласа, 13% опрошенных продолжают пользоваться им в работе.
69% опрошенных сказали, что не видят ничего предосудительного в использовании атласа, 15% признались, что, узнав о происхождении иллюстраций, засомневались в допустимости использования этой книги. 17% сообщили, что не могут решить для себя этот сложный этический вопрос.
17% сообщили, что не могут решить для себя этот сложный этический вопрос.
По словам Сьюзан Маккиннон, за все эти десятилетия, что прошли с момента первого издания атласа, не появилось ни одной книги, которая могла бы хоть как-то сравниться с трудом Пернкопфа и работавших с ним иллюстраторов — и в плане подробности и точности описания строения человеческого тела.
Маккиннон говорит, что при проведении сложных операций помощь атласа Пернкопфа неоценима — потому что на иллюстрациях прорисованы даже самые мелкие периферические нервы.
По словам хирурга, весь медицинский персонал, участвующий в проведении операции, знает о кровавой истории создания книги.
«Когда мне стало известно о темной истории создания этого атласа, я стала его хранить в сейфе в своем кабинете», — говорит Сьюзан Маккиннон.
Автор фото, Washington University/ St. Louis
Подпись к фото,Сьюзан Маккиннон говорит, что при проведении сложных операций помощь атласа Пернкопфа неоценима
В прошлом году специалист по медицинской этике Джозеф Полак и историк медицины, психиатр Майкл Гродин подготовили ответ (на основе иудейской медицинской этики) на вопрос, этично ли использовать атлас, учитывая его темное происхождение.
По мнению Полака и Гродина, иудейская медицинская этика позволяет использовать иллюстрации атласа, если это необходимо для спасения жизни человека. При этом, по их мнению, те, кто пользуется атласом, должны знать историю создания книги. Таким образом, врачи и их пациенты смогут также почтить память жертв нацистов.
В своем ответе Полак и Гродин приводят в пример опыт работы с атласом Сьюзан Маккиннон.
«Во время операции она не могла найти один нерв, хотя она — одна из лучших специалистов в своей области. Пациент сказал ей: отрежьте мне ногу, если вы не можете его найти. Она попросила принести атлас Пернкопфа и нашла нерв за несколько минут — благодаря подробным иллюстрациям атласа», — рассказал Би-би-си Джозеф Полак.
«Сьюзан спросила меня как специалиста по этике об этой ситуации. И я сказал ей, что если обращение к труду Пернкопфа поможет ей вылечить пациента, то тогда нет никаких сомнений в том, что атлас можно использовать», — добавил Полак.
Автор фото, Washington University/ St. Louis
Подпись к фото,Доктор Маккиннон во время операции
После окончания войны Пернкопфа уволили из университета и арестовали. Три года он находился в лагере для военнопленных, однако никаких обвинений ему предъявлено не было.
После освобождения он вернулся в Венский университет и продолжил работу над атласом. Третий том вышел в 1952 году.
Перкопф умер тремя годами позже, незадолго до публикации четвертого тома.
Прошло уже более 60 лет с момента выхода последних томов атласа, однако труд Пернкопфа остается для хирургов одним из самых ценных источников информации об анатомии тела человека, говорит Сабин Хильдебрандт, которая преподает анатомию в университете.
«Так думают те из нас, кто привык обращаться к атласу, когда возникает какой-либо вопрос. Многие врачи говорят, что в хирургии нервов многотомник остается уникальным источником информации, замены которому до сих пор нет», — говорит Хильдебрандт.
«Я обычно не обращаюсь к атласу Пернкопфа на лекциях и семинарах, если у меня нет времени рассказать об истории книги», — говорит она.
Джонатан Айвз, специалист по биоэтике из Бристольского университета, говорит, что атлас удивительно подробный, но его использование вызывает вопросы из-за ужасающих обстоятельств его создания.
«Если мы используем его и получаем некую «выгоду», то это невольно делает нас причастными к этому ужасу. Но вы можете возразить, что если не использовать атлас, то он будет забыт и не будет нам напоминать о том, что произошло», — говорит Айвз.
Для Маккиннон эта книга остается актуальной, несмотря на мрачные страницы истории ее создания.
«Мне как хирургу, желающему соблюдать врачебную этику, кажется, что к этому надо отнестись как к данности: я должна пользоваться теми образовательными ресурсами, которые помогут мне добиться максимально успешного исхода операции. И это именно то, чего от меня ждут мои пациенты», — говорит Маккиннон.
«По своему опыту я могу сказать, что хирургия периферических нервов будет сильно отброшена назад, если мы потеряем эти книги», — добавляет она.
Материал написан на основе статьи корреспондента Би-би-си Кайли Бейкер
Некроз мышц и смерть после инъекции железа — Атлас патологий свиней
Болезни, вызванные бактериями
Актинобациллезная плевропневмония
Атрофический ринит
Болезнь Глессера
Бордетелла бронхисептика
Бруцеллез
Дизентерия свиней
Заражение иерсинией
Илеит
Клостридий новый
Колиэнтерит
Лептоспироз
Микоплазма
Микоплазменный артрит
Отечная болезнь
Пастереллез
Рожа
Сальмонеллез
Сибирская язва
Стрептококковые инфекции
Туберкулез
Экссудативный эпидермит
Энзоотическая пневмония
Энтеритные клостридии
Болезни, вызванные вирусами
Африканская чума свиней
Болезнь «голубого глаза»
Болезнь Ауески
Везикулярная болезнь свиней
Вирус Нипах
Грипп
Классическая чума свиней
Оспа свиней
Парвовирус свиней
Респираторный коронавирус свиней
Ротавирусная инфекция
РРСС
Трансмиссивный гастроэнтерит
Цирковирус свиней
Цитомегаловирус свиней
Энцефаломиокардит
Эпизоотическая диарея свиней
Японский энцефалит В
Болезни, вызванные паразитами
Аскаридоз
Трихиуреаз
Чесотка
Дефицит питательных веществ
Железодефицитная анемия
Остеопороз, дефицит витамина Д
Шелковичное сердце
Интоксикация
Афлатоксикоз
Обезвоживание
Отравление зеараленоном
Отравление фумонизином
Эрготизм
Прочее
Атрезия заднего прохода
Вредные привычки
Выпадение вагины и шейки матки
Выпадение матки
Выпадение прямой кишки
Грыжа
Заворот желудка и кишок
Обморожение
Остеохондроз
Розовый лишай
Синдром кишечного кровотечения
Синдром стресса свиней
Солнечный ожог
Сужение прямой кишки
Тромбоцитопеническая пурпура
Язва желудка
МРТ бедра: Детальный Анатомия
1, Подвздошной мышцы. 2, Прямая мышца живота мышцы. 3, Подвздошная кость. 4, средняя ягодичная мышца 5, Малая ягодичная мышца. 6, Большая ягодичная мышца мышцы.
2, Прямая мышца живота мышцы. 3, Подвздошная кость. 4, средняя ягодичная мышца 5, Малая ягодичная мышца. 6, Большая ягодичная мышца мышцы.
1, мышца—напрягатель широкой фасции бедра. 2, Подвздошных сосудов. 3, Тонкая кишка. 4, Портняжной мышцы. 5, Грушевидная мышца (m. piriformis) 6, Прямая кишка. 7, Малая ягодичная мышца.
1, мышца—напрягатель широкой фасции бедра. 2, Подвздошной мышцы. 3, Прямая мышца живота мышцы. 4, Портняжной мышцы. 5, Малая ягодичная мышца. 6, Большая ягодичная мышца мышцы. 7, Грушевидная мышца (m. piriformis) 8, средняя ягодичная мышца.
1, Нижняя передняя подвздошная ость. 2, Прямая мышца живота мышцы. 3, мышца—напрягатель широкой фасции бедра. 4, средняя ягодичная мышца. 5, Внутренней запирательной мышц. 6, Большая ягодичная мышца мышцы. 7, Малая ягодичная мышца.
1, мышца—напрягатель широкой фасции бедра. 2, Портняжной мышцы. 3, Подвздошно-поясничная мышца. 4, Подвздошных сосудов. 5, Мочевой пузырь. 6, средняя ягодичная мышца. 7, Малая ягодичная мышца. 8, Большая ягодичная мышца мышцы.
2, Портняжной мышцы. 3, Подвздошно-поясничная мышца. 4, Подвздошных сосудов. 5, Мочевой пузырь. 6, средняя ягодичная мышца. 7, Малая ягодичная мышца. 8, Большая ягодичная мышца мышцы.
1, мышца—напрягатель широкой фасции бедра. 2, Портняжной мышцы. 3, Подвздошных сосудов. 4, Мочевой пузырь. 5, средняя ягодичная мышца. 6, Малая ягодичная мышца. 7, Прямая кишка. 8, Большая ягодичная мышца мышцы.
1, Портняжной мышцы. 2, Подвздошных сосудов. 3, Мочевой пузырь. 4, Подвздошно-поясничная мышца. 5, мышца—напрягатель широкой фасции бедра. 6, средняя ягодичная мышца. 7, Крестец. 8, Большая ягодичная мышца мышцы.
1, мышца—напрягатель широкой фасции бедра.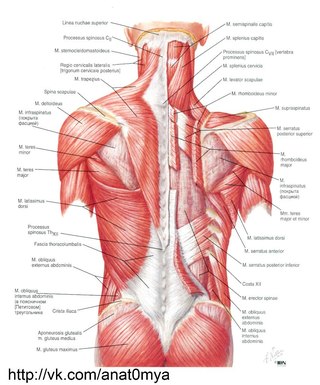 2, Прямая мышцы бедра мышцы. 3, Портняжной мышцы. 4, Мочевой пузырь. 5, Подвздошно-поясничная мышца 6, средняя ягодичная мышца. 7, Внутренней запирательной мышц. 8, Прямая кишка. 9, Большая ягодичная мышца мышцы. 10, Головки бедренной кости.
2, Прямая мышцы бедра мышцы. 3, Портняжной мышцы. 4, Мочевой пузырь. 5, Подвздошно-поясничная мышца 6, средняя ягодичная мышца. 7, Внутренней запирательной мышц. 8, Прямая кишка. 9, Большая ягодичная мышца мышцы. 10, Головки бедренной кости.
1, средняя ягодичная мышца. 2, Прямая мышцы бедра мышцы. 3, Мочевой пузырь. 4, Подвздошно-поясничная мышца. 5, Портняжной мышцы. 6, мышца—напрягатель широкой фасции бедра. 7, Большая ягодичная мышца мышцы. 8, Головки бедренной кости.
1, средняя ягодичная мышца. 2, Прямая мышцы бедра мышцы. 3, Бедренных кровеносных сосудов. 4, Мочевой пузырь. 5, Подвздошно-поясничная мышца. 6, Портняжной мышцы. 7, мышца—напрягатель широкой фасции бедра. 8, Головки бедренной кости. 9, Внутренней запирательной мышц. 10, Большая ягодичная мышца мышцы.
1, Портняжной мышцы. 2, Подвздошно-поясничная мышца. 3, средняя ягодичная мышца. 4, Большая ягодичная мышца мышцы. 5, Внутренней запирательной мышц. 6, Седалищно-прямокишечный ямка. 7, Право головки бедренной кости.
4, Большая ягодичная мышца мышцы. 5, Внутренней запирательной мышц. 6, Седалищно-прямокишечный ямка. 7, Право головки бедренной кости.
1, Прямая мышцы бедра мышцы. 2, Бедренных кровеносных сосудов. 3, Мочевой пузырь. 4, Подвздошно-поясничная мышца. 5, Портняжной мышцы. 6, мышца—напрягатель широкой фасции бедра. 7, средняя ягодичная мышца. 8, Внутренней запирательной мышц. 9, Седалищно-прямокишечный ямка. 10, Большая ягодичная мышца мышцы.
1, мышца—напрягатель широкой фасции бедра. 2, Подвздошно-поясничная мышца. 3, Мочевой пузырь. 4, Бедренных кровеносных сосудов. 5, Портняжной мышцы. 6, Прямая мышцы бедра мышцы. 7, средняя ягодичная мышца. 8, Левая головки бедренной кости. 9, Внутренней запирательной мышц. 10, Quadrilateral plate. 11, Posterior pillar. 12, Большая ягодичная мышца мышцы. 13, Большого вертела.
1, мышца—напрягатель широкой фасции бедра. 2, Прямая мышцы бедра мышцы. 3, Бедренных кровеносных сосудов. 4, Мочевой пузырь. 5, Подвздошно-поясничная мышца. 6, Портняжной мышцы. 7, Шейки бедренной кости. 8, Внутренней запирательной мышц. 9, Большая ягодичная мышца мышцы.
2, Прямая мышцы бедра мышцы. 3, Бедренных кровеносных сосудов. 4, Мочевой пузырь. 5, Подвздошно-поясничная мышца. 6, Портняжной мышцы. 7, Шейки бедренной кости. 8, Внутренней запирательной мышц. 9, Большая ягодичная мышца мышцы.
1, мышца—напрягатель широкой фасции бедра. 2, Прямая мышцы бедра мышцы. 3, Бедренных кровеносных сосудов. 4, Портняжной мышцы. 5, Большая ягодичная мышца мышцы. 6, Внутренней запирательной мышц. 7, квадратная мышца бедра. 8, Большого вертела.
1, Шейки бедренной кости. 2, Бедренных кровеносных сосудов. 3, Мочевой пузырь. 4, Портняжной мышцы. 5, Прямая мышцы бедра мышцы. 6, мышца—напрягатель широкой фасции бедра. 7, Большая ягодичная мышца мышцы. 8, Внутренней запирательной мышц.
1, Прямая мышцы бедра мышцы. 2, Бедренных кровеносных сосудов. 3, Гребенчатая мышца. 4, Портняжной мышцы. 5, Подвздошно-поясничная мышца. 6, квадратная мышца бедра. 7, Внутренней запирательной мышц. 8, Большая ягодичная мышца мышцы.
7, Внутренней запирательной мышц. 8, Большая ягодичная мышца мышцы.
1, мышца—напрягатель широкой фасции бедра. 2, Прямая мышцы бедра мышцы. 3, Портняжной мышцы. 4, Лонного сочленения. 5, Бедренных кровеносных сосудов. 6, Подвздошно-поясничная мышца. 7, квадратная мышца бедра. 8, Внутренней запирательной мышц. 9, Большая ягодичная мышца мышцы.
1, мышца—напрягатель широкой фасции бедра. 2, Прямая мышцы бедра мышцы. 3, Лонного сочленения. 4, Подвздошно-поясничная мышца. 5, квадратная мышца бедра. 6, Наружная запирательная мышца. 7, Внутренней запирательной мышц. 8, Большая ягодичная мышца мышцы.
1, мышца—напрягатель широкой фасции бедра. 2, Прямая мышцы бедра мышцы. 3, Наружная запирательная мышца. 4, Лонного сочленения. 5, Гребенчатая мышца. 6, Подвздошно-поясничная мышца. 7, квадратная мышца бедра. 8, Внутренней запирательной мышц. 9, Большая ягодичная мышца мышцы.
1, мышца—напрягатель широкой фасции бедра. 2, Подвздошно-поясничная мышца. 3, Бедренных кровеносных сосудов. 4, Портняжной мышцы. 5, Прямая мышцы бедра мышцы. 6, квадратная мышца бедра. 7, Наружная запирательная мышца. 8, Седалищный бугор. 9, Полуперепончатой мышцы. 10, Большая ягодичная мышца мышцы.
1, мышца—напрягатель широкой фасции бедра. 2, Подвздошно-поясничная мышца. 3, Гребенчатая мышца. 4, Лонного сочленения. 5, Портняжной мышцы. 6, Прямая мышцы бедра мышцы. 7, латеральная широкая мышца бедра. 8, Наружная запирательная мышца. 9, квадратная мышца бедра. 10, Большая ягодичная мышца мышцы.
1, мышца—напрягатель широкой фасции бедра. 2, Прямая мышцы бедра мышцы. 3, Портняжной мышцы. 4, Гребенчатая мышца. 5, Бедренных кровеносных сосудов. 6, Подвздошно-поясничная мышца. 7, латеральная широкая мышца бедра. 8, Большая ягодичная мышца мышцы.
Мышечная система – 3D Атлас анатомии – Кости и мышцы человеческого тела — Muscular System — 3D Atlas of Anatomy (1.
 3.2)
3.2)Мышечная система – это атлас анатомии нового поколения в 3D, в котором вы найдете интерактивные и очень подробные анатомические модели!
При выборе моделей или значков вам будут показаны термины, относящиеся к любой специфической анатомической части, вы можете выбрать из 10 языков и показ терминов на двух языках одновременно.
Мышечная система – это полезный инструмент для студентов, которые получают образование в сфере медицины и физической культуры, для терапевтов, ортопедов, физиотерапевтов, психотерапевтов, медработников, кинезиологов, медсестер и спортивные тренеров.
ОЧЕНЬ ПОДРОБНЫЕ АНАТОМИЧЕСКИЕ 3D-МОДЕЛИ
• Костная система
• Мышечная система
• Точное 3D-моделирование
• Поверхность скелета с высоким разрешением текстур до стандарта 4K
ПРОСТОЙ И ПОНЯТНЫЙ ИНТЕРФЕЙС.
• Вращайте и приближайте каждую модель в трехмерном пространстве
• Мышцы сгруппированы по слоям, от поверхностных до самых глубоких
• Визуализация мышечных слоев в нескольких или одной режиме
• Возможность спрятать каждую кость или мышцу по отдельности
• Функция фильтра для того, чтобы скрыть или отобразить каждую систему
• Умное вращение, автоматически двигает центр вращения для более легкой навигации
• Интерактивный значок позволяет визуализировать термин, который имеет отношение к каждой анатомической детали
• Скрыть / Показать интерфейс, идеально подходит для использования на смартфонах
МНОГОЯЗЫЧНОСТЬ
• Анатомические термины и интерфейс доступны 10 языках: английский, французский, немецкий, итальянский, португальский, русский, китайский, японский и корейский
• Язык может быть выбран непосредственно через интерфейс приложения
• Анатомические термины могут быть показаны на двух языках одновременно
Мышечная система – это часть коллекции приложений для изучения анатомии человека 3D Атлас анатомии, новых приложений и обновлений, которые были разработаны.
Мышечная система – 3D Атлас анатомии – Кости и мышцы человеческого тела — Muscular System
Мышечная система – это атлас анатомии нового поколения в 3D, в котором вы найдете интерактивные и очень подробные анатомические модели!Вся костно-мышечная система была реконструирована в 3D, вы можете вращать и приближать каждую модель, а также подробно рассматривать модели под любым углом.
При выборе моделей или значков вам будут показаны термины, относящиеся к любой специфической анатомической части, вы можете выбрать из 11 языков и показ терминов на двух языках одновременно.
Мышечная система – это полезный инструмент для студентов, которые получают образование в сфере медицины и физической культуры, для терапевтов, ортопедов, физиотерапевтов, психотерапевтов, медработников, кинезиологов, медсестер и спортивные тренеров.
ОЧЕНЬ ПОДРОБНЫЕ АНАТОМИЧЕСКИЕ 3D-МОДЕЛИ
• Костная система
• Мышечная система
• Точное 3D-моделирование
• Поверхность скелета с высоким разрешением текстур до стандарта 4K
ПРОСТОЙ И ПОНЯТНЫЙ ИНТЕРФЕЙС.
• Вращайте и приближайте каждую модель в трехмерном пространстве
• Разделение по районам для понятной и быстрой визуализации каждой структуры
• Мышцы сгруппированы по слоям, от поверхностных до самых глубоких
• Визуализация мышечных слоев в нескольких или одной режиме
• Возможность спрятать каждую кость или мышцу по отдельности
• Функция фильтра для того, чтобы скрыть или отобразить каждую систему
• Умное вращение, автоматически двигает центр вращения для более легкой навигации
• Интерактивный значок позволяет визуализировать термин, который имеет отношение к каждой анатомической детали
• Скрыть / Показать интерфейс, идеально подходит для использования на смартфонах
• Описания мышц (происхождение, вставка, иннервации, действие), на английском
МНОГОЯЗЫЧНОСТЬ
• Анатомические термины и интерфейс доступны 11 языках: Латинский, английский, французский, немецкий, итальянский, португальский, русский, китайский, японский и корейский
• Язык может быть выбран непосредственно через интерфейс приложения
• Анатомические термины могут быть показаны на двух языках одновременно
Мышечная система – это часть коллекции приложений для изучения анатомии человека 3D Атлас анатомии, новых приложений и обновлений, которые были разработаны.
мышц — Атлас анатомии
Атлас анатомии
2 мышцыМышцы спины: обзор
Мышцы спины делятся на две группы: внешние и внутренние мышцы, которые разделены поверхностным слоем грудопоясничной фасции. Поверхностные внешние мышцы считаются мышцами верхней конечности, которые переместились в спину; эти мышцы обсуждаются в Блоке 4.
Фиг.2.1 Поверхностные (внешние) мышцы спины
Вид сзади. Удалено: Трапеции и широчайшие мышцы спины (справа). Выявлено: Грудно-поясничная фасция. Примечание: Поверхностный слой грудопоясничной фасции усилен апоневротическим началом широчайшей мышцы спины.
Рис. 2.2 Грудно-поясничная фасция
Поперечный разрез, вид сверху. Внутренние мышцы спины изолированы в костно-фиброзном канале, образованном грудопоясничной фасцией, дугами позвонков, а также остистыми и поперечными отростками связанных позвонков. Грудопоясничная фасция состоит из поверхностного и глубокого слоев, которые соединяются на латеральном крае внутренних мышц спины. В области шеи поверхностный слой сливается с затылочной фасцией (глубокий слой), переходя в шейную фасцию (превертебральный слой).
Грудопоясничная фасция состоит из поверхностного и глубокого слоев, которые соединяются на латеральном крае внутренних мышц спины. В области шеи поверхностный слой сливается с затылочной фасцией (глубокий слой), переходя в шейную фасцию (превертебральный слой).
Внутренние мышцы шейного отдела позвоночника
Рис. 2.3 Мышцы в затылочной области
Вид сзади. Удалено: Трапециевидная, грудинно-ключично-сосцевидная, голеностопная и полуостистая мышцы (справа). Выявлено: Затылочные мышцы (справа).
Рис. 2.4 Короткие затылочные мышцы
Вид сзади. См. Рис. 2.6 .
Внутренние мышцы спины
Внешние мышцы спины (трапециевидные, широчайшие мышцы спины, поднимающие лопатки и ромбовидные мышцы) обсуждаются в блоке 4. Задняя зубчатая мышца, считающаяся промежуточной внешней мышцей спины, была включена в состав поверхностных внутренних мышц в этом блоке.
Рис. 2.5 Внутренние мышцы спины
Вид сзади. Последовательное рассечение грудопоясничной фасции, поверхностных внутренних мышц, промежуточных внутренних мышц и глубоких внутренних мышц спины.
Факты о мышцах (I)
Рис. 2.6 Короткие мышцы затылочного и краниовертебрального суставов
Фиг.2,7 Превертебральные мышцы
Факты о мышцах (II)
Внутренние мышцы спины делятся на поверхностный, промежуточный и глубокий слои. Задние зубчатые мышцы — это внешние мышцы спины, иннервируемые брюшными ветвями межреберных нервов, а не спинными ветвями, которые иннервируют внутренние мышцы спины. Они включены сюда, поскольку встречаются при рассечении мускулатуры спины.
Фиг.2,8 Поверхностные внутренние мышцы спины (схема)
Правая сторона, вид сзади.
Рис. 2.9 Промежуточные внутренние мышцы спины (схема)
Правая сторона, вид сзади. Эти мышцы известны как мышцы, выпрямляющие позвоночник.
Рис. 2.10 Поверхностные и промежуточные внутренние мышцы спины
Вид сзади.
Факты о мышцах (III)
Глубокие внутренние мышцы спины делятся на две группы: трансверсоспинальные и глубокие сегментарные мышцы. Transversospinalis мышцы проходят между поперечными и остистыми отростками позвонков.
Рис. 2.11 Transversospinalis мышцы (схема)
Вид сзади.
Рис. 2.12 Глубокие сегментарные мышцы (схема)
Вид сзади.
Рис. 2.13 Глубокие внутренние мышцы спины
Вид сзади.
Новый атлас экспрессии генов в скелетных мышцах человека раскрывает молекулярные изменения, связанные со старением | Скелетная мышца
Delmonico MJ, Harris TB, Visser M, Park SW, Conroy MB, Velasquez-Mieyer P, et al. Продольное исследование мышечной силы, качества и инфильтрации жировой ткани. Am J Clin Nutr.2009. 90 (6): 1579–85. DOI: 10.3945 / ajcn.2009.28047.
PubMed Central CAS Статья PubMed Google Scholar
Диркс А.Дж., Хофер Т., Марцетти Э., Пахор М., Левенбург К. Мутации митохондриальной ДНК, энергетический метаболизм и апоптоз в стареющих мышцах. Aging Res Rev.2006; 5 (2): 179–95. DOI: 10.1016 / j.arr.2006.03.002.
CAS Статья PubMed Google Scholar
Ким Т.Н., Цой КМ. Саркопения: определение, эпидемиология и патофизиология. J Bone Metab. 2013; 20 (1): 1–10. DOI: 10.11005 / jbm.2013.20.1.1.
DOI: 10.11005 / jbm.2013.20.1.1.
PubMed Central Статья PubMed Google Scholar
Goodpaster BH, Park SW, Harris TB, Kritchevsky SB, Nevitt M, Schwartz AV, et al. Потеря силы, массы и качества скелетных мышц у пожилых людей: исследование здоровья, старения и состава тела. J Gerontol A Biol Sci Med Sci.2006. 61 (10): 1059–64.
Артикул PubMed Google Scholar
Зан Дж. М., Сону Р., Фогель Х., Крейн Е., Мазан-Мамчарз К., Рабкин Р. и др. Транскрипционное профилирование старения в мышцах человека выявляет общий признак старения. PLoS Genet. 2006; 2 (7): e115. DOI: 10.1371 / journal.pgen.0020115.eor.
PubMed Central Статья PubMed Google Scholar
de Magalhães JP, Curado J, Church GM.Мета-анализ профилей экспрессии возрастных генов позволяет выявить общие признаки старения. Биоинформатика. 2009. 25 (7): 875–81. DOI: 10.1093 / биоинформатика / btp073.
Биоинформатика. 2009. 25 (7): 875–81. DOI: 10.1093 / биоинформатика / btp073.
PubMed Central Статья PubMed Google Scholar
Буа Е., Джонсон Дж., Хербст А., Делонг Б., Маккензи Д., Саламат С. и др. Мутации с делециями митохондриальной ДНК накапливаются внутриклеточно до вредных уровней в волокнах скелетных мышц пожилого человека. Am J Hum Genet.2006. 79 (3): 469–80. DOI: 10,1086 / 507132.
PubMed Central CAS Статья PubMed Google Scholar
Остин С., Сен-Пьер Дж. PGC1α и митохондриальный метаболизм — новые концепции и актуальность при старении и нейродегенеративных расстройствах. J Cell Sci. 2012; 125 (Pt 21): 4963–71. DOI: 10.1242 / jcs.113662.
CAS Статья PubMed Google Scholar
Mootha VK, Lindgren CM, Eriksson KF, Subramanian A, Sihag S, Lehar J, et al. PGC-1alpha-чувствительные гены, участвующие в окислительном фосфорилировании, координированно подавляются при диабете человека. Нат Жене. 2003. 34 (3): 267–73.
PGC-1alpha-чувствительные гены, участвующие в окислительном фосфорилировании, координированно подавляются при диабете человека. Нат Жене. 2003. 34 (3): 267–73.
CAS Статья PubMed Google Scholar
Патти М.Э., Бьют А.Дж., Кранкхорн С., Куси К., Берриа Р., Кашьяп С. и др. Скоординированное снижение генов окислительного метаболизма у людей с инсулинорезистентностью и диабетом: потенциальная роль PGC1 и NRF1.Proc Natl Acad Sci U S. A. 2003; 100 (14): 8466–71. DOI: 10.1073 / pnas.10320.
PubMed Central CAS Статья PubMed Google Scholar
Bodine SC, Stitt TN, Gonzalez M, Kline WO, Stover GL, Bauerlein R, et al. Путь Akt / mTOR является важным регулятором гипертрофии скелетных мышц и может предотвращать атрофию мышц in vivo. Nat Cell Biol. 2001. 3 (11): 1014–9. DOI: 10.1038 / NCB1101-1014.
CAS Статья PubMed Google Scholar

Ли М.Н., Ха Ш, Ким Дж., Ко А., Ли С.С., Ким Дж. Х. и др. Гликолитический поток передает сигнал mTOR через регуляцию Rheb, опосредованную глицеральдегид-3-фосфатдегидрогеназой. Mol Cell Biol. 2009. 29 (14): 3991–4001. DOI: 10.1128 / MCB.00165-09.
PubMed Central CAS Статья PubMed Google Scholar
Лукк М., Капушески М., Никкила Дж., Паркинсон Х., Гонсалвес А., Хубер В. и др. Глобальная карта экспрессии генов человека. Nat Biotechnol.2010. 28 (4): 322–4. DOI: 10.1038 / nbt0410-322.
PubMed Central CAS Статья PubMed Google Scholar
Чжэн-Брэдли X, Рунг Дж., Паркинсон Х., Бразма А. Крупномасштабное сравнение глобальных паттернов экспрессии генов у человека и мыши. Genome Biol. 2010; 11 (12): R124. DOI: 10.1186 / ГБ-2010-11-12-r124.
PubMed Central CAS Статья PubMed Google Scholar
Ojala KA, Kilpinen SK, Kallioniemi OP. Классификация неизвестных первичных опухолей с помощью метода, основанного на данных, на основе большой справочной базы данных микрочипов. Genome Med. 2011; 3 (9): 63. DOI: 10,1186 / GM279.
PubMed Central Статья PubMed Google Scholar
Килпинен С., Аутио Р., Ояла К., Ильин К., Бухер Е., Сара Х. и др. Систематический биоинформатический анализ уровней экспрессии 17 330 человеческих генов в 9 783 образцах из 175 типов здоровых и патологических тканей.Genome Biol. 2008; 9 (9): R139. DOI: 10.1186 / ГБ-2008-9-9-r139.
PubMed Central Статья PubMed Google Scholar
Rung J, Brazma A. Повторное использование общегеномных данных по экспрессии генов. Nat Rev Genet. 2013. 14 (2): 89–99. DOI: 10,1038 / nrg3394.
CAS Статья PubMed Google Scholar

Рустичи Г., Колесников Н., Брандизи М., Бурдетт Т., Дилаг М., Эмам И. и др.Обновление ArrayExpress — тенденции роста базы данных и ссылки на инструменты анализа данных. Nucleic Acids Res. 2013; 41 (выпуск базы данных): D987–90. DOI: 10.1093 / nar / gks1174.
PubMed Central CAS Статья PubMed Google Scholar
Лахти Л., Эло Л.Л., Аиттокаллио Т., Каски С. Вероятностный анализ надежности зонда в исследованиях дифференциальной экспрессии генов с короткими массивами олигонуклеотидов. IEEE / ACM Trans Comput Biol Bioinform.2011; 8 (1): 217–25. DOI: 10.1109 / TCBB.2009.38.
Артикул PubMed Google Scholar
Лахти Л., Торренте А., Эло Л.Л., Бразма А., Рунг Дж. Полностью масштабируемый онлайн-алгоритм предварительной обработки для коротких атласов олигонуклеотидных микрочипов. Nucleic Acids Res. 2013; 41 (10), с110. DOI: 10.1093 / nar / gkt229.
PubMed Central CAS Статья PubMed Google Scholar
Кауфманн А., Джентльмен Р., Хубер В. arrayQualityMetrics — пакет биопроводников для оценки качества данных микрочипов. Биоинформатика. 2009. 25 (3): 415–6. DOI: 10.1093 / биоинформатика / btn647.
PubMed Central CAS Статья PubMed Google Scholar
Консорциум GTEx. Проект «Экспрессия генотипа-ткани» (GTEx). Нат Жене. 2013. 45 (6): 580–5. 10.1038 / ng.2653.
Артикул Google Scholar
Робинсон MD, Ошлак А. Метод масштабной нормализации для анализа дифференциальной экспрессии данных РНК-seq. Genome Biol. 2010; 11 (3): R25. DOI: 10.1186 / ГБ-2010-11-3-r25.
PubMed Central Статья PubMed Google Scholar
Руди Дж., Валафар Ф. Эмпирическое сравнение кроссплатформенных методов нормализации данных экспрессии генов. BMC Bioinformatics. 2011; 12: 467. DOI: 10.1186 / 1471-2105-12-467.
BMC Bioinformatics. 2011; 12: 467. DOI: 10.1186 / 1471-2105-12-467.
PubMed Central Статья PubMed Google Scholar
Парих Х., Нильссон Э., Линг С., Поулсен П., Альмгрен П., Ниттби Х. и др. Молекулярные корреляты максимального поглощения кислорода и волокон типа 1. Am J Physiol Endocrinol Metab. 2008. 294 (6): E1152–9. DOI: 10.1152 / ajpendo.
.2008.CAS Статья PubMed Google Scholar
Хорн Д., Чжоу В., Тревиссон Е., Аль-Али Х., Харрис Т.К., Сальвиати Л. и др. Консервативный митохондриальный близнец Cx9C белок Cmc2 является гомологом Cmc1, необходимым для биогенеза цитохром с оксидазы.J Biol Chem. 2010. 285 (20): 15088–99. DOI: 10.1074 / jbc.M110.104786.
PubMed Central CAS Статья PubMed Google Scholar
Секерес Ф., Чадт А., Том Р.З., Дешмук А.С., Чибалин А. В., Бьёрнхольм М. и др. Белок TBC1D1, активирующий Rab-GTPase, регулирует метаболизм глюкозы в скелетных мышцах. Am J Physiol Endocrinol Metab. 2012; 303 (4): E524–33. DOI: 10.1152 / ajpendo.00605.2011.
В., Бьёрнхольм М. и др. Белок TBC1D1, активирующий Rab-GTPase, регулирует метаболизм глюкозы в скелетных мышцах. Am J Physiol Endocrinol Metab. 2012; 303 (4): E524–33. DOI: 10.1152 / ajpendo.00605.2011.
CAS Статья PubMed Google Scholar
An D, Toyoda T, Taylor EB, Yu H, Fujii N, Hirshman MF и др. TBC1D1 регулирует транспорт глюкозы, индуцированный инсулином и сокращением, в скелетных мышцах мышей. Сахарный диабет. 2010. 59 (6): 1358–65. DOI: 10.2337 / db09-1266.
PubMed Central CAS Статья PubMed Google Scholar
Ван Н., Ян Ц., Се Ф, Сун Л., Су Х, Ван И и др. Gadd45α: новый ген, связанный с диабетом, потенциально связывающий диабетическую кардиомиопатию и дисфункцию барорефлекса.PLoS ONE. 2012; 7 (12), e49077. DOI: 10.1371 / journal.pone.0049077.
PubMed Central CAS Статья PubMed Google Scholar

Споррер Д., Вебер М., Ваннингер Дж., Вейгерт Дж., Ноймайер М., Штёгбауэр Ф. и др. Адипонектин подавляет активность CD163, клеточные и растворимые формы которого повышаются при ожирении. Eur J Clin Invest. 2009. 39 (8): 671–9. DOI: 10.1111 / j.1365-2362.2009.02170.x.
CAS Статья PubMed Google Scholar
Бонгерс К.С., Фокс Д.К., Эберт С.М., Кункель С.Д., Дайл М.С., Буллард С.А. и др. Денервация скелетных мышц вызывает атрофию скелетных мышц посредством пути, в котором участвуют как Gadd45a, так и HDAC4. Am J Physiol Endocrinol Metab. 2013; 305 (7): E907–15. DOI: 10.1152 / ajpendo.00380.2013.
PubMed Central CAS Статья PubMed Google Scholar
Cheng Y, Wang Y, Li Y, Deng Y, Hu J, Mo X и др. Новый человеческий ген ZNF415 с пятью изоформами ингибирует AP-1- и p53-опосредованную транскрипционную активность.Biochem Biophys Res Commun. 2006; 351 (1): 33–9. DOI: 10.1016 / j.bbrc.2006.09.161.
2006; 351 (1): 33–9. DOI: 10.1016 / j.bbrc.2006.09.161.
CAS Статья PubMed Google Scholar
Møller HJ, Frikke-Schmidt R, Moestrup SK, Nordestgaard BG, Tybjrg-Hansen A. Растворимый в сыворотке CD163 предсказывает риск диабета 2 типа в общей популяции. Clin Chem. 2011; 57 (2): 291–7. DOI: 10.1373 / Clinchem.2010.154724.
Артикул PubMed Google Scholar
Short KR, Bigelow ML, Kahl J, Singh R, Coenen-Schimke J, Raghavakaimal S, et al. Снижение митохондриальной функции скелетных мышц у людей с возрастом. Proc Natl Acad Sci U S. A. 2005; 102 (15): 5618–23. DOI: 10.1073 / pnas.0501559102.
PubMed Central CAS Статья PubMed Google Scholar
Джонсон М.Л., Робинсон М.М., Наир К.С. Старение скелетных мышц и митохондрии. Trends Endocrinol Metab. 2013; 24 (5): 247–56. DOI: 10.1016 / j.tem.2012.12.003.
DOI: 10.1016 / j.tem.2012.12.003.
PubMed Central CAS Статья PubMed Google Scholar
Conley KE, Jubrias SA, Esselman PC. Окислительная способность и старение в мышцах человека. J Physiol. 2000. 526 (Pt 1): 203–10.
PubMed Central CAS Статья PubMed Google Scholar
McDermott-Roe C, Ye J, Ahmed R, Sun XM, Serafín A, Ware J, et al.Эндонуклеаза G представляет собой новый фактор, определяющий гипертрофию сердца и функцию митохондрий. Природа. 2011. 478 (7367): 114–8. DOI: 10,1038 / природа10490.
PubMed Central CAS Статья PubMed Google Scholar
Humphries AD, Streimann IC, Stojanovski D, Johnston AJ, Yano M, Hoogenraad NJ, et al. Рассечение митохондриального пути импорта и сборки Tom40 человека. J Biol Chem. 2005. 280 (12): 11535–43. DOI: 10.1074 / jbc. M413816200.
M413816200.
CAS Статья PubMed Google Scholar
Джозеф А.М., Любичич В., Адхихетти П.Дж., Худ Д.А. Биогенез митохондриального канала Tom40 в скелетных мышцах старых животных и его приспособляемость к хронической сократительной активности. Am J Physiol Cell Physiol. 2010. 298 (6): C1308–14. DOI: 10.1152 / ajpcell.00644.2008.
CAS Статья PubMed Google Scholar
Schaefer L, Ballabio A, Zoghbi HY. Клонирование и характеристика предполагаемого гена синтетазы холоцитохрома с-типа (HCCS) человека, выделенного из критической области микрофтальмии с линейными дефектами кожи (MLS). Геномика. 1996. 34 (2): 166–72. DOI: 10.1006 / geno.1996.0261.
CAS Статья PubMed Google Scholar
Исии К.А., Фумото Т., Иваи К., Такешита С., Ито М., Шимохата Н. и др. Координация захвата PGC-1beta и железа в митохондриальном биогенезе и активации остеокластов. Nat Med. 2009. 15 (3): 259–66. DOI: 10,1038 / нм.1910.
Nat Med. 2009. 15 (3): 259–66. DOI: 10,1038 / нм.1910.
CAS Статья PubMed Google Scholar
Willcox BJ, Donlon TA, He Q, Chen R, Grove JS, Yano K, et al. Генотип FOXO3A тесно связан с долголетием человека. Proc Natl Acad Sci U S. A. 2008; 105 (37): 13987–92. DOI: 10.1073 / pnas.0801030105.
PubMed Central CAS Статья PubMed Google Scholar
Сандри М., Сандри С., Гилберт А., Скурк С., Калабрия Е., Пикард А. и др. Факторы транскрипции Foxo индуцируют связанную с атрофией убиквитинлигазу атрогин-1 и вызывают атрофию скелетных мышц. Клетка. 2004. 117 (3): 399–412.
PubMed Central CAS Статья PubMed Google Scholar
Гамильтон С.Р., Стэплтон Д., О’Доннелл Дж. Б., Кунг Дж. Т., Далал С. Р., Кемп Б. Е. и др. Активирующая мутация в субъединице gamma1 AMP-активируемой протеинкиназы. FEBS Lett. 2001. 500 (3): 163–8.
FEBS Lett. 2001. 500 (3): 163–8.
CAS Статья PubMed Google Scholar
Reznick RM, Zong H, Li J, Morino K, Moore IK, Yu HJ, et al. Связанное со старением снижение активности АМФ-активированной протеинкиназы и митохондриального биогенеза. Cell Metab. 2007. 5 (2): 151–6. DOI: 10.1016 / j.cmet.2007.01.008.
PubMed Central CAS Статья PubMed Google Scholar
Kulkarni SS, Karlsson HK, Szekeres F, Chibalin AV, Krook A, Zierath JR. Подавление ферментов 5′-нуклеотидазы способствует фосфорилированию и метаболизму AMP-активированной протеинкиназы (AMPK) в скелетных мышцах человека и мыши. J Biol Chem. 2011. 286 (40): 34567–74. DOI: 10.1074 / jbc.M111.268292.
PubMed Central CAS Статья PubMed Google Scholar
Рихтер Е.А., Харгривз М. Упражнения, GLUT4 и поглощение глюкозы скелетными мышцами. Physiol Rev.2013; 93 (3): 993–1017. DOI: 10.1152 / Physrev.00038.2012.
Physiol Rev.2013; 93 (3): 993–1017. DOI: 10.1152 / Physrev.00038.2012.
CAS Статья PubMed Google Scholar
Broskey NT, Greggio C, Boss A, Boutant M, Dwyer A, Schlueter L, et al. Митохондрии скелетных мышц у пожилых людей: влияние физической подготовки и физических упражнений. J Clin Endocrinol Metab. 2014. 99 (5): 1852–61. DOI: 10.1210 / jc.2013-3983.
CAS Статья PubMed Google Scholar
Dey BR, Spence SL, Nissley P, Furlanetto RW. Взаимодействие человеческого супрессора передачи сигналов цитокинов (SOCS) -2 с рецептором инсулиноподобного фактора роста-I. J Biol Chem. 1998. 273 (37): 24095–101.
CAS Статья PubMed Google Scholar
Хаддад Ф., Адамс Г.Р. Чувствительные к старению клеточные и молекулярные механизмы, связанные с гипертрофией скелетных мышц. J. Appl Physiol (1985). 2006. 100 (4): 1188–203. DOI: 10.1152 / japplphysiol.01227.2005.
Appl Physiol (1985). 2006. 100 (4): 1188–203. DOI: 10.1152 / japplphysiol.01227.2005.
CAS Статья Google Scholar
Буфорд Т.В., Кук МБ, Уиллоуби Д.С. Изменения экспрессии воспалительных генов в скелетных мышцах человека, вызванные упражнениями с отягощениями. Eur J Appl Physiol. 2009. 107 (4): 463–71. DOI: 10.1007 / s00421-009-1145-z.
CAS Статья PubMed Google Scholar
Атлас мышц — РНКанализ скелетных мышц
Почему это приложение?
Скелетная мышца состоит в основном из мышечных волокон, но также включает в себя кровеносные и лимфатические сосуды, нервы и иммунные клетки.Таким образом, образцы, взятые из биопсии цельной мышцы, представляют собой смесь уровней экспрессии всех различных клеток, составляющих ткань. Поскольку каждый из этих типов клеток потенциально может реагировать на экспериментальные условия, трудно расшифровать вклад мышечных клеток в общий ответ ткани.
Этот инструмент позволяет оценивать уровни мРНК генов в мышечной ткани и сравнивать их с изолированными мышечными волокнами и несколькими первичными типами клеток, присутствующими в мышечной ткани in vivo.Поскольку подходы с использованием первичных клеток полезны для исследования специфической реакции миотрубок, этот инструмент направлен на обеспечение первой оценки относительного уровня экспрессии генов в мышечных клетках по сравнению с другими типами клеток, присутствующими в биопсиях мышц.
В качестве примера известно, что поверхностный маркер CD14 экспрессируется на циркулирующих моноцитах. Действительно, инструмент оценивает, что экспрессия мРНК CD14 выше в крови и первичных макрофагах по сравнению с другими типами клеток. Экспрессия эндотелина (EDN1) ограничена эндотелием, и инструмент действительно обнаруживает, что она намного выше в эндотелиальных клетках по сравнению с мышцами или кровью.Напротив, уровни мРНК тяжелой цепи 2 миозина (MYh3) очень высоки в биоптатах мышц, волокнах и клетках и почти отсутствуют в крови, иммунных и эндотелиальных клетках.
В заключение, этот инструмент полезен для быстрого сравнения обогащения вашего любимого гена одним типом клеток по сравнению с другим. Однако данные должны быть подтверждены в ваших собственных клетках и в ваших условиях с помощью количественной ПЦР или Вестерн-блоттинга.
https://github.com/NicoPillon/Muscle_Atlas
От общедоступных наборов данных к атласу выражений
Этот инструмент был создан путем сбора общедоступных данных массива генов об уровнях экспрессии мРНК в тканях и клетках человека.Для каждого массива сохранялись и обрабатывались только базальные / контрольные условия для определения базовых уровней экспрессии. Затем данные были закодированы и загружены в блестящее приложение, созданное для упрощения опроса базы данных.
Протеом человека в скелетных мышцах
Основная функция скелетных мышц — сокращение, которое обеспечивает устойчивость и движение тела. Скелетная мышца состоит из поперечно-полосатых мышечных клеток, которые сливаются в длинные мышечные волокна.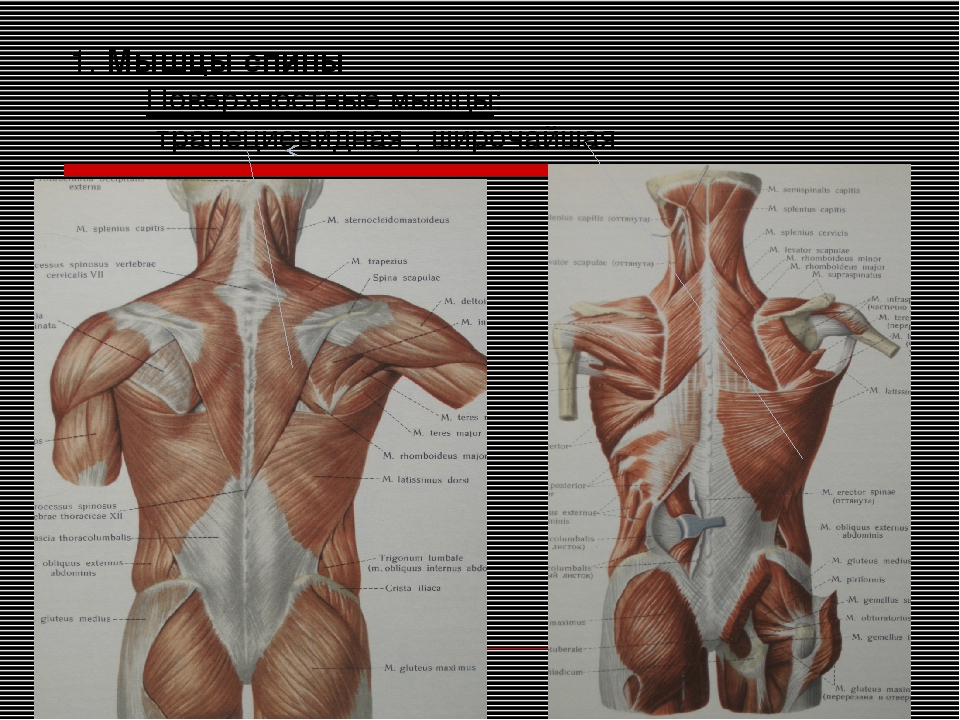 Анализ транскриптома показывает, что 70% (n = 13839) всех белков человека (n = 19670) экспрессируются в скелетных мышцах, и 907 из этих генов демонстрируют повышенную экспрессию в скелетных мышцах по сравнению с другими типами тканей.
Анализ транскриптома показывает, что 70% (n = 13839) всех белков человека (n = 19670) экспрессируются в скелетных мышцах, и 907 из этих генов демонстрируют повышенную экспрессию в скелетных мышцах по сравнению с другими типами тканей.
- 907 повышенных генов
- 111 обогащенных генов
- 202 групповых обогащенных гена
- Скелетная мышца имеет наибольшую групповую экспрессию генов, общих с языком
Транскриптом скелетных мышц
Анализ транскриптома скелетных мышц может быть визуализирован в отношении специфичности и распределения транскрибируемых молекул мРНК (рис. 1).Специфичность показывает количество генов с повышенной или не повышенной экспрессией в скелетных мышцах по сравнению с другими тканями. Повышенное выражение включает три подкатегории типа повышенного выражения:
- Обогащенная ткань: как минимум в четыре раза выше уровень мРНК в скелетных мышцах по сравнению с любыми другими тканями.
- Обогащенная группа: по крайней мере, в четыре раза выше средний уровень мРНК в группе из 2-5 тканей по сравнению с любой другой тканью.

- Улучшенная ткань: по крайней мере, в четыре раза выше уровень мРНК в скелетных мышцах по сравнению со средним уровнем во всех других тканях.
Распределение, с другой стороны, показывает, сколько генов имеют или не имеют обнаруживаемых уровней (NX≥1) транскрибированных молекул мРНК в скелетных мышцах по сравнению с другими тканями. Как видно из Таблицы 1, все гены, повышенные в скелетных мышцах, классифицируются как:
- Обнаружен в одиночном: обнаружен в единственной ткани
- Обнаружен в некоторых: Обнаружен более чем в одной, но менее чем в одной трети тканей
- Обнаружен во многих: обнаружен как минимум в трети, но не во всех тканях
- Обнаружен во всех: Обнаружен во всех тканях
Рисунок 1.(A) Распределение всех генов по пяти категориям на основе специфичности транскрипта в скелетных мышцах, а также во всех других тканях. (B) Распределение всех генов по шести категориям на основе обнаружения транскриптов (NX? 1) в скелетных мышцах, а также во всех других тканях.
Как показано на рисунке 1, 907 генов демонстрируют некоторый уровень повышенной экспрессии в скелетных мышцах по сравнению с другими тканями. Три категории генов с повышенной экспрессией в скелетных мышцах по сравнению с другими органами показаны в таблице 1.В таблице 2 определены 12 генов с наибольшим обогащением в скелетных мышцах.
Таблица 1. Количество генов в подразделяемых категориях повышенной экспрессии в скелетных мышцах.
Таблица 2. 12 генов с наивысшим уровнем экспрессии в скелетных мышцах. «Тканевое распределение» описывает обнаружение транскрипта (NX? 1) в скелетных мышцах, а также во всех других тканях. «мРНК (ткань)» показывает уровень транскрипта в скелетных мышцах в виде значений NX.«Оценка тканевой специфичности (TS)» соответствует кратному изменению между уровнем экспрессии в скелетных мышцах и тканях со вторым по величине уровнем экспрессии.
| Джин | Описание | Распределение тканей | мРНК (ткань) |  «> Оценка тканевой специфичности «> Оценка тканевой специфичности |
|---|---|---|---|---|
| IDI2 | изопентенилдифосфат-дельта-изомераза 2 | Обнаружен в некоторых | 141.6 | 125 |
| DUPD1 | домен фосфатазы двойной специфичности и произомеразы, содержащий 1 | Обнаружен в одиночном | 38,9 | 71 |
| MYh2 | тяжелая цепь миозина 1 | Обнаружен в некоторых | 428,9 | 31 |
| LRRC30 | богатый лейцином повтор, содержащий 30 | Обнаружен в одиночном | 18.7 | 30 |
| MYh5 | тяжелая цепь миозина 4 | Обнаружен в одиночном | 15,1 | 29 |
| SMTNL1 | Smoothelin как 1 | Обнаружен в некоторых | 118,9 | 28 |
| ACTN3 | актинин альфа 3 (ген / псевдоген) | Обнаружен в некоторых | 185,5 | 24 |
| PPP1R27 | Регуляторная субъединица протеинфосфатазы 1 27 | Обнаружен в некоторых | 211. 0 0 | 22 |
| MYADML2 | маркер дифференцировки, связанный с миелоидом, такой как 2 | Обнаружен в некоторых | 44,5 | 20 |
| АНКРД23 | анкириновый повторяющийся домен 23 | Обнаружен в некоторых | 184,5 | 15 |
| UCP3 | разобщающий белок 3 | Обнаружен в некоторых | 99.6 | 15 |
| CHRNA10 | холинергический рецептор никотиновая альфа 10 субъединица | Обнаружен в некоторых | 33,6 | 15 |
Повышенная экспрессия белков в скелетных мышцах
Углубленный анализ повышенных генов в скелетных мышцах с использованием профилей белков на основе антител позволил нам визуализировать паттерны экспрессии этих белков в различных функциональных компартментах, включая белки, связанные с i) сокращением, ii) функцией кальция и iii) ферментативной активностью. .
.
Белки, связанные с сокращением, экспрессируются в скелетных мышцах
Первичные структурные белки в скелетных миоцитах, связанные с сокращением, — это миозиновые и актиновые филаменты, образующие полосатый рисунок, который можно наблюдать при электронной микроскопии. Другое семейство белков, связанных с мышечным сокращением, — это семейство тропонинов, регулирующих связывание миозина с актином посредством конформационных изменений, зависящих от концентрации ионов кальция в клетках. Примеры членов семейств миозина и тропонина, экспрессируемых исключительно в скелетных мышцах, включают MYh3 и TNNT1, при этом MYh3 экспрессируется в быстрых (тип II) волокнах, а TNNT1 — в медленных (тип I) волокнах.Другой пример белка, участвующего в сокращении скелетных мышц, — это миозинсвязывающий белок MYBPC1, который влияет на сокращение за счет образования поперечных мостиков в саркомере.
MYh3
TNNT1
MYBPC1
Белки, связанные с функцией кальция, экспрессируются в скелетных мышцах
Как в сердце, так и в скелетных мышцах сокращение зависит от уровня внутриклеточного кальция. Однако, в отличие от кардиомиоцитов, где высвобождение кальция регулируется путем связывания ионов кальция из внешней среды с потенциалозависимыми кальциевыми каналами, скелетные миоциты накапливают кальций в саркоплазматическом ретикулуме до тех пор, пока нейрональный импульс не вызовет приток кальция вдоль миофиламентов. Три примера, связанных с функцией кальция с избирательной экспрессией в скелетных мышцах, — это RYR1, CASQ1 и JPh2. RYR1 — это рецептор рианодина, действующий как канал высвобождения кальция, в то время как CASQ1 необходим для хранения кальция в саркоплазматическом ретикулуме.JPh2 способствует функциональному взаимодействию между клеточной поверхностью и внутриклеточными каналами высвобождения кальция.
Однако, в отличие от кардиомиоцитов, где высвобождение кальция регулируется путем связывания ионов кальция из внешней среды с потенциалозависимыми кальциевыми каналами, скелетные миоциты накапливают кальций в саркоплазматическом ретикулуме до тех пор, пока нейрональный импульс не вызовет приток кальция вдоль миофиламентов. Три примера, связанных с функцией кальция с избирательной экспрессией в скелетных мышцах, — это RYR1, CASQ1 и JPh2. RYR1 — это рецептор рианодина, действующий как канал высвобождения кальция, в то время как CASQ1 необходим для хранения кальция в саркоплазматическом ретикулуме.JPh2 способствует функциональному взаимодействию между клеточной поверхностью и внутриклеточными каналами высвобождения кальция.
RYR1
CASQ1
JPh2
Белки, связанные с ферментативной активностью, экспрессируемой в скелетных мышцах
Ферментативная активность — важная функция физиологии скелетных мышц, которая связана с различными процессами, такими как метаболизм, накопление и регенерация гликогена. Примеры трех белков, участвующих в ферментативной активности с избирательной экспрессией в скелетных мышцах, включают AMPD1, PYGM и ENO3. AMPD1 — это фермент, участвующий в пуриновом нуклеотидном цикле, и он играет важную роль в энергетическом обмене, в то время как фермент PYGM необходим для метаболизма углеводов и гликогенолиза. ENO3 — это изофермент, который, как предполагается, играет роль в развитии и регенерации мышц, с мутациями, связанными с болезнью накопления гликогена.
Примеры трех белков, участвующих в ферментативной активности с избирательной экспрессией в скелетных мышцах, включают AMPD1, PYGM и ENO3. AMPD1 — это фермент, участвующий в пуриновом нуклеотидном цикле, и он играет важную роль в энергетическом обмене, в то время как фермент PYGM необходим для метаболизма углеводов и гликогенолиза. ENO3 — это изофермент, который, как предполагается, играет роль в развитии и регенерации мышц, с мутациями, связанными с болезнью накопления гликогена.
AMPD1
PYGM
ENO3
Экспрессия гена разделяется между скелетными мышцами и другими тканями
В скелетных мышцах экспрессируется 202 гена, обогащенных группами.Обогащенные группы гены определяются как гены, показывающие в 4 раза более высокий средний уровень экспрессии мРНК в группе из 2-5 тканей, включая скелетные мышцы, по сравнению со всеми другими тканями.
Чтобы проиллюстрировать связь ткани скелетных мышц с другими типами тканей, был создан сетевой график, отображающий количество генов с общей экспрессией между различными типами тканей.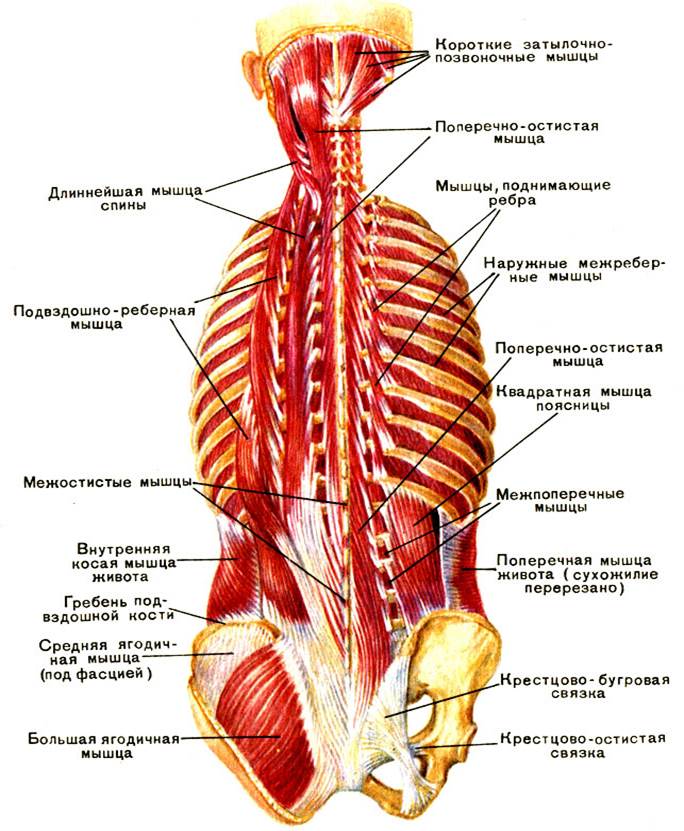
Рис. 2. Интерактивный сетевой график скелетных мышц, обогащенных и группируемых, обогащенных генов, связанных с их соответствующими обогащенными тканями (серые круги).Красные узлы представляют количество генов, обогащенных скелетными мышцами, а оранжевые узлы представляют количество генов, обогащенных группой. Размеры красных и оранжевых узлов связаны с количеством генов, отображаемых в узле. На каждый узел можно щелкнуть, и в результате отображается список всех обогащенных генов, связанных с выделенными краями. Сеть ограничена групповыми обогащенными генами в комбинациях до 5 тканей, но итоговые списки показывают полный набор групповых обогащенных генов в конкретной ткани.
Скелетные мышцы разделяют наибольшую экспрессию генов, обогащенных группой, с сердцем, что ожидается, поскольку и сердце, и скелетные мышцы являются поперечнополосатыми мышцами со многими сходными чертами. Двумя примерами белков с общей экспрессией в сердце и скелетных мышцах являются MYH7 и LDB3. MYH7 связан с сокращением и показывает дифференциальную экспрессию между медленными (тип I) и быстрыми (тип II) мышечными волокнами. LDB3 участвует в организации саркомеров и отчетливо экспрессируется в Z-дисках сердца.
MYH7 связан с сокращением и показывает дифференциальную экспрессию между медленными (тип I) и быстрыми (тип II) мышечными волокнами. LDB3 участвует в организации саркомеров и отчетливо экспрессируется в Z-дисках сердца.
MYH7 — скелетная мышца
MYH7 — сердечная мышца
LDB3 — скелетная мышца
LDB3 — сердечная мышца
Скелетная мышца — один из крупнейших органов человеческого тела, и до 50% общей массы тела составляют скелетные мышцы. Основная функция скелетных мышц — сокращение, которое приводит к движению тела, но также необходимо для осанки и устойчивости тела.В отличие от сердечной мышцы, другой поперечно-полосатой мышцы, похожей по структуре, сокращение скелетных мышц находится под произвольным контролем и инициируется импульсами из мозга. Еще одна важная функция скелетных мышц — регулирование температуры тела. Тепло выделяется, когда мышцы сокращаются и вызывают расширение кровеносных сосудов кожи. Таким образом, скелетные мышцы также участвуют в регуляции кровотока.
Скелетные мышцы вместе с сердечной мышцей состоят из поперечно-полосатой мышечной ткани, образующей параллельные мышечные волокна.Поперечно-полосатая мышечная ткань состоит из миоцитов, расположенных в виде длинных и тонких многоядерных волокон, которые пересекаются правильным рисунком из тонких красных и белых линий, что придает мышце характерный вид и ее название. Существует два типа мышечных волокон (быстрые и медленные) в зависимости от типа присутствующего миозина. Эти типы волокон невозможно различить при обычном окрашивании гематоксилин-эозином (HE).
Развитие и нормальная деятельность скелетных мышц зависят от нервной системы и тесно связаны с ней.Скелетные мышцы прикрепляются к кости и сокращаются добровольно (посредством нервной стимуляции) в отличие от других распространенных типов мышц, то есть сердечной мышцы и гладкой мышцы.
Основным типом клеток скелетных мышц является миоцит. Миоциты сливаются во время развития, образуя большие многоядерные клетки, называемые синцитиями.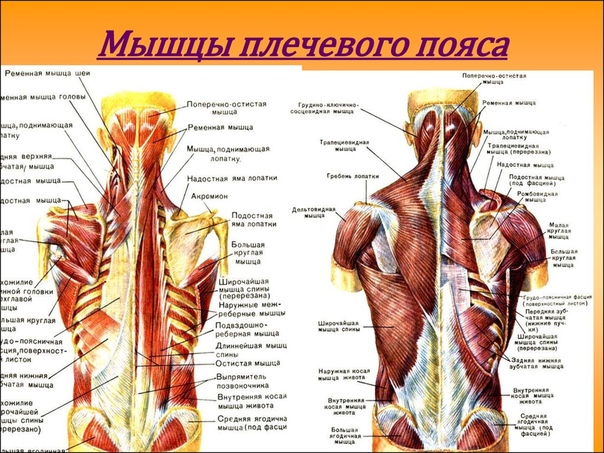 Клетки богаты митохондриями и в значительной степени содержат белки актина и миозина, расположенные в повторяющихся единицах, называемых саркомерами. Гистологически это высокоструктурированное расположение саркомеров выглядит как темные (полосы А) и светлые (полосы I) полосы, которые хорошо видны на микроскопическом изображении.Помимо мышечных волокон, скелетные мышцы также состоят из прилегающих полос соединительной и жировой ткани. Ткань скелетных мышц сильно васкуляризована с тонкой сетью капилляров, проходящих между волокнами.
Клетки богаты митохондриями и в значительной степени содержат белки актина и миозина, расположенные в повторяющихся единицах, называемых саркомерами. Гистологически это высокоструктурированное расположение саркомеров выглядит как темные (полосы А) и светлые (полосы I) полосы, которые хорошо видны на микроскопическом изображении.Помимо мышечных волокон, скелетные мышцы также состоят из прилегающих полос соединительной и жировой ткани. Ткань скелетных мышц сильно васкуляризована с тонкой сетью капилляров, проходящих между волокнами.
Используя световую микроскопию и иммуноокрашивание, мы можем детально изучить и визуализировать сложность скелетных мышц. На видео ниже скелетные мышцы показаны красным цветом, а сложная сеть нервов — бирюзовым. Полная версия видео находится здесь.
Гистологию скелетных мышц человека, включая подробные изображения и информацию о различных типах клеток, можно просмотреть в гистологическом словаре белкового атласа.
Здесь описаны и охарактеризованы гены, кодирующие белок, экспрессируемые в скелетных мышцах, вместе с примерами иммуногистохимически окрашенных срезов ткани, которые визуализируют соответствующие паттерны экспрессии белков генов с повышенной экспрессией в скелетных мышцах.
Профилирование транскриптов было основано на комбинации трех наборов данных транскриптомики (HPA, GTEx и FANTOM5, что соответствует в общей сложности 483 образцам из 37 различных типов нормальных тканей человека. Использовалось окончательное согласованное значение нормализованной экспрессии (NX) для каждого типа ткани) для классификации всех генов в соответствии с тканеспецифической экспрессией на две разные категории на основе специфичности или распределения.
Uhlén M et al., Тканевая карта протеома человека. Science (2015)
PubMed: 25613900 DOI: 10.1126 / science.1260419
Yu NY et al., Дополнение характеристики ткани путем интеграции профилей транскриптомов из Атласа белков человека и консорциума FANTOM5. Nucleic Acids Res. (2015)
Nucleic Acids Res. (2015)
PubMed: 26117540 DOI: 10.1093 / nar / gkv608
Fagerberg L et al., Анализ тканеспецифической экспрессии человека путем полногеномной интеграции транскриптомики и протеомики на основе антител. Протеомика клеток Mol. (2014)
PubMed: 24309898 DOI: 10.1074 / mcp.M113.035600
Lindskog C et al., Протеомы сердечных и скелетных мышц человека, определенные с помощью транскриптомики и профилирования на основе антител. BMC Genomics. (2015)
PubMed: 26109061 DOI: 10.1186 / s12864-015-1686-y
Гистологический словарь — скелетная мышца
Ошибка
Обратная связь
Пожалуйста, расскажите нам о своем опыте работы с AclandAnatomy! Пожалуйста, расскажите нам о своем опыте работы с AclandAnatomy!(1000 осталось символов)
Легкость использования 1 = Не проста в использовании; 5 = Очень проста в использовании| 1 | 2 | 3 | 4 | 5 |
| 1 | 2 | 3 | 4 | 5 |
| 1 | 2 | 3 | 4 | 5 |
| 1 | 2 | 3 | 4 | 5 |
 Д.)?
Д.)?| Да | Нет |
Расскажи нам, кто ты.
| Студент |
| Факультет |
| Профессиональный |
| Другое (укажите) |
| Да | Нет |
Ваш отзыв был успешно отправлен.
В настоящее время мы не можем получить ваш отзыв. Пожалуйста, попробуйте еще раз в другой раз.
Пожалуйста, войдите, чтобы оставить отзыв.
Уже подписчик?
ВойтиОбязательный Обязательный
Забыл пароль? Введите код доступа Войдите через: Открыть Афины | Шибболет ×(PDF) Атлас скелетных мышц человека определяет траектории стволовых клеток и клеток-предшественников в процессе развития и от человеческих плюрипотентных стволовых клеток
Borchin, B. , Чен, Дж., И Барбери, Т. (2013). Получение и FACS-опосредованная очистка
, Чен, Дж., И Барбери, Т. (2013). Получение и FACS-опосредованная очистка
предшественников скелетных мышц PAX3 + / PAX7 + из человеческих стволовых клеток pluripo-
. Отчеты о стволовых клетках 1, 620–631.
Brent, A.E., и Табин, C.J. (2002). Регуляция развития производных сомита —
факторов: мышцы, хрящи и сухожилия. Curr. Opin. Genet. Dev. 12, 548–557.
Батлер А., Хоффман П., Смиберт П., Папалекси Э. и Сатия Р. (2018).
Объединение транскриптомных данных отдельных клеток в различных условиях, технологиях и видах.Nat. Biotechnol. 36, 411–420.
Цао, Дж., Шпильманн, М., Цю, X., Хуанг, X., Ибрагим, Д.М., Хилл, А.Дж., Чжан, Ф.,
,Мундлос, С., Кристиансен, Л., Стимерс, Ф.Дж. , и другие. (2019). Одноклеточный транскрипционный ландшафт органогенеза млекопитающих. Природа 566, 496–502.
Castiglioni, A., Hettmer, S., Lynes, M.D., Rao, T.N., Tchessalova, D., Sinha, I.,
Lee, B.T., Tseng, Y.H., and Wagers, A.J. (2014). Выделение предшественников, которые
проявляют миогенную / остеогенную бипотентность in vitro, путем сортировки активируемых флуоресценцией клеток
из мышц плода человека. Отчеты о стволовых клетках 2, 92–106.
Отчеты о стволовых клетках 2, 92–106.
Cerletti, M., Shadrach, J.L., Jurga, S., Sherwood, R., and Wagers, A.J. (2008).
Регуляция и функция стволовых клеток скелетных мышц. Харб Холодного источника.
Symp. Quant. Биол. 73, 317–322.
Чал Дж. И Пурки
´, О. (2017). Создание мышц: скелетный миогенез in vivo
и in vitro. Разработка 144, 2104–2122.
Чал, Дж., Огинума, М., Аль Танури, З., Гоберт, Б., Сумара, О., Хик, А.,
Буссон, Ф., Zidouni, Y., Mursch, C., Moncuquet, P., et al. (2015).
Дифференциация плюрипотентных стволовых клеток в мышечное волокно для моделирования мышечной дистрофии Дюшенна
. Nat. Biotechnol. 33, 962–969.
Чал, Дж., Аль Танури, З., Хестин, М., Гоберт, Б., Айвио, С., Хик, А., Черриер, Т.,
, Несмит, А.П., Паркер, К.К., и Пурки
´, О. (2016). Получение человеческих
мышечных волокон и сателлитных клеток из человеческих плюрипотентных стволовых клеток
in vitro.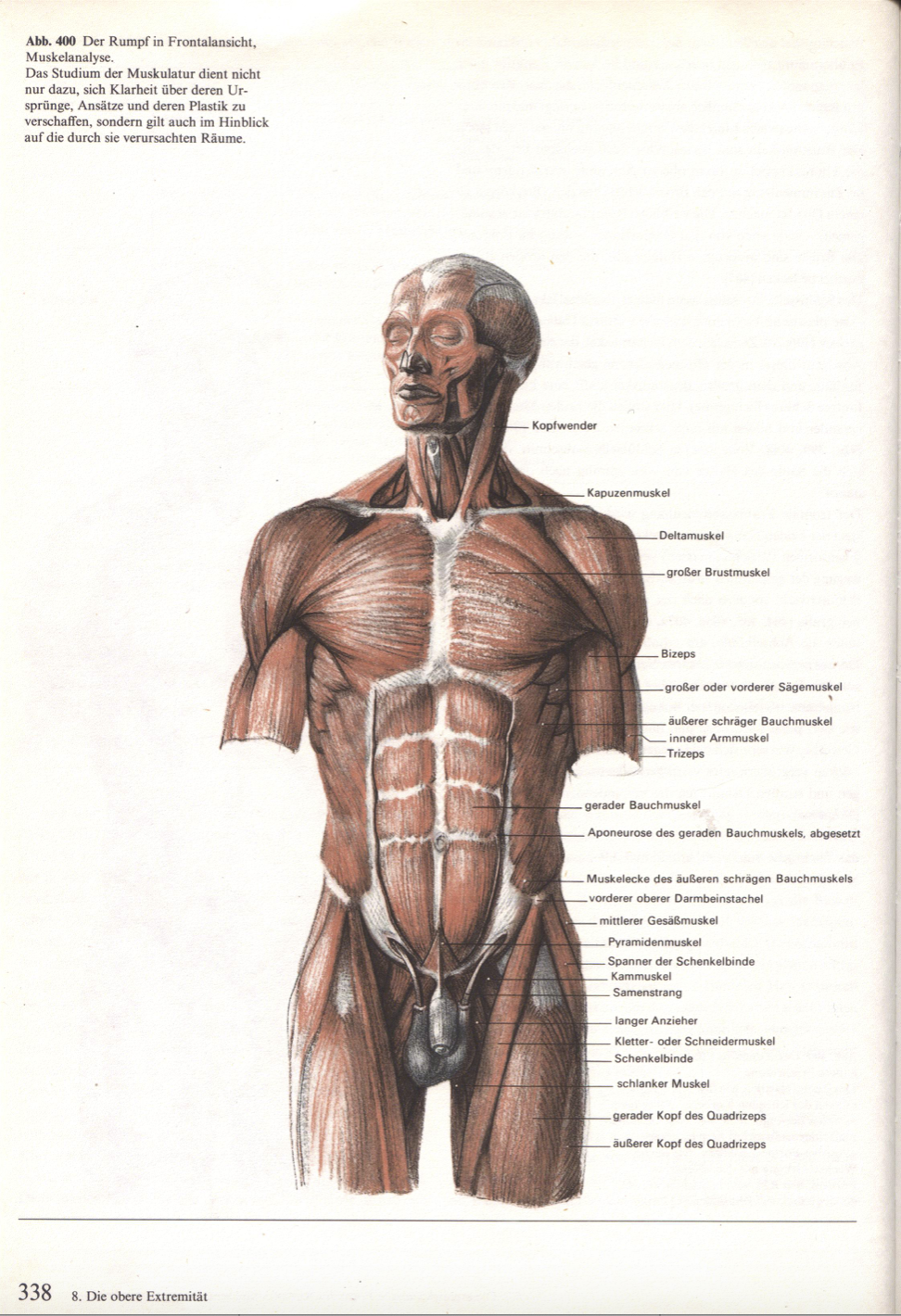 Nat. Protoc. 11, 1833–1850.
Nat. Protoc. 11, 1833–1850.
Чемберс, С.М., Фазано, К.А., Папапетру, Э.П., Томишима, М., Саделайн,
М., и Студер, Л. (2009). Высокоэффективная нейронная конверсия человеческих ES
и iPS клеток за счет двойного ингибирования передачи сигналов SMAD. Nat. Biotechnol. 27,
275–280.
Чавес, А., Шейман, Дж., Вора, С., Прюитт, Б.В., Таттл, М., П.Р. Айер, Э., Лин, С.,
Киани, С., Гусман, С.Д., Виганд, DJ, et al. (2015). Высокоэффективное транскрипционное программирование, опосредованное Cas9-
.Nat. Методы 12, 326–328.
Chen, J.F., Tao, Y., Li, J., Deng, Z., Yan, Z., Xiao, X., and Wang, D.Z. (2010).
microRNA-1 и microRNA-206 регулируют пролиферацию и дифференцировку r-
сателлитных клеток скелетных мышц путем репрессии Pax7. J. Cell Biol. 190, 867–879.
Cheung, M., Tai, A., Lu, P.J. и Cheah, K.S. (2019). Приобретение мультипотентных
и мигрирующих клеток нервного гребня в эволюции позвоночных. Curr. Opin. Genet. Dev.
Genet. Dev.
57, 84–90.
De Micheli, A.J., Laurilliard, E.J., Heinke, C.L., Ravichandran, H., Fraczek, P.,
,Soueid-Baumgarten, S., De Vlaminck, I., Elemento, O., and Cosgrove, B.D.
(2020). Одноклеточный анализ иерархии мышечных стволовых клеток выявляет типичные сигналы гетеро-
, участвующие в регенерации скелетных мышц. Cell
Rep. 30, 3583–3595.e5.
Делль’Орсо, С., Хуан, А.Х., Ко, К.Д., Наз, Ф., Перованович, Дж., Гутьеррес-Крус, Г.,
Фенг, X., и Сарторелли, В. (2019). Анализ одиночных клеток скелетных
мышечных стволовых клеток взрослых мышей в гомеостатических и регенеративных условиях. Разработка
146, dev174177.
Фламини В., Гадиали Р.С., Антчак П., Ротвелл А., Тернбулл Дж. Э. и
Писконти А. (2018). Ниша сателлитных клеток регулирует баланс между дифференцировкой
миобластов и самообновлением посредством p53. Отчеты о стволовых клетках 10,
970–983.
Фу Ю., Сандер Дж.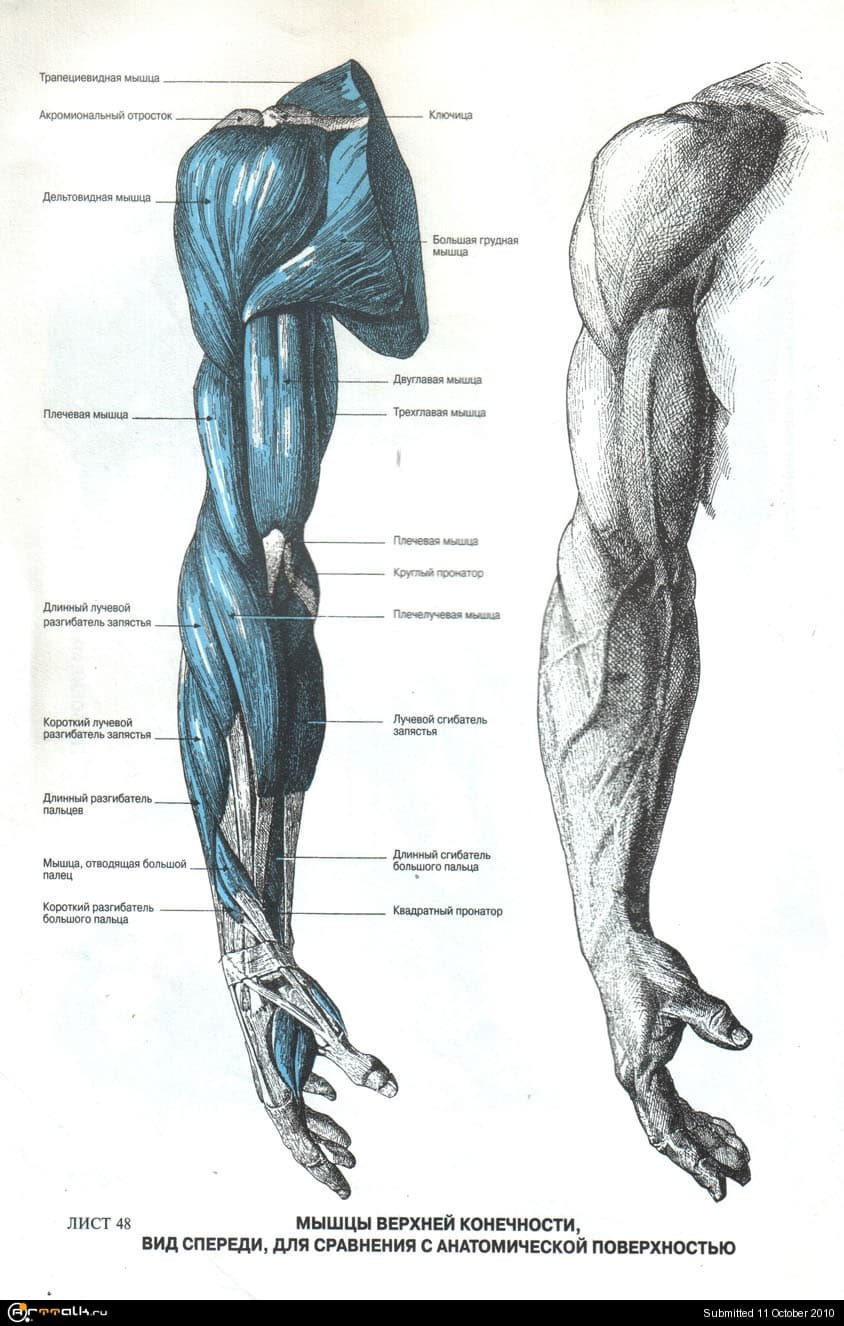 D., Reyon, D., Cascio, V.M., и Joung, J.K. (2014).
D., Reyon, D., Cascio, V.M., и Joung, J.K. (2014).
Повышение специфичности нуклеазы CRISPR-Cas с помощью усеченных направляющих РНК.
Нат. Biotechnol. 32, 279–284.
Джордани, Л., Хе, Г.Дж., Негрони, Э., Сакаи, Х., Лоу, Дж.М.С., Сиу, М.М., Ван, Р.,
Корно, А., Таджбахш, С., Чунг, Т.Х., и Ле Гран, Ф. (2019). Высокомерная одноклеточная картография
выявила новые резидентные в скелетных мышцах
популяции клеток. Мол. Cell 74, 609–621.e6.
Gopinath, S.D., Webb, A.E., Brunet, A., and Rando, T.A. (2014). FOXO3 pro-
находится в состоянии покоя в взрослых мышечных стволовых клетках в процессе самообновления. Отчеты о стволовых клетках 2, 414–426.
Грос, Дж. И Табин, К.Дж. (2014). Формирование зачатка конечностей позвоночных инициируется
локализованным эпителиально-мезенхимальным переходом. Science 343, 1253–1256.
Haghverdi, L., Buettner, F., and Theis, F.J. (2015). Карты диффузии для анализа данных дифференцировки с высокой размерностью
отдельных клеток.
2989–2998.
Hayashi, K., and Ozawa, E. (1995). Миграция миогенных клеток из сомитов
индуцируется контактом ткани с медиальной областью дермы презумптивной конечности мезо-
у куриных эмбрионов. Разработка 121, 661–669.
Хикс, М.Р., Хисеродт, Дж., Парас, К., Фудзивара, В., Эскин, А., Ян, М., Си, Х.,
Янг, К.С., Евсеенко, Д., Нельсон, С.Ф. , и другие. (2018). ERBB3 и NGFR
маркируют отдельные клетки-предшественники скелетных мышц в человеческом развитии и
hPSC.Nat. Cell Biol. 20, 46–57.
Джо, А.В., Йи, Л., Натараджан, А., Ле Гранд, Ф., Со, Л., Ван, Дж., Рудницки, М.А.,
и Росси, Ф.М. (2010). Повреждение мышцы активирует резидентные фибро / адипогенные гениторы pro-
, которые способствуют миогенезу. Nat. Cell Biol. 12, 153–163.
Ким, Дж., Магли, А., Чан, С.С.К., Оливейра, В.К.П., Ву, Дж., Дараби, Р., Киба, М.,
и Перлингейро, Р.С.Р. (2017). Расширение и очистка имеют решающее значение для терапевтического применения
миогенных предшественников плюрипотентных стволовых клеток.
Отчеты о стволовых клетках 9, 12–22.
Kivel €
a, R., Salmela, I., Nguyen, Y.H., Petrova, T.V., Koistinen, H.A., Wiener, Z.,
and Alitalo, K. (2016). Фактор транскрипции Prox1 важен для дифференцировки сателлитных клеток
и регуляции типа мышечных волокон. Nat. Commun. 7, 13124.
Koopman, R., Ly, C.H., and Ryall, J.G. (2014). Метаболическая связь с истощением и регенерацией скелетных мышц. Фронт. Physiol. 5, 32.
Лангмид, Б., и Зальцберг, С. (2012). Быстрое выравнивание с пропуском считывания с
Bowtie 2. Нат. Методы 9, 357–359.
Ли, X., Чжао, X., Fang, Y., Jiang, X., Duong, T., Fan, C., Huang, C.C., and Kain,
S.R. (1998). Получение дестабилизированного зеленого флуоресцентного белка в качестве репортера транскрипции транскрипции
. J. Biol. Chem. 273, 34970–34975.
Machado, L., Esteves de Lima, J., Fabre, O., Proux, C., Legendre, R., Szegedi,
A., Varet, H., Ingerslev, LR, Barre
`s , Р. , Relaix, F., и Mourikis, P. (2017). В
, Relaix, F., и Mourikis, P. (2017). В
фиксация на месте переопределяет покой и раннюю активацию стволовых клеток скелетных мышц
. Cell Rep. 21, 1982–1993.
Macosko, EZ, Basu, A., Satija, R., Nemesh, J., Shekhar, K., Goldman, M.,
Tirosh, I., Bialas, AR, Kamitaki, N., Martersteck, EM , и другие. (2015). Профилирование экспрессии отдельных клеток с высоким пар-
аллелем в масштабе всего генома с использованием нанолитровой капли —
лет. Cell 161, 1202–1214.
Магли, А., и Perlingeiro, R.R.C. (2017). Спецификация миогенного предшественника
из плюрипотентных стволовых клеток. Семин. Cell Dev. Биол. 72, 87–98.
Мали, П., Янг, Л., Эсвелт, К.М., Аач, Дж., Гуэль, М., ДиКарло, Дж. Э., Норвилл, Дж. Э.,
и Черч, Г. (2013). РНК-управляемая инженерия генома человека с помощью Cas9.
Наука 339, 823–826.
МакКоннелл, Б. Б., Янг, В. В. (2010). Млекопитающие Kr €
благоприятных факторов в
здоровье и болезни. Physiol.Ред. 90, 1337–1381.
Physiol.Ред. 90, 1337–1381.
Мессина, Г., Биресси, С., Монтеверде, С., Магли, А., Кассано, М., Перани, Л.,
Ронкалья, Э., Тальяфико, Э., Старнес, Л., Кэмпбелл , CE и др. (2010). Nfix reg-
производит специфичную для плода транскрипцию в развивающихся скелетных мышцах. Cell 140,
554–566.
Murmann, O.V., Niggli, F., and Sch €
afer, B.W. (2000). Клонирование и характеристика промотора PAX7 человека. Биол. Chem. 381, 331–335.
Нойфельд, С.Дж., Ван Ф. и Кобб Дж. (2014). Генетические взаимодействия между генами
Shox2 и Hox во время регионального роста и развития конечности мыши
. Генетика 198, 1117–1126.
Огинума, М., Монкуке, П., Сюн, Ф., Кароли, Э., Чал, Дж., Геворкия, К.,
и Пурки
´, О. (2017). Градиент гликолитической активности координирует передачу сигналов FGF
и Wnt во время удлинения оси тела у эмбрионов амниот.
Дев. Cell 40, 342–353.e10.
Ох, Ю. и Джанг, Дж. (2019). Направленная дифференцировка плюрипотентных стволовых клеток с помощью
Направленная дифференцировка плюрипотентных стволовых клеток с помощью
факторов транскрипции. Мол. Ячейки 42, 200–209.
Pala, F., Di Girolamo, D., Mella, S., Yennek, S., Chatre, L., Ricchetti, M., и
Tajbakhsh, S. (2018). Определенные метаболические состояния определяют судьбу стволовых клеток скелетных мышц
во время пренатального и постнатального миогенеза. J. Cell Sci. 131,
jcs212977.
ll
Resource
Cell Stem Cell 27, 1–19, 2 июля 2020 г. 17
Цитируйте эту статью в прессе как: Xi et al., Атлас скелетных мышц человека определяет траектории стволовых клеток и клеток-предшественников в процессе развития
и от Human Pluripotent Stem Cells, Cell Stem Cell (2020), https://doi.org/10.1016/j.stem.2020.04.017
Атлас метиломов ДНК в жировой и мышечной тканях свиней
Образцы и их фенотипы, связанные с ожирением
В этом исследовании мы выбрали три породы свиней на основе известной истории формирования пород и измерения фенотипов, связанных с ожирением (см. Дополнительные методы ).Порода Ландрас была выбрана в Европе более 100 лет для уменьшения ожирения, в то время как порода Ронгчанг была выбрана из-за чрезмерной жировой прослойки. Тибетская порода — это почти дикая порода, которая почти не подвергалась искусственному отбору. В среднем у взрослых самок процент жира выше, чем у самцов, когда они достигают половой зрелости в возрасте 210 дней. Чтобы исследовать половые различия, мы также разделили мужчин и женщин в сравнении. Как и ожидалось, плотность тела, которая отрицательно коррелирует с процентом жира, показала значительную разницу между тремя породами (двухфакторный дисперсионный анализ (ANOVA), P B = 6.98 × 10 −10 ) и между самцом и самкой (двухфакторный дисперсионный анализ, P S = 0,02; рис. 1а). Измерение показателей метаболизма в сыворотке также выявило такой же ранжирование (дополнительный рисунок S1).
Дополнительные методы ).Порода Ландрас была выбрана в Европе более 100 лет для уменьшения ожирения, в то время как порода Ронгчанг была выбрана из-за чрезмерной жировой прослойки. Тибетская порода — это почти дикая порода, которая почти не подвергалась искусственному отбору. В среднем у взрослых самок процент жира выше, чем у самцов, когда они достигают половой зрелости в возрасте 210 дней. Чтобы исследовать половые различия, мы также разделили мужчин и женщин в сравнении. Как и ожидалось, плотность тела, которая отрицательно коррелирует с процентом жира, показала значительную разницу между тремя породами (двухфакторный дисперсионный анализ (ANOVA), P B = 6.98 × 10 −10 ) и между самцом и самкой (двухфакторный дисперсионный анализ, P S = 0,02; рис. 1а). Измерение показателей метаболизма в сыворотке также выявило такой же ранжирование (дополнительный рисунок S1).
( a ) Разница в плотности тела у свиней Ландрас (L), тибетской (T) и Rongchang (R), а также между самцом (M) и самкой (F). Двусторонний дисперсионный анализ с повторными измерениями ( n = 9 на породу и пол). «B» и «S» означают породу и пол соответственно.Значения являются средними ± стандартное отклонение. ( b ) Источники тканей: три SAT (ASA: брюшная подкожная жировая ткань, ILB: внутренний слой спинного жира, ULB: верхний слой спинного жира), четыре VATs (GOM: большой сальник, MAD: мезентериальный жир, RAD: забрюшинный жир. , PAD: жировая ткань перикарда), межмышечная жировая ткань (IAD) и два SMT (LDM: longissimus dorsi, мышца, и PMM: psoas, большая мышца). ( c ) Разница в составе жирных кислот. НЖК, МНЖК и ПНЖК означают насыщенные, мононенасыщенные и полиненасыщенные жирные кислоты соответственно.Трехфакторный дисперсионный анализ с повторными измерениями ( n = 9 на породу, пол на ткань). (SFA: P B = 2,46 × 10 −7 , P S = 1,35 × 10 −6 , P T = 0,99, P B × S = 0,19, P B × T = 0,96, P S × T = 0,69, P B × S × T = 0,77; MUFA: P B = 0,006, P S = 0,0004, P T = 0.98, P B × S = 0,99, P B × T = 0,99, P S × T = 0,93, P B × S × T = 0,77; и PUFA: P B = 0,0008, P S = 0,14, P T = 0,98, P B × S = 0,03, P B × T = 0,99, P S × T = 0,56, P B × S × T = 0,099). ( d ) Разница в объеме адипоцитов. Трехфакторный дисперсионный анализ с повторными измерениями ( n = 9 на породу, пол на ткань).«Т» означает ткань. ( P B <10 −16 , P S = 10 −16 , P T = 6,74 × 10 −12 , P B × S <10 −16 , P B × T = 0,29, P S × T = 0,36, P B × S × T = 0,99). Значения являются средними ± стандартное отклонение. ( e ) Разница в площади поперечного сечения миофибрилл. Трехфакторный дисперсионный анализ с повторными измерениями ( n = 9 на породу, пол на ткань; P B <10 −16 , P S = 0.005, P T = 9,66 × 10 −12 , P B × S = 0,44, P B × T = 0,01, P S × T = 0,583, P B × S × T = 0,07). ( f ) Разница в соотношении типов миофибрилл. Трехфакторный дисперсионный анализ с повторными измерениями ( n = 9 на породу для каждого пола на ткань; P B = 4,42 × 10 −10 , P S = 5,45 × 10 −9 , P T <10 −16 , P B × S = 0.004, P B × T = 0,02, P S × T = 1,61 × 10 −5 , P B × S × T = 0,04).
Для изучения регуляции адипоцитов в различных анатомических точках мы взяли образцы восьми АТ из разных областей тела (рис. 1b), которые показали разный состав жирных кислот (рис. 1c) и значительно различались объемы адипоцитов (трехфакторный ANOVA, P T = 6,74 × 10 −12 ) среди трех пород (трехфакторный дисперсионный анализ, P B <10 −16 ) и между самцами и самками (трехфакторный дисперсионный анализ, P S = 10 −16 ; рис.1d; Дополнительный рис. S2). Мы также взяли образцы двух SMT, белой мышцы longissimus dorsi, (LDM) и красной поясничной мышцы, основной мышцы (PMM; рис. 1b), представляющих два разных типа волокон, из которых PMM имеет более высокий процент капилляров, миоглобина, липидов и митохондрии 17 . По сравнению с PMM, LDM имеет более высокую площадь поперечного сечения миофибрилл (трехфакторный дисперсионный анализ , P T = 9,66 × 10 −12 ; рис. 1e; дополнительный рис. S2) и соотношение быстрых и медленных сокращений миофибрилл. (Трехфакторный дисперсионный анализ , P T <10 −16 ; рис.1f; Дополнительный рис. S2). Также наблюдается значительное расхождение в площади поперечного сечения миофибры (трехфакторный ANOVA , P B <10 −16 , P S = 0,005) и соотношении типов миофибр (трехфакторный дисперсионный анализ ANOVA, P B = 4,42 × 10 −10 , P S = 5,45 × 10 −9 ) среди трех пород и между двумя полами. Эти фенотипические различия для AT и SMT между породами, полами и анатомическим расположением подразумевают внутренние эпигеномные различия.
Пейзаж метиломов ДНК
Мы сгенерировали в общей сложности 1381 ГБ данных секвенирования метилированной ДНК (MeDIP-seq) из 180 образцов (~ 7,67 ГБ на образец), из которых 1067 ГБ (77,3%) чистых считываний были выровнены на геном свиньи. После удаления неоднозначно сопоставленных чтений и чтений, которые могли быть получены из повторяющихся клонов, мы использовали 993 ГБ (71,9%) однозначно выровненных недубликатных чтений в следующем анализе (дополнительная таблица S1). Чтобы избежать ложноположительных результатов при обогащении, нам потребовалось не менее десяти чтений для определения метилированного CpG в образце.В среднем 16,1% CpG были покрыты этим порогом (дополнительный рисунок S3).
Измерение уровня метилирования ДНК по хромосомам показало, что Х-хромосома глобально гиперметилирована у женщин по сравнению с мужчинами (рис. 2а), что можно объяснить инактивацией Х-хромосомы у женщин 18 . Путем сравнения уровня метилирования ДНК между каждой парой образцов мы обнаружили переменные скорости корреляции в разных категориях (рис. 2b). Биологические реплики сильно коррелировали друг с другом (медиана Пирсона r = 0.95 для SMT и r = 0,94 для AT), что предполагает как экспериментальную надежность, так и эпигенетическую согласованность в пределах одной и той же группы породы / пола / типа ткани. Коэффициенты корреляции были относительно ниже между мужчинами и женщинами (медиана Пирсона r = 0,92 для SMT и r = 0,91 для AT) и еще ниже между различными анатомическими точками (медиана r = 0,91 для SMT по Пирсону, r = 0,89 для между AT и SMT и r = 0,87 для AT) и между разными породами (медиана Пирсона r = 0.88 для AT и r = 0,84 для SMT), что указывает на значительные биологические различия в последних категориях.
Рисунок 2: Хромосомные профили метиломов AT и SMT свиней и их изменчивость.( a ) Распределение уровня метилирования ДНК в геноме свиньи. Для сравнения скоростей метилирования ДНК среди образцов глубина считывания была нормализована по общему среднему количеству считываний в каждой группе, а затем было использовано скользящее окно размером 1 МБ для сглаживания распределения. Отношение CpG o / e , плотность SNP, гена, повтора и CGI рассчитывали с помощью скользящего окна размером 1 МБ.( b ) Коробчатые диаграммы парной корреляции Пирсона по скорости метилирования между образцами. Скорость корреляции между каждыми двумя выборками рассчитывалась с помощью скользящего окна размером 1 кб. Затем эти коэффициенты корреляции были сгруппированы в следующие категории: биологические реплики, тот же пол, те же ткани, те же породы и между жировой и мышечной тканями. Поскольку Х-хромосома имеет явно более высокий уровень метилирования у женщин, чем у мужчин, мы исключили его из анализа между полами. Прямоугольники обозначают межквартильный размах (IQR) между первым и третьим квартилями (25-й и 75-й процентили соответственно), а внутренняя линия обозначает медиану.Усы обозначают самые низкие и самые высокие значения в пределах 1,5-кратного IQR из первого и третьего квартилей, соответственно. Выбросы за пределами усов показаны черными точками.
Мы заметили, что уровень метилирования отрицательно коррелирует с длиной хромосомы ( r = -0,614, P = 0,005) и положительно с содержанием GC ( r Пирсона = 0,784, P = 7 × 10 -5. ), плотность повторов ( r Пирсона = 0,336, P = 0,159), плотность гена ( r Пирсона = 0.535, P = 0,018), и особенно с наблюдаемым превышением ожидаемого отношения количества CpG (CpG o / e ) ( r Пирсона = 0,902, P = 1,33 × 10 −7 ) (рис. 3а), что согласуется с предыдущими отчетами 19 . Более подробный анализ показал, что области с содержанием GC около 46% и соотношением CpG o / e около 0,35, как правило, имеют более высокий уровень метилирования (рис. 3b, c), где среднее содержание GC в геноме свиньи составляет 40%. и отношение CpG o / e равно 0.21 (рис. 3г, д). Тем не менее, нет значимой корреляции между содержанием GC и соотношением CpG o / e в геномной шкале ( r Пирсона = 0,10, P = 0,106; рис. 3f). Анализ показал, что соотношение CpG o / e имеет более высокую скорость корреляции с уровнем метилирования ДНК, чем с содержанием GC. Плотность однонуклеотидного полиморфизма (SNP) также положительно коррелировала с уровнем метилирования ( r Пирсона = 0,597, P = 0,007; рис. 3а). Хотя была обнаружена существенная ассоциация плотности SNP и уровня метилирования ДНК 20 , механизм до сих пор неясен.Кроме того, богатая генами субтеломерная область (7 МБ от каждой теломеры) имеет значительно более высокое метилирование в большинстве (~ 80%, 15 из 19) хромосом (тест Стьюдента t , P <0,01; рис. 3g) .
Рис. 3. Уровень метилирования ДНК с геномными особенностями.( a ) Корреляция Пирсона между уровнем метилирования ДНК и особенностями аутосом свиньи (хромосомы 1–18) и половой хромосомы X. Глубина считывания была нанесена на график в зависимости от длины, содержания GC, плотности повторов, плотности гена, CpG соотношение o / e и плотность SNP отдельной хромосомы.Линия представляет линейную регрессию. Значения являются средними ± стандартное отклонение ( n = 180). ( b — e ) Секвенирование считывает распределение относительно соотношения CpG o / e и содержания GC. Считывает распределение по содержанию GC ( b ) и соотношению ( c ) CpG o / e по всем образцам. Значения являются средними ± стандартное отклонение ( n = 180). Коробчатая диаграмма содержания GC ( d ) и соотношения ( e ) CpG o / e всего генома в окнах размером 1 kb. Прямоугольники обозначают межквартильный размах (IQR) между первым и третьим квартилями (25-й и 75-й процентили соответственно), а внутренняя линия обозначает медиану.Усы обозначают самые низкие и самые высокие значения в пределах 1,5-кратного IQR из первого и третьего квартилей, соответственно. Выбросы за пределами усов показаны черными точками. Содержание GC 46–47% и соотношение 0,35–0,36 CpG o / e соответствуют максимальному% от общего числа считываний, то есть 7,52 ± 0,49 и 2,03 ± 0,07% соответственно. Геномная медиана содержания GC (40–41%) и отношения CpG o / e (0,21–0,22) соответствуют 4,59 ± 0,63 и 1,50 ± 0,17% от общего числа считываний, соответственно. ( f ) График соотношения содержания GC и CpG o / e окон размером 1 т.п.н. в геноме свиньи.Геном свиньи был разделен на 2 262 494 окна размером 1 т.п.н., и эти окна использовались для расчета коэффициента корреляции Пирсона ( r ) между содержанием GC и соотношением CpG o / e . Красная линия представляет собой линейную регрессию. ( г ) Уровни метилирования в субтеломерных (до 7 Mb от каждой теломер) и не субтеломерных областях в каждой хромосоме. (Тест Стьюдента t , ** P <0,01). Значения являются средними ± стандартное отклонение ( n = 180).
Характеристика DMR
Мы использовали статистику для измерения изменений скорости метилирования и определили DMR для разных пород (B-DMR), полов (S-DMR) и тканей (T-DMR; см. Дополнительные методы).Высокая корреляция (среднее значение Пирсона r = 0,994) между количеством DMR, количеством CpG в DMR и длиной DMR подразумевает, что обнаружение DMR в областях разной длины и количество встроенных CpG в него не было смещенным. Количество DMR значительно варьировалось между категориями (например, 387 мышечных S-DMR против 218 623 мышечных B-DMR; Таблица 1).
Макроскопическое отображение DMR вдоль хромосом показывает, что богатые DMR области также преимущественно имеют более высокие отношения CpG o / e (~ 0.35), чем в среднем по геному (0,21; рис. 4а). Более 20% DMR расположены в субтеломерных областях, которые занимают только 11,76% всего генома. Среди 282 генов свиней, которые были ортологами известных генов, связанных с ожирением человека, 2,12,13,21 , 223 (~ 80%) находились в пределах установленных нами DMR (рис. 4a; дополнительные данные 1 и 2), что предполагает что DMR имеют высокую ассоциацию с известными генами, связанными с ожирением, и эти гены могут играть функциональную роль в развитии ожирения путем изменения скорости метилирования.
Рис. 4. Распределение дифференциально метилированных областей по всему геному.( a ) Круговое представление распределения DMR по всему геному. Эта визуализация была создана с использованием программного обеспечения Circos 58 . Крайний круг отображает хромосомы свиньи с масштабом и отношением CpG o / e в ячейках 1 Мб. Во втором круге показаны 282 гена свиньи, которые являются ортологами хорошо аннотированных генов, связанных с ожирением человека (дополнительные данные 1), из которых 223 (79%) гена, перекрывающие определенные DMR, выделены синим цветом, а остальные 59 генов (21 %) красным цветом (Дополнительные данные 2).Кружки с третьего по девятый представляют семь категорий DMR (A: мышечные B-DMR, B: жировые T-DMR, C: жировые B-DMR, D: жировые и мышечные T-DMR, E: мышечные T-DMR, F: S-DMR жировой ткани и G: S-DMR мышц). Высота столбцов гистограммы указывает количество CpG в DMR. Интервалы с богатым DMR (глубокий цвет, χ 2 -тест, P <0,001) и плохим DMR (светлый цвет, χ 2 -тест, P > 0,001) определяются следующим образом: сравнивая со средним геномом.TES: конечный сайт транскрипции. ( b ) Все 17 930 промоторов в геноме свиньи были классифицированы на три типа на основе представления CpG. HCP (синий, n = 7 249), ICP (желтый, n = 6 629) и LCP (красный, n = 4052). ( c ) Диаграмма Венна, показывающая распределение всех 38 778 CGI в геноме свиньи среди 5 классов CGI. CGI промотора ( n = 7,126), внутригенные CGI ( n = 13 611), CGI с 3′-транскриптом ( n = 2 305), межгенные CGI ( n = 16,954) и промотор miRNA CG25 908 = 169) были определены в соответствии с их геномным местоположением.Есть перекрытия между пятью классами CGI из-за перекрывающейся аннотации генов в определенной координате генома. ( d ) Процент CpG в DMR в каждом из 31 геномного элемента. Статистическая значимость сравнения трех промоторов miRNA (D, I, P) рассчитывалась с помощью однофакторного дисперсионного анализа с повторными измерениями, а других — с помощью двустороннего дисперсионного анализа с повторными измерениями. ( e ) Распределение отношения CpG o / e для CGI и их берегов в пяти классах CGI.Для определения разницы в соотношении CpG o / e между CGI и их берегами использовался критерий суммы рангов Вилкоксона.
Затем мы рассмотрели DMR в 31 категории функциональных геномных элементов. Мы разделили промоторы на три типа в соответствии с представлением CpG, как описано ранее 22 (Рис. 4b), а также классифицировали CpG-островки (CGI) на пять классов в соответствии с их геномным расположением, как описано ранее 23 (см. Методы; Рис. . 4c). DMR чаще встречаются в промежуточном промоторе CpG (ICP), чем в областях промотора с высоким CpG (HCP) и промотора с низким CpG (LCP) (двусторонний ANOVA, P = 0.056; Рис. 4г). Класс ICP содержит множество слабых CGI 24 (<500 п.н., имеют умеренное богатство CpG и / или имеют содержание GC ниже 55%). Этот результат подтвердил предыдущие открытия, что слабые CGIs более предрасположены к регуляции с помощью метилирования ДНК, а предпочтительное нацеливание на слабые CGIs является общим феноменом у млекопитающих 22 . Гиперметилирование промоторов играет критическую роль в подавлении экспрессии генов, но в телах генов оно также важно для регуляции альтернативных промоторов и предотвращения ложной инициации транскрипции 23 .Интересно, что области первого экзона имеют самые низкие DMR в теле гена (Fig. 4d), что может быть из-за перекрывания некоторых функциональных мотивов между проксимальной областью промоторов и первыми экзонами. Кроме того, дистальные (D) области промоторов мРНК и микроРНК (miRNA) имеют больше DMR, чем промежуточные (I) и проксимальные (P) области (рис. 4d), что позволяет предположить, что изменения в метилировании в D-областях промоторов могут быть более распространенным механизмом для создания вариабельности транскрипции.Мы обнаружили, что большинство DMR расположены на берегах CGI, а не в CGI (рис. 4d) во всех пяти классах геномных элементов (двусторонний дисперсионный анализ, P = 0,002), что согласуется с предыдущими сообщениями 25,26, 27 . Отношение CpG o / e для большинства берегов CGI составляет около 0,35–0,36, тогда как CGI имеют отношения CpG o / e , намного превышающие это пороговое значение (критерий суммы рангов Вилкоксона, P <10 −16 ; рис. 4e), как показано на графике распределения считываний в зависимости от отношения CpG o / e на рис.3c.
Метилирование промоторов и репрессия транскрипции
Мы исследовали корреляцию между скоростью метилирования в промоторах и уровнем экспрессии ассоциированных мРНК и миРНК (Рис. 5; Дополнительный Рис. S4). Считается, что метилирование ДНК в промоторах — только один из нескольких механизмов регуляции экспрессии генов; следовательно, логично, что не все гены имеют коррелированные паттерны метилирования и экспрессии. Порядок уровня корреляции в промоторах мРНК, от высокого к низкому, был HCP, ICP, LCP и P, I, D (рис.5), который подтвердил предыдущий отчет 28 . Тем не менее, поскольку DMR обогащены ICP и D-областями промоторов (рис. 4d), было больше пар мРНК-DMR, показывающих корреляцию в ICP (8257), чем в HCP (6217) и LCP (3099), и больше пар в D (9313), чем в I (4923) и P (3337) областях (рис. 5). Кроме того, он показал, что различие в содержании CpG промоторов оказывает более сильное влияние на экспрессию мРНК (двухфакторный дисперсионный анализ, P = 5,56 × 10 -4 ), чем расстояние до сайта начала транскрипции (TSS) областей. в промоторах (двусторонний дисперсионный анализ, P = 0.07).
Фиг. 5. Процент мРНК и миРНК, уровень экспрессии которых отрицательно коррелирует с уровнем метилирования промотора.Использовали только 3074 зонда, однозначно представляющих 3074 мРНК и 611 уникально картированных генов miRNA с данными высокой достоверности экспрессии из тех же образцов для MeDIP-seq (см. Дополнительные методы). Пары мРНК / миРНК-DMR, которые имеют отрицательную регуляцию вместе с гиперметилированием промотора или повышающую регуляцию вместе с гипометилированием промотора, были приняты как имеющие отрицательную корреляцию между экспрессией и метилированием.В скобках указано общее количество идентифицированных пар мРНК / миРНК-DMR. Серые столбцы представляют процент пар мРНК / миРНК-DMR, которые демонстрируют обратную взаимосвязь в общих парах мРНК / миРНК-DMR. Статистическая значимость рассчитывалась с помощью двухфакторного дисперсионного анализа без повторных измерений.
Была обнаружена корреляция между экспрессией miRNA и скоростью метилирования в P-областях промоторов miRNA ( r Пирсона = -0,368, P = 4,44 × 10 -8 ), но почти не было корреляции в I ( r Пирсона). = -0.116, P = 0,146) и D (Pearson’s r = 0, P = 0,996) областей промоторов miRNA (дополнительный рисунок S4). Первичные транскрипты miRNA могут иметь длину в несколько тысяч оснований и включают предшественник стволовой петли длиной ~ 70 нуклеотидов (pre-miRNA) 29 . Хотя мало известно о TSS первичных транскриптов miRNA 30 , результаты здесь подтверждают, что метилирование ДНК в 5′-положении выше pre-miRNAs может играть роль в подавлении транскрипции зрелых miRNA.
Метилирование у берегов CGI имело более сильную (Пирсона r = -0,146, P = 6,15 × 10 -29 ) корреляцию с экспрессией мРНК, чем в CGI (Пирсона r = -0,128.
Промоторные DMR лучше всего различают породы и ткани
Чтобы проанализировать, проявляют ли DMR какие-либо особенности породы, пола и / или анатомического расположения, мы выполнили неконтролируемую кластеризацию для всех образцов с использованием DMR каждой категории геномных элементов. B-DMR жировой и мышечной ткани в промоторах хорошо сгруппированы по породам (рис.6а, б). Кластеризация образцов по соответствующей экспрессии мРНК в целом аналогична (дополнительный рис. S5), что указывает на постоянную взаимосвязь между метилированием ДНК в промоторах и экспрессией генов. Кластеризация с помощью B-DMR на берегах CGI может сгруппировать большинство образцов от каждой породы, но не так отчетливо, как с помощью B-DMR в промоторах. Это предполагает, что, хотя метилирование в берегах CGI важно для регуляции экспрессии генов 25,26,27 , различия в метилировании промоторов являются лучшими предикторами различий между породами.Кластеризация с помощью B-DMRs других геномных элементов еще менее различима, предполагая, что большая часть метилирования в этих геномных элементах может иметь слабую или не иметь прямой связи с функциональной дивергенцией трех пород.
Рисунок 6: Иерархическая кластеризация образцов с использованием DMR в промоторах.( a ) Кластеризация образцов жировой ткани с использованием 1 994 B-DMR жировой ткани в промоторах. ( b ) Кластеризация мышечных образцов с использованием 4816 мышечных B-DMR в промоторах. По определению, три породы свиней полностью разделены.Три основные подгруппы на радиальных дендрограммах полностью соответствуют породе свиней, независимо от пола и типов тканей. ( c ) Кластеризация образцов жировой ткани с использованием 4673 T-DMR жировой ткани в промоторах приводит к тому, что большинство из восьми вариантов жировых образцов отличаются друг от друга. Цифры в скобках показывают количество DMR, используемых при кластеризации.
B-DMR в промоторах AT показали, что порода ронгчанг и тибетская породы ближе друг к другу, чем свинья ландрас (рис.6а). Тот же анализ мышечной ткани показал, что порода ландрас ближе к тибетской, чем порода ронгчанг (рис. 6b). Одинаковый паттерн дистанционных взаимоотношений трех пород отражен соответствующей кластеризацией данных экспрессии мРНК (дополнительный рисунок S5). Различные модели кластеризации можно объяснить заметными фенотипическими изменениями между дикими тибетскими породами, более стройными породами свиней Ландрас и жирными породами свиней Ронгчанг из-за противоположного направления разведения, что приводит к различиям не только на генетическом уровне, но и в эпигенетическом состоянии а также потенциальные взаимодействия генотип-эпигенотип 32 .
Кроме того, T-DMRs в промоторах могут в значительной степени кластеризовать образцы ткани одного и того же типа вместе (Fig. 6c), указывая тем самым, что метилирование промотора также коррелирует с распределением жировой ткани в анатомических местах. Хорошо известно, что НДС имеют внутренние особенности, отличные от SAT, и более тесно связаны с метаболическими факторами риска ожирения, чем SAT 4,6,7,8 . Интересно, что межмышечный жир (IAD), который откладывается между мышечными пучками, был больше похож на VATs с точки зрения метилирования.Это наблюдение предполагает, что ДИА может быть новым фактором риска заболеваний, связанных с ожирением. Перикардиальная жировая ткань (ЗПА) вокруг коронарных артерий является более высоким коррелятивным фактором риска сердечно-сосудистых заболеваний, чем другие НДС, и, хотя грудная ЗПА имеет общее эмбриональное происхождение с другими абдоминальными НДС — висцеральной мезодермой 33 , мы наблюдали значительные локальные специфические различия в скорость метилирования между ними (рис. 6в). Типы тканей также лучше распознаются T-DMR в промоторах, чем в других геномных элементах (дополнительный рис.S5). Ожидается, что метилирование Х-хромосомы между мужчиной и женщиной будет значительным из-за преобладающего эффекта инактивации Х-хромосомы у женщин 18 . Кластеризация S-DMR по полу была менее отчетливой после удаления DMR на Х-хромосоме (дополнительный рис. S5).
Гены, участвующие в фенотипической дивергенции
Чтобы изучить связь дифференциального метилирования в промоторных областях с фенотипической дивергенцией, мы сначала исследовали взаимосвязь между метилированием ДНК на промоторах и данными экспрессии известных генов, связанных с ожирением, полученных с помощью MassArray и количественной ПЦР. (q-ПЦР).Например, FTO (ген, связанный с жировой массой и ожирением) — это ген, однозначно связанный с ожирением, и он экспрессируется повсеместно 34,35 . От более поджарых ландрасов, диких тибетцев до жирных пород ронгчанг, а также для жировой и мышечной ткани, FTO гиперметилирован в области D промотора с более низким уровнем экспрессии гена (рис. 7a). Тот факт, что уровень метилирования самый высокий у свиней Ландрас и самый низкий у свиней Ронгчанг, согласуется с наблюдением, что потеря экспрессии и / или функции FTO защищает от ожирения и приема пищи 35 . ATP1B1 , который кодирует повсеместно экспрессируемую β-субъединицу Na + / K + АТФазы, необходим для правильного клеточного позиционирования АТФазы и ее стабильности. Снижение активности АТФазы предшествует ожирению и гиперинсулинемии, влияя на термогенез и энергетический баланс 36 . COL8A2 , который кодирует цепь α2 коллагена типа VIII, необходим для расширения мезангиального матрикса, а также для гиперцеллюлярности. Отсутствие COL8A2 обеспечивает ренопротектор при диабетической нефропатии 37 .И ATP1B1 , и COL8A2 имеют гиперметилирование в I области промотора и более низкий уровень экспрессии генов, который более выражен в VATs и IAD, чем SATs (рис. 7b), что позволяет предположить, что гиперметилирование в промоторах этих двух генов является потенциальными биомаркерами висцеральное ожирение высокого риска.
Рис. 7. Примеры генов, связанных с ожирением, и функциональных категорий генов, показывающих дифференциальное метилирование ДНК в промоторах.( a ) Жировой и мышечный B-DMR в промоторе FTO .Верхние панели, верхняя половина: метилирование CpG. Каждая точка представляет уровень метилирования (глубину считывания MeDIP-seq) образца на данном сайте CpG. Кривые показали среднее значение по образцам. Две вертикальные пунктирные линии обозначили границы идентифицированного DMR. Нижняя половина: динуклеотиды CpG (черные отметки на оси X), плотность CpG (серая линия), TSS (черная стрелка), экзоны и интроны (черные и белые квадраты, соответственно). Знаки плюс и минус обозначают транскрипцию смыслового и антисмыслового гена. Вторые панели: проверка индивидуального метилирования CpG с помощью MassArray (отображение в желтой рамке на верхней панели).Третьи панели: диаграмма разброса и линия тренда (корреляция Пирсона), показывающая корреляцию между логарифмическими 2 отношениями экспрессии мРНК с микрочипа и CpG-метилированием DMR из MeDIP-seq. Нижние панели: проверка уровней экспрессии мРНК с помощью q-PCR. Полоски представляют средний уровень экспрессии. ( b ) T-DMR жировой ткани в промоторах ATP1B1 и COL8A2 . ( c ) Жировая S-DMR в промоторах miRNA mir-99b, let-7e и mir-125a. ( d ) Десять основных категорий GO (генная онтология) и путей, обогащенных промоторами B-DMR жировой ткани.Анализ обогащения выполняли с использованием программного обеспечения DAVID 59 (см. Дополнительные методы). Оценка EASE, указывающая на значимость сравнения, была рассчитана с использованием модифицированного точного критерия Фишера с поправкой Бенджамини. BP: биологический процесс, MF: молекулярная функция.
Мы также обнаружили корреляцию между метилированием в промоторе и экспрессией генов, а также разумную ассоциацию с породой и дивергенцией анатомического местоположения для многих других генов с известной ролью в отложении жира и росте мышц.Например, ESD , который увеличивает экспрессию в предрасположенных к ожирению моделях 38 ; PPP1R3C , который действует против внутримиоцеллюлярного накопления липидов и снижает циркулирующий лептин и триглицериды 39 ; GHSR , который способствует высвобождению гормона роста и увеличению мышечной, но не жировой массы у субъектов с ожирением 40 ; LIPA , который ингибирует внутримышечные запасы липидов 41 ; MC4R , который подавляет прием пищи и предотвращает гиперинсулинемию и гиперглицинемию 42 ; и PROX1 , который предотвращает лимфатические сосудистые дефекты, вызывающие ожирение у взрослых 43 (дополнительный рис.S6). Гены, предпочтительно экспрессируемые только в жировой (например, HEXB и HTR2A ) или мышечной (например, ACE , PRKAR1A и PRKCQ ) тканях, были подтверждены метилированием промотора и данными экспрессии генов, как колодец (дополнительный рис. S6). Полный список генов-кандидатов, связанных с ожирением, которые мы собрали вместе с их паттерном метилирования ДНК в промоторах, представлен в дополнительных данных 1 и 2.
Кроме того, из 2311 генов / ~ 282.Область локусов количественных признаков (QTL) размером 57 Mb, собранная из 901 суженных (<2 Mb) QTL, влияющих на упитанность и качество свинины, в базе данных PigQTL 44 ; 1669 (72,22%) генов перекрываются с определенными DMR (дополнительные данные 3, 4, 5). Эта высокая согласованность подчеркивает потенциал идентификации областей-кандидатов или генов количественных признаков (таких как ожирение) на основе данных о метилировании ДНК по всему геному, таких как недавно разработанный анализ QTL метилирования 15 .Примечательно, что из 77 предполагаемых генов, расположенных в этой области QTL, 66 (85,71%) перекрываются с определенными нами DMR. Уровень метилирования промоторов этих генов сильно обратно коррелировал с экспрессией генов, предполагая, что эти не охарактеризованные гены, кодирующие белок, могут участвовать в отложении жировой ткани и росте мышц. Типичные примеры показаны на дополнительном рисунке S7.
Кроме того, были идентифицированы многочисленные miRNAs, известные или потенциальные роли в ожирении (дополнительные данные 6).Мы обнаружили S-DMR с гиперметилированием у мужчин по сравнению с женщинами. Этот S-DMR расположен в промоторной области кластера miRNA, который включает соседние miR-99b, let-7e и miR-125a (рис. 7c). Хотя не существует предшествующих доказательств прямой связи этих трех miRNA с ожирением, ключевые функции и мишени этих miRNA связаны с подавлением рака простаты у мужчин 45 и рака груди у женщин 46 и, следовательно, потенциально способствуют половым различиям в развитии ожирения.
Чтобы идентифицировать новые гены, потенциально ответственные за фенотипические различия, мы выполнили анализ обогащения для генов с DMR в промоторах (дополнительный рисунок S8; дополнительные данные 7 и 8). Как и ожидалось, наиболее обогащенные функциональные категории генной онтологии B-DMR жировой ткани в промоторах были связаны с патогенезом ожирения, такими как «гомеостаз стерола, липидов, холестерина», «активность ингибитора липазы», «сахарный диабет I типа» и « дислипидемия »(рис. 7г). Примечательно, что мультигенное семейство глутатионтрансферазы и цитохрома P450, две важные группы многофункциональных детоксифицирующих ферментов, ответственных за метаболизм ряда ксенобиотических соединений , 47,48 , были среди обогащенных жировой B-DMR в промоторах (рис.7г). Ферменты «транс-1,2-дигидробензол-1,2-диолдегидрогеназы», которые также участвуют в метаболизме эндокринно-разрушающих ксенобиотиков 49 , были повсеместно идентифицированы среди T-DMR, обогащенных жировой тканью, в промоторах (дополнительные данные 8). Свиньи, как и люди, подвергаются все большему воздействию ксенобиотиков из окружающей среды в результате проглатывания зараженной пищи или воды, вдыхания загрязненного воздуха или даже воздействия на кожу. Была предложена связь между воздействием ксенобиотиков, разрушающих эндокринную систему, и ожирением 50 .Наше открытие предполагает, что изменения скорости метилирования ДНК генов, кодирующих детоксифицирующие ферменты, вызванные загрязнителями, могут потенциально объяснить патогенность ожирения, вызванного химическими эндокринными разрушителями окружающей среды.
Наш анализ также выявил множество других категорий функциональных генов, которые потенциально участвовали в регуляции жировой ткани и мышц (дополнительные данные 8). Например, категории связанных с иммунитетом генов, в том числе «путь передачи сигналов рецептора RIG-I», «связывание рецептора интерферона-α / β», «цитотоксичность, опосредованная естественными клетками-киллерами» и «процессинг и презентация антигена», были идентифицированы среди обогащенные мышечными B-DMR промоторами, что согласуется с предыдущими открытиями иммунной дисфункции, вызванной ожирением 51 .
